Back to Journals » International Journal of General Medicine » Volume 17
Association Between the miR-100 rs1834306 A>G Polymorphism and Susceptibility to Venous Malformation
Authors Wu G, Lin X ![]() , Jiang H, Liu Z
, Jiang H, Liu Z ![]()
Received 26 September 2023
Accepted for publication 3 February 2024
Published 9 February 2024 Volume 2024:17 Pages 509—515
DOI https://doi.org/10.2147/IJGM.S441542
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Woon-Man Kung
Guitao Wu, Xi Lin, Hua Jiang, Zhenyin Liu
Department of Interventional Radiology and Vascular Anomalies, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou, Guangdong, 510623, People’s Republic of China
Correspondence: Zhenyin Liu, Department of Interventional Radiology and Vascular Anomalies, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, 9 Jinsui Road, Guangzhou, Guangdong, 510623, People’s Republic of China, Email [email protected]
Background: Venous malformation is related to genes and results in functional and morphologic anomalies. Genetic variations affecting the development of vessel endothelial cells are unclear. Therefore, this study aimed to investigate the potential value of the miR-100 rs1834306 A>G polymorphism as a marker of susceptibility to venous malformation.
Methods: In this case–control study in southern Chinese children, we collected blood samples from 1158 controls and 1113 patients with venous malformation. TaqMan genotyping of miR-100 rs1834306 A>G was performed by real-time fluorescent quantitative polymerase chain reaction.
Results: Multivariate logistic regression analysis showed that there was no significant association between the presence of the miR-100 rs1834306 A>G polymorphism and susceptibility to venous malformation by evaluating the values of pooled odds ratios and 95% confidence intervals. Similarly, among different sites, rs1834306 A>G was also not associated with venous malformation.
Conclusion: Our results suggest that the miR-100 rs1834306 A>G polymorphism is not associated with susceptibility to venous malformation in southern Chinese children. These results need to be further confirmed by investigating a more diverse ethnic population of patients with venous malformations.
Keywords: vascular malformation, susceptibility, miR-100, polymorphism
Introduction
Vascular malformation refers to a group of congenital defects in vascular development that manifest as diffuse or local structural abnormalities later in life, and its etiology is unknown.1 Vascular malformation results in functional and morphologic anomalies of the affected vascular areas with consequent hemodynamic aberration.1 As a result, vascular malformation patients are symptomatic, although some lesions remain quiescent for many years.1,2 Recent studies have proven that genetic mutations are responsible for vascular pathogenesis in many patients.2 Vascular malformations include artery malformations, venous malformations (VMs), capillary malformations and lymphatic malformations.2,3 Venous malformation is the most common form of vascular malformation.4 VM in different areas results in different symptoms, such as pain and swelling when it occurs in the limbs and disfigurement when it occurs in the face.3 VM can be life threatening when it occurs in important organs such as the pharynx, larynx, brain and heart.3,5 The disease has negative effects on patients’ daily lives, and identification of the main molecules involved in VM is urgent for improving clinical outcomes.6 A growing number of genes and genetic pathways that play a role in endothelial cell proliferation have been confirmed to be related to the pathogenesis of VM.7–9 However, the exact mechanism underlying VM is not currently clear. Most VMs are caused by mutations in genes involved in one of the following three signaling pathways: PIK3CA, MAPK and G-protein-coupled receptor signaling.10 Tie2 mutations are now known to also be found in patients with sporadic venous malformations, and it is recognized that some patients also have mutations in PIK3CA or Akt, which are downstream molecules of Tie2.11
The phenotype of germline mutations is determined by penetrance and expressivity, and is influenced by epigenetic factors. Methylation of specific genes is significantly correlated with the occurrence of VMs.10,12 The phenotypic spectrum of somatic mutations is wide and depends on variant allele frequency, timing during embryogenesis, cell type(s) involved and type of mutation. It has been shown that in vascular endothelial cells, methyl-binding proteins bind to methylated promoters of endothelial nitric oxide synthase (eNOS) and vascular endothelial growth factor receptor 2 (VEGFR2).13 Ablation of methyl-binding proteins in these cells leads to activation of eNOS and VEGFR2 gene expression and stimulates the proangiogenetic signaling pathway.14 However, the role of epitranscriptomics in this pathogenesis requires further research.
Epitranscriptomics refers to RNA functional alterations or modifications in the transcriptome that do not change the ribonucleotide sequence.15 MicroRNAs (miRNAs) are a class of single-stranded, small molecule RNAs that are derived from noncoding regions of DNA.15 These miRNAs can act as oncogenes or tumor suppressors to regulate a variety of biological processes, such as development, inflammation, and tumorigenesis.16 miR-100 is a potent suppressor of endothelial adhesion molecule expression, resulting in attenuated leukocyte-endothelial interactions in vitro and in vivo.17 Leonhard et al18 reported that neovascularization during graft-versus-host disease is regulated by miR-100. Grundmann et al19 indicated that miR-100 has an antiangiogenic function and represses mTOR signaling in endothelial and vascular smooth muscle cells. Recent research has revealed that local miR-100 expression is inversely correlated with patient inflammatory cell levels.17,20 Polymorphisms in miRNAs play an important role in and have potential value for diagnosing patients early. Developing interventions that target these polymorphisms can reduce the development and susceptibility of disease. However, polymorphisms of miR-100 have not been investigated in VM. A polymorphism of miR-100 may affect the migration and proliferation of embryonic endothelial cells and may be an underlying cause of VM. Therefore, we aimed to investigate the relationship between the miR-100 rs1834306 A>G single nucleotide polymorphism (SNP) and the risk of VM in southern Chinese children in a case–control study.
Methods
Participants
Our study was prospectively planned and conducted as a case–control study. We included southern Chinese children with VM (1113 cases) and healthy controls (1158 cases) who were recruited from the Guangzhou Women and Children’s Medical Center, mainly between January 2015 and March 2021. Blood samples were collected from both children. The healthy controls were children who were matched geographically and ethnically with the children with VM and had no prior history of VM. This study was approved by the institutional review board of Guangzhou Women and Children’s Medical Center (Ethical approval No. 201943800). All patients were diagnosed with venous malformation via physical examination and clinical imaging diagnosis. Clinical information was collected from medical records, and all subjects or their guardians signed informed consent forms. Informed consent was obtained from all participants as per as the rules of the Helsinki Declaration.
Genotyping
On the basis of our previous criteria, in the present case‒control study, we isolated the potential rs1834306 A>G polymorphic locus of miR-100. Using a TIANamp Blood DNA Kit (DP348; TianGen Biotech Co., Ltd., Beijing, China), DNA was extracted from peripheral blood leukocytes from all participants. The DNA samples were diluted to 10 ng/μL in 96-well plate and stored at −20°C for later use. They were then added to a TaqMan Genotyping PCR PreMix (probe (FP211); TianGen Biotech Co., Ltd.) in 384-well plates. Real-time fluorescent quantitative polymerase chain reaction was used to genotype the miR-100 rs1834306 A>G polymorphism. At least 10% of samples were randomly selected for repeated genotyping, and the same results were obtained.
Statistical Analysis
Differences in demographic characteristics and distribution of genotype frequency between patients with VM and healthy controls were assessed by using the bilateral χ2 test. Hardy–Weinberg equilibrium was used to assess the genotype frequencies of the goodness-of-fit χ2 test in the healthy controls. Measurement data are presented as the mean ± standard deviation. Multivariate logistic regression was used to adjust for the effects of age and sex. The association between the miR-100 rs1834306 A>G polymorphism and susceptibility to VM was estimated by multivariate analysis. Logistic regression analysis using odds ratios (ORs) and 95% confidence intervals (CIs) was performed. Furthermore, stratified analyses were performed to calculate different subtypes of VM based on the VM site of the limbs. All statistical analyses and tests were performed using SAS software (version 9.4; SAS Institute, Cary, NC, USA). Statistical significance was set at P<0.05.
Results
Baseline Demographics
We studied 1113 southern Chinese children with VM (mean age: 68.98±37.08 months; 54.90% boys) and 1158 healthy controls (mean age: 22.22 ± 19.98 months; 56.82% boys).
Association of the miR-100 rs1834306 A>G with Susceptibility to VM
Using real-time fluorescent quantitative polymerase chain reaction, we successfully genotyped the miR-100 rs1834306 A>G polymorphism in 1113 patients and 1158 controls. The distribution of the genotype frequency of the miR-100 rs1834306 A>G was consistent with the Hardy–Weinberg equilibrium (P=0.120) in healthy controls. We found that the genotype results in patients with VM were not significantly different compared with those of the healthy controls. Therefore, none of the rs1834306 genotypes were associated with susceptibility to VM (Table 1).
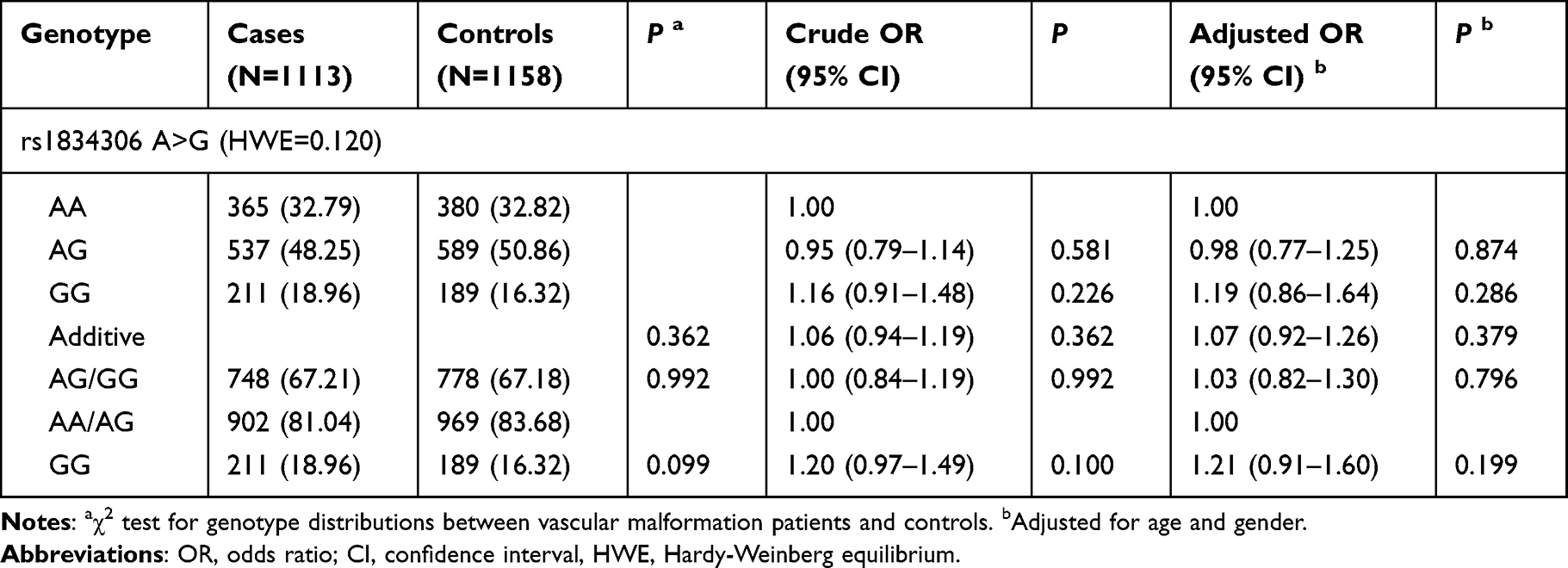 |
Table 1 Association Between miR-100 rs1834306 A>G Polymorphism and Vascular Malformation Susceptibility |
Stratification Analysis of the Risk Between the miR-100 rs1834306 A>G and Different VM Sites
According to different VM sites, VM was divided into 6 different subtypes, including head, neck, trunk, upper limbs, lower limbs and multiple venous malformations. After adjusting for age and sex, we found that there were no significant associations between the miR-100 rs1834306 A>G and the six VM subtypes (Table 2). This finding suggests that the miR-100 rs1834306 A>G is not associated with susceptibility to VM.
 |
Table 2 Stratification Analysis for the Association Between miR-100 rs1834306 A>G and Vascular Malformation Susceptibility (by Site) |
Discussion
Although genetic data from numerous studies suggest that genetic abnormalities trigger VM pathogenesis, the exact molecular pathways responsible for the development of these lesions remain to be elucidated.2,19,21 In our case–control study of southern Chinese children, we investigated the association between a miR-100 rs1834306 A>G and susceptibility to VM. To the best of our knowledge, the miR-100 rs1834306 A>G has not been examined in any previous studies on VM. Our study showed that the miR-100 rs1834306 A>G was not significantly associated with susceptibility to VM in southern Chinese children.
Genetic mutations affecting the proliferation, migration, adhesion, differentiation, and survival of endothelial cells and the integrity of the extracellular matrix are believed to play a role in the pathogenesis of VM.22 An increased number of genetic mutations have been discovered in vascular anomalies via targeted deep sequencing. For example, both noncomplicated cutaneous capillary malformation and capillary malformation associated with Sturge‒Weber syndrome were found to have the same GNAQ mutations.2,23–25 Similarly, mutations in AKT1, AKT2, and AKT3 have also been discovered in patients with megalencephaly and venous and capillary malformation.26,27 Many studies have investigated the potential functional genetic variation sites and their genetic susceptibilities to VM. Cottrell et al28 suggested that PIK3R1 variation underlies vascular malformations and overgrowth. Al-Olabi et al29 discovered multiple mosaic-activating variants in 4 genes, KRAS, NRAS, BRAF, and MAP2K1, of the RAS/MAPK pathway, which is commonly activated in cancer and responsible for germline RASopathies. Furthermore, PIK3CA mutations were also discovered in patients with congenital lipomatous overgrowth, vascular malformations, epidermal nevi and scoliosis/skeletal/spinal anomalies (CLOVES) syndrome, capillary vascular malformation of the lower lip (CLAPO) syndrome, capillary malformation with megalencephaly, Klippel-Trenaunay syndrome, or mucocutaneous venous malformations.30–32 These studies indicate that SNPs may affect the expression and function of target genes and may contribute to VM susceptibility. Investigating the effect of SNPs and genetic mutations on the pathogenesis of VM will contribute to a better understanding of the etiology of this disease.
A growing amount of research has shown that noncoding RNAs are potential markers of disease risk and prognosis in VMs.33,34 MiR-21 expression is markedly decreased in skin specimens from patients with VM compared with skin specimens from healthy individuals, and miR-21 positively regulates the expression of collagens in human umbilical vein endothelial cells (HUVECs) and shows a positive association with the TGF-β/Smad3 pathway in VM tissues.35,36 MiR-145 expression was positively correlated with TGF-β expression and perivascular α-SMA cell coverage in VM tissue samples.36 Moreover, miR-18a-5p was reported to regulate the activation of P53 signaling pathway constituents and consequently regulate proliferation, migration, invasion and angiogenesis.37
Nonetheless, whether miR-100 is involved in the pathogenesis of VM has yet to be determined. At present, several studies have concentrated on the effect of miR-100 on the vasculature. Yu et al38 discovered that miR-100, along with other miRNAs, exerts considerable influences on circulatory systems in cancellous bone by targeting cytokines and enzymes such as catalase, fibroblast growth factor and nerve growth factor. Furthermore, miR-100 plays a role in disrupting the formation of vessels by negatively regulating angiopoietin 2, which inhibits encapsulated tumor clusters.39 Moreover, overexpression of miR-100 attenuates endothelial cell dysfunction by targeting HIPK2 after hypoxia and reoxygenation treatment.20 Grundmann et al19 proved that miR-100 has an antiangiogenic function and represses mTOR signaling in endothelial and vascular smooth muscle cells. Collectively, these reports raised the question of whether miR-100 can directly regulate the migration or differentiation of endothelial cells in the pathogenesis of VM.
However, our results are not completely consistent with those of previous studies. Previous studies concentrated on the effect of miR-100 on rats or in vitro experiments, while our study used a population analysis. As a result of the possible differences in allele frequencies and SNP-mediated allelic imbalance among different populations, only the rs1834306 A>G from the miR-100 was selected for association analysis in the current study. Additional gene polymorphisms should be further investigated in future studies. Our research does have some limitations that should be mentioned. First, only one polymorphism in the miR-100 was selected for this study, and the combined effects of miR-100 rs1834306 A>G in combination with other polymorphisms in related genes on VM were not studied. Second, as a hospital-based retrospective investigation, a certain degree of admission bias, such as information gathering and population selection, is unavoidable. Third, lifestyle and environmental factors were not taken in account, and those factors may hide the real association between miR-100 and VM. Fourth, our conclusion cannot be speculated outward into other ethnicities, because all our subjects were recruited from the Han population. Fifth, more functional experiments are required to confirm the outcome of this investigation.
In summary, the miR-100 rs1834306 A>G is not associated with the risk of VM in southern Chinese children. The role of other SNPs in the pathogenesis of VM needs to be further examined. Future research should study target genetic variants identified in VM to identify their role and clinical effect in VM pathogenesis. These investigations may ultimately lead to better screening programs and prevention of VM.
Abbreviations
VM, venous malformation; PIK3CA, phosphatidylinositol 3-kinase catalytic subunit alpha; MAPK, Mitogen-activated protein kinase; eNOS, endothelial nitric oxide synthase; VEGFR2, vessel endothelial growth factor receptor 2; miRNA, microRNA; OR, odd ratio; CI, confidence interval; SNP, single nucleotide polymorphism; GNAQ, G protein guanine nucleotide binding protein alpha subunit q; HUVECs, human umbilical vein endothelial cells; HIPK2, Homeodomain-Interacting Protein Kinase 2.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The study received the approval of the Ethics Committee of the Children’s Medical Center and was conducted according to the principles expressed in the Declaration of Helsinki of 1975, as revised in 2001. Informed consent was acquired from each participant and their parents included in the study.
Acknowledgments
We thank the Clinical Biological Resource Bank of Guangzhou Women and Children’s Medical Center for providing the clinical sample.
Author Contributions
Zhenyin Liu conceived and designed the work, Guitao Wu conducted the statistical analysis and prepared the Tables, Xi Lin and Guitao Wu conducted the experiments and wrote the manuscript, Xi Lin and Hua Jiang collected the samples and clinical data. All authors reviewed and approved the final version of the manuscript. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was funded by Guangzhou Women and Children’s Medical Center Hospital Fund (No: IP-2019-015).
Disclosure
The authors declare no competing interests with this study.
References
1. Carqueja IM, Sousa J, Mansilha A. Vascular malformations: classification, diagnosis and treatment. Int Angiol. 2018;37(2):127–142. doi:10.23736/S0392-9590.18.03961-5
2. Duffy K. Genetics and syndromes associated with vascular malformations. Pediatr Clin North Am. 2010;57(5):1111–1120. doi:10.1016/j.pcl.2010.07.001
3. Huang JT, Liang MG. Vascular malformations. Pediatr Clin North Am. 2010;57(5):1091–1110. doi:10.1016/j.pcl.2010.08.003
4. Lee BB. Venous malformation and haemangioma: differential diagnosis, diagnosis, natural history and consequences. Phlebology. 2013;28(Suppl 1):176–187. doi:10.1177/0268355513475960
5. Redondo P, Aguado L, Martinez-Cuesta A. Diagnosis and management of extensive vascular malformations of the lower limb: part I. Clinical diagnosis. J Am Acad Dermatol. 2011;65(5):893–906; quiz 907–898. doi:10.1016/j.jaad.2010.12.047
6. Kim H, Joh J, Labropoulos N. Characteristics, clinical presentation, and treatment outcomes of venous malformation in the extremities. J Vasc Surg Venous Lymphat Disord. 2022;10(1):152–158. doi:10.1016/j.jvsv.2021.05.011
7. Wassef M, Blei F, Adams D, et al. Vascular anomalies classification: recommendations from the international society for the study of vascular anomalies. Pediatrics. 2015;136(1):e203–214. doi:10.1542/peds.2014-3673
8. Limaye N, Wouters V, Uebelhoer M, et al. Somatic mutations in angiopoietin receptor gene TEK cause solitary and multiple sporadic venous malformations. Nat Genet. 2009;41(1):118–124. doi:10.1038/ng.272
9. Nguyen HL, Boon LM, Vikkula M. Vascular anomalies caused by abnormal signaling within endothelial cells: targets for novel therapies. Semin Intervent Radiol. 2017;34(3):233–238. doi:10.1055/s-0037-1604296
10. Hoeger PH. Genes and phenotypes in vascular malformations. Clin Exp Dermatol. 2021;46(3):495–502. doi:10.1111/ced.14513
11. Kunimoto K, Yamamoto Y, Jinnin M. ISSVA classification of vascular anomalies and molecular biology. Int J Mol Sci. 2022;23(4):2358. doi:10.3390/ijms23042358
12. Chen X, Liu Y, Zhou S, et al. Methylation of the CDKN2A gene increases the risk of brain arteriovenous malformations. J Mol Neurosci. 2019;69(2):316–323. doi:10.1007/s12031-019-01360-6
13. Rao X, Zhong J, Zhang S, et al. Loss of methyl-CpG-binding domain protein 2 enhances endothelial angiogenesis and protects mice against hind-limb ischemic injury. Circulation. 2011;123(25):2964–2974. doi:10.1161/CIRCULATIONAHA.110.966408
14. Thomas JM, Surendran S, Abraham M, Rajavelu A, Kartha CC. Genetic and epigenetic mechanisms in the development of arteriovenous malformations in the brain. Clin Clin Epigenet. 2016;8(1):78. doi:10.1186/s13148-016-0248-8
15. Piletic K, Kunej T. MicroRNA epigenetic signatures in human disease. Arch Toxicol. 2016;90(10):2405–2419. doi:10.1007/s00204-016-1815-7
16. Mohr AM, Mott JL. Overview of microRNA biology. Semin Liver Dis. 2015;35(1):3–11. doi:10.1055/s-0034-1397344
17. Pankratz F, Hohnloser C, Bemtgen X, et al. MicroRNA-100 suppresses chronic vascular inflammation by stimulation of endothelial autophagy. Circ Res. 2018;122(3):417–432. doi:10.1161/CIRCRESAHA.117.311428
18. Leonhardt F, Grundmann S, Behe M, et al. Inflammatory neovascularization during graft-versus-host disease is regulated by alphav integrin and miR-100. Blood. 2013;121(17):3307–3318. doi:10.1182/blood-2012-07-442665
19. Grundmann S, Hans FP, Kinniry S, et al. MicroRNA-100 regulates neovascularization by suppression of mammalian target of rapamycin in endothelial and vascular smooth muscle cells. Circulation. 2011;123(9):999–1009. doi:10.1161/CIRCULATIONAHA.110.000323
20. Zheng H, Sun Y, Shu X, Gao Q, Chen X. Overexpression of microRNA-100-5p attenuates the endothelial cell dysfunction by targeting HIPK2 under hypoxia and reoxygenation treatment. J Mol Histol. 2021;52(5):1115–1125. doi:10.1007/s10735-021-10002-4
21. Vikkula M, Boon LM, Carraway KL, et al. Vascular dysmorphogenesis caused by an activating mutation in the receptor tyrosine kinase TIE2. Cell. 1996;87(7):1181–1190. doi:10.1016/S0092-8674(00)81814-0
22. Snellings DA, Hong CC, Ren AA, et al. Cerebral cavernous malformation: from mechanism to therapy. Circ Res. 2021;129(1):195–215. doi:10.1161/CIRCRESAHA.121.318174
23. Wetzel-Strong SE, Detter MR, Marchuk DA. The pathobiology of vascular malformations: insights from human and model organism genetics. J Pathol. 2017;241(2):281–293. doi:10.1002/path.4844
24. Huang L, Couto JA, Pinto A, et al. Somatic GNAQ mutation is enriched in brain endothelial cells in Sturge-Weber syndrome. Pediatr Neurol. 2017;67:59–63. doi:10.1016/j.pediatrneurol.2016.10.010
25. Sundaram SK, Michelhaugh SK, Klinger NV, et al. GNAQ mutation in the venous vascular malformation and underlying brain tissue in Sturge-Weber syndrome. Neuropediatrics. 2017;48(5):385–389. doi:10.1055/s-0037-1603515
26. Rodriguez-Laguna L, Ibanez K, Gordo G, et al. CLAPO syndrome: identification of somatic activating PIK3CA mutations and delineation of the natural history and phenotype. Genet Med. 2018;20(8):882–889. doi:10.1038/gim.2017.200
27. Castillo SD, Vanhaesebroeck B, Sebire NJ. Phosphoinositide 3-kinase: a new kid on the block in vascular anomalies. J Pathol. 2016;240(4):387–396. doi:10.1002/path.4802
28. Cottrell CE, Bender NR, Zimmermann MT, et al. Somatic PIK3R1 variation as a cause of vascular malformations and overgrowth. Genet Med. 2021;23(10):1882–1888. doi:10.1038/s41436-021-01211-z
29. Al-Olabi L, Polubothu S, Dowsett K, et al. Mosaic RAS/MAPK variants cause sporadic vascular malformations which respond to targeted therapy. J Clin Invest. 2018;128(4):1496–1508. doi:10.1172/JCI98589
30. Greene AK, Goss JA. Vascular anomalies: from a clinicohistologic to a genetic framework. Plast Reconstr Surg. 2018;141(5):709e–717e. doi:10.1097/PRS.0000000000004294
31. Queisser A, Boon LM, Vikkula M. Etiology and Genetics of Congenital Vascular Lesions. Otolaryngol Clin North Am. 2018;51(1):41–53. doi:10.1016/j.otc.2017.09.006
32. Kangas J, Natynki M, Eklund L. Development of molecular therapies for venous malformations. Basic Clin Pharmacol Toxicol. 2018;123(Suppl 5):6–19. doi:10.1111/bcpt.13027
33. Erkinova SA, Sokolova EA, Orlov KY, et al. Angiopoietin-Like Proteins 4 (ANGPTL4) gene polymorphisms and risk of brain arteriovenous malformation. J Stroke Cerebrovasc Dis. 2018;27(4):908–913. doi:10.1016/j.jstrokecerebrovasdis.2017.10.033
34. Si Y, Huang J, Li X, et al. AKT/FOXO1 axis links cross-talking of endothelial cell and pericyte in TIE2-mutated venous malformations. Cell Commun Signal. 2020;18(1):139. doi:10.1186/s12964-020-00606-w
35. Xia HF, Ren JG, Zhu JY, et al. Downregulation of miR-145 in venous malformations: its association with disorganized vessels and sclerotherapy. Eur J Pharm Sci. 2017;100:126–131. doi:10.1016/j.ejps.2017.01.019
36. Zhu J, Tang Z, Ren J, et al. Downregulation of microRNA-21 contributes to decreased collagen expression in venous malformations via transforming growth factor-beta/Smad3/microRNA-21 signaling feedback loop. J Vasc Surg Venous Lymphat Disord. 2022;10(2):469–481 e462. doi:10.1016/j.jvsv.2021.08.020
37. Zhang L, Wang D, Wang Z, et al. MiR-18a-5p acts as a novel serum biomarker for venous malformation and promotes angiogenesis by regulating the thrombospondin-1/P53 signaling axis. Am J Transl Res. 2021;13(10):11271–11286.
38. Yu Q, Guo W, Cheng L, Lu Y, Li P. 甾体类激素影响骨内微血管内皮细胞表达谱与转录组的初探 [Preliminary study of impact of steroids on expression profile and transcriptome of bone microvascular endothelial cells]. Zhonghua Yi Xue Za Zhi. 2014;94(48):3817–3820. Chinese.
39. Zhou HC, Fang JH, Shang LR, et al. MicroRNAs miR-125b and miR-100 suppress metastasis of hepatocellular carcinoma by disrupting the formation of vessels that encapsulate tumour clusters. J Pathol. 2016;240(4):450–460. doi:10.1002/path.4804
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Genetic Variations of AKT1 are Associated with Risk Screening for Non-Alcoholic Fatty Liver Disease

Ding Y, Tang Z, Zhang R, Zhang M, Guan Q, Zhang L, Wang H, Chen Y, Zhang W, Wang J
Risk Management and Healthcare Policy 2023, 16:1365-1376
Published Date: 26 July 2023