")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 15
Association study of the excitatory amino acid transporter 2 (EAAT2) and glycine transporter 1 (GlyT1) gene polymorphism with schizophrenia in a Polish population
Authors Merk W , Kucia K , Mędrala T , Kowalczyk M, Owczarek A, Kowalski J
Received 16 November 2018
Accepted for publication 14 February 2019
Published 24 April 2019 Volume 2019:15 Pages 989—1000
DOI https://doi.org/10.2147/NDT.S194924
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Roger Pinder
Wojciech Merk,1 Krzysztof Kucia,1 Tomasz Mędrala,1 Małgorzata Kowalczyk,2 Aleksander Owczarek,3,4 Jan Kowalski2
1Department of Psychiatry and Psychotherapy, School of Medicine in Katowice, Medical University of Silesia, Katowice, Poland; 2Department of Medical Genetics, School of Pharmacy with the Division of Laboratory Medicine in Sosnowiec, Medical University of Silesia, Katowice, Poland; 3Division of Statistics, Department of Instrumental Analysis, School of Pharmacy with the Division of Laboratory Medicine in Sosnowiec, Medical University of Silesia, Katowice, Poland; 4Department of Instrumental Analysis, School of Pharmacy with the Division of Laboratory Medicine in Sosnowiec, Medical University of Silesia, Katowice, Poland
Background: Excitatory amino acid transporter 2 encoded by SLC1A2 is responsible for approximately 90% of glutamate uptake. Glycine transporter 1, encoded by SLC6A9, is responsible for maintaining a low concentration of the N-methyl-D-aspartate receptor (NMDAR) co-agonist – glycine in the synaptic cleft, suggesting its participation in the development of the NMDARs hypofunction described in schizophrenia.
Aim: The aim of this study was to evaluate whether the functional polymorphism-181 A/C (rs4354668) of the SLC1A2 and the rs2486001 (IVS3+411 G/A) in the SLC6A9 are involved in schizophrenia development and its clinical picture in the Polish population.
Methods: The study group consisted of 393 unrelated Caucasian patients (157 [39.9%] females and 236 [60.1%] males; mean age 41±12) diagnosed with schizophrenia according to the DSM-5, and 462 healthy controls. The results of the Positive and Negative Syndrome Scale (PANSS) were presented in the five-dimensional model. Polymorphisms of SLC1A2 and SLC6A9 were genotyped with the use of PCR-RFLP assay.
Results: There were no statistically significant differences in the frequency of genotypes and alleles between the patients and controls for SLC1A2 and SLC6A9 polymorphisms in either the entire sample or after stratification according to gender. In the haplotype analysis, men with CA haplotype had more than 1.5 higher risk to develop schizophrenia than women (OR=1.63 [95% CI=1.17–2.27, p<0.05]). The influence of gender, genotypes of both analyzed polymorphisms and gender x genotype interactions on individual dimensions of the PANSS scale has not been observed. Also, there was no association of either polymorphism with suicide attempts.
Conclusion: The results of the present study did not indicate an association of polymorphism-181 A/C (rs4354668) in SLC1A2 and rs2486001 in SLC6A9 with onset of schizophrenia and its psychopathology in a Polish population.
Keywords: schizophrenia, glutamate system, excitatory amino acid transporter 2, glycine transporter 1, polymorphism, PANSS
Background
Schizophrenia, in contrast to Einheitspsychose, is regarded today as a heterogeneous clinical syndrome or collection of symptoms, including a number of changes in the cognitive, behavioral and emotional domains with poorly defined etiology. Various hypotheses have been proposed over the years to explain its pathogenesis. The etiology of schizophrenia is considered as an interaction of many factors, including genetic, and the heritability is estimated to be approximately 80%.1 The growing body of evidence drawn from post-mortem, neuroimaging and genetic studies stresses the role of the glutamate system and its interconnections with other neurotransmitter systems in the etiology of schizophrenia.
Glutamate is the major excitatory neurotransmitter and plays a key role in central nervous system (CNS) activation.2 More than 40% of CNS neurons use glutamate as a neurotransmitter, especially the pyramidal cells of the cortex.3 The possible role of glutamate in the etiopathogenesis of schizophrenia was based on clinical observations of phencyclidine (PCP) and ketamine action. Both substances induce not only schizophrenia-like positive and negative symptoms, but also lead to cognitive impairments in healthy volunteers4–6 and worsening of cognitive functions in patients with schizophrenia.7 Based on the psychomimetic properties of the N-methyl-D-aspartate receptor (NMDAR) antagonists, a decreased activity of the glutamate system was proposed to be present in schizophrenia. However, other studies have shown elevated glutamate levels in the striatum and prefrontal cortex (PFC) of those taking PCP, which increased its availability for the α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) and correlated with the severity of cognitive dysfunction.8 The AMPAR blocade reverses these effects, as well as the presynaptic metabotropic glutamate receptors 2 (mGluR2) and 3 (mGluR3) agonists.5,8 Therefore, activation of the glutamatergic system with AMPAR may be a link in the etiopathogenesis of schizophrenia.
Large-scale genome-wide association studies (GWASs) on schizophrenia have identified many low penetrance risk variants related to synaptic plasticity, neuron differentiation, neurotransmission, and immunity.9–11 The latest GWAS conducted by the Psychiatric Genomic Consortium (PGC) which combined several independent GWAS datasets in the largest resource available to date with 36 989 cases and 113 075 controls revealed the association between SCZ phenotype and a number of different loci containing genes involved in glutamatergic transmission.12 This group includes the following genes: GRM3 of the metabotropic glutamate receptor mGluR3, GRIN2A of the NMDAR subunit NR2A, GRIA1 of the glutamate receptor 1 (GluR1, GluA1) – a subunit of an AMPAR, SRR – serine racemase which catalyzes L-serine racemization to D-serine, an essential co-agonist and activator of NMDAR, CLCN3 of the voltage-gated chloride channel localized to glutamatergic synapses in the hippocampus and SLC38A7 encoding SNAT7, an L-glutamine preferring neuronal amino acid transporter that may be important for the reuptake and recycling of glutamate.12 In addition, GWAS that investigated cognitive phenotypes impaired in schizophrenia has been conducted. It was found that genetic variants related to cognitive trait impairment in schizophrenia are involved in the NMDA glutamate network.13 Associations between schizophrenia and genetic variants, such as rare SNPs, as well as copy number variants in the GRIN3A, which encodes the NR3A subunit of the NMDAR, have been also reported.14,15
Transport of glutamate from the synaptic cleft is a necessary mechanism for maintaining neuronal activity. Reuptake of glutamate is mediated by endothelial proteins - excitatory amino acid transporters (EAAT).16 Removal of glutamate prevents excessive binding to receptors and formation of free radicals.17 Interruption of this process may lead to excessive stimulation of glutamatergic receptors and excitotoxicity.18 The EAAT family contains 5 subtypes, numbered EAAT1–EAAT5 sequentially. EAAT 1 to 4 are located in the brain, whereas EAAT5 is found exclusively in the retina.19 EAAT 1 and 2 are mainly expressed in astrocytes, although the presence of EAAT2 was also found in neurons and oligodendrocytes.20,21 EAAT2, encoded by the SLC1A2, is responsible for approximately 90% of cerebral glutamate uptake.22 Blockade of glutamate uptake in oligodendrocyte cultures led to cell death, demyelination and astrocyte damage.23 Bauer et al24 documented lower glycosylation of EAAT1 and EAAT2 in the dorsolateral prefrontal cortex (DLPFC) and cingulate cortex of schizophrenia patients that could explain their reduced ability of glutamate reuptake from the synaptic cleft. Shan et al25 found decreased expression of EAAT1 and EAAT2 protein in the superior temporal gyrus and decreased EAAT2 protein in the hippocampus in schizophrenia. Rao et al26 reported higher protein and mRNA levels for EAAT1, EAAT3 and EAAT4 in DLPFC in schizophrenic patients, while EAAT2 expression levels remained unchanged. A decreased EAAT2 expression in parahippocampal and DLPFC of patients with schizophrenia was also observed.27 Based on these findings the SLC1A2 encoding EAAT2 became a candidate gene. The SLC1A2 consists of 11 exons coding for proteins and extends to over 50 kb of genomic DNA on chromosome 11p13-12.28 The functional significance of the polymorphism at position −181 in the promoter region of the SLC1A2, analyzed in this paper, was described by Mallolas et al29. The presence of this variant within the regulatory site for the activator protein 2 was found to prevent binding of this transcription factor, creating a new binding site for the repressor transcription factor GC-binding factor 2. As a consequence, the expression of the SLC1A2 has been altered, and the presence of C allele resulted in a 30% reduction in promoter activity compared with A allele.
Glycine plays an important role in the CNS development, and it is also a required co-agonist along with glutamate for NMDAR activation process. The availability of glycine, as well as its uptake from the synaptic cleft, is regulated by endothelial glycine transporters (GlyT). Two variants, known as Type 1 (GlyT1) and Type 2 (GlyT2), have been described. GlyT1 is widely expressed in glial cells of the PFC, hippocampus, hypothalamus, olfactory bulb and retina.30 It is composed of 12 endothelial domains responsible for glycine uptake, which ends the stimulation process.31 Heterozygous rodents with decreased expression of the GlyT1 gene exhibited increased NMDAR activity in the hippocampus and improved memory capacity and were also protected against post-methamphetamine sensory gating disorders, suggesting that GlyT1 inhibitors may improve cognitive performance and have antipsychotics properties. Postmortem studies of schizophrenia patients showed increased levels of glycine and serine in the medial temporal lobe.32 Administration of glycine to schizophrenia patients alleviated negative symptoms of the disease.33 Antipsychotics (such as haloperidol, chlorpromazine and clozapine) are uncompetitive GlyT1 antagonists in therapeutic doses, but their effect is assessed as poor.34 Monotherapy of patients with sarcosine, a GlyT1 inhibitor, has resulted in significant improvements in both negative symptoms and cognitive impairment.35 Due to the role of GlyT1 in regulation of NMDAR activity, the gene encoding GlyT1 seems to be a promising candidate gene in schizophrenia. The SLC6A9 for the GlyT1 transporter consists of 17 exons and is located on the short arm of chromosome 1 (1p31.3).36
Taking into account the results of previous reports pointed at the potential role of EAAT2 and GlyT1 in schizophrenia, the current study focuses on examining the association of the −181 A/C (rs4354668) polymorphism of the SLC1A2 encoding EAAT2 and the rs2486001 (IVS3+411 G/A) polymorphism in the SLC6A9 encoding GlyT1 with schizophrenia and its severity in Polish population.
Methods
The relationship between those two polymorphisms and schizophrenia has been assessed based on a comparison of genotype and allele distribution between the study and control group. We also examined the possible connection between some clinical variables (age of onset, severity and psychopathology of schizophrenia symptoms and history of suicide attempts) and the studied polymorphisms.
Subjects
The study group consisted of 393 unrelated Caucasian patients with a diagnosis of schizophrenia (157 [39.9%] females and 236 [60.1%] males; mean age 41±12, median 41, range 18–73) recruited from inpatients treated at the Department of Psychiatry and Psychotherapy, Medical University of Silesia in Katowice and the Neuropsychiatric Hospital in Lubliniec. All patients fulfilled the DSM-5 (Diagnostic and Statistical Manual of Mental Disorders, 5th Edition) diagnostic criteria for schizophrenia. Exclusion criteria involved: psychotic disorders other than schizophrenia, mood disorders, anxiety and stress-related disorders, substance-related and addictive disorders (excluding caffeine and nicotine), behavioral and personality disorders, neurological illness, endocrine disorders and autoimmune disease. The mean age of onset of schizophrenia, defined as the age at which the first psychotic symptoms appeared, was 26 years (SD=7, range 13–54). This information was acquired from patients` medical records. The mean duration of schizophrenia was 16 years (SD=10, range 1–47). The Positive and Negative Syndrome Scale (PANSS)37 was used by clinically trained psychiatrists to assess the severity of the illness. The results of the PANSS scale were presented in the five-dimensional model presented by van der Gaag et al38 which includes positive (POS), negative (NEG), disorganization (DIS), excitation (EXC) and emotional distress (EMO) dimension. The numerical values of these dimensions were calculated according to the formula given by the authors using all 30 PANSS symptoms.38 Suicide attempts made by patients in the past were classified as violent and nonviolent. Nonviolent methods included drug overdose, gas poisoning and melting, all others were classified as violent methods according to the division made in other studies.39 Suicide attempts have been reported in 75 patients (19.1%), including 40 (53.3%) classified as violent. Positive family history was diagnosed in 95 (24.2%) of schizophrenia patients. All of the patients were assessed to be capable of understanding the study before written consent was obtained.
The control group comprised 462 healthy unrelated individuals (238 women [51.5%]; mean age 40 years [SD=9, range 23–68 years]) recruited from among the volunteer blood donors at the Regional Centre of Blood Donation and Treatment in Katowice. Exclusion criteria for controls were: current psychiatric problems, any other neurological disorders, family history of schizophrenia (verified by direct interview), and chronic and acute physical illness (infections, autoimmune disease).
The study population was homogeneous and composed of the Polish Caucasian residents of the Silesia region. The study was approved by the Bioethics Committee of the Medical University of Silesia.
SNP choice and genotyping with PCR-RFLP
Information related to the SNPs was obtained from the National Center for Biotechnology Information, dbSNP (
Genomic DNA was isolated from whole peripheral blood using phenol-chloroform method. DNA extracts were qualitatively evaluated by electrophoresis in 1.5% agarose gel and quantitated spectrophotometrically using BioPhotometer plus (Eppendorf AG, Hamburg, Germany).
The genotypes for two SNPs: −181 A/C (rs4354668) in the SLC1A2 and rs2486001 (IVS3+411 G/A) in the SLC6A9 were determined by the polymerase chain reaction-restriction enzyme fragment length polymorphism (PCR-RFLP) assay under modified conditions according to the method described before.40,41 Briefly, the regions spanning polymorphisms were amplified using the following published primers: SLC1A2 5ʹ-GAGCGGCGGGGCCTCTTTTC-3ʹ and 5ʹ-TGCAGCCGCTGCCACCTGTG-3ʹ;41 SLC6A9 5ʹ-TTCTATTCCCTGGGGTTCAGCA-3ʹ and 5ʹ-AGCCTGGGCTGAGGCACACCAC-3ʹ.40 The PCR cycling conditions were an initial denaturation step at 95 ºC for 5 mins. followed by 35 cycles under the following conditions: 94°C for 30 s, 62°C for 30 s (SLC1A2) and 60°C for 30 s (SLC6A9) and final elongation at 72°C for 10 mins. Amplification was performed using a G-STORM GS1 thermal cycler (Gene Technologies Ltd, Essex, UK) with a Taq polymerase (Epicentre, Biotechnologies, Madison, WI, USA) according to the manufacturer`s instructions in a reaction mix with a total volume of 25 µL. Subsequently, amplified PCR products were digested with the appropriate restriction enzymes (Thermo Fisher Scientific, Waltham, MA, USA) and separated on 2% agarose gels stained with ethidium bromide. The SLC1A2 −181 A/C PCR products (381 bp) were digested with BcnI (NciI). An undigested product od 381 bp for the A allele and two products of 262 bp and 119 bp for the C allele were observed. The SLC6A9 IVS3+411 G/A PCR products (210 bp) were digested with Eco47I (AvaII). An uncut product of 210 bp for the G allele and two fragments of 123 bp and 87 bp for the A allele were visible.
The overall success rate of genotyping was over 95%. Samples with missing genotypes have been removed from the analysis. For quality control of genotyping, approximately 5% of the samples were randomly selected to be retested and the concordance of duplicated genotypes was 100%.
Statistical analysis
The results were presented as the mean ± SD in case of data with normal distribution and as median with IQR in case of data with skewed distribution. Qualitative data were expressed as percentage values. The Shapiro–Wilk test as well as quantile-quantile plot was used to evaluate data distribution. The association between the genotypes, gender, PANSS dimensions and age of onset was done with two-way ANOVA and contrast analysis. The homogeneity of variance was assessed by the Levene test. Comparison of nominal and order variables was performed using the χ2 test. Assessment of the relationship between occurrence of a particular allele, genotype, haplotype or gender and the occurrence of schizophrenia/suicide attempt was based on the OR with corresponding CIs and statistical significance. In case of genotypes analysis, the Hardy–Weinberg equilibrium (HWE) analysis was done. Then, the logistic regression model for five classic inheritance models (codominant, dominant, recessive and dominating and log-additive), linkage disequilibrium and haplotype analysis were performed. The impact of gender, age and time of onset on schizophrenia/suicide was assessed with the univariable logistic regression model. Adjustment of scoring distribution in individual dimensions of the PANSS scale was realized with the regression model with Epanecznikov kernel function. Correlation between time of onset and dimensions of the PANSS scale was done with the Pearson correlation coefficient. All p-values were two-tailed, and p<0.05 was established as statistically significant. The results were analyzed using the Statistica 10.0 version PL software (StatSoft, Cracow, Poland;
Results
Comparison of genotype and allele distributions between study and control group
The two-way ANOVA showed a statistically significant effect on the age by: group (patients/control) (F=4,5; p<0,05) and gender (F=20.7; p<0.001). There was also an interaction between the group and the gender of the subjects (F=31.1; p<0.001). There were no statistically significant differences in age (p=0.87) in the control group, but in the study group women were statistically significantly older (45±12 vs 38±12; p<0.001). There was a statistically significantly higher number of women in the control group than in the study group (51.5% vs 39.9%, χ2=11.4, p<0.001).
We compared the distribution of genotypes and alleles for the polymorphism −181 A/C (rs4354668) of the SLC1A2 and rs2486001 of the SLC6A9 in both the patients and control groups. There were no statistically significant differences in the frequency of genotypes and alleles between the patients and controls for SLC1A2 and SLC6A9 polymorphisms in either the entire sample or after stratification according to gender (Tables 1 and 2). However, in the subgroup of women, a trend for an effect of −181 A/C genotype on the development of schizophrenia was observed. .
 | Table 1 SLC1A2 (rs4354668) SNP genotypes and allele distribution in patient and control groups |
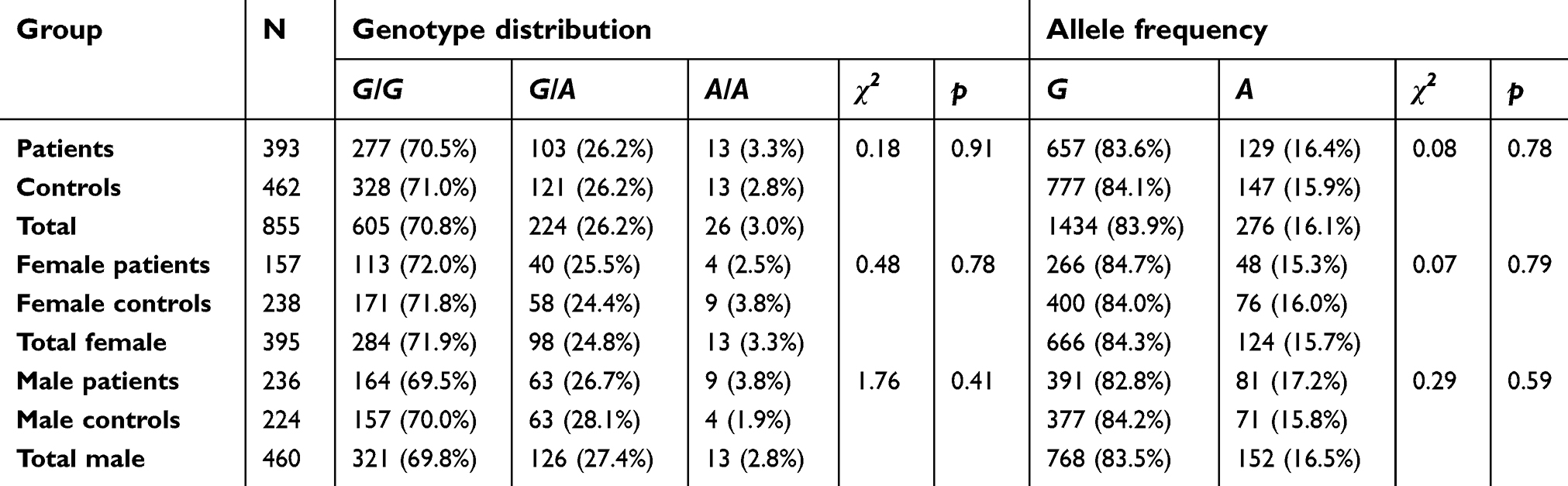 | Table 2 SLC6A9 (rs2486001) SNP genotypes and allele distribution in patient and control group |
The genotype distributions of 2 SNPs did not depart significantly from the HWE both in the patients (−181 A/C [p=0.61], rs2486001 [p=0.38]) and in the controls (−181 A/C [p=0.34], rs1008438 [p=0.65]).
We further tested the potential associations between schizophrenia and individual polymorphisms based on different genetic inheritance models (co-dominant, dominant, recessive, over-dominant and additive) in the entire group, and separately in the group of women and men. No significant differences were found for any of the models analyzed.
The analysis showed no linkage disequilibrium between the two polymorphisms neither in the entire sample (D′=0.0242; r=0.0124; P=0.608) nor assessed separately in the female (D′=0.0580, r=−0.0209, P=0.556) and male group (D′=0.0777, r=0.0397, P=0.229). The results of the haplotype associations tests (in the entire group with men/women comparison, and also separately in women and men group) showed no statistically significant higher risk for schizophrenia development for three haplotypes in relation to the most frequent haplotype AG. However, men with CA haplotype had more than 1.5 higher risk to develop schizophrenia than women (OR=1.63 [95% CI=1.17–2.27, p<0.05]).
Correlation between gender, genotypes and clinical presentation of the disease
The results of the ANOVA showed a statistically significant impact of gender on the onset of first schizophrenia episode (F=25.5; p<0.001), but no statistically significant effect of genotype of −181 A/C polymorphism in the SLC1A2 (F=0.43, p=0.652), or gender x genotype interaction (F=0.10; p=0.906). The onset of schizophrenia occurred earlier in men than in women (24±6 vs 28±8 years). For rs2486001 polymorphism in the SLC6A9, such analysis was not performed due to the small number of patients with A/A genotype.
There was no effect of gender or genotypes of both analyzed polymorphisms on the individual dimensions of the PANSS scale (Table 3). There was a significant weak positive correlation between the age of onset and negative symptoms dimension values (r=0.118; p<0.05) and a significant weak negative correlation between the age of onset and the score in the emotional distress dimension (r=−0.117; p<0.05).
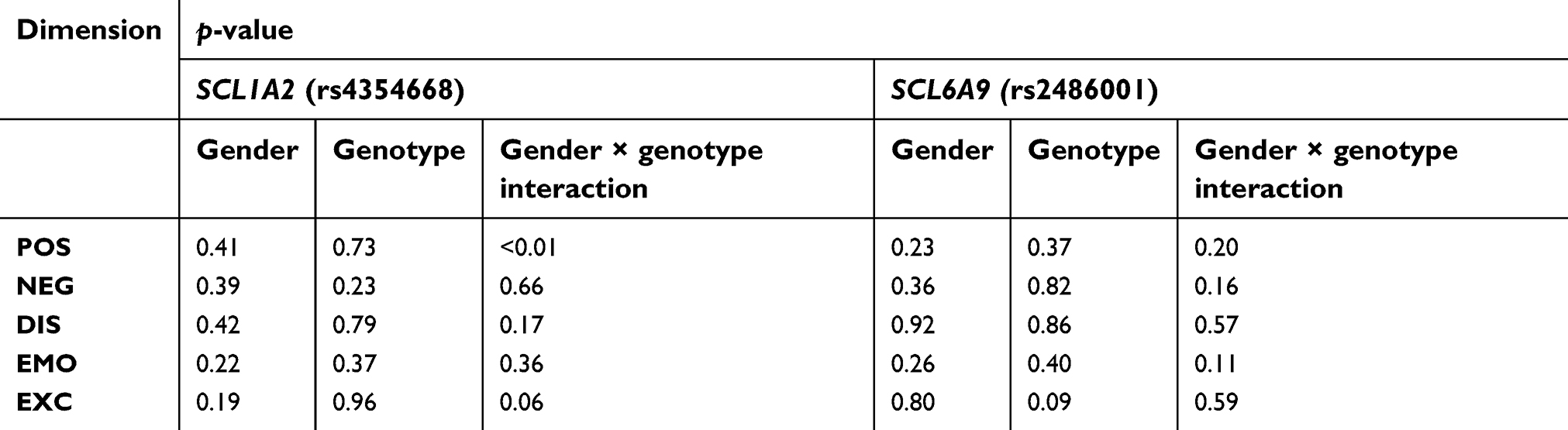 | Table 3 The results of two–way ANOVA (gender and genotype distribution) for PANSS dimensions |
Analysis of suicide attempts
The distribution of genotypes and alleles in the group of patients with a history of suicide attempts and nonsuicidal group was analyzed. There were no statistically significant differences in the genotype and allele frequencies between groups for both analyzed polymorphisms. Results of five different logistic regression models, evaluating the distribution of the genotypes combination and ORs for suicide attempt, showed no statistical significance for any of the models presented for genotypes of both polymorphisms. Also, no statistical significant higher risk of suicidal attempt was found for three haplotypes in relation to the most frequent haplotype AG.
Individuals who had a suicide attempt were younger with a shorter duration of illness. There were no statistically significant differences in family history, gender, and time of schizophrenia onset. These individuals had higher scores in EMO dimension of the PANSS scale. For other dimensions, there were no statistically significant differences (Table 4).
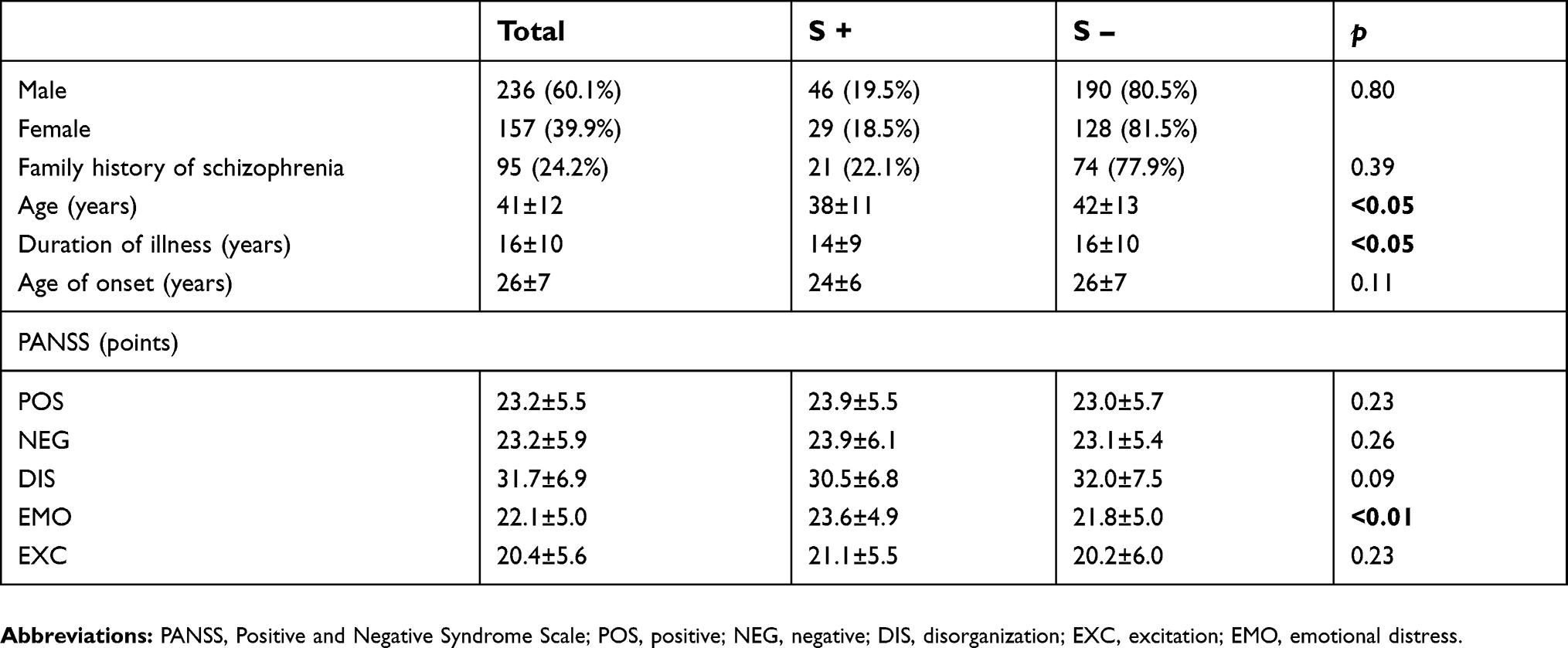 | Table 4 Analysis of the clinical data and PANSS dimensions with respect to suicide attempts |
Our analysis showed that the risk of committing suicide attempt decreases with age of onset (OR=0.973; 95% CI: 0.953–0.994; p<0.05). The Hosmer–Lemeshov match test proved to be statistically insignificant, confirming the good fit of the logistic regression model (p=0.116).
Also the risk of suicide attempt decreases with each year of schizophrenia (OR=0.971, 95% CI: 0.945–0.997, p<0.05). The Hosmer–Lemeshov match test proved to be statistically insignificant, confirming the good fit of the logistic regression model (p=0.391).
Discussion
Analysis of the distribution of alleles and genotypes for the −181 A/C polymorphism of the SLC1A2 among schizophrenic patients and healthy volunteers showed no statistically significant differences, both in the whole group and after stratification according to gender. This indicates a lack of association of the mentioned polymorphism with the risk of developing schizophrenia in the Polish population. One of the polymorphisms analyzed in our study: −181 T/G (lead strand DNA) (rs4354668) was also evaluated in a group of 211 Italian Caucasians diagnosed with schizophrenia. Due to the lack of a control group, only the distribution of individual genotypes was given: 77 patients with genotype T/T (36.5%), 88 T/G (41.7%) and 46 G/G (21.8%). The results obtained in the current work can be confronted only in terms of the frequency of individual genotypes. The distribution of genotype and alleles in this study, including a twice larger group of schizophrenic patients (AA 32.8%, AC 47.8%, CC 19.4%) and alleles (A 56.7% and C 43.3%), was slightly different, with a predominance of AC heterozygotes and a lower percentage of AA and CC homozygotes. At the same time, the frequency of the C allele (43.3%) was slightly higher than in the study of Italian researchers (42%),42 where the frequency was almost the same as in our control group (A 58.7%, C 41.3%) and the European population (T 58%, G 42%).43 In the association study of −181 T/G polymorphism conducted in the group of Asian schizophrenic patients, statistically significant differences in the distribution of both genotypes and alleles between the study and control groups were found. It was demonstrated that the presence of the G allele was associated with an increased risk of schizophrenia in the Chinese population. In further analysis, it was noted that the polymorphism rs4354668 shows a similar association with schizophrenia regardless of the gender of the examined persons.44 In another association studies of the SLC1A2 with schizophrenia in the Japanese population, for none of the analyzed polymorphisms no statistically significant differences in the distribution of both alleles and genotypes between the patients and control groups could be demonstrated.45,46 In conclusion, in contrast to the obtained results in the study of the Chinese population, in which the −181 A/C polymorphism of the SLC1A2 was associated with schizophrenia, in our study of the Caucasian schizophrenia patients. a similar compound with the disease was not found.
In the current study, the polymorphism rs2486001 in the SLC6A9 was analyzed. There were no statistically significant differences in the prevalence of genotypes and alleles between schizophrenic patients and healthy volunteers, both in the whole group and after stratification according to gender.
To the best of our knowledge, this is the first association study of SLC6A9 polymorphisms and schizophrenia in Caucasian individuals. Similarly, no studies have been published on association of rs2486001 polymorphism of the SLC6A9 with schizophrenia among Asian patients. Few published association studies of polymorphisms of the GlyT1 encoding gene, not analyzed in this paper, apply only to schizophrenia patients of Asian ethnicity. Deng et al47 studied 3 SNPs (rs783307, rs2248829 and rs7555) in the SLC6A9, without noticeable differences in the distribution of alleles and genotypes between the groups for any of them. Also, Tsai et al48 in the study of 4 polymorphisms (rs1766967, rs16831541, rs2248632 and rs2248253) of the SLC6A9 did not obtain statistically significant differences in the distribution of genotypes and alleles. The authors concluded that there is no evidence of association of the SLC6A9 with schizophrenia in the Chinese population. Similar conclusions are derived from the study of Korean schizophrenia patients with no difference in genotypic and allelic distributions for rs2248829 of the SLC6A9.49 A study of the Japanese population indicated a significant association of rs2486001 polymorphism with a higher risk of developing post-amphetamine psychosis.40
As described previously (see background), a number of different loci related to glutamatergic transmission have been identified by GWAS.10 GWAS is an epidemiological study conducted on a very large population, but often heterogeneous in contrast to candidate gene case–control association studies. Because genetic marker frequencies can differ between individuals with different ethnic background, population stratification can lead to false-positive associations and/or mask true associations. It should be emphasized that our study was carried out on a highly homogeneous population, which minimizes the risk of false results due to population differences. Moreover, an important feature of GWAS is stringent selection criteria of significance which may lead to false-negative results. For this reason, the direct comparison of our moderate sample size case–control study with large-scale GWAS or meta-analyses will be rather difficult. However, data obtained in our study may be used in the future meta-analyses of association studies.
Numerous studies indicate differences in the clinical picture and course of schizophrenia between men and women,50,51 which emphasizes slightly higher risk among men in whom the disease develops earlier. Men are also more likely to abuse psychoactive substances. Milder course of the disease among women means longer periods of remission and less risk of recurrence. Some authors highlight greater severity of negative symptoms of schizophrenia in men and affective and positive symptoms in women. There is also evidence of gender differences in gene expression in the PFC of schizophrenia patients.52 We analyzed the effect of gender and genotype of the −181 A/C polymorphism on the age of onset in the schizophrenia group. Statistically significant earlier age of onset among men compared to women (24±6 vs 28±8 years) was observed. There were no statistically significant effects of genotypes of the −181 A/C polymorphism or gender/genotype interaction on the analyzed parameter. Studies of other authors examining the −181 T/G polymorphism (T/G: DNA of the leading strand) of the SLC1A2 contain averaged clinical data with no significant differences between the groups (homozygous for allele T and homo- and heterozygous for allele G) with respect to analyzed variables.53,54 Poletti et al42 also describes averaged clinical data and mentions the lack of statistically significant differences between GG, GT and TT genotypes. Deng et al45 omit the course of the disease. Nagai et al46 showed no statistically significant differences in the distribution of genotypes in the 6 SNPs of the SLC1A2 in individual subtypes of schizophrenia, or their effects on clinical variables such as time of onset and duration of illness, duration of untreated psychosis or number of relapses. Regarding −181 A/C polymorphism of the SLC1A2 the presence of allele A in healthy Japanese, especially in women, was associated with higher transporter activity and thus lower glutamate plasma levels, and promoted the occurrence of personality traits such as introversion, independence and emotional coldness.41 Another study of −181 A/C polymorphism of the SLC1A2 in Caucasian patients diagnosed with bipolar disorder showed higher recurrence rates in homozygotes for the polymorphic allele.55 Earlier studies already pointed to the genetic similarity and co-occurrence of candidate genes between bipolar disorder and schizophrenia. Shao et al56 showed an altered expression of the SLC1A2 in the DLPFC of patients in the bipolar and schizophrenia group. Impairment of glutamate uptake due to dysfunction or decreased expression of EAAT2 has been postulated in the pathogenesis of many other neurodegenerative diseases such as Alzheimer’s disease, Huntington’s disease or amyotrophic lateral sclerosis.57
In case of SNP rs2486001 of the SLC6A9, the analysis of influence of gender and genotype on the age of onset of schizophrenia was not performed due to the small number of subjects with the A/A genotype in the study group.. In the previous association studies of SLC6A9 polymorphisms in schizophrenia among patients of Asian ethnicity, no clinical data were available (onset and duration of illness).
Due to the heterogeneous nature of schizophrenia, the PANSS scale has been introduced as an operatized, standardized research instrument that should provide a balanced representation of positive and negative symptoms, their relative proportions and their relationship to global psychopathology. Initial groups of symptoms: positive, negative and general do not fully reflect the complexity of the disease, which resulted in the development of the 3-, 4-, and then 5-dimensional PANSS scale. The dimension description of schizophrenia was supported by two main premises: it is complete, comprehensive and does not lead to the loss of any part of the clinical picture.58 The 5-dimensional PANSS structure is more accurate reflecting clinical reality, and this model has also been most frequently used in studies over the past decade.59 The 5-dimensional model proposed by van der Gaag38 used in this paper seems to comprehensively describe the clinical picture of schizophrenia. So far, no association studies of the glutamatergic system with schizophrenia which use this model have been carried out. The ANOVA results for both examined SNPs showed no effect of gender and genotype on the values of individual dimensions of PANSS. However, a significant weak positive correlation was found between the age of the onset and the magnitude of the negative dimension, and a significant negative for the emotional distress dimension. In other studies analyzing EAAT2 gene polymorphisms in the Caucasian population, no gender division was introduced, and each author provided a collective value of the PANSS scale using a classical three-dimensional division. This prevents direct reference to the results obtained by these researchers. In the association study of −181 A/C polymorphism of the SLC1A2 among Asian schizophrenia patients the PANSS has not been used,44 this is the case also in studies including other SNPs of the gene.45,46
In the current work, cognitive functions were not evaluated with neuropsychological tests in the study group; however, according to other researchers, the disorganization dimension could be identified as describing some aspects of cognitive dysfunctions. Van der Gaag38 in the 5-dimensional model of the PANSS scale includes symptoms that are considered to be determinants of cognitive impairment in the DIS dimension, although he does not use a similar comparison. In the results of the present study, genotypes and alleles of both examined polymorphisms were not related to the values of disorganization in the study group. In the Asian population, Zhang et al44 found a statistically significant influence of the −181 A/C polymorphism of the SLC1A2 on the results in WCST. Homozygotes of the dominant T allele obtained better results in comparison to persons with the polymorphic G allele, both in the patients and in the control group. The direct translation of disorganization dimension into a determinant of patients cognitive functioning would be a too reductionist approach, especially in the case of incompatibility of individual authors as to the quantity and quality of symptoms forming this dimension.
The distribution of genotypes and alleles in the group of patients with a history of suicide attempts and nonsuicidal group did not show statistically significant differences in the genotype and allele frequencies between groups for both analyzed polymorphisms. So far, no studies have been published which would describe the influence of polymorphisms of glutamatergic system components genes on the risk of suicide attempts in the group of schizophrenia patients. On the other hand, our results are consistent with the existing data, which include younger age, symptoms of depression, sense of hopelessness or guilt, anxiety, insomnia and emotional tension as the risk factors for a suicidal attempt in schizophrenia. The older age at the time of the onset of schizophrenia is considered to be poorly related, while the duration of the disease on the risk of suicide is ambiguous.60 Among Korean patients diagnosed with schizophrenia, there was a significant correlation of the occurrence of suicidal thoughts with the following symptoms of the PANSS scale: G1, G2, G3, G6.61 All of them are included in the EMO dimension of the model we used in our analysis.
This study shows several limitations which include the relatively small sample size of the study group, which may contribute to false-negative or false-positive results. Another limitation may be the less number of female patients included in the study. Neuropsychological tests of cognitive functioning, which could enrich the results and indicate possible association with polymorphisms, were also not performed. Element hampering comparison of the obtained results is the choice of a 5-dimensional model of the PANSS scale proposed by van der Gaag.38 Some of the presented results tended to be statistically significant, and it is not known whether increasing the size of the studied population could contribute to the statistical significance of the examined parameters. However, we conducted our study on a homogenous patient group of Caucasian ethnicity, with the presence of a control group. So far, no association studies of SLC6A9 polymorphisms in schizophrenia regarding Caucasian patients have been published. Previous association studies of SLC1A2 polymorphism in schizophrenia were performed only in patients. We broadly analyzed the psychopathology of schizophrenia in association with polymorphisms, as well as their impact on suicide attempts. Further analyses performed with larger samples are still needed. It would be also interesting to analyze the impact of the polymorphisms on other clinical features (number of episodes and their duration and duration of complete/incomplete remission) of schizophrenia.
Conclusions
In conclusion, the current study showed no association of polymorphism −181 A/C (rs4354668) in SLC1A2 and rs2486001 polymorphism in SLC6A9 with susceptibility to schizophrenia development. The presence of haplotype CA in men was associated with an increased risk of developing schizophrenia compared to women. There was no effect of genotypes of both studied polymorphisms on the PANSS scale factors. There was also no association of both polymorphisms with attempting suicide by patients. The results of the present study did not indicate an association of studied polymorphisms with onset of schizophrenia and its psychopathology in a Polish Caucasian population.
Abbreviations list
AMPAR, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor; DLPFC, dorsolateral prefrontal cortex; DSM-5, Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition); EAAT, excitatory amino acid transporter; GlyT, glicyne transporter; NMDAR, N-methyl-D-aspartate receptor; PANSS, Positive and Negative Syndrome Scale; PCP, phencyclidine; PCR-RFLP, polymerase chain reaction–restriction fragment length polymorphism; PFC, prefrontal cortex; WCST, Wisconsin Card Sorting Test.
Ethics approval and informed consent
The research was approved by the Bioethical Commission of the Medical University of Silesia in Katowice (under approval number KNW/0022/KB1/34/14 from 22/04/2014).
All participants provided written informed consent, in accordance with the Declaration of Helsinki.
Data availability
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors would like to thank the patients for their cooperation in this study, without whom this work would not be possible. This work was supported by a grant from Medical University of Silesia (KNW-1-101/N/3/0). The university had no role in study design, in the collection, analysis and interpretation of data, in the writing of the report, or in the decision to submit the paper for publication.
This paper was presented at the World Psychiatric Association 17th World Congress of Psychiatry, Berlin, October 8–10, 2017, as a poster presentation. The poster’s abstract was not published in any journal.
Author contributions
WM, KK and TM conceptualized and designed the study, provided DNA samples and clinical data, and contributed equally to drafting and revising the manuscript. AO contributed to interpreting the data and statistical analyses. MK and JK selected the SNPs, performed genotyping and prepared the genetic analysis. All authors contributed toward data analysis, drafting and critically revising the paper, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sullivan PF, Kendler KS, Neale MC. Schizophrenia as a complex trait: evidence from a meta-analysis of twin studies. Arch Gen Psychiat. 2003;60:1187–1192. doi:10.1001/archpsyc.60.12.1187
2. Stahl SM. Essential Psychopharmacology.
3. Fairman WA, Amara SG. Functional diversity of excitatory amino acid transporters: ion channel and transport modes. Am J Physiol. 1999;277:481–486. doi:10.1152/ajpheart.1999.277.5.H2002
4. Javitt DC, Zukin SR. Recent advances in the phencyclidine model of schizophrenia. Am J Psychiatry. 1991;148:1301–1308. doi:10.1176/ajp.148.10.1301
5. Krystal JH, Perry EB
6. Krystal JH, Karper LP, Seibyl JP, et al. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch Gen Psychiatry. 1994;51:199–214. doi:10.1001/archpsyc.1994.03950030035004
7. Lahti AC, Weiler MA, Tamara Michaelidis BA, Parwani A, Tamminga CA. Effects of ketamine in normal and schizophrenic volunteers. Neuropsychopharmacology. 2001;25:455–467. doi:10.1016/S0893-133X(01)00243-3
8. Moghaddam B, Adams B, Verma A, Daly D. Activation of glutamatergic neurotransmission by ketamine: a novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. J Neurosci. 1997;17:2921–2927. doi:10.1523/JNEUROSCI.17-08-02921.1997
9. Stefansson H, Ophoff RA, Steinberg S, et al. Common variants conferring risk of schizophrenia. Nature. 2009;460(7256):744–747. doi:10.1038/nature08186
10. Ripke S, Sanders AR, Kendler KS, et al. Genome-wide association study identifies five new schizophrenia loci. Nat Genet. 2011;43:969–976. doi:10.1038/ng.940
11. Schijven D, Kofink D, Tragante V, et al. Comprehensive pathway analyses of schizophrenia risk loci point to dysfunctional postsynaptic signaling. Schizophr Res. 2018;199:195–202. doi:10.1016/j.schres.2018.03.032
12. Ripke S, Neale BM, Corvin A, et al. Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511:421–427. doi:10.1038/nature13595
13. Ohi K, Hashimoto R, Ikeda M, et al. Glutamate networks implicate cognitive impairments in schizophrenia: genome-wide association studies of 52 cognitive phenotypes. Schizophr Bull. 2015;41(4):909–918. doi:10.1093/schbul/sbu171
14. Tarabeux J, Kebir O, Gauthier J, et al. Rare mutations in N-methyl-D-aspartate glutamate receptors in autism spectrum disorders and schizophrenia. Transl Psychiatry. 2011;1:e55. doi:10.1038/tp.2011.52
15. Van Horn MR, Sild M, Ruthazer ES. D-serine as a gliotrans- mitter and its roles in brain development and disease. Front Cell Neurosci. 2013;7:39.
16. Rose EM, Koo JC, Antflick JE, Ahmed SM, Angers S, Hampson DR. Glutamate transporter coupling to Na, K-ATPase. J Neurosci. 2009;29(25):8143–8155. doi:10.1523/JNEUROSCI.1081-09.2009
17. Danbolt NC. Glutamate uptake. Prog Neurobiol. 2001;65(1):1–105. doi:10.1016/S0301-0082(00)00067-8
18. Nakagawa T, Kaneko S. SLC1 glutamate transporters and diseases: psychiatric diseases and pathological pain. Curr Mol Pharmacol. 2013;6(2):66–73. doi:10.2174/18744672113069990033
19. Arriza JL, Eliasof S, Kavanaugh MP, Amara SG. Excitatory aminoacid transporter 5, a retinal glutamate transporter coupled to a chloride conductance. Proc Natl Acad Sci USA. 1997;94:4155–4160. doi:10.1073/pnas.94.8.4155
20. Lauriat TL, Dracheva S, Chin B, Schmeidler J, McInnes LA, Haroutunian V. Quantitative analysis of glutamate transporter mRNA expression in prefrontal and primary visual cortex in normal and schizophrenic brain. Neuroscience. 2006;(137):843–851. doi:10.1016/j.neuroscience.2005.10.003.
21. Sheldon AL, Robinson MB. The role of glutamate transporters in neurodegenerative diseases and potential opportunities for intervention. Neurochem Int. 2007;51:333–355. doi:10.1016/j.neuint.2007.03.012
22. Rothstein JD, Martin L, Levey AI, et al. Localization of neuronal and glial glutamate transporters. Neuron. 1994;13:713–725. doi:10.1016/0896-6273(94)90038-8
23. Domercq M, Etxebarria E, Perez-Samartin A, Matute C. Excitotoxic oligodendrocyte death and axonal damage induced by glutamate transporter inhibition. Glia. 2005;52:36–46. doi:10.1002/glia.20221
24. Bauer D, Haroutunian V, Meador-Woodruff JH, et al. Abnormal glycosylation of EAAT1 and EAAT2 in prefrontal cortex of elderly patients with schizophrenia. Schizophr Res. 2010;117:92–98. doi:10.1016/j.schres.2009.07.025
25. Shan D, Lucas EK, Drummond JB, et al. Abnormal expression of glutamate transporters in temporal lobe areas in elderly patients with schizophrenia. Schizophr Res. 2013;144:1–8. doi:10.1016/j.schres.2012.12.019
26. Rao JS, Kellom M, Reese EA, et al. Dysregulated glutamate and dopamine transporters in postmortem frontal cortex from bipolar and schizophrenic patients. J Affect Disord. 2012;136:63–71. doi:10.1016/j.jad.2011.08.017
27. Ohnuma T, Tessler S, Arai H, Faull RL, McKenna PJ, Emson PC. Gene expression of metabotropic glutamate receptor 5 and excitatory amino acid transporter 2 in the schizophrenic hippocampus. Brain Res Mol Brain Res. 2000;85(1–2):24–31. doi:10.1016/S0169-328X(00)00222-9
28. Meyer T, Ludolph AC, Morkel M, Hagemeier C, Speer A. Genomic organization of the human excitatory amino acid transporter gene GLT-1. Neuroreport. 1997;8:775–777. doi:10.1097/00001756-199702100-00039
29. Mallolas J, Hurtado O, Castellanos M, et al. A polymorphism in the EAAT2 promoter is associated with higher glutamate concentrations and higher frequency of progressing stroke. J Exp Med. 2006;203(3):711–717. doi:10.1084/jem.20051979
30. Zafra F, Gomeza J, Olivares L, Aragón C, Giménez C. Regional distribution and developmental variation of the glycine transporters GLYT1 and GLYT2 in the rat CNS. Eur J Neurosci. 1995;7(6):1342–1352. doi:10.1111/j.1460-9568.1995.tb01125.x
31. Danysz W, Parsons AC. Glycine and N-methyl-D-aspartate receptors: physiological significance and possible therapeutic applications. Pharmacol Rev. 1998;50:597–664.
32. Waziri R, Baruah S, Sherman AD. Abnormal serine-glycine metabolism in the brains of schizophrenics. Schizophr Res. 1993;8(3):233–243. doi:10.1016/0920-9964(93)90021-A
33. Heresco-Levy U, Javitt DC. Comparative effects of glycine and D-cycloserine on persistent negative symptoms in schizophrenia: a retrospective analysis. Schizophr Res. 2004;66(2–3):89–96. doi:10.1016/S0920-9964(03)00129-4
34. Williams JB, Mallorga PJ, Conn PJ, Pettibone DJ, Sur C. Effects of typical and atypical antipsychotics on human glycine transporters. Schizophr Res. 2004;71(1):103–112. doi:10.1016/j.schres.2004.01.013
35. Tsai G, Lane HY, Yang P, Chong MY, Lange N. Glycine transporter I inhibitor, N-methylglycine (sarcosine), added to antipsychotics for the treatment of schizophrenia. Biol Psychiatry. 2004;55:452–456. doi:10.1016/j.biopsych.2003.09.012
36. Nelson N. The family of Na+/Cl- neurotransmitter transporters. J Neurochem. 1998;71(5):1785–1803. doi:10.1046/j.1471-4159.1998.71051785.x
37. Kay SR, Fiszbein A, Opler LA. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull. 1987;13(2):261–276. doi:10.1093/schbul/13.2.261
38. van der Gaag M, Hoffman T, Remijsen M, et al. The five-factor model of the positive and negative syndrome scale II: a ten-fold cross-validation of a revised model. Schizophr Res. 2006;85:280–287. doi:10.1016/j.schres.2006.03.021
39. Dumais A, Lesage AD, Lalovic A, et al. Is violent method of suicide a behavioral marker of lifetime aggression? Am J Psychiatry. 2005;162(7):1375–1378. doi:10.1176/appi.ajp.162.7.1375
40. Morita Y, Ujike H, Tanaka Y, et al. The glycine transporter 1 gene (GLYT1) is associated with methamphetamine-use disorder. Am J Med Genet B Neuropsychiatr Genet. 2008;147B(1):54–58. doi:10.1002/ajmg.b.30565
41. Matsumoto Y, Suzuki A, Ishii G, Oshino S, Otani K, Goto K. The −181 A/C polymorphism in the excitatory amino acid transporter-2 gene promoter affects the personality trait of reward dependence in healthy subjects. Neurosci Lett. 2007;427(2):99–102. doi:10.1016/j.neulet.2007.09.015
42. Poletti S, Radaelli D, Bosia M, et al. Effect of glutamate transporter EAAT2 gene variants and gray matter deficits on working memory in schizophrenia. Eur Psychiatry. 2014;29(4):219–225. doi:10.1016/j.eurpsy.2013.07.003
43.
44. Zhang B, Guan F, Chen G, et al. Common variants in SLC1A2 and schizophrenia: association and cognitive function in patients with schizophrenia and healthy individuals. Schizophr Res. 2015;169(1–3):128–134. doi:10.1016/j.schres.2015.10.012
45. Deng X, Shibata H, Ninomiya H, et al. Association study of polymorphisms in the excitatory amino acid transporter 2 gene (SLC1A2) with schizophrenia. BMC Psychiatry. 2004;4:21. doi:10.1186/1471-244X-4-21
46. Nagai Y, Ohnuma T, Karibe J, et al. No genetic association between the SLC1A2 gene and Japanese patients with schizophrenia. Neurosci Lett. 2009;463(3):223–227. doi:10.1016/j.neulet.2009.07.092
47. Deng X, Sagata N, Takeuchi N, et al. Association study of polymorphisms in the neutral amino acid transporter genes SLC1A4, SLC1A5 and the glycine transporter genes SLC6A5, SLC6A9 with schizophrenia. BMC Psychiatry. 2008;8:58. doi:10.1186/1471-244X-8-58
48. Tsai SJ, Cheng CY, Hong CJ, Liao DL, Liou YJ. Polymorphisms in glycine transporter with schizophrenia. Neuropsychopharmacol Hung. 2006;8(1):17–21.
49. Kang WS, Park HJ, Park JK, Cho AR, Kim JW. Association study between solute carrier family 6, member 9 (SLC6A9) gene and schizophrenia in Korean population (abstract). 10th World Congress of Biological Psychiatry. 2011. doi: 10.3284/wfsbp.2011.1
50. Abel KM, Drake R, Goldstein JM. Sex differences in schizophrenia. Int Rev Psychiatry. 2010;22(5):417–428. doi:10.3109/09540261.2010.515205
51. Ochoa S, Usall J, Cobo J, Labad X, Kulkarni J. Gender differences in schizophrenia and first-episode psychosis: a comprehensive literature review schizophrenia research and treatment. Schizophr Res Treat. 2012;2012:916198.
52. Qin W, Liu C, Sodhi M, Lu H. Meta-analysis of sex differences in gene expression in schizophrenia. BMC Syst Biol. 2016;10(1):100–106. doi:10.1186/s12918-016-0304-1
53. Spangaro M, Bosia M, Zanoletti A, et al. Cognitive dysfunction and glutamate reuptake: effect of EAAT2 polymorphism in schizophrenia. Neurosci Lett. 2012;522(2):151–155. doi:10.1016/j.neulet.2012.06.030
54. Spangaro M, Bosia M, Zanoletti A, et al. Exploring effects of EAAT polymorphisms on cognitive functions in schizophrenia. Pharmacogenomics. 2014;15(7):925–932. doi:10.2217/pgs.14.42
55. Dallaspezia S, Poletti S, Lorenzi C, et al. Influence of an interaction between lithium salts and a functional polymorphism in SLC1A2 on the history of illness in bipolar disorder. Mol Diagn Ther. 2012;16(5):303–309. doi:10.1007/s40291-012-0004-5
56. Shao L, Vawter MP. Shared gene expression alterations in schizophrenia and bipolar disorder. Biol Psychiatry. 2008;64(2):89–97. doi:10.1016/j.biopsych.2007.11.010
57. Kim K, Lee SG, Kegelman TP, et al. Role of excitatory amino acid transporter-2 (EAAT2) and glutamate in neurodegeneration: opportunities for developing novel therapeutics. J Cell Physiol. 2011;226(10):2484–2493. doi:10.1002/jcp.22609
58. Łoza B. Nowe wymiary leczenia schizofrenii. Psychiatria. 2005;1:1–8.
59. Wallwork RS, Fortgang R, Hashimoto R, Weinberger DR, Dickinson D. Searching for a consensus five-factor model of the positive and negative syndrome scale for schizophrenia. Schizophr Res. 2012;137(1–3):246–250. doi:10.1016/j.schres.2012.01.031
60. Hor K, Taylor M. Suicide and schizophrenia: a systematic review of rates and risk factors. J Psychopharmacol. 2010;24:81–90. doi:10.1177/1359786810385490
61. Kim SW, Kim SJ, Mun JW, et al. Psychosocial factors contributing to suicidal ideation in hospitalized schizophrenia patients in Korea. Psychiatry Invest. 2010;7(2):79–85. doi:10.4306/pi.2010.7.2.79
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.