Back to Journals » Nature and Science of Sleep » Volume 18

Associations Between Social Jetlag and Gut Microbiome Profiles in Adolescents
Authors Gong Q, Ding K, Lou W, Wang J ![]() , Bian G
, Bian G
Received 24 February 2026
Accepted for publication 29 April 2026
Published 27 May 2026 Volume 2026:18 603131
DOI https://doi.org/10.2147/NSS.S603131
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Ahmed BaHammam
Qinghai Gong,1,* Keqin Ding,1,* Wangwei Lou,2 Jinwen Wang,3 Guolin Bian1
1Ningbo Municipal Center for Disease Control and Prevention, Ningbo, Zhejiang, People’s Republic of China; 2Ninghai Country Center for Disease Control and Prevention, Ninghai, Zhejiang, People’s Republic of China; 3Department of Endocrinology and Metabolism, The Affiliated People’s Hospital of Ningbo University, Ningbo, Zhejiang, People’s Republic of China
*Qinghai Gong and Keqin Ding contributed equality to this work
Correspondence: Guolin Bian, Email [email protected]
Background: The microbiota-gut-brain axis plays a vital role in health, with growing evidence linking SJL to the gut microbiome. In this study, the relationship between self-reported SJL and gut microbiome composition is evaluated in an adolescent population.
Methods: A total of 120 healthy adolescents (69 females, 51 males; mean age 12.69± 0.65 years) were included. We investigated the associations of self-reported SJL with gut microbial α-diversity, the Firmicutes/Bacteroidetes (F/B) ratio, specific bacterial taxa, and functional gene diversity.
Results: Participants had an average SJL of 0.87± 0.73 h, with 25.0% experiencing SJL ≥ 2 h/d. Although continuous SJL was negatively correlated with α-diversity at the family level (Shannon and Simpson indices, all P< 0.05), no significant differences were found between the SJL < 2 and ≥ 2 h/d groups. Several taxa across multiple taxonomic levels (e.g. Bacteroidia and Bacteroidales) were associated with SJL (P< 0.05). Differential abundance was observed for multiple families and genera between these two SJL groups. After FDR correction (q≤ 0.05), only the glutamate dehydrogenase gene gdh_K00262 was significantly enriched in the high SJL group (q=0.05).
Conclusion: SJL is associated with shifts in functionally relevant gut taxa, rather than a global ecological restructuring, in adolescents. Future work is needed to decipher the mechanisms of this microbiome-SJL interaction and its health significance.
Keywords: gut microbiome, sleep jetlag, adolescents
Introduction
SJL (SJL)-the persistent discordance between biologically driven circadian timing and externally dictated sleep-wake patterns-has escalated into a global adolescent health epidemic.1,2 Recent epidemiological data from China reveal that 61.2% of school-age adolescents endure SJL ≥1 hour, while a substantial subset (17.8%) grapple with more severe circadian disruption exceeding 2 hours.1 Excessive SJL has multifaceted adverse effects on the health of children and adolescents, including obesity,3 cardiovascular morbidity,4 dysregulated dietary patterns,5 and psychological and behavioral problems.6,7 It has also been linked to impairments in cognitive function,8 memory,9 attention,10 and academic performance.11 Thus, SJL represents a pervasive and modifiable health risk factor for adolescent well-being.
Growing evidence indicates that circadian rhythms are closely linked to the composition and function of the gut microbiota.12–14 Even minor disruptions to circadian patterns can influence microbial community structure.15 For instance, a study conducted within the ZOE PREDICT 1 cohort revealed that a 90-minute difference in sleep midpoint between workdays and free days was associated with significant variations in the abundance of 17 gut bacterial species.16 It has been proposed that SJL may induce a state of systemic circadian disorder, potentially mediated or amplified by dysregulation of the gut-brain axis.17–19 Therefore, the gut microbiome may act both as a target of SJL-induced disruption and as a key mediator in translating circadian misalignment into broader physiological and neuropsychiatric consequences. Adolescence is a period of active maturation of both the circadian system and the gut microbiome, The circadian timing system undergoes significant changes during pubertal development, contributing to characteristic shifts in sleep-wake patterns.20,21 The gut microbiome continues to develop toward an adult-like profile throughout adolescence, with its composition and function remaining dynamic until stability is achieved in early adulthood.22,23 Although adult studies have investigated the SJL-gut microbiome nexus, this area remains largely unexplored in adolescents- a demographic marked by heightened vulnerability to circadian disruption. Whether conclusions from adult research extend to this developmental window, where both circadian systems and gut microbial communities undergo maturation, remains uncertain. Addressing this research gap, the primary objective of this study was to investigate the relationship between SJL and gut microbiome composition in an adolescent cohort. Secondary objectives were to examine associations with alpha-diversity, relative abundances of key bacterial taxa, and functional gene pathways. Emerging evidence suggests that circadian disruption can alter gut microbial nitrogen metabolism,24 which in turn affects host metabolic health and intestinal barrier function. We hypothesized that elevated SJL (SJL) would correspond to a perturbed microbial profile, reflected in diminished alpha-diversity and altered abundances of specific taxa and functional pathways using the NCycDB database.
Methods
Study Design and Participants
In December 2024, we recruited a non-random sample of early adolescents (12–14 years) from a single public primary school located in the central urban area of Cixi, Zhejiang Province, China. All participants were in apparently good health. Participants were excluded if they (i) declined assent or whose guardians withheld consent, (ii) carried physician-diagnosed chronic conditions (eg., metabolic, endocrine, genetic, cardiac, or neurodevelopmental disorders, severe malnutrition, or anatomical deformities), or (iii) had taken any medications known to affect gut microbiota composition, such as antibiotics, proton pump inhibitors, laxatives, and probiotic supplements. The final analysis study included 120 healthy student participants. Detailed information regarding the purpose and procedures of the population-based study was provided to all children and their parents or legal guardians, from whom written informed consent was obtained. The research protocol received ethical approval from the Ethics Committee of the Ningbo Municipal Center for Disease Control and Prevention (Approval No. 202313). Each participant completed a comprehensive questionnaire covering demographic characteristics, general health status, sleep behaviors, and prior medical history.
SLJ Assessment
As reported in our previous study, SJL was estimated from self-reported sleep–wake schedules.25 Participants answered two questions for both weekdays and weekends: (1)”What time do you usually go to bed? (2)”What time do you usually wake up?” The SJL value, quantified in hours, was computed as the difference between the sleep midpoints on weekdays and weekends.26 Mid-sleep time was calculated as bedtime plus half of the total sleep duration, for weekdays and weekends, respectively.27 Consistent with previous research, an SJL ≥ 2 h was used to classify participants into high-SJL and low-SJL groups.2
Gut Microbiome Sample Collection and Sequencing
Participants collected morning stool samples at home using EasySampler kits (ALPCO) and transferred them into tubes pre-filled with DNA/RNA Shield (Zymo Research). Immediately after collection, samples were stored in liquid nitrogen and transported within 24 hours to the research center in liquid nitrogen containers. Upon arrival, samples were immediately transferred and stored at −80°C until DNA extraction. A total of n=120 stool samples were collected and processed for metagenomic sequencing. DNA extraction was performed following the standard protocols of BMKCloud (Biomarker Technologies Corporation). DNA concentration was quantified using a Qubit fluorometer (Thermo Fisher Scientific), and purity was assessed by OD260/280 and OD260/230 ratios using a NanoDrop spectrophotometer (Thermo Fisher Scientific). Samples were required to meet the following criteria: DNA concentration ≥10 ng/μL, total DNA ≥0.1 μg, OD260/280 within 1.8–2.0, and OD260/230 ≥1.8. Microbial DNA was isolated from 200 mg of faecal material with the PowerSoil® DNA Isolation kit (MO BIO) following the manufacturer’s high-throughput protocol. DNA concentration and purity were assessed with a Qubit 2.0 fluorometer (Thermo Fisher). Genomic DNA was randomly fragmented, and libraries were prepared with the TrueLib DNA Library Rapid Prep Kit for Illumina (ExCell Bio). Paired-end sequencing (2×150 bp, 500 bp inserts) was performed on an Illumina NovaSeq 6000 at Biomarker Technologies (Beijing). Taxonomic classification, functional annotation (Nitrogen Cycle Database), and assembly procedures followed previously published pipelines.28,29 The detailed workflow was as follows: (i) Quality filtering: Raw reads were filtered using fastp (v0.23.2) with parameters as implemented in the BMKCloud platform (date accessed: 15/04/2025). Low-quality or ambiguous reads were removed. (ii) Host read removal: Filtered reads were aligned to the human reference genome (GRCh38) using Bowtie2 (v2.4.4); mapped reads were discarded. (iii) Metagenomic assembly: Clean reads were assembled using MEGAHIT (v1.2.9) with default parameters. Contigs <300 bp were filtered out. Assembly quality was assessed using QUAST (v5.0.2). (iv) Gene prediction and non‑redundant gene set construction: Coding regions were predicted using MetaGeneMark (v3.26) with default parameters. Redundancy was removed using MMseqs2 (v12-113e3) with 90% sequence identity and 80% coverage thresholds. (v) Functional annotation: Predicted genes were annotated using the NR, GO, KEGG, eggNOG,Pfam and SwissProt databases as implemented in the BMKCloud platform (date accessed: 15/04/2025). To specifically investigate nitrogen metabolism, a pathway of interest given the hypothesized link between circadian disruption and microbial nitrogen cycling, we performed targeted functional annotation using the NCycDB database via the BMKCloud platform. NCycDB is a manually curated database of nitrogen cycle-related genes, comprising 84,759 (95% identity) and 219,146 (100% identity) representative sequences across 68 gene subfamilies covering eight nitrogen cycle processes.24,30 All raw metagenomic data were deposited in the National Center for Biotechnology Information Sequence Read Archive (https://www.ncbi.nlm.nih.gov/) under BioProject accession number PRJNA1403232.
Statistical Analysis
Continuous variables are presented as mean and standard deviation, median and interquartile range, and categorical variables as frequencies (%). Mann–Whitney U-tests (two-tailed) were employed to compare bacterial relative abundances, alpha-diversity indices, and functional gene counts between high and low SJL groups. Partial Spearman correlations evaluated associations between continuous SJL values and alpha-diversity metrics (Chao1, Shannon, Simpson, Invsimpson). For partial correlation analyses of individual bacterial taxa, taxa were included if they met the following criteria: ubiquity ≥80% (present in at least 96 of 120 participants) and mean relative abundance ≥2% at their respective taxonomic levels. To evaluate the adequacy of the sample size for detecting differences in alpha-diversity between the high and low SJL groups, we conducted a post-hoc power analysis using the observed effect size for the Shannon diversity index (Cohen’s d = 0.28). The analysis was performed using the pwr package in R (version 4.2.0), with α set at 0.05 (two-tailed). The achieved power was 0.32, indicating insufficient power to detect a small-to-moderate effect. Differential taxa at the genus level were identified with LEfSe (|LDA score| > 2) and the Kruskal–Wallis test (P < 0.05). Alpha-, beta-, and functional-diversity analyses were conducted on the BMK Cloud platform.31 We also performed differential abundance analysis for functional genes with the non-parametric Mann–Whitney U-test, and applied false discovery rate (FDR) correction to adjust for multiple testing. All of the other analyses were carried out using the R software (version 4.1.0), and a two-sided P ≦ 0.05 was considered statistically significant.
Results
Participants Characteristics
The general characteristics of the study participants are shown in Table 1. The mean age of the 120 adolescents (69 females/51 males) was 12.69±0.65 years. The average SJL was 0.87±0.73 h, and 30 participants (25.0%) reported SJL≥ 2hours/day. In addition, the average sleep duration was 8.32±0.83 h.
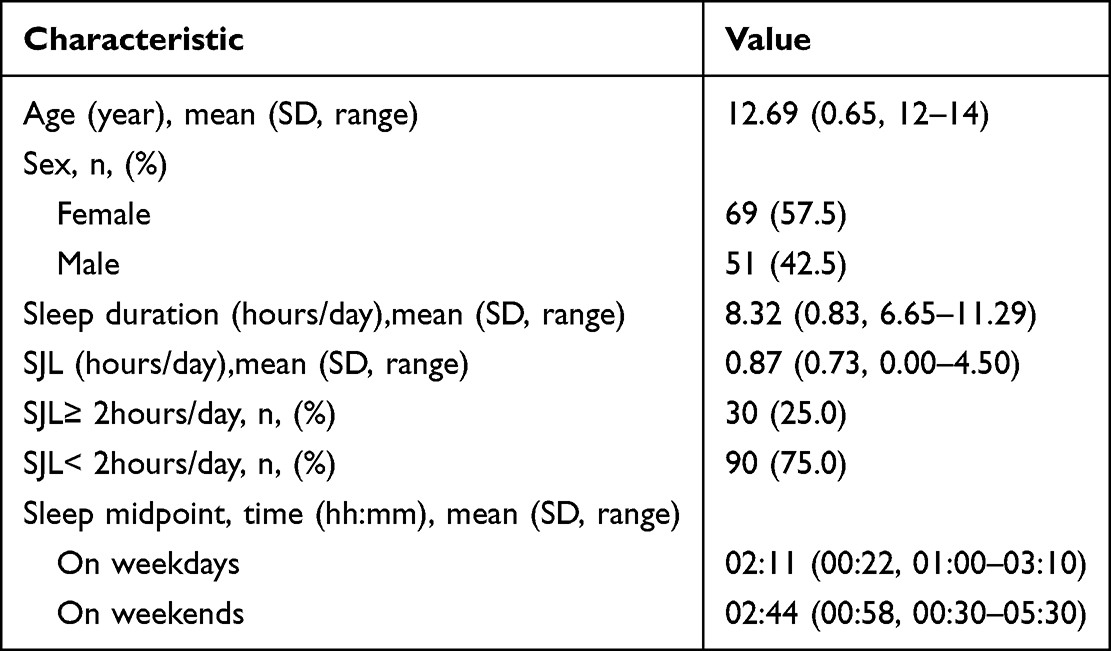 |
Table 1 Participant Characteristics and Sleep Measures (n=120) |
Relationships Between SJL with Gut Microbiome Variables
Descriptive statistics for bacterial taxa that met the inclusion criteria at each taxonomic level are listed in Table 2. The analysis revealed significant negative associations between α-diversity (assessed using the Shannon and Simpson) and SJL at the bacterial family level (Shannon: Mid (P25-P75)=2.08 (1.72–2.29), r=−0.183, Simpson: Mid (P25-P75)=0.80 (0.71–0.85), r=−0.198, all P < 0.05). Descriptive statistics for bacterial taxa that met the inclusion criteria at each taxonomic level are listed in Table 3. Two of the taxa, Bacteroidia (class) and Bacteroidales (order) were associated with SJL (Bacteroidia: Mid (P25-P75)=40.8 1(24.24–58.02), r= 0.209; Bacteroidales: Mid (P25-P75)=41.11 (24.29–58.14), r= 0.210, all P< 0.05). Beta diversity was assessed using weighted UniFrac distance, and the principal coordinate analysis (PCoA) plot showed no significant separation between the high and low SJL groups (PERMANOVA, R2=0.01, P=0.2, Figure 1). No differences in α-diversity of the fecal microbiota were observed between subjects who were classified in the SJL <2 h/d and SJL ≥2 h/d groups (all P>0.05, Figure 1).
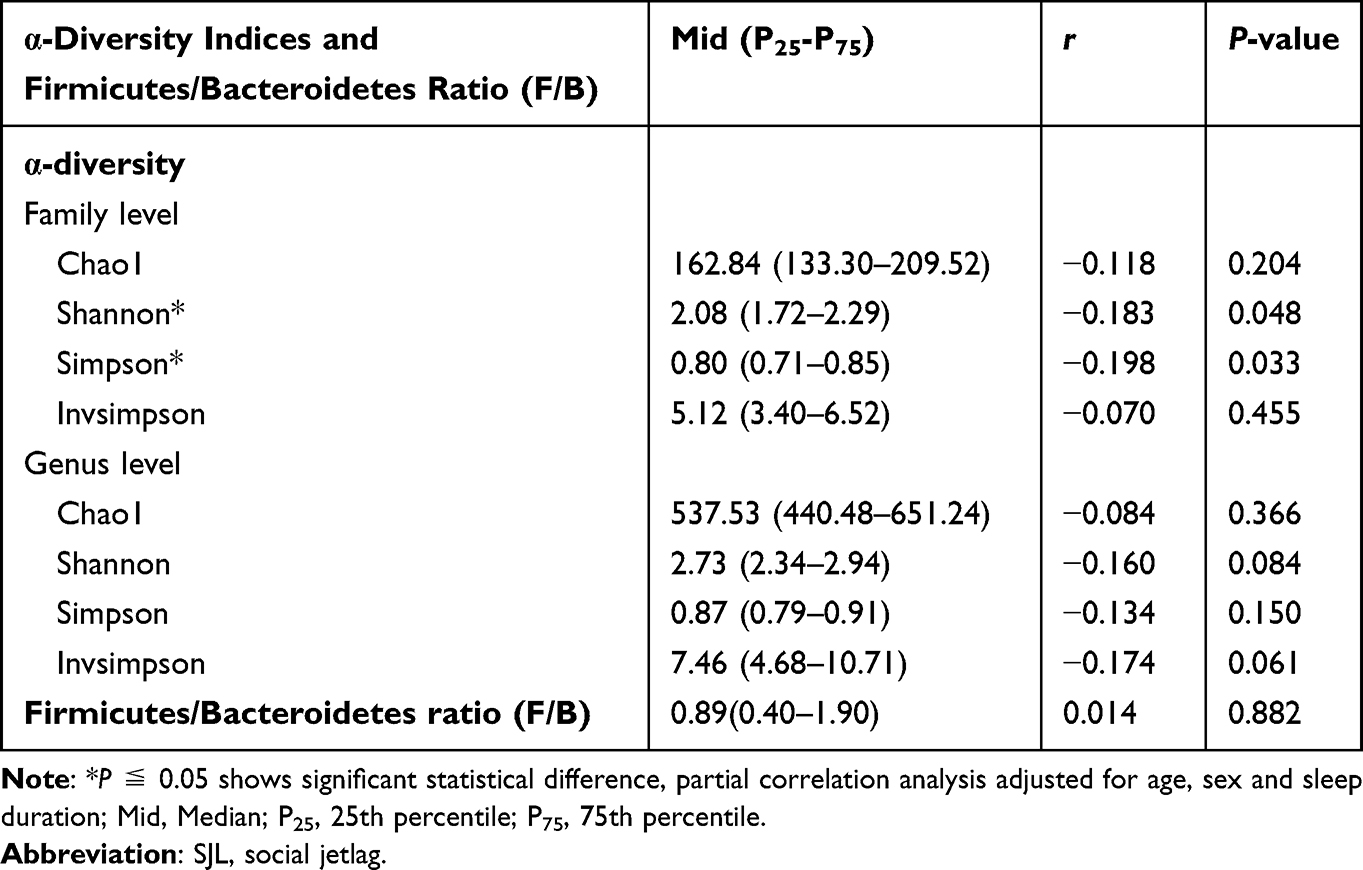 |
Table 2 Partial Correlations of α-Diversity Indices and Firmicutes/Bacteroidetes Ratio (F/B) with the SJL |
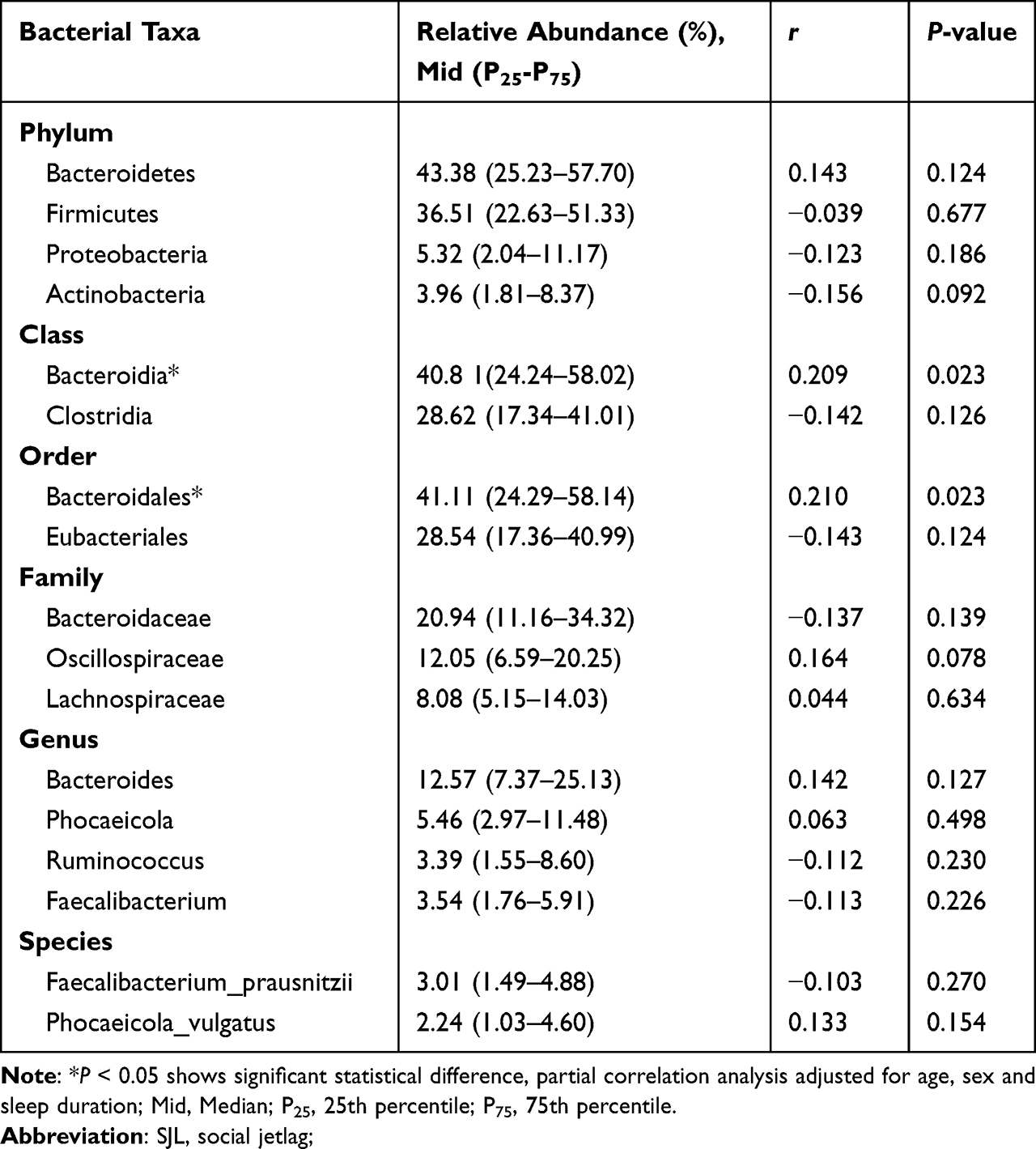 |
Table 3 Partial Correlations of Select Bacterial Taxa with the SJL |
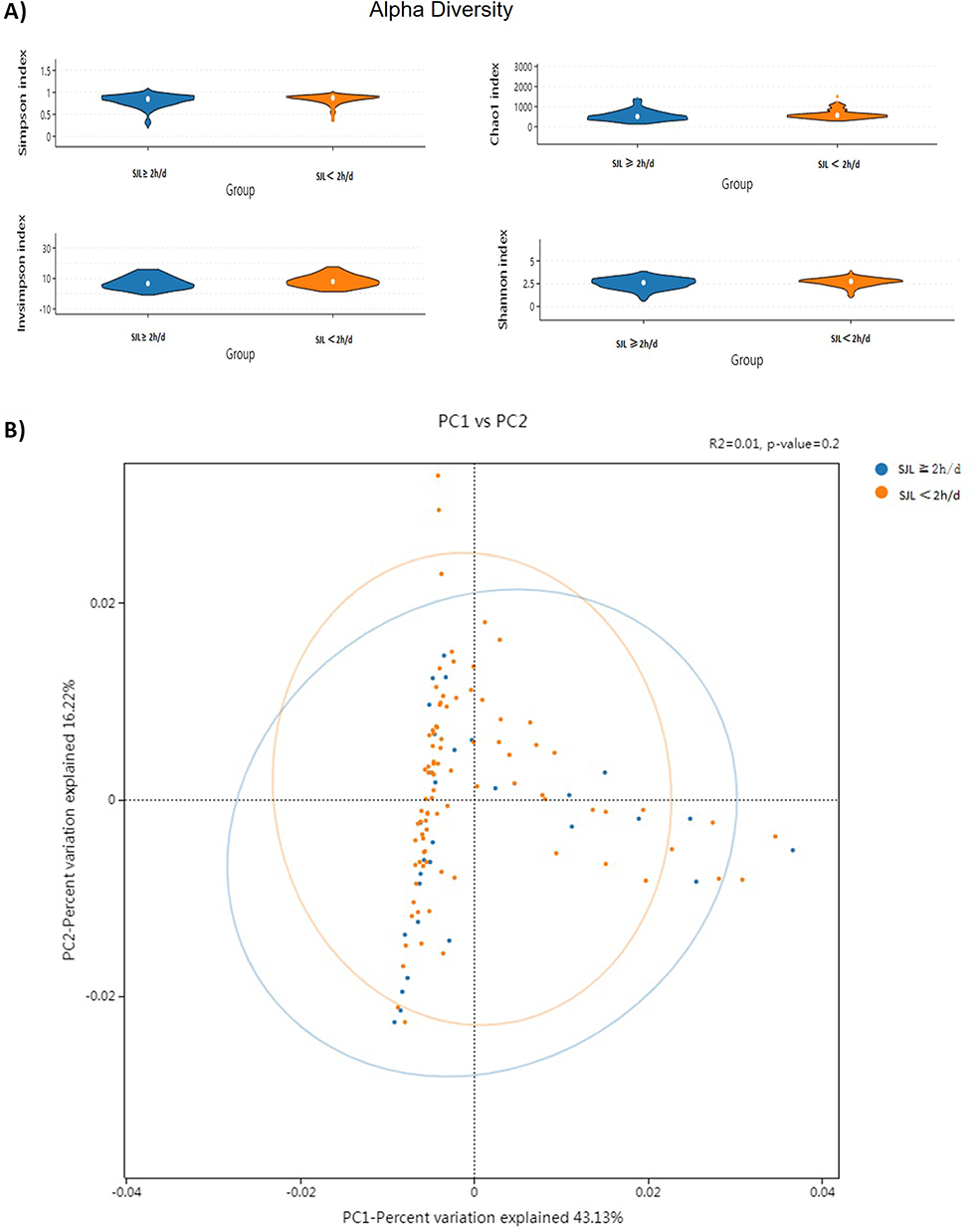 |
Figure 1 Gut microbiome diversity in adolescents with low (< 2 h/d) and high (≥ 2 h/d) social jetlag (SJL). (A) Alpha diversity indices at the genus level. Violin plots compare the two groups (Mann–Whitney U-test). (B) Beta diversity. Principal component analysis (PCA) ordination based on weighted UniFrac distance visualizes community composition differences. Statistical testing was performed using PERMANOVA (weighted UniFrac distance) (PERMANOVA, R2= 0.01, P = 0.2). Points represent samples colored by SJL group (high vs. low). |
Comparison of Bacterial Composition Between the Groups
At the family taxonomic level, Ruminococcaceae showed a tendency toward greater relative abundance (LDA SCORE >3, P <0.05) in subjects with SJL ≥2 h/d than in those with SJL <2 h/d. Conversely, the abundances of Atopobiaceae, Coprobacillaceae, Peptostreptococcaceae, Campylobacteraceae, Treponemataceae, and Clostridiaceae were all observed to be higher in the SJL <2 h/d group (Figure 2A and B). At the genus level, multiple taxa such as Ligilactobacillus, Acidovorax, Eisenbergiella, Olsenella, Coprobacillus, Campylobacter, Finegoldia, and Treponema exhibited an upward trend in relative abundance (|LDA SCORE| >2, P <0.05) within the SJL <2 h/d group (Figure 3A and B).
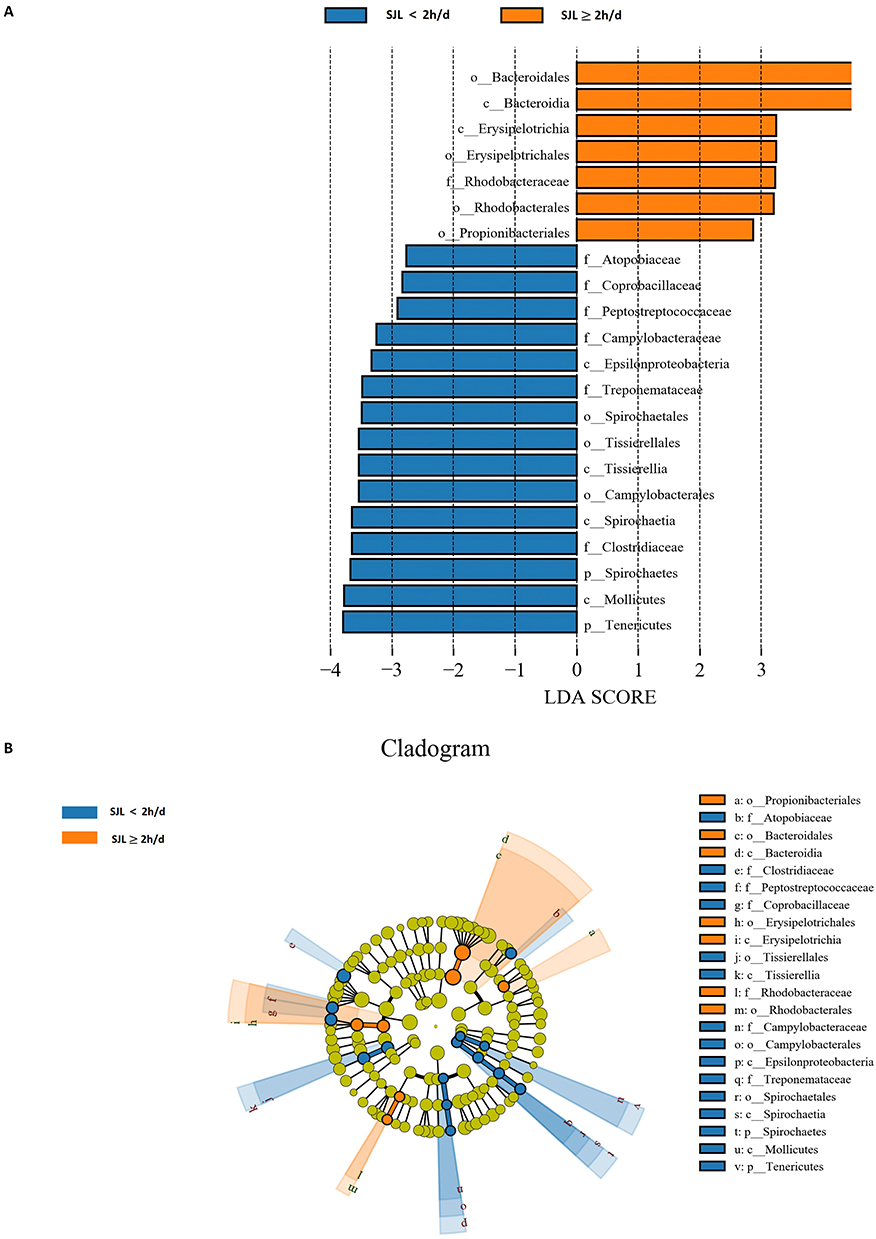 |
Figure 2 Specific bacterial taxa at family level between SJL (SJL)≧2h/d (Orange) and SJL < 2h/d (blue) groups. (A) Linear discriminant analysis effect size (LEfSe) analysis revealed significant differences in the fecal microbiota between the two groups. Taxa with an linear discriminant analysis (|LDA|) score > 2 and a Kruskal–Wallis test p < 0.05 are listed. (B) The cladogram, constructed using the Linear discriminant analysis effect size (LEfSe) method, indicates the phylogenetic distribution of fecal microbiota associated with the two SJL groups. Abbreviations: f, family level; c, class level; o, order level; p, phylum level. |
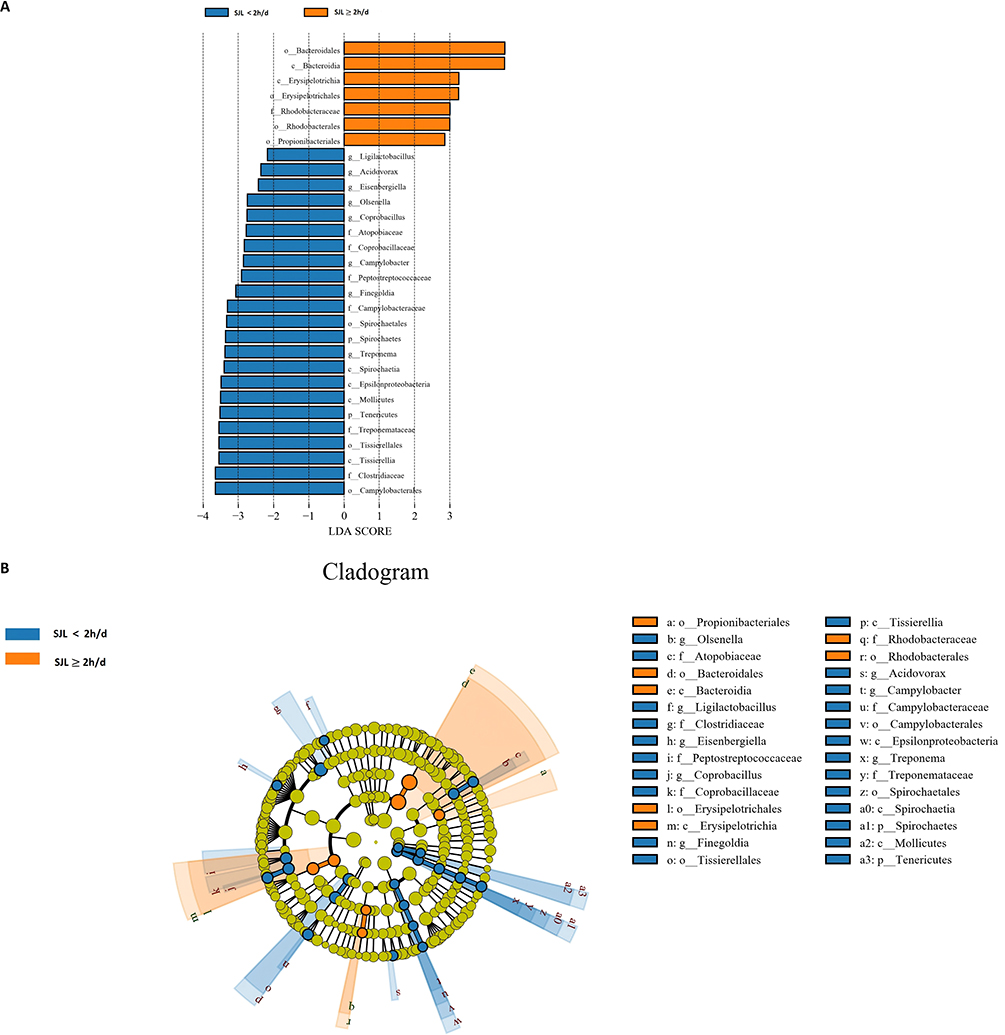 |
Figure 3 Differential bacterial taxa at the genus level between groups with social jetlag (SJL) ≥ 2 h/d and < 2 h/d. (A) Linear discriminant analysis effect size (LEfSe) analysis identified taxa with significant differences in fecal microbiota between the two groups (SJL ≥ 2 h/d: Orange; SJL < 2 h/d: blue). Taxa meeting the criteria of linear discriminant analysis (|LDA|) score > 2 and a Kruskal–Wallis test p < 0.05 are listed are listed. (B) Cladogram generated by LEfSe illustrates the phylogenetic distribution of fecal microbiota associated with each group. Abbreviations: p, phylum; c, class; o, order; f, family; g, genus. |
Differential Abundance Analysis of Functional Genes
To further investigate the microbial nitrogen metabolism pathways influenced by SJL, functional genes were aligned with the specialized Nitrogen Cycle Database (Ncyc orthology). The abundance of 57 microbial nitrogen metabolism key genes was compared between the normal and high SJL groups using the non-parametric Mann–Whitney U-test, with multiple comparisons adjusted by false discovery rate (FDR) correction. After this stringent adjustment (q ≤ 0.05), only the gdh_K00262 gene reached the threshold for significance (q = 0.05). However, this boundary finding should be interpreted with caution, as it shows a higher abundance in the high SJL group (q = 0.05). However, four other genes-ureA, ureC, gdh_k15371, and nosZ-which showed nominally significant differences in raw p - values (all P < 0.05), did not retain statistical significance after FDR correction (all q > 0.05). All gene-level summary statistics-including median abundance, interquartile range, uncorrected P-values and FDR-adjusted q-values are provided in Supplementary Table 1.
Discussion
This study revealed that SJL in children was significantly associated with certain gut microbiota. Although it did not affect the alpha or beta diversity of the gut microbial community between the groups with SJL ≥ 2 h/d and those below this threshold, notably, a distinct pattern of differential microbial abundance was observed between the SJL groups. To the best of our knowledge, this study therefore provides the first metagenomic evidence linking SJL to pediatric gut microbiota disruption.
Our findings suggest that the association between SJL and gut microbiota is weak and not holistic. A monophyletic assemblage comprising the class Bacteroidia and the order Bacteroidales showed concomitant elevation alongside increasing SJL. This pattern points to a potential shift in the gut microbiota toward a greater abundance of Bacteroides in individuals with elevated SJL. Although Bacteroides members are renowned for metabolic versatility and resilient gut colonisation,32,33 several strains can ignite mucosal inflammation under host-specific contexts.34 Therefore, an increase in these taxa in individuals with SJL ≥ 2 h/d might indicate a microbiota configuration that is primed to promote or exacerbate inflammatory processes, potentially linking disrupted sleep patterns to adverse health outcomes. This phenomenon may be mediated through two primary pathways. Firstly, SJL often correlates with erratic eating patterns and poor diet quality, which are key determinants of microbial community structure.16 Secondly, direct circadian disruption impairs essential gut functions, including motility, secretion, and local immune surveillance, all of which have a profound influence on the microbial environment.35 Consequently, this SJL-associated dysbiosis could be a significant contributor to the metabolic and inflammatory comorbidities observed in individuals with irregular sleep - wake cycles.
We observed a disproportionate expansion of the Ruminococcaceae lineage within the high-SJL microbiome. This echoes a recent meta-analysis that likewise catalogued a rise in the same family under chronic sleep curtailment conditions.36 Although these organisms are among the principal colonic producers of short-chain fatty acids (notably butyrate), which support epithelial turnover and mucosal integrity,37,38 heir role in health is context-dependent. Specific species such as Ruminococcus gnavus have been linked to inflammatory conditions, and recent evidence suggests that their enrichment may reflect shifts in bile acid metabolism rather than a uniform health benefit.39 Whether the observed expansion of Ruminococcaceae in our circadian-misaligned cohort stabilizes barrier function or, conversely, contributes to low-grade inflammation will require isolate-level metagenomics and gnotobiotic modeling.40
At genus-level resolution, subjects with low SJL scores carried a taxonomically broader assemblage. Among the expanded genera, Ligilactobacillus-whose members are routinely used as live biotherapeutics—was markedly enriched, suggesting a community structure that favors intestinal stability.14 Inaddition, the abundance of Olsenella and Coprobacillus genera also increased in individuals with low SJL, but their specific functions in gut homeostasis remain unclear and require further investigation. However, the increased abundance of Campylobacteraceae (genus Campylobacter), Treponemataceae (genus Treponema), and Peptostreptococcaceae (potentially including Finegoldia) in the low SJL group presents a paradoxical finding. For individuals with low SJL, which reflects stable metabolic and circadian rhythms, these microbial taxa often display behavior characterized by conditional pathogenicity or functional duality.41 The observed pattern indicates that individuals with regular sleep -wake cycles may have a different, and not universally beneficial, ecological network of gut microbiota. It is possible that in the context of overall metabolic and circadian stability, these taxa exist as commensals without provoking disease, a state potentially disrupted by the physiological stress of high SJL.
In our study, the divergent microbial profiles appear to stem from the manifold physiological and behavioral consequences of SJL. First, SJL frequently accompanies dietary shifts-lower overall diet quality, erratic eating schedules, and diminished consumption of microbiota-supportive items such as nuts. Nutrients act as immediate growth substrates for bacteria, while the temporal structure of meals entrains microbial oscillations in peripheral tissues.16,42 Second, disruption of circadian clocks intrinsically perturbs fundamental host processes governing the intestinal milieu, motility, barrier integrity, and mucosal immunity.43–45 Such alterations modify local nutrient flux, oxygen levels, and antimicrobial-peptide release, thereby opening niches that selectively promote the bloom of particular taxa.46,47 In summary, our findings corroborate earlier evidence that circadian misalignment constitutes a key behavioral driver of microbiota structure.16
Our present study specifically identified an up-regulation of the microbial glutamate dehydrogenase gene gdh_K00262, which is a phylogenetically widespread and biochemically characterized genetic marker for a critical step in microbial nitrogen metabolism,48 in individuals exhibiting high SJL. This observation suggests a targeted functional change in gut microbiota nitrogen metabolism. Given the established evidence that SJL alters gut microbial composition and that microbiota functions exhibit circadian rhythms, this shift in gdh_K00262 may represent a metabolic adaptation to host circadian disruption. Although the functional consequences of this alteration remain to be determined, we hypothesize that it could modulate microbial glutamate synthesis and, potentially, influence the availability of neuroactive precursors such as GABA via the microbiota-gut-brain axis.49,50 This speculative mechanistic link between SJL and neurobehavioral outcomes warrants investigation in future studies integrating metabolomic, neuroimaging, and behavioral assessments. It should be noted that the significant association for gdh_K00262 was observed at the conventional FDR threshold of 0.05. Given the exploratory nature of this analysis and the potential for type I error, this finding requires replication in larger, independent cohorts.
Furthermore, the microbial signature associated with lower SJL (<2 h/d) is notable It is characterized by an increased abundance of taxa like the probiotic genus Ligilactobacillus51 and the health-associated families Atopobiaceae52 and Coprobacillaceae,53 suggesting a more beneficial gut microbiota profile with stable sleep-wake cycles. In contrast, the high SJL group showed a trend only toward enriching Ruminococcaceae,54 a key SCFA producer. Although SCFAs are advantageous, this selective increase may reflect an adaptive response to circadian disruption. Consequently, the physiological meaning of this specific microbial shift requires confirmation through future research.
This study is subject to a number of limitations that must be recognised. Firstly, the relatively small sample size within each SJL subgroup, notably the underrepresentation of the lower SJL cohort. A post-hoc power analysis revealed that the study was underpowered (power = 0.32) to detect the observed effect size for alpha-diversity. Therefore, the null findings may reflect insufficient statistical power rather than a true absence of association. Future studies with larger, more balanced samples are needed to more definitively evaluate the relationship between SJL and gut microbiota diversity. Secondly, the cross-sectional design precludes causal inference. It remains unclear whether SJL drives the microbial changes or if a pre-existing gut microbiota composition predisposes individuals to greater circadian misalignment. Future longitudinal and interventional studies are needed to establish causality. Thirdly, the reliance on child self - report without parental verification may have further contributed to measurement error. Additionally, we did not use objective sleep assessment tools such as wrist-worn actigraphy, which would have provided more precise estimates of sleep timing and enabled the validation of the self-reported measures. Future studies that incorporate both subjective and objective sleep assessments, along with parental verification when appropriate, are warranted to more accurately capture SJL in adolescent populations. Fourth, detailed dietary intake data were not collected. We cannot rule out the possibility that unmeasured dietary differences may have confounded the observed SJL-microbiome associations. Future studies should incorporate detailed dietary assessments to better isolate the independent effects of circadian disruption. Lastly, by sampling exclusively in winter, we may have introduced uncontrolled confounding related to seasonal factors, including reduced daylight duration, altered dietary patterns, and decreased outdoor physical activity. These season-specific conditions may limit the applicability of our findings to other seasons or geographic regions with different climate patterns. Future multi-site studies employing year-round recruitment across diverse geographic and climatic settings are warranted to validate and extend our observations.
Conclusion
In summary, this study reveals an association between SJL and specific bacterial taxa in the gut microbiota of children. Our findings suggest that SJL may not induce wholesale restructuring of the overall microbial community, but rather exerts targeted modulation of select functional taxa. These observations indicate that even in the absence of broad shifts in community structure, SJL can still perturb gut microecology by influencing key bacterial populations. However, several limitations should be considered when interpreting these findings. The relatively small sample size, particularly in the high SJL group, resulted in limited statistical power, which may have contributed to null findings for some comparisons. The reliance on self‑reported sleep timing introduces potential recall and reporting bias, and the absence of comprehensive dietary data limits our ability to control for important confounders. Additionally, the single site, single season, and cross-sectional design precludes inferences about causality or temporal dynamics. Future studies with larger, more balanced samples, objective sleep and dietary assessments, and longitudinal designs are needed to confirm and extend these findings, and to clarify the mechanistic pathways linking SJL to child health outcomes.
Data Sharing Statement
The sequencing datasets from this study have been deposited in the NCBI Sequence Read Archive (SRA). The accession number for this data is PRJNA1403232.
Ethics Approval and Consent to Participate
The protocol for the research project had been approved by the Ningbo CDC Institutional Review Board (No. 202313) and had therefore been performed following the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments. Both the participant and his/her parents or guardian have signed written informed consent forms, and all information related to participants and schools is confidential.
Acknowledgments
Thanks to the Center for Disease Control and Prevention of Cixi County for their support of this study. Thanks to all the participating children and their parents/guardians from Cixi, Ningbo, Zhejiang, China.
Author Contributions
Guolin Bian: Conceptualization, Funding acquisition, Project administration
Qinghai Gong: Data curation, Funding acquisition, Methodology, Writing – original draft, Writing – review & editing
Keqin Ding: Data curation, Investigation, Writing – review & editing
Jinwen Wang: Formal analysis
Wangwei Lou: Validation
All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by Zhejiang Provincial Natural Science Foundation (No. LTGY24H260007), Ningbo Public Welfare Science and Technology Key Project (No.2025S036), Disease Control and Prevention of Zhejiang Provincial (No. 2025JK070), Medical and Health Science Program of Zhejiang Province (No. 2025HY1038) and Ningbo Top Medical and Health Research Program (No.2023020713).
Disclosure
The authors have no conflicts of interest to disclose.
References
1. Li M, Zhang Y, Huang M, et al. Prevalence, correlates, and mental health outcomes of social jetlag in Chinese school-age adolescents: a large-scale population-based study. Sleep Med. 2024;119:424–12. doi:10.1016/j.sleep.2024.05.039
2. Sun S, Yang Y, Yu F, et al. Social jetlag and depressive symptoms among young people: a systematic review and meta-analysis. BMC Psychiatry. 2025;25(1):664. doi:10.1186/s12888-025-07066-x
3. Higgins S, Stoner L, Black K, et al. Social jetlag is associated with obesity-related outcomes in 9-11-year-old children, independent of other sleep characteristics. Sleep Med. 2021;84:294–302. doi:10.1016/j.sleep.2021.06.014
4. Cui L, Li J, Lou X, et al. Associations between sleep characteristics and risk for high blood pressure among students aged 9-18: a cross-sectional study in China. Sleep Med. 2023;107:72–80. doi:10.1016/j.sleep.2023.04.004
5. Arab A, Karimi E, Garaulet M, Scheer F. Social jetlag and dietary intake: a systematic review. Sleep Med. Rev. 2023;71:101820. doi:10.1016/j.smrv.2023.101820
6. Henderson SEM, Brady EM, Robertson N. Associations between social jetlag and mental health in young people: a systematic review. Chronobiol. Int. 2019;36(10):1316–1333. doi:10.1080/07420528.2019.1636813
7. Chen CX, TMH Li, Zhang J, et al. The impact of sleep-corrected social jetlag on mental health, behavioral problems, and daytime sleepiness in adolescents. Sleep Med. 2022;100:494–500. doi:10.1016/j.sleep.2022.09.027
8. Yang FN, Picchioni D, Duyn JH. Effects of sleep-corrected social jetlag on measures of mental health, cognitive ability, and brain functional connectivity in early adolescence. Sleep. 2023;46(12):zsad259. doi:10.1093/sleep/zsad259
9. Deibel SH, Lewis LM, Cleary J, Cassell TTS, Skinner DM, Thorpe CM. Unpredictable mealtimes rather than social jetlag affects acquisition and retention of hippocampal dependent memory. Behav Processes. 2022;201:104704. doi:10.1016/j.beproc.2022.104704
10. Diogo FMC, Bessa ZCM, Galina SD, et al. Sex differences in temporal sleep patterns, social jetlag, and attention in high school adolescents. Sleep Sci. 2024;17(2):e125–e133. doi:10.1055/s-0043-1777831
11. Li AR, Thomas ML, Gonzalez MR, et al. Greater social jetlag predicts poorer NIH Toolbox crystallized cognitive and academic performance in the Adolescent Brain Cognitive Development (ABCD) study. Chronobiol. Int. 2024;41(6):829–839. doi:10.1080/07420528.2024.2353848
12. Matenchuk BA, Mandhane PJ, Kozyrskyj AL. Sleep, circadian rhythm, and gut microbiota. Sleep Med. Rev. 2020;53:101340. doi:10.1016/j.smrv.2020.101340
13. Lotti S, Dinu M, Colombini B, Amedei A, Sofi F. Circadian rhythms, gut microbiota, and diet: possible implications for health. Nutr Metab Cardiovasc Dis. 2023;33(8):1490–1500. doi:10.1016/j.numecd.2023.05.009
14. Dos Santos A, Galiè S. The microbiota-gut-brain axis in metabolic syndrome and sleep disorders: a systematic review. Nutrients. 2024;16(3):390. doi:10.3390/nu16030390
15. Yang Y, Hernandez MC, Chitre S, Jobin C. Emerging roles of modern lifestyle factors in microbiome stability and functionality. Curr. Clin. Microbiol. Rep. 2025;12(1):1–11. doi:10.1007/s40588-025-00242-3
16. Bermingham KM, Stensrud S, Asnicar F, et al. Exploring the relationship between social jetlag with gut microbial composition, diet and cardiometabolic health, in the ZOE PREDICT 1 cohort. Eur J Nutr. 2023;62(8):3135–3147. doi:10.1007/s00394-023-03204-x
17. Tofani GSS, Leigh SJ, Gheorghe CE, et al. Gut microbiota regulates stress responsivity via the circadian system. Cell Metab. 2025;37(1):138–153. doi:10.1016/j.cmet.2024.10.003
18. Yang R, Wang X, Yang J, et al. Perturbations in the microbiota-gut-brain axis shaped by social status loss. Commun. Biol. 2025;8(1):401. doi:10.1038/s42003-025-07850-1
19. Savvidis C, Maggio V, Rizzo M, Zabuliene L, Ilias I. The gut microbiota axis in social jetlag: a novel framework for metabolic dysfunction and chronotherapeutic innovation. Medicina. 2025;61(9):1630. doi:10.3390/medicina61091630
20. Olejniczak I, Pilorz V, Oster H. Circle(s) of life: the circadian clock from birth to death. Biology. 2023;12(3):383. doi:10.3390/biology12030383
21. Carson MD, Westwater C, Novince CM. Adolescence and the microbiome: implications for healthy growth and maturation. Am J Pathol. 2023;193(12):1900–1909. doi:10.1016/j.ajpath.2023.07.004
22. Mancabelli L, Milani C, De Biase R, et al. Taxonomic and metabolic development of the human gut microbiome across life stages: a worldwide metagenomic investigation. mSystems. 2024;9(4):e0129423. doi:10.1128/msystems.01294-23
23. Laue HE, Willis AD, Wang F, et al. Early-life and concurrent predictors of the healthy adolescent microbiome in a cohort study. Genome med. 2025;17(1):50. doi:10.1186/s13073-025-01481-1
24. Parkar SG, Kalsbeek A, Cheeseman JF. Potential role for the gut microbiota in modulating host circadian rhythms and metabolic health. Microorganisms. 2019;7(2):41. doi:10.3390/microorganisms7020041
25. Yang Y, Li SX, Zhang Y, et al. Chronotype is associated with eating behaviors, physical activity and overweight in school-aged children. Nutr J. 2023;22(1):50. doi:10.1186/s12937-023-00875-4
26. Wittmann M, Dinich J, Merrow M, Roenneberg T. Social jetlag: misalignment of biological and social time. Chronobiol. Int. 2006;23(1–2):497–509. doi:10.1080/07420520500545979
27. Roenneberg T, Allebrandt KV, Merrow M, Vetter C. Social jetlag and obesity. Curr bio. 2012;22(10):939–943. doi:10.1016/j.cub.2012.03.038
28. Liu L, Wu P, Chen F, et al. Multi-omics analyses reveal that the gut microbiome and its metabolites promote milk fat synthesis in Zhongdian yak cows. PeerJ. 2022;10:e14444. doi:10.7717/peerj.14444
29. Chen B, Sun L, Zeng G, et al. Gut bacteria alleviate smoking-related NASH by degrading gut nicotine. Nature. 2022;610(7932):562–568. doi:10.1038/s41586-022-05299-4
30. Tu Q, Lin L, Cheng L, Deng Y, He Z. NCycDB: a curated integrative database for fast and accurate metagenomic profiling of nitrogen cycling genes. Bioinformatics. 2019;35(6):1040–1048. doi:10.1093/bioinformatics/bty741
31. Guo X, Su L, Shi M, et al. Network Pharmacology and Transcriptomics to Explore the Pharmacological Mechanisms of 20(S)-Protopanaxatriol in the Treatment of Depression. Int J Mol Sci. 2024;25(14).
32. McCallum GA-O, Tropini CA-O. The gut microbiota and its biogeography. Nat Rev Microbiol. 2024;22(2):105–118. doi:10.1038/s41579-023-00969-0
33. Liu H, Shiver AL, Price MN, et al. Functional genetics of human gut commensal Bacteroides thetaiotaomicron reveals metabolic requirements for growth across environments. Cell Rep. 2021;34(9):108789. doi:10.1016/j.celrep.2021.108789
34. Spindler MP, Siu S, Mogno I, et al. Human gut microbiota stimulate defined innate immune responses that vary from phylum to strain. Cell Host Microbe. 2022;30(10):1481–1498. doi:10.1016/j.chom.2022.08.009
35. Puri S, Shaheen M, Grover B. Nutrition and cognitive health: a life course approach. Front Public Health. 2023;11:1023907. doi:10.3389/fpubh.2023.1023907
36. Supasitdikul T, Mazariegos JRR, Nhat NN, et al. Sleep deprivation alters gut microbiome diversity and taxonomy: a systematic review and meta-analysis of human and rodent studies. J. Sleep Res. 2025;35:e70125. doi:10.1111/jsr.70125
37. Li Y, Shao L, Mou Y, Zhang Y, Ping YA-O. Sleep, circadian rhythm and gut microbiota: alterations in Alzheimer’s disease and their potential links in the pathogenesis. Gut Microbes. 2021;13(1):1957407. doi:10.1080/19490976.2021.1957407
38. Hays KE, Pfaffinger JM, Ryznar R. The interplay between gut microbiota, short-chain fatty acids, and implications for host health and disease. Gut Microbes. 2024;16(1):2393270. doi:10.1080/19490976.2024.2393270
39. Andreu-Sánchez S, Blanco-Míguez A, Wang D, et al. Global genetic diversity of human gut microbiome species is related to geographic location and host health. Cell. 2025;188(15):3942–3959. doi:10.1016/j.cell.2025.04.014
40. Vico-Oton E, Volet C, Jacquemin N, et al. Strain-dependent induction of primary bile acid 7-dehydroxylation by cholic acid. BMC Microbiol. 2024;24(1):286. doi:10.1186/s12866-024-03433-y
41. Burnham PM, Hendrixson DR. Campylobacter jejuni: collective components promoting a successful enteric lifestyle. Nat Rev Microbiol. 2018;16(9):551–565. doi:10.1038/s41579-018-0037-9
42. Moreira Gobis ML, de Souza-Silva TG, de Almeida Paula HA, de Almeida Paula HA. The impact of a western diet on gut microbiota and circadian rhythm: a comprehensive systematic review of in vivo preclinical evidence. Life Sci. 2024;349:122741. doi:10.1016/j.lfs.2024.122741
43. Parasram KA-O, Zuccato A, Shin M, et al. The emergence of circadian timekeeping in the intestine. Nat Commun. 2024;15(1):1788. doi:10.1038/s41467-024-45942-4
44. Post ZA-OX, Zilberstein NF, Keshavarzian A. The circadian rhythm as therapeutic target in inflammatory bowel disease. J Can Assoc Gastroenterol. 2024;8(Suppl 2):S27–S35. doi:10.1093/jcag/gwae027
45. Duboc H, Coffin B, Siproudhis L. Disruption of circadian rhythms and gut motility: an overview of underlying mechanisms and associated pathologies. J Clin Gastroenterol. 2020;54(5):405–414. doi:10.1097/MCG.0000000000001333
46. Srivastav S, Biswas A, Anand A. Interplay of niche and respiratory network in shaping bacterial colonization. J Biol Chem. 2024;301(1):108052.
47. Cullen TW, Schofield WB, Barry NA, et al. Gut microbiota. Antimicrobial peptide resistance mediates resilience of prominent gut commensals during inflammation. Science. 2015;347(6218):170–175. doi:10.1126/science.1260580
48. Frey B, Varliero G, Qi W, Stierli B, Walthert L, Brunner I. Shotgun metagenomics of deep forest soil layers show evidence of altered microbial genetic potential for biogeochemical cycling. Front Microbiol. 2022;13:828977. doi:10.3389/fmicb.2022.828977
49. Toledo MA-O, Martínez-Martínez SA-O, Van Hul M, et al. Rapid modulation of gut microbiota composition by hypothalamic circuits in mice. Nat Metab. 2025;7(6):1123–1135.
50. Cesaro A, Broset E, Salimando GJ, et al. Using synthetic biology to understand the gut-brain axis. Cell Rep Phy Sci. 2025;6(3):102458.
51. Patra S, Pradhan B, Roychowdhury A. Complete genome sequence, metabolic profiling and functional studies reveal Ligilactobacillus salivarius LS-ARS2 is a promising biofilm-forming probiotic with significant antioxidant, antibacterial, and antibiofilm potential. Front Microbiol. 2025;16:1535388.
52. Shao J, Su T, Wang J, et al. Mediators linking gut microbiota and sporadic Creutzfeldt-Jakob disease: a Mendelian randomization study. AMB Express. 2025;15(1):120. doi:10.1186/s13568-025-01932-3
53. Huang R, Zhang J, Sun M, et al. Oat β-glucan enhances gut barrier function and maintains intestinal homeostasis in naturally aging mice. Int J Biol Macromol. 2025;305(1):141129. doi:10.1016/j.ijbiomac.2025.141129
54. Dassa B, Borovok I, Ruimy-Israeli V, et al. Rumen cellulosomics: divergent fiber-degrading strategies revealed by comparative genome-wide analysis of six ruminococcal strains. PLoS One. 2014;9(7):e99221. doi:10.1371/journal.pone.0099221
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.