Back to Journals » Neuropsychiatric Disease and Treatment » Volume 11
Association between toll-like receptor 4 expression and symptoms of major depressive disorder
Authors Wu M, Huang T, Huang K, Huang Y, Hung Y ![]()
Received 12 May 2015
Accepted for publication 12 June 2015
Published 30 July 2015 Volume 2015:11 Pages 1853—1857
DOI https://doi.org/10.2147/NDT.S88430
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Wai Kwong Tang
Ming-Kung Wu,1 Tiao-Lai Huang,1,2 Kai-Wei Huang,3 Ya-Ling Huang,1 Yi-Yung Hung1,4
1Department of Psychiatry, Kaohsiung Chang Gung Memorial Hospital and Chang Gung University College of Medicine, 2Genomic and Proteomic Core Laboratory, Department of Medical Research, Kaohsiung Chang Gung Memorial Hospital, 3Department of Nursing, Kaohsiung Chang Gung Memorial Hospital and Chang Gung University College of Medicine, 4Graduate Institute of Clinical Medical Sciences, Chang Gung University, College of Medicine, Kaohsiung, Taiwan
Background: In our previous study, toll-like receptor 4 (TLR4) mRNA expression level was associated with severity of major depressive disorder (MDD) evaluated with the 17-item Hamilton Depression Rating Scale (HAMD-17). However, there are few studies that have investigated the relationship between symptoms of MDD and changes in TLR4 expression. Therefore, the aim of the present study was to further analyze the association between subscales of HAMD-17 and TLR4.
Methods: Fifty-one patients with MDD (15 male and 36 female) participated in this study. HAMD-17 was used to assess the symptoms of major depression. The mRNA expression levels of TLR4 were examined in parallel with a housekeeping gene, using real-time polymerase chain reaction. A stepwise linear regression forward model was used to evaluate the relationships between items of HAMD-17 and TLR4 expression.
Results: Some sickness behavior-associated symptoms, including suicide, somatic symptoms of anxiety, or performance of work and activities, were not associated with TLR4 expression. However, psychological signs of anxiety and loss of weight in HAMD-17 can predict the expression level of TLR4.
Conclusion: Our results suggest a significant association between anxiety, body weight loss, and TLR4 mRNA levels in patients with MDD. Larger longitudinal studies combining both subjective and objective measures of depression are needed to clarify the link between TLR4 and symptoms of depression.
Keywords: major depressive disorder, innate immune, toll-like receptor, anxiety
Introduction
Major depressive disorder (MDD) is one of the most serious psychiatric diseases throughout the world, and causes numerous socioeconomic burdens. The most serious consequence of MDD is increased risk of suicide. The etiology and pathophysiology of MDD still remains unclear at the present time. There is evidence from numerous studies suggesting that the etiology of MDD is heterogeneous. Despite the hypothesis of disturbances in neurotransmitter systems and alterations in levels of neurotrophic factors, there are an increasing number of reports to suggest that immune abnormalities also play a role in MDD. This is not only supported by animal studies, but has also been shown in human investigations. In a recent review, whereby the literature on this subject was summarized, it was suggested that inflammatory activity can increase anhedonia-like behavior in animal models.1 Similarly, in humans, another meta-analysis (conducted by Dowlati et al2) found that the proinflammatory cytokines, tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6), were significantly higher in depressed than in control subjects. Similar changes in C-reactive protein in depressive patients were also found in another meta-analysis.3 Moreover, psychosocial and physiological stressors – which are two of the most frequently mentioned etiological factors in MDD – can induce the production of proinflammatory mediators, such as IL-1 and reactive oxygen species, and hypothalamic–hypophyseal–adrenal axis disturbances in the brain.4 Moreover, disturbances of the immune system impaired neurotransmitter metabolism and neural circuits that regulate mood.5 For example, increases in cytokine and inflammatory mediators decrease the production of serotonin.6 All the evidence indicates that the alterations – either primary or secondary to outside stimulation – affect internal regulation in the brain and result in mood abnormalities.
The toll-like receptors (TLRs) are one of the protein types expressed in the circumventricular organs and the choroid plexus.7 TLRs are considered one of the pattern recognition proteins of the innate immune response, which can defend against external microorganisms8 and recognize endogenous ligands, to mediate the inflammatory response to stress.9 The TLR family contains various subtypes, including TLR2 (which binds with bacterial lipoproteins and lipoteichoic acid), TLR3 (which recognizes double-stranded RNA), and TLR7 (which recognizes single-stranded DNA).10 Because of their potential roles in neuropsychiatric disease, TLRs are attracting increasing attention. Alterations in TLRs have been found in psychiatric disorders, such as increased expression in TLR3, TLR4, TLR5, and TLR7 in patients with MDD,11 reduced mRNA expression in schizophrenia,12 and enhanced response to ligand in psychosis.13 Moreover, in addition to alterations in TLRs, changes in adaptor proteins of TLRs have also been found. Indeed, in a report by Hajebrahimi et al14 the adaptor proteins of TLRs were found to be significantly elevated in depressed patients. In addition, genetic polymorphism of TLR4 has been investigated in bipolar disorder, which revealed that the rs1927914 AA and rs11536891 TT genotypes were more frequent in bipolar disorder patients than in controls.15
The TLR family member, TLR4, has received the most attention in recent decades. TLR4, initially discovered for its role in the recognition of bacterial lipopolysaccharide (LPS), has been found to have a regulatory role in the adrenal response to stressful inflammatory stimuli.16,17 Recently, researchers have started trying to link TLR4 with neuropsychiatric disorders. For example, McKernan et al13 pointed out that there is an enhanced response in TLR2 and TLR4 activation in schizophrenia and bipolar disorder.
Since the “leaky gut” hypothesis was put forward, the link between TLR4 and MDD is starting to become more well established.18 The “leaky gut” hypothesis states that the translocation of enterobacteria induces and activates the inflammatory and oxidative/nitrosative stress pathways through TLR4 signaling. In a recent report by Garate et al19 it was suggested that LPS from Gram-negative bacteria translocating through the gastrointestinal tract due to an increase in its permeability are responsible for TLR4 activation found in the brain. This reaction would result in increases in inflammatory mediators in the central nervous system. Moreover, in humans, another study attempted to investigate the differences in TLR4 in MDD patients, and to assess alterations with cognitive behavioral therapy alone. In this report, it was found that the expression of TLR4 was significantly higher in MDD patients than in healthy controls, and that expression was decreased with a course of cognitive behavioral therapy alone.20 In our previous report, TLR4 was shown to be the only member of this protein family to have significant association with MDD severity.11 Here, we performed further investigations to determine the associations between items of the 17-item Hamilton Depression Rating Scale (HAMD-17) and TLR4.
Methods
Subjects
In an extension of our previously published study,11 we recruited MDD patients through the Structured Clinical Interview for DSM-IV Axis I Disorders, conducted by a senior psychiatrist (Hung YY) from August 2013 to December 2014, at Kaohsiung Chang Gung Memorial Hospital Medical Center in Taiwan. Institutional Review Board approval was obtained from the hospital ethics committee (101-5012A3, 103-5114B) and informed consent was obtained for each participant. We used the HAMD-17 to evaluate the symptoms and severity of MDD.21 The exclusion criteria were 1) alcohol dependence and 2) any history of immune inflammatory disease. In order to exclude any chronic physical illness, all patients received routine physical examination, chest X-rays, electrocardiogram, and routine blood tests. At the same time, patients were checked for acute infections or allergic reactions at the time of entering this study.
Sample preparation and analysis
Blood samples were collected through venous drawing (5 mL) and stored in PAXgene blood RNA tubes in the early morning, after patients had fasted for 9 hours. The tubes were stored in a −80°C freezer immediately after blood was drawn, and the samples remained in the freezer until they were assayed. We used TRIzol reagent to isolate the RNA and performed real-time polymerase chain reaction (RT-PCR) using a set of primers for TLR4 (sense 5′-TGAGCAGTCGTGCTGGTATC-3′, antisense 5′-CAGGGCTTTTCTGAGTCGTC-3′) (Lopez et al22). The QuantiTect Reverse Transcription Kit (Qiagen, Venlo, the Netherlands) was used for the reverse transcription of RNA. The PTC-200 apparatus (Bio-Rad Laboratories, Hercules, CA, USA) and the QuantiTect SYBR Green PCR kit (Qiagen) were applied for the complete analysis. We used glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as a housekeeping gene to assess the relative abundance of mRNA.
Statistical procedure
A stepwise multiple linear regression forward model was used to judge the subscores of HAMD-17 that correlated with the TLR4 levels. All results are presented as mean ± standard deviation. Statistical significance was set at P<0.05. All statistical analyses were performed through the Statistical Product and Service Solutions (SPSS, IBM Corporation, Armonk, NY, USA), version 22.
Results
Epidemiologic data
Altogether, 51 patients with MDD were recruited. Table 1 shows baseline characteristics regarding participants’ sex, age, and body mass index (BMI) (15 male/36 female, mean age, 46.25±10.04 years; BMI, 23.90±4.96 kg/m2). Smoking (17 patients), education duration (11.24±2.96 years), HAMD-17 score (27.20±5.40), and TLR4 relative expression (1.50±1.82) are also shown in Table 1. Table 2 shows the average scores of different HAMD-17 items.
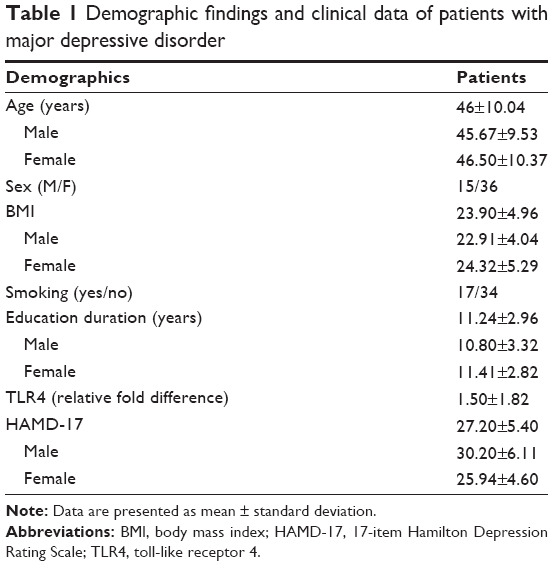 | Table 1 Demographic findings and clinical data of patients with major depressive disorder |
 | Table 2 Scores of different items in HAMD-17 |
Cox regression of TLR4
Through the cox regression model, we were able to determine whether the subscores of HAMD-17 had significant associations with the TLR4 mRNA levels (Table 3). Within the subscores of HAMD-17, the scores for psychological anxiety and loss of weight showed significant associations with peripheral TLR4 mRNA levels (P=0.25 in the model of psychological anxiety only; P=0.01 in the model for combination of items of psychological anxiety and loss of weight).
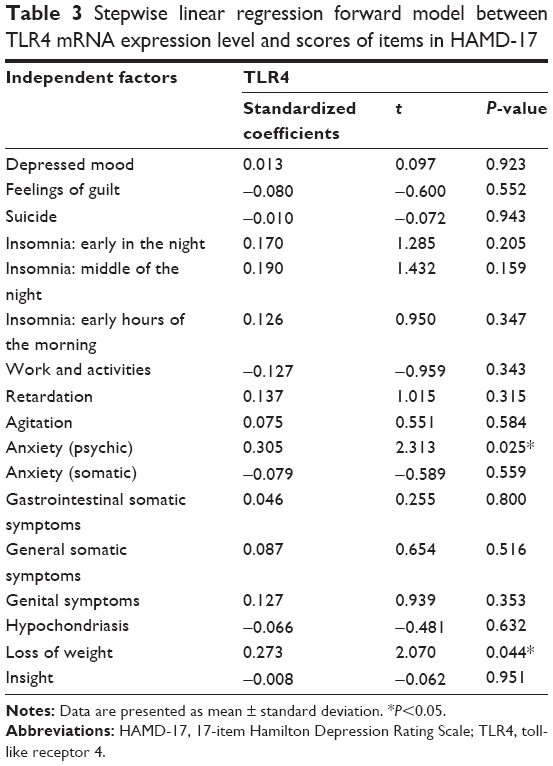 | Table 3 Stepwise linear regression forward model between TLR4 mRNA expression level and scores of items in HAMD-17 |
Discussion
To our knowledge, this is the first study to investigate the association with peripheral TLR4 mRNA levels and specific HAMD-17 subscores. The main results of our study are that psychological anxiety and loss of weight predict peripheral TLR4 mRNA expression.
In the present study, we show that anxiety has a significant association with peripheral TLR4 mRNA expression. The immune system in the gastroenteric environment plays an important role in body weight and psychological anxiety. Gastric and duodenal ulcers were found to have a correlation with the Hamilton anxiety scale in a cross-sectional study, in a population of 2,000 students at Belgrade University.23 In recent reviews, exposure to physical, physiological, as well as psychological, stressors was found to have numerous impacts on homeostasis of the intestinal microbiologic environment.24 Moreover, disruption of the intestinal microbiologic environment resulted in overproduction of inflammatory mediators and behavioral abnormalities, including anxiety-like behaviors.25,26 There are also other reports suggesting a relationship between anxiety and the innate immune system, which may partially support the results of the present study. In animal studies, lack of TLR4 is related to a reduction in anxiety-related behaviors induced by alcohol withdrawal.27 In contrast, administration of LPS or endogenous ligands of TLR4 such as high-mobility group box-1 (HMGB1) and heat shock proteins, which could activate the innate immune system through the TLR4 pathway, was found to induce sickness behaviors in an animal model,28–30 which mimicked anxiety and loss of body weight in our study.
In the present study, suicidality was not associated with TLR4 levels. In postmortem studies that used brain tissue from suicide victims, some significant findings were reported on alterations in immune activities and TLR4 levels in these individuals. Steiner et al31 demonstrated significant microgliosis in the dorsolateral prefrontal cortex (DLPFC) of suicide patients, compared with controls. One report discussed the differences in TLR4 in the postmortem brain of depressed suicide victims and depressed nonsuicide subjects. The authors suggest that both mRNA and protein expression of TLR4 are significantly increased in DLPFC of depressed suicide victims, compared with controls, which may be related to an abnormality in TLR4 expression.32 Thus, a possible cause for the difference, compared with previous studies, may be that patients who commit suicide differ from those who do not commit suicide. In a previous follow-up study, patients who committed suicide during follow-up had significantly higher Hamilton Depression Rating Scale scores for suicidality.33
There are several limitations of the present study. Firstly, the effects of distribution of different types of leukocytes on TLR4 mRNA expression were unclear based on the present work, as TLR4 mRNA expression was summated for the different cell types in the blood. The decreased percentages of lymphocytes reported in a previous study may cause deviation.34 Further studies with cell-type specific designs are therefore necessary in the future. Secondly, cytokines were not analyzed is this study. There is evidence that sickness behaviors, including weight loss and anxiety, are mediated through the effects of proinflammatory cytokines, such as IL-1, TNFα, and IL-6.7,35 The association between TLR4 and weight loss or anxiety may be mediated by cytokines. Thirdly, an increase in the content of TLR-specific mRNA may not, as a necessary consequence, cause an increase in TLR-specific surface expression of proteins. The dissociation between the protein and mRNA for receptors was noted in patients with sepsis, and in some cell lines.36,37 Finally, the sample size of the current study was very small; thus, further analyses using larger sample sizes and healthy controls are needed to avoid type I error and confirm these results.
In conclusion, our report highlights the significant association between anxiety, body weight loss, and TLR4 mRNA levels in patients with MDD. These findings contribute to evidence on the important role of TLR4 in anxiety-related behavior. However, further investigations and carefully controlled studies are still needed, to provide more evidence on the mechanism of TLR4-associated sickness behavior in patients with MDD.
Acknowledgment
This study was supported by grants CMRPG-8D1361 from Kaohsiung Chang Gung Memorial Hospital, Taiwan.
Disclosure
The authors report no conflicts of interest in this work.
References
De La Garza R 2nd. Endotoxin- or pro-inflammatory cytokine-induced sickness behavior as an animal model of depression: focus on anhedonia. Neurosci Biobehav Rev. 2005;29(4–5):761–770. | ||
Dowlati Y, Herrmann N, Swardfager W, et al. A meta-analysis of cytokines in major depression. Biol Psychiatry. 2010;67(5):446–457. | ||
Howren MB, Lamkin DM, Suls J. Associations of depression with C-reactive protein, IL-1, and IL-6: a meta-analysis. Psychosom Med. 2009;71(2):171–186. | ||
Myint AM, Schwarz MJ, Muller N. The role of the kynurenine metabolism in major depression. J Neural Transm. 2012;119(2):245–251. | ||
Capuron L, Miller AH. Immune system to brain signaling: neuropsychopharmacological implications. Pharmacol Ther. 2011;130(2):226–238. | ||
Miller AH, Maletic V, Raison CL. Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol Psychiatry. 2009;65(9):732–741. | ||
Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. 2008;9(1):46–56. | ||
Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4(7):499–511. | ||
Piccinini AM, Midwood KS. DAMPening inflammation by modulating TLR signalling. Mediators Inflamm. 2010;2010. pii:672395. | ||
Kawai T, Akira S. TLR signaling. Semin Immunol. 2007;19(1):24–32. | ||
Hung YY, Kang HY, Huang KW, Huang TL. Association between toll-like receptors expression and major depressive disorder. Psychiatry Res. 2014;220(1–2):283–286. | ||
Chang SH, Chiang SY, Chiu CC, et al. Expression of anti-cardiolipin antibodies and inflammatory associated factors in patients with schizophrenia. Psychiatry Res. 2011;187(3):341–346. | ||
McKernan DP, Dennison U, Gaszner G, Cryan JF, Dinan TG. Enhanced peripheral toll-like receptor responses in psychosis: further evidence of a pro-inflammatory phenotype. Transl Psychiatry. 2011;1:e36. | ||
Hajebrahimi B, Bagheri M, Hassanshahi G, et al. The adapter proteins of TLRs, TRIF and MYD88, are upregulated in depressed individuals. Int J Psychiatry Clin Pract. 2014;18(1):41–44. | ||
Oliveira J, Busson M, Etain B, et al. Polymorphism of Toll-like receptor 4 gene in bipolar disorder. J Affect Disord. 2014;152–154:395–402. | ||
Caso JR, Pradillo JM, Hurtado O, et al. Toll-like receptor 4 is involved in subacute stress-induced neuroinflammation and in the worsening of experimental stroke. Stroke. 2008;39(4):1314–1320. | ||
Zacharowski K, Zacharowski PA, Koch A, et al. Toll-like receptor 4 plays a crucial role in the immune-adrenal response to systemic inflammatory response syndrome. Proc Natl Acad Sci U S A. 2006;103(16):6392–6397. | ||
Maes M. Depression is an inflammatory disease, but cell-mediated immune activation is the key component of depression. Prog Neuropsychopharmacol Biol Psychiatry. 2011;35(3):664–675. | ||
Garate I, Garcia-Bueno B, Madrigal JL, et al. Origin and consequences of brain Toll-like receptor 4 pathway stimulation in an experimental model of depression. J Neuroinflamm. 2011;8:151. | ||
Keri S, Szabo C, Kelemen O. Expression of Toll-Like Receptors in peripheral blood mononuclear cells and response to cognitive-behavioral therapy in major depressive disorder. Brain Behav Immunity. 2014;40:235–243. | ||
Hamilton M. A rating scale for depression. J Neurol Neurosurg Psychiatry. 1960;23:56–62. | ||
López J, Fernández-Pisonero I, Dueñas AI, et al. Viral and bacterial patterns induce TLR-mediated sustained inflammation and calcification in aortic valve interstitial cells. Int J Cardiol. 2012;158(1):18–25. | ||
Stojanovic-Tasic M, Grgurevic A, Cvetkovic J, Grgurevic U, Trajkovic G. Association between somatic diseases and symptoms of depression and anxiety among Belgrade University students. Med Glas (Zenica). 2014;11(2):373–378. | ||
Maes M, Kubera M, Leunis JC. The gut-brain barrier in major depression: intestinal mucosal dysfunction with an increased translocation of LPS from gram negative enterobacteria (leaky gut) plays a role in the inflammatory pathophysiology of depression. Neuroendocrinol Lett. 2008;29(1):117–124. | ||
Bailey MT. Influence of stressor-induced nervous system activation on the intestinal microbiota and the importance for immunomodulation. Adv Exp Med Biol. 2014;817:255–276. | ||
Fetissov SO, Dechelotte P. The new link between gut-brain axis and neuropsychiatric disorders. Curr Opin Clin Nutr Metab Care. 2011;14(5):477–482. | ||
Pascual M, Balino P, Aragon CM, Guerri C. Cytokines and chemokines as biomarkers of ethanol-induced neuroinflammation and anxiety-related behavior: role of TLR4 and TLR2. Neuropharmacology. 2015;89:352–359. | ||
Bassi GS, Kanashiro A, Santin FM, et al. Lipopolysaccharide-induced sickness behaviour evaluated in different models of anxiety and innate fear in rats. Basic Clin Pharmacol Toxicol. 2012;110(4):359–369. | ||
Wu TY, Liu L, Zhang W, et al. High-mobility group box-1 was released actively and involved in LPS induced depressive-like behavior. J Psychiatr Res. 2015;64:99–106. | ||
Joshi JC, Ray A, Gulati K. Effects of morphine on stress induced anxiety in rats: role of nitric oxide and Hsp70. Physiol Behav. 2015;139:393–396. | ||
Steiner J, Bielau H, Brisch R, et al. Immunological aspects in the neurobiology of suicide: elevated microglial density in schizophrenia and depression is associated with suicide. J Psychiatr Res. 2008;42(2):151–157. | ||
Pandey GN, Rizavi HS, Ren X, Bhaumik R, Dwivedi Y. Toll-like receptors in the depressed and suicide brain. J Psychiatr Res. 2014;53:62–68. | ||
Schneider B, Philipp M, Muller MJ. Psychopathological predictors of suicide in patients with major depression during a 5-year follow-up. Eur Psychiatry. 2001;16(5):283–288. | ||
Kronfol Z, Nasrallah HA, Chapman S, House JD. Depression, cortisol metabolism and lymphocytopenia. J Affect Disord. 1985;9(2):169–173. | ||
Maes M, Berk M, Goehler L, et al. Depression and sickness behavior are Janus-faced responses to shared inflammatory pathways. BMC Med. 2012;10:66. | ||
Cohen J. The immunopathogenesis of sepsis. Nature. 2002;420(6917):885–891. | ||
Erdinest N, Aviel G, Moallem E, et al. Expression and activation of toll-like receptor 3 and toll-like receptor 4 on human corneal epithelial and conjunctival fibroblasts. J Inflamm. 2014;11(1):3. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.