Back to Journals » Drug Design, Development and Therapy » Volume 20
Astragaloside IV Alleviates Lupus Nephritis by Inhibiting Podocyte Ferroptosis via the PI3K/AKT/Nrf2 Pathway
Authors Tang Z, Cheng L ![]() , Li M
, Li M ![]() , Huang C
, Huang C ![]()
Received 5 March 2026
Accepted for publication 22 May 2026
Published 28 May 2026 Volume 2026:20 607142
DOI https://doi.org/10.2147/DDDT.S607142
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Tamer Ibrahim
Zhongfu Tang,1,* Lili Cheng,1,* Ming Li,1,2 Chuanbing Huang1,2
1Department of Rheumatology, The First Affiliated Hospital of Anhui University of Traditional Chinese Medicine, Hefei, People’s Republic of China; 2Key Laboratory of Xin’an Medicine, Ministry of Education, Anhui University of Chinese Medicine, Hefei, People’s Republic of China
*These two authors contributed equally to this work
Correspondence: Chuanbing Huang, Department of Rheumatology, the First Affiliated Hospital of Anhui University of Traditional Chinese Medicine, No. 117 Meishan Road, Shushan District, Hefei, Anhui, People’s Republic of China, Email [email protected]
Purpose: Lupus nephritis (LN) is an immune complex-mediated glomerulonephritis caused by systemic lupus erythematosus (SLE) involving the kidneys. Astragaloside IV (AS-IV) is the main active ingredient of the traditional Chinese medicine Astragalus membranaceus, belonging to the cycloartane-type triterpenoid saponins, and has anti-inflammatory and antioxidant pharmacological effects. This study mainly explores the therapeutic potential and molecular mechanism of AS-IV for LN.
Methods: Using MRL/lpr mice as an in vivo model, the effects of AS-IV on the histopathology and ultrastructure of renal tissues were evaluated by histopathological staining and transmission electron microscopy (TEM). Immunofluorescence and biochemical kits were used to assess the changes in renal damage indicators. Network pharmacology and molecular docking were employed to reveal the molecular targets of AS-IV in the intervention of lupus nephritis (LN). In vitro, a puromycin aminonucleoside (PAN)-induced ferroptosis model of MPC-5 cells was established to evaluate the effects of AS-IV on ferroptosis and the PI3K/AKT/Nrf2 pathway in MPC-5 cells, and further verification was carried out through rescue experiments.
Results: AS-IV can improve the renal histopathology of MRL/lpr mice, reduce the glomerular score, perivascular wall score, foot process fusion rate, and electron-dense deposit density, and decrease the levels of 24hPRO, UTPCR, and UACR, showing a certain protective effect on the kidneys. Meanwhile, AS-IV can upregulate the damage-related proteins in MPC-5 cells, increase the levels of factors related to the PI3K/AKT/Nrf2 pathway, GPX4, SLC7A11, DHODH, FSP1, GSH, and SOD in MPC-5 cells, and reduce the levels of Fe2+, MDA, and ROS. Rescue experiments showed that knockdown of PI3K can reverse the inhibitory effect of AS-IV on ferroptosis in MPC-5 cells.
Conclusion: AS-IV can inhibit podocyte ferroptosis by regulating the PI3K/AKT/Nrf2 axis, reduce the degree of kidney damage, and thus relieve LN. This also indicates that AS-IV has certain therapeutic potential in the treatment of LN.
Keywords: lupus nephritis, Astragaloside IV, podocytes, ferroptosis, renal damage
Introduction
Systemic lupus erythematosus (SLE) is a chronic autoimmune disease characterized by the production of autoantibodies, deposition of immune complexes, and multi-organ damage.1 The clinical manifestations of SLE are diverse, including skin erythema, joint pain, oral ulcers, and kidney damage.2 Among them, the kidney is one of the most commonly affected organs. Approximately 40%-60% of SLE patients will develop lupus nephritis (LN), which has become the main cause of death in SLE patients and end-stage renal disease (ESRD).3 The pathological mechanism of LN is complex, involving glomerular damage mediated by immune complexes, activation of the complement system, infiltration of inflammatory cells, and podocyte damage.4 Although glucocorticoids and immunosuppressants (such as cyclophosphamide and mycophenolate mofetil) are still the standard treatment regimens for LN, some patients still have the risk of treatment resistance or recurrence, and long-term use of these drugs may lead to serious adverse reactions, such as infection and osteoporosis.5 Therefore, exploring safe and effective therapies based on new targets is an urgent need for the clinical management of LN. The core pathological feature of LN is the deposition of immune complexes in the glomerular basement membrane, which triggers complement activation and an inflammatory cascade reaction.6 In recent years, studies have found that in addition to the abnormal activation of T and B cells, which have been traditionally focused on, podocytes, as key components of the glomerular filtration barrier, are directly involved in the development of proteinuria and the process of glomerulosclerosis through their structural and functional damage.7 Podocytes in LN show fusion, detachment, and apoptosis. The mechanism of podocyte damage involves oxidative stress imbalance. The excessive accumulation of reactive oxygen species (ROS) promotes the release of inflammatory factors through the NLRP3 inflammasome pathway, forming a vicious cycle.8,9
In recent years, ferroptosis, as a new type of regulated cell death, has gradually attracted attention for its role in lupus nephritis (LN). Ferroptosis is characterized by the iron-dependent accumulation of lipid peroxidation and is mainly regulated by glutathione peroxidase 4 (GPX4) and the cystine/glutamate antiporter (composed of SLC7A11 and SLC3A2).10 In LN, podocytes, as a key component of the glomerular filtration barrier, are closely related to proteinuria and glomerulosclerosis when damaged.11 Previous studies have found that in the LN podocyte injury model, the expression of GPX4 is down-regulated, and the lipid peroxidation products MDA and ROS are significantly increased, suggesting that podocyte ferroptosis is involved in the progression of LN.12 In addition, the abnormal expression of ferroptosis-related genes ACSL4 and FSP1 further exacerbates podocyte injury.13 Therefore, targeting and inhibiting podocyte ferroptosis may become a new direction for the treatment of LN. Existing studies have shown that the PI3K/AKT/Nrf2 axis plays an important role in various diseases, especially in inhibiting ferroptosis.14 Activating the PI3K/AKT pathway can promote the nuclear translocation of Nrf2, thereby up-regulating the expression of antioxidant enzymes and inhibiting the occurrence of ferroptosis.15 Specifically, the activation of the PI3K/AKT pathway can phosphorylate Nrf2, releasing it from the Keap1-Nrf2 complex, allowing it to enter the nucleus and bind to the ARE, initiating the transcription of downstream antioxidant genes. These antioxidant genes include GPX4, SLC7A11, GSH synthetase, etc., which can scavenge lipid peroxides and ROS, thus inhibiting the occurrence of ferroptosis.16 However, there is still a lack of systematic research on the specific role of the PI3K/AKT/Nrf2 axis in podocyte ferroptosis in LN.
Astragalus membranaceus is a medicinal herb commonly used in traditional Chinese medicine for immune regulation and kidney protection. Its active ingredient, astragaloside IV (AS-IV), belongs to the cycloartane-type triterpenoid saponins and has various pharmacological effects such as anti-inflammation, anti-oxidation, and anti-fibrosis.17 Previous studies have shown that AS-IV can regulate the ITGB1/PTK2/p38 axis and inhibit the immune inflammatory response, thereby alleviating the progression of SLE. However, its effect on podocyte ferroptosis has not been elucidated.18 Notably, AS-IV has been shown to activate the PI3K/AKT pathway in kidney damage, suggesting that it may intervene in the ferroptosis process in LN through a similar mechanism.19 In addition, some studies have found that AS-IV effectively alleviates diabetic nephropathy through the SIRT1/PGC1α/Nrf1 axis, reducing phenyl sulfate-induced oxidative stress and mitochondrial dysfunction in podocytes.20 However, there is currently no direct evidence indicating whether AS-IV regulates podocyte ferroptosis through the PI3K/AKT/Nrf2 axis.
Based on the above background, this study aims to explore the therapeutic potential of AS-IV for LN and its molecular mechanism. We hypothesize that AS-IV can upregulate the expression of GPX4 and SLC7A11 by activating the PI3K/AKT/Nrf2 pathway, inhibit podocyte ferroptosis, and thereby alleviate renal injury in LN. To verify this hypothesis, we first used MRL/lpr mice as a classic SLE model to evaluate the effects of AS-IV on renal histopathology and ultrastructure, and detected the degree of renal damage through biochemical indicators. Secondly, we revealed the potential molecular targets of AS-IV in the intervention of LN through network pharmacology and molecular docking techniques. Finally, we constructed an in vitro model of ferroptosis in MPC-5 cells induced by puromycin aminonucleoside (PAN).21 We evaluated the effects of AS-IV on ferroptosis in MPC-5 cells and the PI3K/AKT/Nrf2 pathway through various experimental techniques. In conclusion, this study not only provides a theoretical basis for the treatment of LN with AS-IV, but also opens up new ideas for the development of natural drugs targeting ferroptosis.
Materials and Methods
Animals and Grouping
Female MRL/lpr mice and C57BL/6 mice, aged 7 weeks and weighing 18–22 g, were purchased from Shanghai Slack Laboratory Animal Co., Ltd., with the license number SCXK (Shanghai) 2022–0004. All animals were housed in the SPF-grade conditions of the Animal Experiment Platform of the Artificial Intelligence Research Institute of Hefei Comprehensive National Science Center. The housing temperature was maintained at 25 ± 2 °C, the relative humidity at 50 ± 10%, and free access to water and food was provided. A 12 h light/dark cycle was maintained. All mice were housed in a uniform environment for 7 weeks before the experiment began. All animal experiments were strictly conducted in accordance with the “Regulations on the Administration of Laboratory Animals of the People’s Republic of China” and the “Ethical Guidelines” and were approved by the Animal Ethics Committee of Anhui University of Chinese Medicine (Approval No.: AHUCM-mouse-2024200). Based on the significance level (α= 0.05) and test power (1-β = 0.8), by referring to relevant literature and pre-experimental data, and using the G*Power software to calculate the experimental sample size, the minimum sample size for each group is 8. Fifty MRL/lpr mice were randomly divided into five groups: MRL/lpr group (Model, n = 10), low-dose AS-IV group (AS-IV-L, n = 10), medium-dose AS-IV group (AS-IV-M, n = 10), high-dose AS-IV group (AS-IV-H, n = 10), and prednisone group (Pred, n = 10). Ten C57BL/6 mice served as the blank control group (Control). Mice in the Pred group were given intragastric administration of 4.55 mg/kg/d prednisone (purchased from Tianjin Tianyao Pharmaceutical Co., Ltd., Batch No.: 20,250,511). Mice in the AS-IV-L, AS-IV-M, and AS-IV-H groups were given intragastric administration of 10 mg/kg/d, 20 mg/kg/d, and 40 mg/kg/d of AS-IV (purchased from Shanghai Yuanye Biotechnology Co., Ltd., Batch No.: 2,025,212,005), respectively. Mice in the Control group and the MRL/lpr group were given an equal volume of normal saline by intragastric administration. Intragastric administration was performed once a day for 8 weeks. During the intervention period, the urine of the mice was collected weekly, and the body weight of the mice was measured. After 8 weeks of administration, the mice were anesthetized by intraperitoneal injection of 3% sodium pentobarbital. Subsequently, euthanasia was carried out by inhaling carbon dioxide (the initial gas flow rate was maintained at the container volume × 50%/min), and death was confirmed by observing the cessation of breathing. Finally, the kidney tissues of mice in each group were collected.
Histopathology
Kidney tissues were fixed in 4% paraformaldehyde (Sigma, Cat. No.: P6148) for 48 h, then rinsed with running water. They were dehydrated with ethanol, cleared in xylene (Sigma, Cat. No.: 247,865), embedded in paraffin, and then subjected to sectioning and dewaxing procedures. The sections were stained with hematoxylin-eosin (HE)22 and periodic acid-Schiff (PAS)23 respectively, and observed and recorded under an optical microscope (Olympus, CX3 - URA2). Meanwhile, the glomerular score24 and perivascular wall score25 were calculated. Glomerular scoring calculation method: Randomly select 15 glomeruli for observation and scoring. The scoring indicators include cell proliferation, basement membrane thickening, crescent formation, and degree of sclerosis. Then calculate the average value of the scores of these glomeruli, which is the glomerular score of the kidney tissue. Calculation method for perivascular wall score: Select 10 blood vessels for observation and scoring. The scoring indicators include inflammatory cell infiltration, blood vessel wall thickening, degree of fibrosis, and thrombus formation. Calculate the average score of these blood vessels.
Observation of the Ultrastructure of Glomeruli by Transmission Electron Microscopy
Place the kidney tissue in 2.5% glutaraldehyde (Thermo Fisher, Cat. No.: 16310) for the first fixation, then transfer it to a 1% osmium tetroxide solution (Ted Pella, Cat. No.: 18451) for secondary fixation. After that, perform gradient ethanol dehydration, resin infiltration, and embedding to prepare ultrathin sections. Stain the sections with an aqueous solution of uranyl acetate (Polysciences, Cat. No.: 24,300–1) for 30 min and lead citrate (Ted Pella, Cat. No.: 18310) for 5 min. Then, thoroughly rinse with distilled water to avoid stain crystallization. Observe and record the glomerular structure using a transmission electron microscope (JEOL, JEM-2100). Meanwhile, calculate the foot - process fusion rate and the deposition density of electron-dense materials.26
Biochemical Kit Detection
Collect the urine of mice. Use the urine protein quantitative kit (Nanjing Jiancheng Bioengineering Institute, catalog number: C035-2-1) to detect the 24-hour urine protein quantification (24hPRO), the total urine protein kit (Nanjing Jiancheng Bioengineering Institute, catalog number: A045-4) to detect the total urine protein content, the urine albumin kit (Nanjing Jiancheng Bioengineering Institute, catalog number: A028-2-1) to detect the urine albumin, and the urine creatinine kit to detect the urine creatinine content. Calculate the urine total protein-creatinine ratio (UTPCR) and the urine albumin-creatinine ratio (UACR). The specific operation process refers to the kit instructions.
Cell Culture and PAN Stimulation
The mouse immortalized podocyte cell line MPC-5 was purchased from the Cell Bank of the Chinese Academy of Sciences (Catalog No.: KCB200678Y). The cryopreserved MPC-5 cell tube was quickly taken out from the liquid nitrogen and immediately placed in a 37°C water bath, and gently shaken until it was completely thawed. MPC-5 cells were routinely cultured in high-glucose DMEM medium containing 10% fetal bovine serum and 1% penicillin/streptomycin in a constant temperature and humidity incubator at 37°C with 5% CO2. When the cells reached 80–90% confluence, subculture was carried out. An appropriate amount of 0.25% trypsin-EDTA solution (Gibco, Catalog No.: 25200056) was added to cover the cell surface, and the cells were placed in a 37°C incubator for digestion for 1–2 min. Then, an appropriate amount of cell cryopreservation solution was added to resuspend the cells. The cell suspension was aliquoted into cryopreservation tubes, placed in a −80°C refrigerator overnight, and then transferred to liquid nitrogen for long-term storage. MPC-5 cells were stimulated with different concentrations of puromycin aminonucleoside (PAN, Sigma-Aldrich, Catalog No.: P7130-25MG) for 24 h to screen the optimal stimulation concentration of PAN for subsequent experiments.
Cell Viability Was Detected by CCK-8 Assay
The effects of different treatments on the viability of MPC-5 cells were evaluated using the CCK-8 method. MPC-5 cells were resuspended in 0.25% trypsin (Shanghai Beyotime Biotechnology Co., Ltd., Cat. No.: C0201), and the density was adjusted to 5×104 cells. The cells were seeded into 96-well plates, with 100 μL of cell suspension added to each well. The 96-well plates were placed in a CO2 incubator and incubated at 37°C with 5% CO2 for 24 h. Six replicate samples were set for each condition. During the last 2 hours of culture, 10 μL of CCK-8 solution (Dojindo, Cat. No.: CK04) was added to each well, and the absorbance of each well was measured at OD 450 nm using a microplate reader (Rayto, RT-6100).
Determination of Intracellular Fe2+
The Fe2+ levels in each group of cells were measured using the ferrous ion colorimetric assay kit (Elabscience, Catalog No.: E-BC-K773-M). The specific operations are described in the kit instructions. The OD values of each well were measured at 593 nm using a microplate reader, and the Fe2+ content in MPC-5 cells was calculated.
Cellular ROS Measurement
Collect MPC-5 cells in the logarithmic growth phase and wash them twice with PBS. Digest the cells with trypsin and resuspend them in serum-free medium. Dilute DCFH-DA (Beyotime, Cat. No.: S0033S) to 10 μM and add it to the serum-free medium. Add 1 mL of the staining solution to each group of cells, mix well, and incubate at 37°C for 30 min. Resuspend the cells in 500 μL of PBS. Set up the flow cytometer (Beckman, CytoFLEX), use a 488 nm laser for excitation, and detect the fluorescence signal with a 530/30 nm filter. Analyze the data using FlowJo_V10 software and calculate the fluorescence intensity of ROS-positive cells.
Determination of Intracellular MDA, GSH and SOD
Collect MPC-5 cells from each group, lyse the cells and centrifuge to collect the supernatant. Then, use the MDA kit (Nanjing Jiancheng Bioengineering Institute, catalog number: A003-1), GSH kit (Nanjing Jiancheng Bioengineering Institute, catalog number: A006-2-1), and SOD kit (Nanjing Jiancheng Bioengineering Institute, catalog number: A001-3) to detect the contents of MDA, GSH, and SOD respectively. The specific operation process is detailed in the kit instructions.
Network Pharmacology and Molecular Docking
The potential target genes of Astragaloside IV were sourced from the SwissTargetPrediction database (http://swisstargetprediction.ch/)27 and the PharmMapper database (http://lilab-ecust.cn/pharmmapper/submitfile.html).28 The target genes of lupus nephritis were collected from the DisGeNET database (https://www.disgenet.org/),29 the GeneCards database (https://www.genecards.org/),30 and the OMIM database (https://www.omim.org/).31 The STRING (https://cn.string-db.org/) was used to construct the PPI network, and the visualization was performed through Cytoscape (V3.10.2). The Metascape database (http://metascape.org)32 was used for gene ontology (GO) analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. The molecular structures of PI3K (PDB: 1E8U), AKT (PDB: 3O96), and Nrf2 (PDB: 5CGJ) were downloaded from the PDB database (https://www.rcsb.org/),33 respectively, and the three - dimensional structure of Astragaloside IV was downloaded from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). The AutoDock Vina software (V1.5.7, The Scripps Research Institute) was used for molecular docking, and the PyMOL software (V2.5.4, DeLano Scientific LLC) was used to visualize the docking results.
Cell Transfection
After digesting and counting MPC-5 cells, seed them into a 6-well cell culture plate at a density of 5×104 cells per well. Add 2 mL of DMEM medium containing 10% FBS to each well and culture the cells in an incubator for 24 hours to ensure that the cell confluence reaches 70% at the time of transfection. For the overexpression group, add 50 μL of Opti-MEM serum-free medium and 2 μg of plasmid to each well (the experimental group uses the pcDNA3.1-P53 plasmid, and the control group uses the pcDNA3.1 empty vector plasmid). For the knockdown group, add 50 μL of Opti-MEM serum-free medium and 50 nM siRNA to each well (the experimental group uses siRNA-PI3K, and the control group uses non-specific siRNA). Disperse the mixture evenly and slowly drop it onto the cells in the 6-well plate. Then place the 6-well plate in a 37°C incubator for 5 h. Aspirate the medium containing the transfection complex and replace it with fresh complete medium, and continue culturing for 24 h.
Detect Protein Levels by Western Blotting
Collect MPC-5 cells from each group, add 5X SDS-PAGE protein loading buffer. Place the cooled protein samples into the loading wells of the SDS-PAGE gel and conduct protein electrophoresis for 1 h. Incubation with primary antibodies: Add p-PI3K (Affinity, AF3242, 1:1000), PI3K (Affinity, AF6241, 1:1000), AKT (CST, 4691S, 1:1000), P-AKT (CST, 4060S, 1:1000), Nrf2 (Bioss, bs-1074R, 1:500), GPX4 (abclonal, A1933, 1:50,000), Synaptophysin (bioss, bsm-52379R, 1:1000), Nephrin (Affinity, DF7951, 1:3000), Podocin (bioss, bs-6597R, 1:1000), α-actinin-4 (bioss, bs-1741R, 1:1000), WT-1 (bioss, bs-6983R, 1:1000), SLC7A11 (Proteintech, 26864-1-AP, 1:4000), and GAPDH (Zsbio, TA-08, 1:5000). Then add the HRP-labeled secondary antibody (1:20000) and incubate at room temperature for 1.2 h. Use the ECL chemiluminescence kit (Biosharp, BL520A) to take photos with a chemiluminescence imaging system and save them. Use Image J software for gray - scale analysis.
Detection of Relative Gene Expression Levels by RT-qPCR
Collect the cell pellet, add 1 mL of TRIzol for lysis, perform layering and RNA precipitation to complete RNA extraction. Conduct an RT reaction to obtain cDNA. Use a PCR instrument to perform RT-qPCR reaction detection. The relative expression level of mRNA is calculated using the 2−ΔΔCt method. All experiments were repeated three times. The primer sequences are as follows: PI3K (forward sequence: 5’- CACCTTAAATGGTGAGCACG −3’ and reverse sequence: 5’- TTCGTCAGTTCTCCTGGATG −3’), AKT (forward sequence: 5’- CCAGGGGAGGATGTTTCTAC −3’ and reverse sequence: 5’- GGTCGCGTCAGTCCTTAATA −3’), GPX4 (forward sequence: 5’- TCCGCCTCTAAGTTCTTGTC −3’ and reverse sequence: 5’- GCGGAGAACTAGGAGATAGC −3’), β-actin (forward sequence: 5’- AGTGTGACGTTGACATCCGT −3’ and reverse sequence: 5’- TGCTAGGAGCCAGAGCAGTA −3’).
Immunofluorescence Staining
Fix the cell samples, then perform antigen retrieval. Add 0.3% Triton X-100 (Ebiogo, Catalog No.: B025) for permeabilization and blocking at room temperature. Add the primary antibodies dropwise: IgG (Wuhan Sanying, 30000-0-AP, 1:500), C3 (Affinity, DF13224, 1:200), SLC7A11 (abclonal, A2413, 1:200), DHODH (Bioss, bs-14305R, 1:500), GPX4 (Wuhan Sanying, 67763-1-Ig, 1:2000), FSP1 (abclonal, A22278, 1:500), and incubate at 37°C for 60 min Add the secondary antibody dropwise: add an appropriate amount of HRP - labeled secondary antibody reagent (Fuzhou Maixin, Catalog No.: KIT5030). Add an appropriate amount of DAPI staining solution to counter-stain the sections. Observe and collect images under a fluorescence microscope (Olympus, BX43).
Statistical Analysis
GraphPad Prism V9.5.1 (GraphPad Software) and SPSS Statistics V27.0 (IBM Corp.) softwares were used for data statistical analysis and visualization. Each experiment was independently repeated at least three times. Measurement data were expressed as mean ± standard deviation. The Shapiro–Wilk test and Levene’s test were used to assess the normality and homogeneity of variance of quantitative data, respectively. A two-tailed Student’s t-test was used to compare the two groups, as the data were normally distributed. When the data from multiple groups met the assumptions of normality and homogeneity of variance, a one-way analysis of variance (ANOVA) with Dunnett’s post-hoc test was applied. If these assumptions were violated, the Kruskal–Wallis test was employed, followed by Dunn’s post-hoc test for pairwise comparisons. A P-value < 0.05 was considered statistically significant.
Results
Astragaloside IV Can Improve Renal Damage in MRL/Lpr Mice
To explore the improvement effect of Astragaloside IV on kidney damage in MRL/lpr mice. In vivo animal experiments were conducted (Figure 1A). HE and PAS staining were used to observe the histopathology of kidney tissues, transmission electron microscopy was used to observe the ultrastructure of glomeruli, and immunofluorescence was used to detect the deposition of IgG and C3 in the kidneys. HE staining showed that in the Model group, there was obvious intracapillary cell proliferation and mesangial proliferation, accompanied by a large number of lymphocyte infiltrations. In the AS-IV-L, AS-IV-M, AS-IV-H groups and the Pred group, cell proliferation and mesangial thickening were alleviated compared with the Model group, and lymphocyte infiltration was reduced. PAS staining showed that in the Model group, there was obvious basement membrane thickening and the formation of many crescents. In the AS-IV-L, AS-IV-M, AS-IV-H groups and the Pred group, basement membrane thickening was alleviated compared with the Model group, and the distribution of crescents was reduced. Among them, the Pred group had the best intervention effect (Figure 1B). Transmission electron microscopy results showed that in the Control group, glomerular mesangial cells were star-shaped with multiple protrusions of different lengths, and the structures of dense plaques and dense bodies could be seen. In the Model group, electron-dense deposits were observed in the glomerular mesangial area, accompanied by mesangial cell proliferation and matrix increase. After Astragaloside IV intervention, electron-dense deposits in the glomerular mesangial area and mesangial cell proliferation were alleviated compared with the Model group (Figure 1C). At the same time, Astragaloside IV could reduce the glomerular score, perivascular wall score, foot process fusion rate and electron - dense deposit density in MRL/lpr mice (P < 0.05, P < 0.01, Figure 1D–G). The results of biochemical kit detection showed that Astragaloside IV reduced the levels of 24hPRO, UTPCR and UACR in MRL/lpr mice (P < 0.01, Figure 2A–C). Immunofluorescence results showed that Astragaloside IV could reduce the deposition of IgG and C3 in the kidneys of MRL/lpr mice (P < 0.05, P < 0.01, Figure 2D and E). The above results indicate that Astragaloside IV can improve the degree of kidney damage in MRL/lpr mice, and its intervention effect is comparable to that of prednisone.
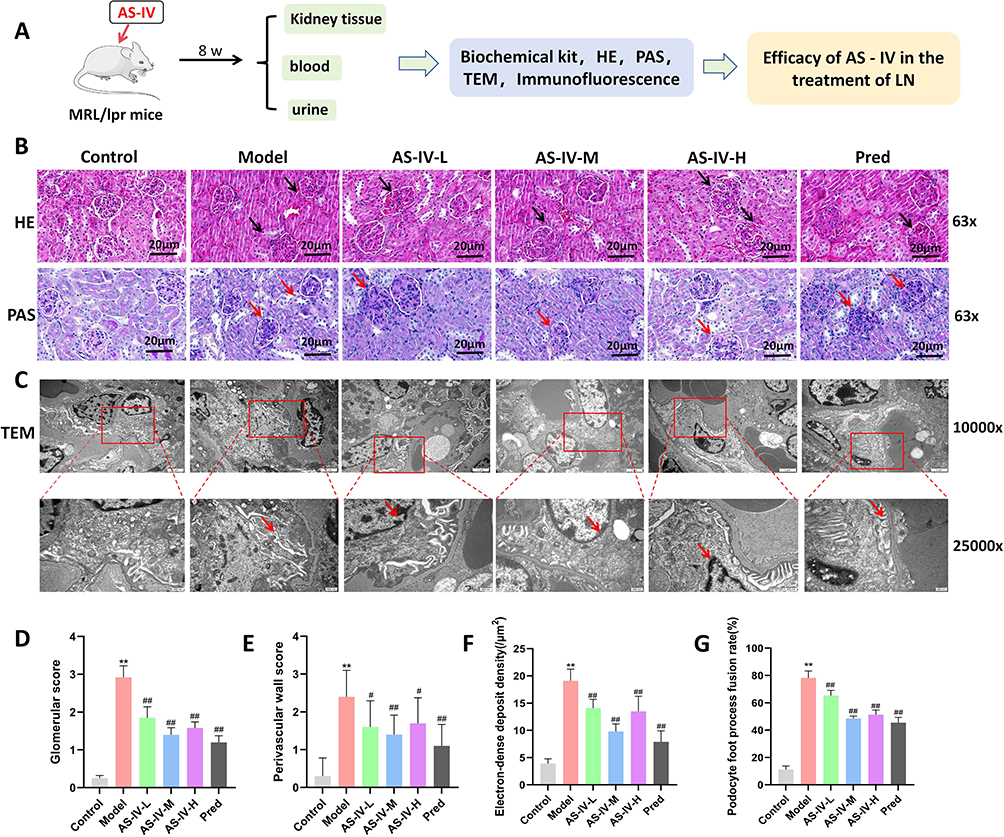 |
Figure 1 Effects of Astragaloside IV on renal histopathology and ultrastructure in MRL/lpr mice. (A) Flow chart of in vivo experiments. (B) HE staining and PAS staining of renal tissues in each group of mice. (C) Transmission electron microscopy was used to observe the ultrastructure of glomeruli in each group of mice. (D and E) Glomerular scores and perivascular wall scores were calculated (n = 10). (F and G) The deposition density of electron-dense materials and the foot process fusion rate were calculated (n = 10). The red and black arrows represent the lesion sites of the glomeruli, and the red marked box represents the magnified area. Compared with the Control group, **P < 0.01; compared with the Model group, ##P < 0.01, #P < 0.05. All data were from at least three repeated experiments, and each repeated experiment was independently detected three times. |
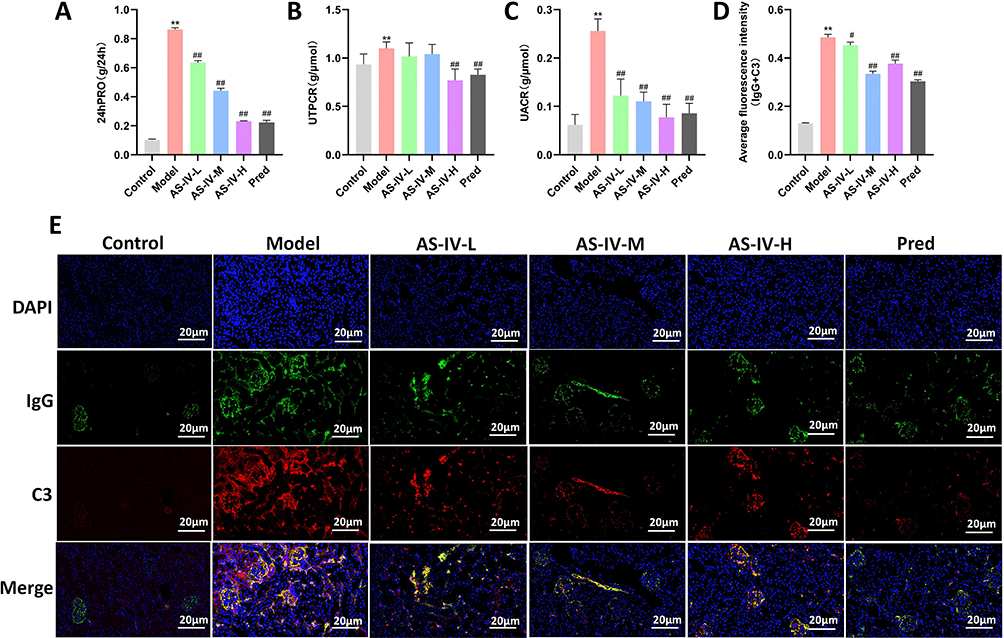 |
Figure 2 Effects of Astragaloside IV on renal damage-related indicators and IgG and C3 deposition in MRL/lpr mice. (A–C) Biochemical kits were used to detect 24hPRO, UTPCR, and UACR in mice of each group (n = 10). (D and E) Immunofluorescence staining was used to detect the deposition of IgG and C3 in renal tissues (n = 3). Compared with the Control group, **P < 0.01; compared with the Model group, ##P < 0.01, #P < 0.05. All data were from at least three repeated experiments, and each repeated experiment was independently detected three times. |
Network Pharmacological Analysis of the Potential Targets of Astragaloside IV in Lupus Nephritis (LN)
To explore the molecular mechanism of Astragaloside IV in intervening in LN, network pharmacology and molecular docking techniques were used for analysis. It was found that there were 35 intersection targets between Astragaloside IV and LN. Through PPI network analysis, 13 core targets were obtained, namely AKT1, PI3KCA, NFE2L2, KEAP1, etc. (Figure 3A and B). The results of KEGG and GO enrichment analysis showed that the mechanism of action of Astragaloside IV was related to cell ferroptosis, chemokine activity, and the PI3K/AKT pathway (Figure 3C and D). Finally, molecular docking technology was used to analyze the binding ability of Astragaloside IV with PI3K, AKT, and Nrf2. The results showed that the lowest binding energies of Astragaloside IV with PI3K, AKT, and Nrf2 were −8.8 kcal/mol, −8.0 kcal/mol, and −10.9 kcal/mol respectively (Figure 3E), suggesting that Astragaloside IV had strong binding ability with PI3K, AKT, and Nrf2. The above results indicate that Astragaloside IV can act on the PI3K/AKT/Nrf2 signaling axis.
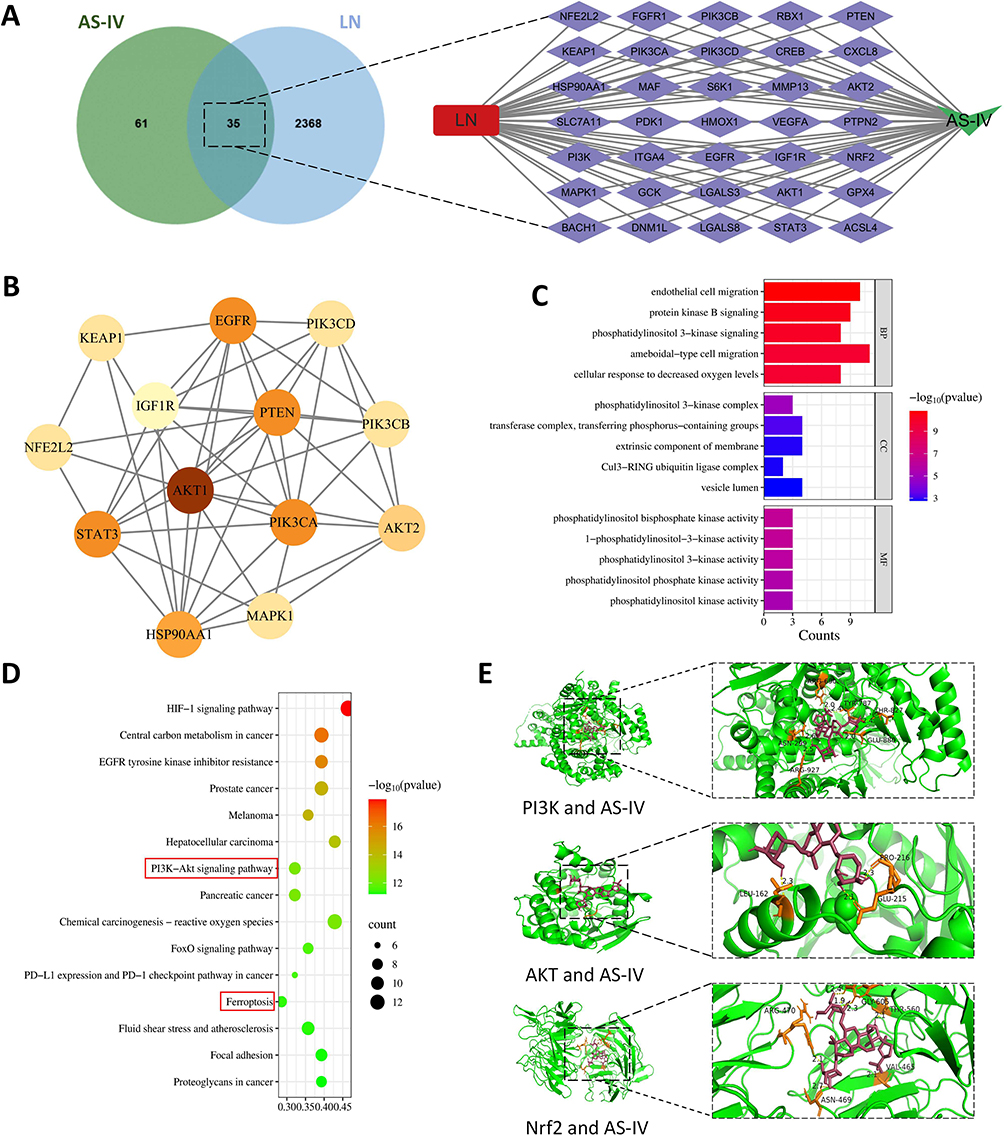 |
Figure 3 Potential targets of Astragaloside IV acting on LN analyzed by network pharmacology. (A) Common targets of Astragaloside IV and LN and the drug-disease-target network diagram. (B) Construction of the PPI network and screening of core targets. (C and D) Results of GO and KEGG analyses. (E) Molecular docking results of Astragaloside IV with PI3K, AKT, and Nrf2. |
Effects of Astragaloside IV on MPC-5 Podocyte Injury and Oxidative Stress
In the in vitro experiment, MPC-5 podocytes were used as the research object, and the specific process is shown in Figure 4A. First, to screen the optimal concentration of PAN for inducing MPC-5 podocytes, the CKK-8 method was used to detect the cell viability after 24 hours of induction with different concentrations (0, 15, 25, 50, 75 μg/mL) of PAN. The results showed that the optimal induction concentration of PAN was 50 μg/mL (Figure 4B). The CKK-8 method was used to detect the cell viability after 24 hours of intervention with different concentrations (0, 20, 40, 60, 80 μM) of Astragaloside IV. The results showed that the optimal intervention concentration of Astragaloside IV was 40 μM (Figure 4C). The cells were divided into four groups, in which Ferrostatin-1 (Fer-1), an inhibitor of ferroptosis, was used as the positive control group. The levels of proteins related to MPC-5 podocyte injury were detected by Western blotting. The results showed that after the intervention of Astragaloside IV and Fer-1, the expressions of Nephrin, Podocin, Synaptopodin, WT-1, and α-actinin-4 proteins were increased (P < 0.05, P < 0.01, Figure 4D–I). The SOD level was detected using an ELISA kit, and the ROS level was detected by flow cytometry. The results showed that Astragaloside IV increased the SOD level and decreased the ROS level (P < 0.01, Figure 4J–L), which was consistent with the intervention effect of Fer-1. The above results indicate that Astragaloside IV can alleviate MPC-5 podocyte injury and oxidative stress levels.
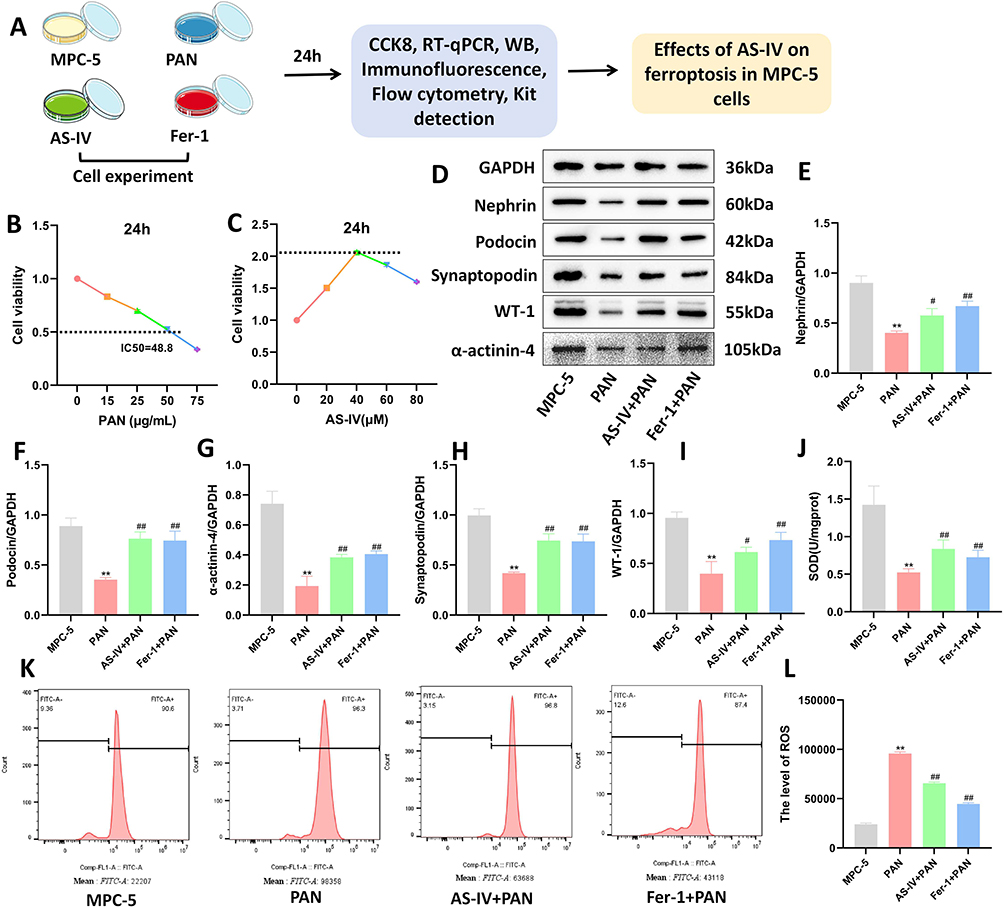 |
Figure 4 Effects of Astragaloside IV on MPC-5 podocyte injury and oxidative stress. (A) Flow chart of in vitro cell experiments. (B) Screening of the optimal concentration of PAN for inducing MPC-5 cells. (C) Screening of the optimal intervention concentration of Astragaloside IV. (D–I) Western blotting was used to detect the expression levels of proteins related to MPC-5 podocyte injury (n = 10). (J) ELISA kit was used to detect the intracellular SOD level (n = 10). (K and L) Flow cytometry was used to detect the intracellular ROS level (n = 3). Compared with the MPC-5 group, **P < 0.01; compared with the PAN group, ##P < 0.01, #P < 0.05. All data were from at least three repeated experiments, and each repeated experiment was independently detected three times. |
Effects of Astragaloside IV on the PI3K/AKT/Nrf2 Axis and Ferroptosis in MPC-5 Podocytes
To explore the effects of Astragaloside IV on the PI3K/AKT/Nrf2 axis and ferroptosis in MPC-5 podocytes. Western blotting, immunofluorescence, and ELISA kits were used to detect the expression of factors related to the PI3K/AKT/Nrf2 axis and ferroptosis in MPC-5 podocytes. The results showed that, compared with the MPC-5 group, after PAN induction, the levels of factors related to the intracellular PI3K/AKT/Nrf2 axis, GPX4, SLC7A11, DHODH, FSP1, and GSH decreased, while the levels of Fe2+, and MDA increased. After Astragaloside IV intervention, the levels of p-PI3K, p-AKT, Nrf2, GPX4, DHODH, FSP1, and SLC7A11 increased (P < 0.05, P < 0.01, Figures 5A–H). Meanwhile, Astragaloside IV could reduce the levels of Fe2+ and MDA in renal podocytes and increase the level of GSH (P < 0.01, Figures 5I–K). The above results indicate that Astragaloside IV has a certain regulatory effect on the PI3K/AKT/Nrf2 axis and can inhibit ferroptosis in MPC-5 podocytes.
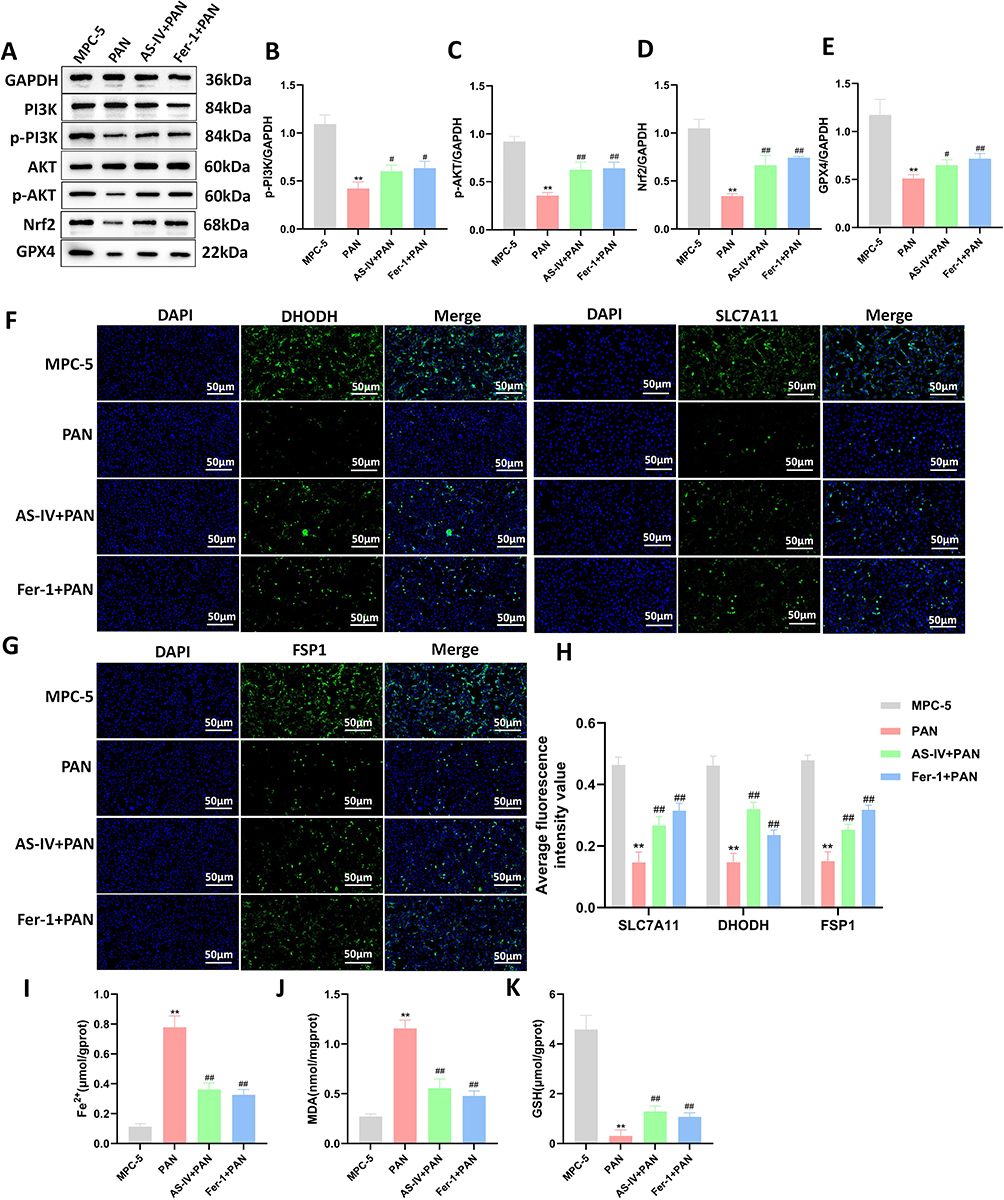 |
Figure 5 Effects of Astragaloside IV on the PI3K/AKT/Nrf2 axis and ferroptosis in MPC-5 podocytes. (A–E) Western blotting was used to detect the expression levels of PI3K, p-PI3K, AKT, p-AKT, Nrf2, and GPX4 proteins (n = 10). (F–H) Immunofluorescence staining was used to detect the expression levels of DHODH, SLC7A11, and FSP1 in MPC-5 cells of each group (n = 3). (I) The intracellular Fe2+ content (n = 10). (J and K) ELISA kits were used to detect the intracellular MDA and GSH levels (n = 10). Compared with the MPC-5 group, **P < 0.01; compared with the PAN group, ##P < 0.01, #P < 0.05. All data were from at least three repeated experiments, and each repeated experiment was independently detected three times. |
Overexpression of PI3K Can Enhance the Inhibitory Effect of Astragaloside IV on Ferroptosis of MPC-5 Podocytes
To further explore whether PI3K is the key target of Astragaloside IV, pcDNA3.1-PI3K plasmid was transfected into MPC-5 cells on the basis of Astragaloside IV intervention. The cells were divided into PAN group, AS-IV + PAN group, AS-IV + PAN + pc3.1-NC group, and AS-IV + PAN + pc3.1-PI3K group. The levels of Fe2+, MDA, GSH, and SOD in each group of cells were detected using an ELISA kit. The results showed that compared with the AS-IV + PAN + pc3.1-NC group, after overexpression of PI3K, the level of GSH increased, while the levels of Fe2+ and MDA decreased (P < 0.01, Figures 6A–D). The level of ROS was detected by flow cytometry. After overexpression of PI3K, the intracellular ROS level decreased (P < 0.01, Figures 6E and F). Immunofluorescence staining was used to detect the expression of GPX4 and FSP1. The results showed that after overexpression of PI3K, the expression of GPX4 and FSP1 increased (P < 0.01, Figures 6G and H). Western blotting was used to detect the expression of proteins related to the PI3K/AKT/Nrf axis. The results showed that compared with the AS-IV + PAN + pc3.1-NC group, after overexpression of PI3K, the levels of p-PI3K, p-AKT, Nrf2, and SLC7A11 increased (P < 0.01, P < 0.05, Figures 6I–M). RT-qPCR was used to detect the expression of genes related to the PI3K/AKT/Nrf axis. The results showed that compared with the AS-IV + PAN + pc3.1-NC group, the mRNA levels of PI3K, AKT, and Nrf2 in the AS-IV + PAN + pc3.1-PI3K group increased (P < 0.01, Figure 6N). The above results indicate that overexpression of PI3K can enhance the inhibitory effect of Astragaloside IV on ferroptosis of MPC-5 podocytes.
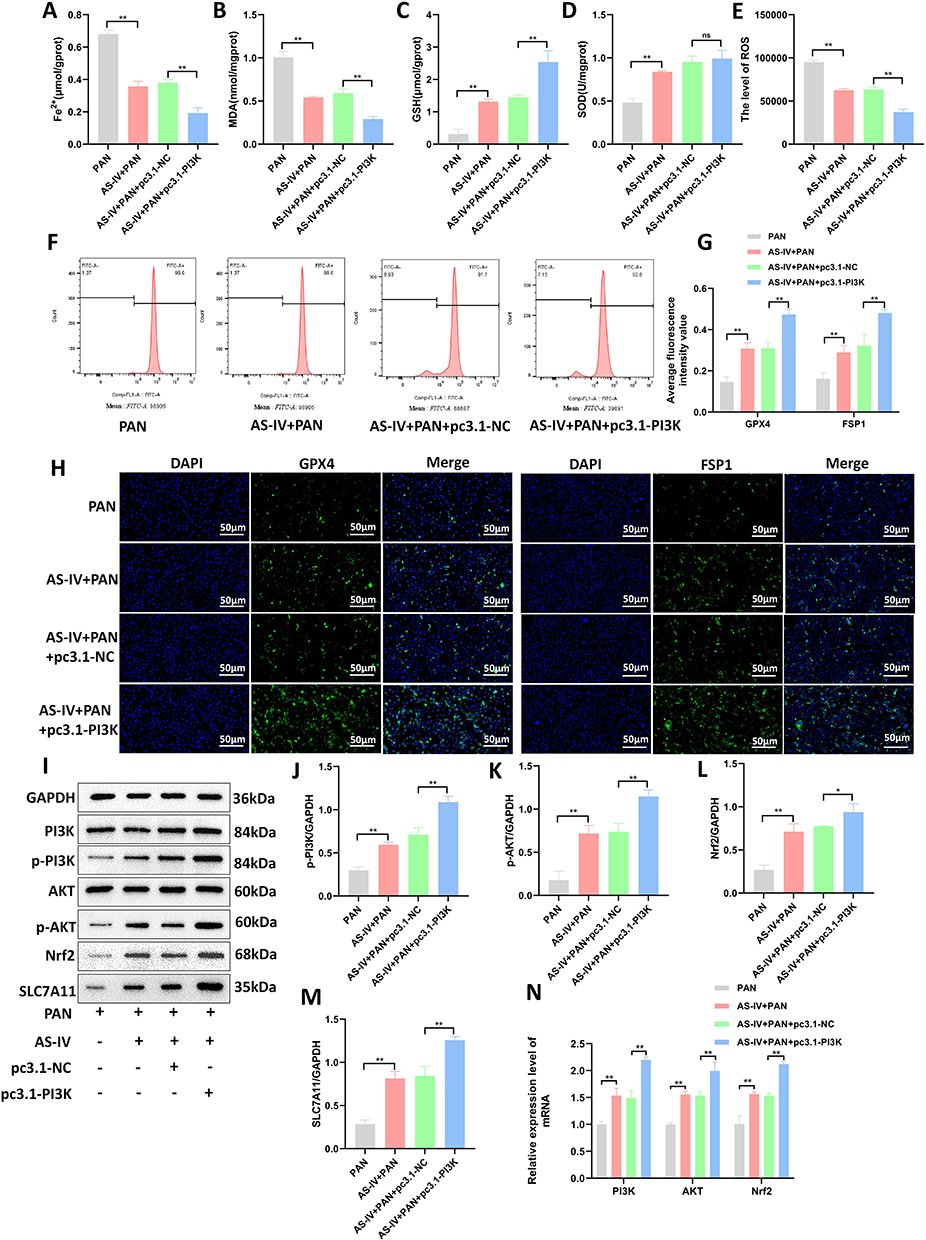 |
Figure 6 Overexpression of PI3K can enhance the inhibitory effect of Astragaloside IV on ferroptosis of MPC-5 podocytes. (A) Intracellular Fe2+ content (n = 10). (B–D) ELISA kits were used to detect the levels of MDA, GSH, and SOD in cells (n = 10). (E and F) Flow cytometry was used to detect the intracellular ROS levels (n = 3). (G and H) Immunofluorescence staining was used to detect the expression levels of GPX4 and FSP1 in MPC-5 cells of each group (n = 3). (I–M) Western blotting was used to detect the expression levels of PI3K, p-PI3K, AKT, p-AKT, Nrf2, and SLC7A11 proteins (n = 10). (N) RT-qPCR was used to detect the expression levels of PI3K, AKT, and Nrf2 mRNAs (n = 10). **P < 0.01, *P < 0.05. All data were from at least three repeated experiments, and each repeated experiment was independently detected three times. |
Knockdown of PI3K Can Reverse the Inhibitory Effect of Astragaloside IV on Ferroptosis of MPC-5 Podocytes
To further verify whether the treatment of Astragaloside IV affects ferroptosis of MPC-5 podocytes by up-regulating the expression of PI3K, we transfected siRNA-PI3K into MPC-5 podocytes on the basis of Astragaloside IV intervention. Immunofluorescence staining was used to detect the levels of SLC7A11 and DHODH in each group of cells. The results showed that compared with the AS-IV + PAN + siRNA-NC group, after knocking down PI3K, the level of SLC7A11 and DHODH decreased (P < 0.01, Figures 7A and B). Flow cytometry was used to detect the level of intracellular ROS, and ELISA kits were used to detect the levels of Fe2⁺, MDA, GSH, and SOD. The results showed that compared with the AS-IV + PAN + siRNA-NC group, the levels of ROS, Fe2⁺, and MDA in the AS-IV + PAN + siRNA-PI3K group increased, while the levels of GSH and SOD decreased (P < 0.01, Figures 7C–H). Western blotting was used to detect the expression of proteins in the PI3K/AKT/Nrf2 axis, and SLC7A11 in each group of cells. The results showed that after knocking down PI3K, the levels of p-PI3K, p-AKT, Nrf2, and SLC7A11 proteins in MPC-5 podocytes decreased (P < 0.01, P < 0.05, Figures 7I–M). The results of RT-qPCR showed that after knocking down PI3K, the relative expression levels of PI3K, AKT, and Nrf2 mRNAs decreased (P < 0.01, Figure 7N). The above results indicate that knockdown of PI3K can reverse the inhibitory effect of Astragaloside IV on ferroptosis of MPC-5 podocytes. At the same time, these results also verify that Astragaloside IV can regulate the PI3K/AKT/Nrf2 axis to inhibit ferroptosis of renal podocytes.
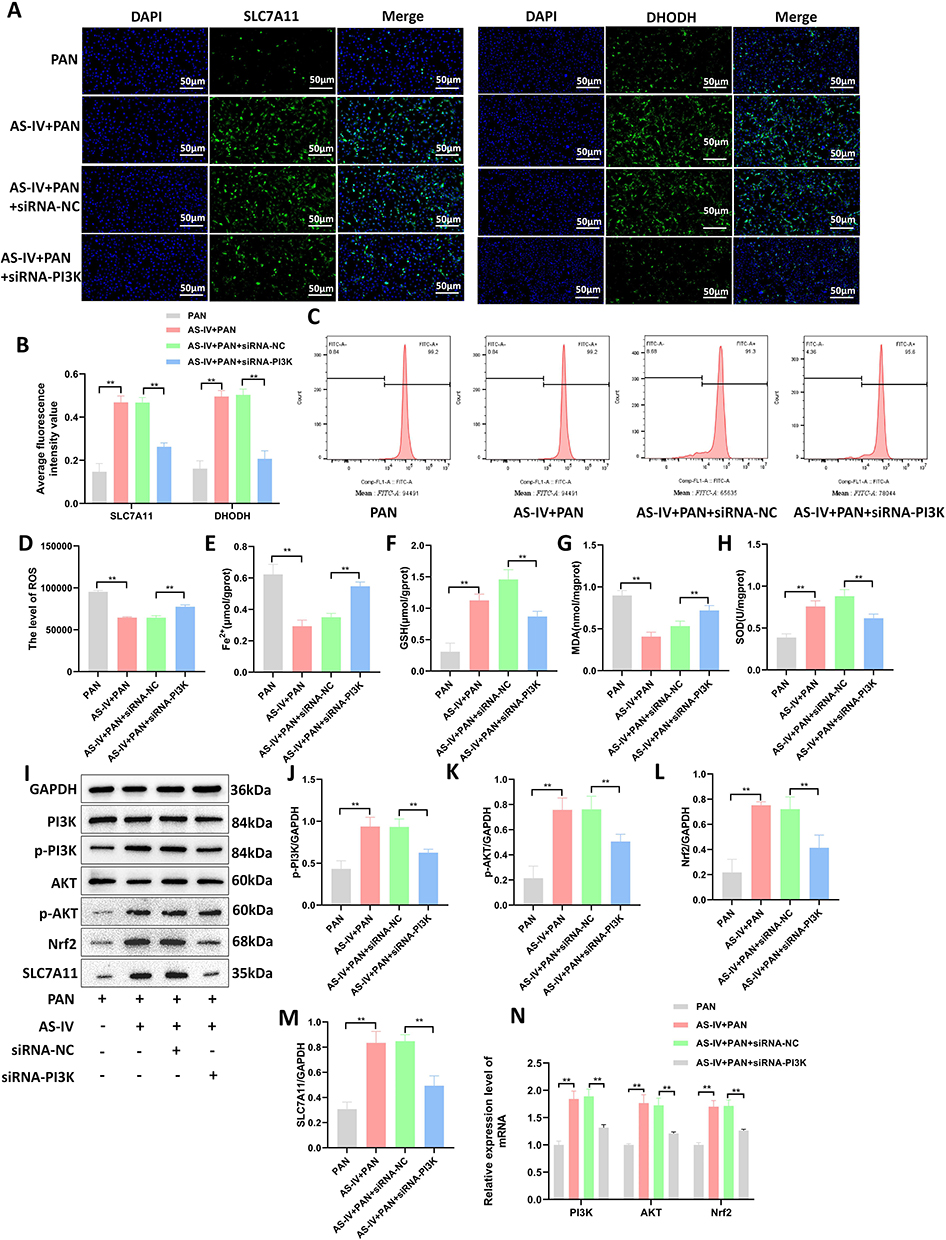 |
Figure 7 Knockdown of PI3K can reverse the inhibitory effect of Astragaloside IV on ferroptosis in MPC-5 cells. (A and B) Immunofluorescence staining was used to detect the expression levels of SLC7A11 and DHODH in MPC-5 cells of each group (n = 3). (C and D) Flow cytometry was used to detect the intracellular ROS levels (n = 3). (E) The intracellular Fe2+ content (n = 10). (F–H) ELISA kits were used to detect the levels of MDA, GSH, and SOD in cells (n = 10). (I–M) Western blotting was used to detect the expression levels of PI3K, p-PI3K, AKT, p-AKT, Nrf2, and SLC7A11 proteins (n = 10). (N) RT-qPCR was used to detect the expression levels of PI3K, AKT, and Nrf2 mRNAs (n = 10). **P < 0.01, *P < 0.05. All data were from at least three repeated experiments, and each repeated experiment was independently detected three times. |
Discussion
As one of the most severe complications of SLE, LN has complex pathological features, involving multiple mechanisms such as immune complex deposition, inflammatory cell infiltration, and damage to intrinsic glomerular cells.25,34 Although standard immunosuppressive therapy has achieved certain efficacy, some patients still face problems such as poor efficacy, high recurrence rate, and drug side effects. Therefore, finding new treatment strategies with high efficiency and low toxicity is an important direction in current LN research. This study focused on AS-IV, the main active ingredient of the traditional Chinese medicine Astragalus membranaceus. Through in vitro and in vivo experiments, we systematically explored its therapeutic potential for LN and its molecular mechanism, and revealed the key role of AS-IV in protecting the kidneys by activating the PI3K/AKT/Nrf2 signaling axis to inhibit podocyte ferroptosis.
In this study, the therapeutic effect of AS-IV was first evaluated using the classic spontaneous model of LN, MRL/lpr mice. Mice of this strain have pathological features highly similar to those of human LN, including immune complex deposition, proliferative glomerular lesions, inflammatory cell infiltration, and significant proteinuria, etc.35 Our histopathological results (HE and PAS staining) clearly showed that AS-IV treatment significantly improved the disordered glomerular structure, mesangial matrix hyperplasia, basement membrane thickening, and vascular wall lesions, and reduced the comprehensive pathological score. Transmission electron microscopy (TEM) observation further confirmed that AS-IV could effectively reduce the degree of podocyte foot process fusion and the deposition density of electron-dense materials in the subendothelial and mesangial areas of the glomerular basement membrane (GBM). These morphological evidences directly reflect the repair effect of AS-IV on renal structural damage. Meanwhile, the biochemical indicators related to renal function (24hPRO, UTPCR, UACR) also showed a significant downward trend in the AS-IV treatment group. Proteinuria is an important marker of LN activity and the degree of renal injury. The reduction of proteinuria is consistent with the improvement of histopathology, jointly indicating the definite protective effect of AS-IV on the kidneys in LN. This finding is consistent with the protective effects of Astragalus membranaceus and its active ingredient AS-IV in kidney diseases (such as diabetic nephropathy and renal fibrosis) reported in many previous studies.36 For example, AS-IV exerts antioxidant and anti-inflammatory effects in diabetic nephropathy by inhibiting FATP2-mediated fatty acid transport, thereby reducing tubular damage.37 AS-IV can improve podocyte pyroptosis in diabetic nephropathy by regulating the SIRT6/HIF-1α pathway, thereby reducing renal injury.38 These results further support the application potential of AS-IV in the field of LN treatment. Clinically, the improvement of renal function is crucial for the prognosis of LN patients. Reducing the level of urinary protein can reduce the burden on the kidneys and delay the progression of kidney diseases. Therefore, AS-IV is expected to become an effective drug for the treatment of LN, providing a new option for improving the renal function of patients.
Ferroptosis is a newly discovered type of cell death in recent years, characterized by the accumulation of intracellular iron ions and an increase in lipid peroxidation.39 In LN, ferroptosis of podocytes may be one of the important mechanisms leading to kidney injury. As a key component of the glomerular filtration barrier, the injury and loss of podocytes are the core links in the occurrence of proteinuria and the progression of glomerulosclerosis.40 An increasing amount of evidence indicates that ferroptosis is involved in the process of podocyte injury.41,42 In this study, the PAN was used in vitro to induce the mouse podocyte cell line MPC-5 to establish a ferroptosis model. PAN is a classic podocyte toxic agent that can induce podocyte injury and apoptosis.43 However, our experimental results show that PAN treatment can also significantly induce characteristic changes of ferroptosis in MPC-5 cells, with a significant increase in the intracellular Fe2+ level, an increase in MDA content, and a down-regulation of the expression of key antioxidant enzymes GPX4 and SLC7A11, leading to the depletion of reduced glutathione (GSH) and the burst of ROS. At the same time, the expression of ferroptosis suppressor protein 1 (FSP1) and dihydroorotate dehydrogenase (DHODH) also changes significantly. The changes in these indicators together confirm that PAN has successfully induced ferroptosis in MPC-5 cells. However, this study lacks the morphological structure of MPC-5 cells under electron microscopy. In subsequent studies, the ultrastructure of MPC-5 mitochondria will be observed, which can more intuitively evaluate the effect of AS-IV in inhibiting ferroptosis. Remarkably, AS-IV treatment can effectively reverse the ferroptosis phenotype induced by PAN, significantly reducing the levels of intracellular Fe2+, MDA, and ROS; at the same time, it up-regulates the protein expression of GPX4, SLC7A11, FSP1, and DHODH, restores GSH reserves, and enhances the activity of SOD. These results strongly indicate that AS-IV effectively inhibits PAN-induced podocyte ferroptosis by multi-target regulation of the key links of ferroptosis. This discovery not only provides a new perspective for understanding podocyte injury in LN but also provides an important mechanistic explanation for the protection of podocytes by AS-IV.
To deeply explore the molecular targets of AS-IV in inhibiting podocyte ferroptosis, this study adopted a method combining network pharmacology prediction with molecular docking verification. The prediction results showed that the PI3K/AKT signaling pathway and its downstream Nrf2 may be the key targets for AS-IV to intervene in LN. Nrf2 is the main regulator of the cellular antioxidant response and can transcriptionally activate the expression of a series of antioxidant and anti-ferroptosis genes, including GPX4, SLC7A11, and HO-1.44 As an important intracellular survival signaling pathway, the activation of the PI3K/AKT pathway has been proven to phosphorylate and stabilize Nrf2, promoting its nuclear translocation and transcriptional activity.45 This study found that AS-IV can upregulate the levels of factors related to the PI3K/AKT/Nrf2 pathway in MPC-5 cells. This result indicates that AS-IV can activate PI3K, enabling it to catalyze the production of PIP3. PIP3 then recruits and activates AKT. Activated AKT can phosphorylate Nrf2, promoting its translocation from the cytoplasm to the nucleus and enhancing the binding ability of Nrf2 to the ARE, thereby upregulating the expression of antioxidant genes. GPX4 is an important antioxidant enzyme that can reduce lipid peroxides to the corresponding alcohols, thus preventing the occurrence of lipid peroxidation.46 In this study, after AS-IV treatment, the level of GPX4 in MPC-5 cells was significantly increased, indicating that AS-IV enhances the cellular antioxidant capacity and reduces the production and accumulation of ROS by activating the PI3K/AKT/Nrf2 axis. The results of this study showed that AS-IV can reduce the levels of Fe2⁺, MDA, and ROS in MPC-5 cells, while increasing the levels of GPX4, SLC7A11, DHODH, FSP1, GSH, and SOD. Among them, SLC7A11 is a cystine/glutamate antiporter that can transport extracellular cystine into the cell for the synthesis of GSH.47 GSH is a coenzyme of GPX4 and is crucial for maintaining the activity of GPX4. Sufficient GSH can provide substrates for GPX4, enabling it to better exert its antioxidant effect, inhibit the occurrence of lipid peroxidation, and further inhibit podocyte ferroptosis.48
To establish the central role of PI3K in this mechanism, we conducted a crucial rescue experiment. After specifically knocking down the expression of PI3K in MPC-5 cells using siRNA, the activation effect of AS-IV on the PI3K/AKT/Nrf2 pathway was significantly weakened, manifested as a decrease in the levels of p-AKT and Nrf2. Meanwhile, the up-regulatory effect of AS-IV on GPX4/SLC7A11 and the down-regulatory effect on Fe2+/MDA/ROS were partially or even completely reversed. This result provides direct causal evidence, demonstrating that PI3K is an indispensable upstream target for AS-IV to exert its anti-ferroptosis effect, and the PI3K/AKT/Nrf2 axis constitutes the core signaling pathway through which AS-IV inhibits podocyte ferroptosis.
This study is the first to link the protective effect of the active ingredient AS-IV from traditional Chinese medicine with ferroptosis, an emerging cell death mechanism in podocytes, and elucidate the core signaling pathway (PI3K/AKT/Nrf2) of its action, providing a brand - new molecular framework for understanding the renal protective effect of AS-IV. This not only enriches the theoretical understanding of the pathogenesis of LN, emphasizes the role of ferroptosis in podocyte injury, but also expands the pharmacological mechanism of AS-IV. As a natural compound, the advantage of AS-IV lies in its multi-target action.49 In addition to the PI3K/AKT/Nrf2/GPX4/SLC7A11 pathway focused on in this study, the known anti-inflammatory and immunomodulatory effects of AS-IV may also synergistically contribute to its overall protective effect on LN. For example, AS-IV may indirectly reduce the oxidative stress stimulation on podocytes by inhibiting the release of inflammatory factors in renal tissues, forming an “upstream” inhibition of ferroptosis. This multi-target characteristic may be the basis for its curative effect. This study provides a solid pre-clinical experimental basis for AS-IV as an adjuvant or alternative therapeutic drug for LN. Compared with currently commonly used immunosuppressants, AS-IV is derived from natural plants, with relatively high long-term safety and potentially fewer side effects. It exerts its effect through a relatively specific mechanism of inhibiting podocyte ferroptosis, which may help to more precisely protect the glomerular filtration barrier and reduce proteinuria.
Although this study has achieved a series of important findings, there are still some limitations that need to be addressed and deepened in future research. In in-vivo studies, the MRL/lpr mouse model was used. Although this model can spontaneously develop LN, its genetic background is single, and there are certain differences in the disease progression compared with human LN. In in-vitro studies, an immortalized MPC-5 was used, and its biological behavior may be different from that of primary podocytes or human podocytes. Future research should consider verification in primary podocytes, conditional gene-knockout mice, and cells derived from LN patients.
Conclusion
In summary, this study confirmed through in vivo and in vitro experiments that AS-IV can inhibit podocyte ferroptosis by regulating the PI3K/AKT/Nrf2 axis, alleviate the degree of kidney damage, and thus relieve LN (Figure 8). This discovery not only provides new insights into the pathogenesis of LN but also lays an important theoretical foundation for the development of novel LN treatment strategies based on natural products and targeting podocyte protection. However, this study has certain limitations, and further functional verification experiments are needed to clarify its mechanism of action. Future research should focus on overcoming the limitations of existing models, exploring the detailed mechanisms of AS-IV, and promoting its translation into clinical applications.
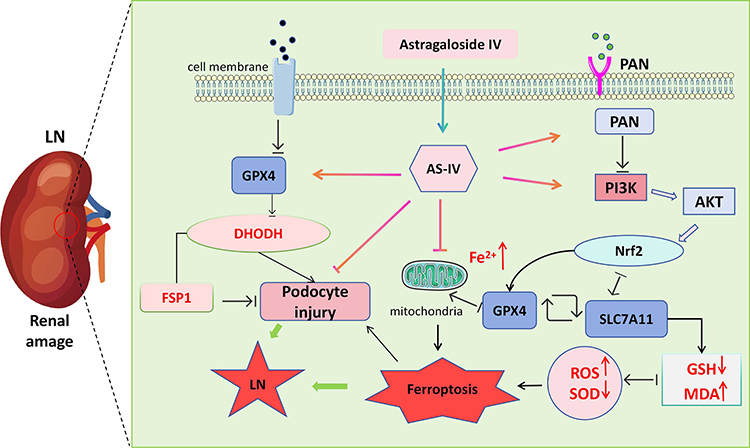 |
Figure 8 Schematic diagram of the mechanism by which Astragaloside IV inhibits ferroptosis of renal podocytes and improves lupus nephritis through the PI3K/AKT/Nrf2 axis. |
Data Sharing Statement
The data can be obtained by contacting the corresponding author.
Ethical Approval
All animal experiments were approved by the Animal Ethics Committee of Anhui University of Traditional Chinese Medicine (No. AHUCM-mouse-2024200). The human data in this study were sourced exclusively from public databases with open licenses permitting unrestricted reuse. Per China’s National Science and Technology Ethics Committee guidelines, such data usage is exempt from ethical review (available at https://www.gov.cn/zhengce/zhengceku/2023-02/28/content_5743658.htm). Ethical exemption was confirmed by The First Affiliated Hospital of Anhui University of Chinese Medicine.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the National Natural Science Foundation of China (No.82574970), Clinical Medical Research Transformation Special Project of Anhui Province (No.202304295107020114 and No.202304295107020115).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Xia XY, Liu ST, Song ZC, et al. Sexual status of female systemic lupus erythematosus patients: a qualitative study. Lupus. 2026:9612033261429133. doi:10.1177/09612033261429133
2. Luo M, He H, Chen L, Yuan G, Yang L, He F. Systemic lupus erythematosus combined with Castleman disease: a case report. Wien Klin Wochenschr. 2025. doi:10.1007/s00508-025-02682-9
3. Hocaoǧlu M, Valenzuela‐Almada MO, Dabit JY, et al. Incidence, prevalence, and mortality of lupus nephritis: a population-based study over four decades using the lupus midwest network. Arthritis Rheumatol. 2023;75(4):567–19. doi:10.1002/art.42375
4. De Vriese AS, Sethi S, Fervenza FC. Lupus nephritis: redefining the treatment goals. Kidney Int. 2025;107(2):198–211. doi:10.1016/j.kint.2024.10.018
5. Dobrowolski C, Lao SM, Kharouf F, et al. Lupus nephritis II: treatment and monitoring. Adv Clin Chem. 2025;126:121–154. doi:10.1016/bs.acc.2025.01.001
6. Niu Y, Jin Y, Hao Y, et al. Paeonol interferes with lupus nephritis by regulating M1/M2 polarization of macrophages. Mol Immunol. 2024;169:66–77. doi:10.1016/j.molimm.2024.03.004
7. Wang L, Tian H, Wang H, et al. Disrupting circadian control of autophagy induces podocyte injury and proteinuria. Kidney Int. 2024;105(5):1020–1034. doi:10.1016/j.kint.2024.01.035
8. Lv C, Cheng T, Zhang B, Sun K, Lu K. Triptolide protects against podocyte injury in diabetic nephropathy by activating the Nrf2/HO-1 pathway and inhibiting the NLRP3 inflammasome pathway. Ren Fail. 2023;45(1):2165103. doi:10.1080/0886022x.2023.2165103
9. Zhao X-Y, Li -S-S, He Y-X, et al. SGLT2 inhibitors alleviated podocyte damage in lupus nephritis by decreasing inflammation and enhancing autophagy. Ann Rheum Dis. 2023;82(10):1328–1340. doi:10.1136/ard-2023-224242
10. Li J, Jia Y-C, Ding Y-X, Bai J, Cao F, Li F. The crosstalk between ferroptosis and mitochondrial dynamic regulatory networks. Int J Biol Sci. 2023;19(9):2756–2771. doi:10.7150/ijbs.83348
11. Ye B, Chen B, Guo C, et al. C5a-C5aR1 axis controls mitochondrial fission to promote podocyte injury in lupus nephritis. Mol Ther. 2024;32(5):1540–1560. doi:10.1016/j.ymthe.2024.03.003
12. Liu C, Gan Y-H, Yong W-J, et al. OTUB1 regulation of ferroptosis and the protective role of ferrostatin-1 in lupus nephritis. Cell Death Dis. 2024;15(11):791. doi:10.1038/s41419-024-07185-5
13. Du L, Guo C, Zeng S, Yu K, Liu M, Li Y. Sirt6 overexpression relieves ferroptosis and delays the progression of diabetic nephropathy via Nrf2/GPX4 pathway. Ren Fail. 2024;46(2):2377785. doi:10.1080/0886022x.2024.2377785
14. Gao N, Huang Z, Xie J, et al. Cryptotanshinone alleviates cerebral ischemia reperfusion injury by regulating ferroptosis through the PI3K/AKT/Nrf2 and SLC7A11/GPX4 signaling pathway. J Ethnopharmacol. 2025;348:119800. doi:10.1016/j.jep.2025.119800
15. Hu N, Li Y, Zhang G, et al. PF4 regulates neuronal ferroptosis in cerebral hemorrhage through CXCR3/PI3K/AKT/Nrf2 pathway. Biomole Biomed. 2024;25(6):1322–1334. doi:10.17305/bb.2024.11415
16. Liu L, Yang S, Wang H. α-Lipoic acid alleviates ferroptosis in the MPP + -induced PC12 cells via activating the PI3K/Akt/Nrf2 pathway. Cell Biol Int. 2021;45(2):422–431. doi:10.1002/cbin.11505
17. Liang Y, Chen B, Liang D, et al. Pharmacological effects of Astragaloside IV: a review. Molecules. 2023;28(16):6118. doi:10.3390/molecules28166118
18. Tang Z, Cheng L, Li M, Huang C. Astragaloside IV alleviates systemic lupus erythematosus by modulating ITGB1/PTK2/p38 pathway: integrated network pharmacology and experimental validation. Drug Des Devel Ther. 2025;19:8967–8984. doi:10.2147/dddt.S554510
19. Qin L-Y, Guan P, Wang J-X, et al. Therapeutic potential of Astragaloside IV against adriamycin-induced renal damage in rats via ferroptosis. Front Pharmacol. 2022;13:812594. doi:10.3389/fphar.2022.812594
20. Li L, Zou J, Zhou M, et al. Phenylsulfate-induced oxidative stress and mitochondrial dysfunction in podocytes are ameliorated by Astragaloside IV activation of the SIRT1/PGC1α /Nrf1 signaling pathway. Biomed Pharmacother. 2024;177:117008. doi:10.1016/j.biopha.2024.117008
21. Liu C, Liu X, Wang Y, et al. Mesenchymal stromal cells reduce ferroptosis of podocytes by activating the Nrf2/HO-1/GPX4 pathway in lupus nephritis. Int Immunopharmacol. 2025;153:114537. doi:10.1016/j.intimp.2025.114537
22. Dunn C, Brettle D, Cockroft M, Keating E, Revie C, Treanor D. Quantitative assessment of H&E staining for pathology: development and clinical evaluation of a novel system. Diagn Pathol. 2024;19(1):42. doi:10.1186/s13000-024-01461-w
23. Giardino L, Generali L, Del Fabbro M, Tartaglia GM, Bidossi A, Savadori P. Detection of bacteria in dental samples using the Periodic acid-Schiff (PAS) histological stain. Micron. 2023;172:103498. doi:10.1016/j.micron.2023.103498
24. Stefan G, Stancu S, Zugravu A, Petre N, Mandache E, Mircescu G. Towards a simplified renal histopathological prognostic score in glomerular nephropathies. Histopathology. 2020;77(6):926–935. doi:10.1111/his.14175
25. Liu Y, Wang X, Huang M, et al. METTL3 facilitates kidney injury through promoting IRF4-mediated plasma cell infiltration via an m6A-dependent manner in systemic lupus erythematosus. BMC Med. 2024;22(1):511. doi:10.1186/s12916-024-03735-y
26. Nezamuldeen L, Mal W, Al Zahrani RA, Jambi S, Jafri MS. Renal-AI: a deep learning platform for multi-scale detection of renal ultrastructural features in electron microscopy images. Diagnostics. 2026;16(2). doi:10.3390/diagnostics16020264
27. Daina A, Michielin O, Zoete V. SwissTargetPrediction: updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019;47(W1):W357–w364. doi:10.1093/nar/gkz382
28. Wang X, Shen Y, Wang S, et al. PharmMapper 2017 update: a web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017;45(W1):W356–w360. doi:10.1093/nar/gkx374
29. Piñero J, Ramírez-Anguita JM, Saüch-Pitarch J, et al. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020;48(D1):D845–d855. doi:10.1093/nar/gkz1021
30. Barshir R, Fishilevich S, Iny-Stein T, et al. GeneCaRNA: a comprehensive gene-centric database of human non-coding RNAs in the GeneCards suite. J Mol Biol. 2021;433(11):166913. doi:10.1016/j.jmb.2021.166913
31. Amberger JS, Bocchini CA, Scott AF, Hamosh A. OMIM.org: leveraging knowledge across phenotype–gene relationships. Nucleic Acids Res. 2019;47(D1):D1038–d1043. doi:10.1093/nar/gky1151
32. Zhou Y, Zhou B, Pache L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019;10(1):1523. doi:10.1038/s41467-019-09234-6
33. Zardecki C, Dutta S, Goodsell DS, Lowe R, Voigt M, Burley SK. PDB −101: educational resources supporting molecular explorations through biology and medicine. Protein Sci. 2022;31(1):129–140. doi:10.1002/pro.4200
34. Cheng C, Li B, Li J, et al. Multi-stain deep learning prediction model of treatment response in lupus nephritis based on renal histopathology. Kidney Int. 2025;107(4):714–727. doi:10.1016/j.kint.2024.12.007
35. Chen K, Deng Y, Shang S, et al. Complement factor B inhibitor LNP023 improves lupus nephritis in MRL/lpr mice. Biomed Pharmacother. 2022;153:113433. doi:10.1016/j.biopha.2022.113433
36. Zhu H, Zheng L, Zhao K, et al. Astragaloside IV improves diabetic kidney disease by regulating NLRP3 inflammasome. J Diabetes Res. 2025;2025(1):3340719. doi:10.1155/jdr/3340719
37. Wang J, Wang L, Feng X, et al. Astragaloside IV attenuates fatty acid-induced renal tubular injury in diabetic kidney disease by inhibiting fatty acid transport protein-2. Phytomedicine. 2024;134:155991. doi:10.1016/j.phymed.2024.155991
38. Zhang M, Liu W, Liu Y, et al. Astragaloside IV inhibited podocyte pyroptosis in diabetic kidney disease by regulating SIRT6/HIF-1α axis. DNA Cell Biol. 2023;42(10):594–607. doi:10.1089/dna.2023.0102
39. Ye Z, Xie B, Tao Y, Xiao D. Mechanism of ferroptosis and its role in disease development. Int J Biol Sci. 2025;21(12):5328–5360. doi:10.7150/ijbs.102859
40. Zhong S, Wang N, Zhang C. Podocyte death in diabetic kidney disease: potential molecular mechanisms and therapeutic targets. Int J Mol Sci. 2024;25(16):9035. doi:10.3390/ijms25169035
41. Gong Q, Lai T, Liang L, Jiang Y, Liu F. Targeted inhibition of CX3CL1 limits podocytes ferroptosis to ameliorate cisplatin-induced acute kidney injury. Mol Med. 2023;29(1):140. doi:10.1186/s10020-023-00733-3
42. Hu H, Liang W, Ding G. Podocyte metabolic reprogramming and targeted therapy. J Am Soc Nephrol. 2026;37(3):619–633. doi:10.1681/asn.0000000917
43. Liu Y, Li S, Rong W, et al. Podocyte-released migrasomes in urine serve as an indicator for early podocyte injury. Kidney Diseases. 2020;6(6):422–433. doi:10.1159/000511504
44. Ma S, Qin J, Zhang Y, et al. Disrupting mitochondrial dynamics attenuates ferroptosis and chemotoxicity via upregulating NRF2-mediated FSP1 expression. Cell Rep. 2025;44(9):116234. doi:10.1016/j.celrep.2025.116234
45. Wen X, Tang S, Wan F, Zhong R, Chen L, Zhang H. The PI3K/Akt-Nrf2 signaling pathway and mitophagy synergistically mediate hydroxytyrosol to alleviate intestinal oxidative damage. Int J Biol Sci. 2024;20(11):4258–4276. doi:10.7150/ijbs.97263
46. Xie Y, Kang R, Klionsky DJ, Tang D. GPX4 in cell death, autophagy, and disease. Autophagy. 2023;19(10):2621–2638. doi:10.1080/15548627.2023.2218764
47. Zhang H, Pan J, Huang S, et al. Hydrogen sulfide protects cardiomyocytes from doxorubicin-induced ferroptosis through the SLC7A11/GSH/GPx4 pathway by Keap1 S-sulfhydration and Nrf2 activation. Redox Biol. 2024;70:103066. doi:10.1016/j.redox.2024.103066
48. Zhang M, Zheng H, Zhu X, et al. Synchronously evoking disulfidptosis and ferroptosis via systematical glucose deprivation targeting SLC7A11/GSH/GPX4 antioxidant axis. ACS Nano. 2025;19(14):14233–14248. doi:10.1021/acsnano.5c00730
49. Yao M, Zhang L, Wang L. Astragaloside IV: a promising natural neuroprotective agent for neurological disorders. Biomed Pharmacother. 2023;159:114229. doi:10.1016/j.biopha.2023.114229
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Comprehensive Analysis of Ferroptosis Markers in Lupus Nephritis Based on Bioinformatics Analysis and Experimental Validation
Zhang S, Hu W, Huang C, Lin X, Chen X
Journal of Inflammation Research 2025, 18:10855-10871
Published Date: 12 August 2025
Machine Learning-Based Identification and Experimental Validation of Hub Ferroptosis-Related Cuproptosis Genes in Lupus Nephritis
Zhang S, Hu W, Zhang Y, Huang C, He Z, Xu J, Lin S, Yang B, Chen X
Journal of Inflammation Research 2025, 18:11335-11353
Published Date: 18 August 2025
CYBB-Mediated Ferroptosis Drives Podocyte Injury in Lupus Nephritis and Represents a Therapeutic Target
Wang B, Jiang H, Wu D, Liu M, Chen H, Zhao S, Yan Y, Li S, Fang X, Wu R
Journal of Inflammation Research 2026, 19:575320
Published Date: 22 January 2026