Back to Journals » Neuropsychiatric Disease and Treatment » Volume 19
Association Between Single Nucleotide Polymorphisms Within lncRNA NONHSAT102891 and Depression Susceptibility in a Chinese Population
Authors Liang P, Sun Y, Li Y, Liang Y ![]()
Received 14 October 2022
Accepted for publication 13 January 2023
Published 2 February 2023 Volume 2023:19 Pages 293—302
DOI https://doi.org/10.2147/NDT.S393498
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yu-Ping Ning
Peng Liang, Yingjie Sun, Yue Li, Yundan Liang
Department of Pathology and Pathophysiology, School of Basic Medical Sciences, Chengdu Medical College, Chengdu, People’s Republic of China
Correspondence: Yundan Liang, Department of Pathology and Pathophysiology, School of Basic Medical Sciences, Chengdu Medical College, Chengdu, Sichuan, 610500, People’s Republic of China, Tel/Fax +86-28-62739308, Email [email protected]
Background: Depression is among the most common psychiatric disorders, and is a leading cause of the global disease burden. Its pathophysiological mechanism is unclear, which limits the development of therapeutic strategies. Long non-coding RNA (lncRNA) single nucleotide polymorphisms (SNPS) may be related. In this study, we aimed to determine the effects of the rs2242385, rs155979, rs3762983, and rs3762984 polymorphisms in the lncRNA NONHSAT102891 on depression susceptibility in a Chinese population.
Methods: We conducted a case-control study in a cohort of 480 patients with depression and 329 healthy controls, and performed genotyping by gene sequencing ii.
Results: The rs155979 GC genotype was significantly associated with increased risk of depression compared with healthy controls. Stratified analysis showed a 2.08-fold increased risk of suicide in patients with rs155979 GC or GG genotype. The rs2242385, rs3762983, and rs3762984 polymorphisms were not significantly associated with the risk of depression. Haploid analysis showed linkage disequilibrium between rs155979, rs3762983, and rs3762984, and the CCG haplotype reduced the risk of depression.
Limitations: The study sample was relatively small, and was restricted to patients from central and southern China. Further, only peripheral blood was used for DNA extraction.
Conclusion: The rs155979 polymorphism may be associated with the occurrence of depression in the Chinese population. However, further studies are needed to verify the reliability of our results in large populations and different ethnic groups.
Keywords: depression, SNP, LncRNA
Introduction
Depression is a common recurrent mental disorder, with a lifetime risk is nearly 20%.1 It is characterized by persistent sadness and a lack of interest or pleasure in previously beneficial or enjoyable activities.2 Patients with depression have about 3 times as many suicidal ideations in their lifetime as healthy people.3 In 2008, the WHO listed major depression as the third leading cause of the global disease burden; it will likely be the first by 2030.4 Despite significant advances in our understanding of the pathophysiological mechanism of depression, with many biological, genetic, environmental, psychological, and other factors being linked to its development,1 the exact mechanism underlying this disease remains unclear. Further research on the underlying mechanism is essential for the development to effective therapeutic strategies.
Long non-coding RNAs (lncRNAs) are groups of non-coding RNAs >200 nucleotides in length that are transcribed but not translated into proteins.5 Their functions are mainly related to DNA methylation, gene silencing or activation, transcription and post-transcriptional regulation, and epigenetics.6 lncRNAs are involved in many diseases, including depression and suicidal behavior.7 The lncRNA NONHSAT102891 in peripheral blood molecular cells of patients with depression was significantly down-regulated compared with normal controls.8 LncRNA NONHSAT102891 is located on chromosome 5 (CHR5:95768987–95770845).
Single nucleotide polymorphisms (SNPs) refer to the DNA sequence polymorphisms caused by a single base difference on the DNA sequence of different individuals at the genome level, which is the most major manifestation of genetic differences between individuals or populations. It accounts for more than 90% of the genetic polymorphisms in the human genome. Variation of any base in the human genome can occur in coding, non-coding and gene spacing regions. Many psychiatric disorders have been linked to SNPS. Milad Heidari Nia et al found that intron GABRB2rs1816072 and rs12187676 polymorphisms were associated with schizophrenia risk in an Iranian population sample.9 Saman Sargazi et al found that rs4705342T/C and rs4705343T/C polymorphisms in the miR-143 promoter region were associated with schizophrenia susceptibility in Iranian population samples.10 Saman Sargazi et al showed that HOTAIR polymorphisms of long-stranded non-coding RNA are associated with susceptibility to bipolar disorder.11 Mohammad Ali Mokhtari et al showed that genetic polymorphisms in miR-137 and its target genes TCF4 and CACNA1C lead to the risk of bipolar disorder.12
lncRNA expression and function are affected by single nucleotide polymorphism and have a correlation with the pathogenesis of mental diseases. To date, some studies have explored the relationship between SNPs in lncRNA and susceptibility to depression. ArezouSayad et al reported that SNP of rs1899663 in lncRNA HOX transcription antisense intergenic RNA (HOTAIR) was associated with depression susceptibility in an Iranian population.13 Weipeng Liu et al reported that the expression of lncRNA (LINC01360) was affected by rs70959274 Alu polymorphism and was related to the pathogenesis of depression.14 Yanni Zeng et al showed that the brain-specific long non-coding RNA RP1-269M15.3 in the basal ganglia of the nucleus accumbens was significantly regulated by Major Depressive Disorder (MDD)-related SNP across the 24 KB region of the TOX2 gene.15
In this hospital-based case-control study, we genotyped SNPS of rs2242385, rs155979, rs3762983 and rs3762984 in lncRNA NONHSAT102891 blood DNA samples from 480 patients with depression and 329 controls from the general population. To examine its association with depression susceptibility in the Chinese population.
Materials and Methods
Subjects
Between March 2018 and December 2019, 480 patients with depression who met the diagnostic criteria of The Diagnostic and Statistical Manual of Mental Disorders (DSM-IV) were recruited at Jining Psychiatric Hospital and Yunnan Mental Health Center.329 healthy controls were recruited from individuals who came to the hospital for health checkups during the same period. The study was approved by the ethics committee of Chengdu Medical College. After being informed of the purpose of the study, all subjects signed a complete written consent form. Eligible patients had a total score of ≥20 on the 24-item Hamilton Depression Scale (HAMD-24) at screening and baseline visit. Exclusion criteria including lack of signed consent to participate in the evaluation, thyroid dysfunction, history of nervous system diseases, neurodegenerative diseases (such as Parkinson’s disease, epilepsy, Alzheimer’s disease), diagnoses of other mental disorders (such as schizophrenia, divided emotional disorder, or bipolar disorder), serious physical disease (acute or chronic), drug or substance abuse, cognitive impairment, and breastfeeding or pregnancy. We also included cardiovascular disease, kidney or liver disease, obesity, abnormal body mass index (BMI), autoimmune disease, infectious disease, and uncontrolled endocrine disease as additional exclusion criteria for participants in this study. The clinical data of patients, including age, sex, pulse rate, age at onset, suicide attempt, whether the patient was a first-time patient, and family history, were obtained from medical records. Healthy controls had the same exclusion criteria as did the patients. The mean age of healthy controls (105 males and 224 females) was 44.0 ± 16.9 years (Table 1). Frequency matching was performed between the control group and the cases according to gender, age, and place of residence. Quanto software 1.2.3 (University of Southern California, Los Angeles, CA, USA) was used to calculate the power of sample size. When the RG value was set to 1.6, the genetic power of the four SNPS was greater than 80% under a dominant model.16
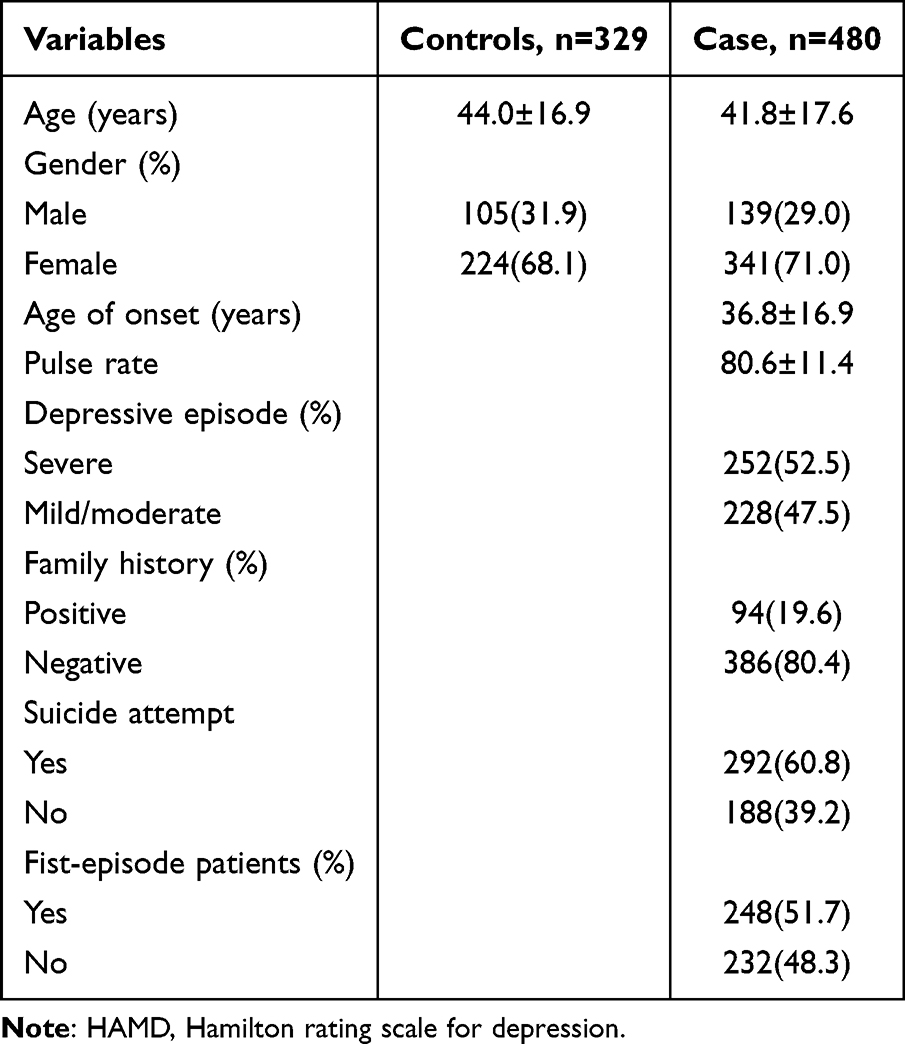 |
Table 1 Characteristics of the Study Population |
SNPs Selection
UCSC was used to search for SNPs associated with depression in the 2kb region downstream of lncRNA NONHSAT102891 transcription start site, and loci with secondary allelic frequency greater than 10% in Asian population were selected as follows: rs2242385, rs155979, rs3762983 and rs3762984. (Figure 1)
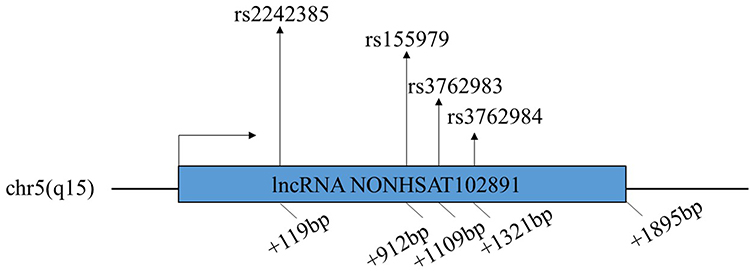 |
Figure 1 Schematic diagram of rs2242385, rs155979, rs3762983, and rs3762984 on the lncRNA NONHSAT102891. |
Genotyping
Human blood collection: 2–3 mL whole blood samples from all subjects were collected using an EDTA anticoagulant tube and stored in −20°C freezer.
DNA extraction: A whole blood genomic DNA rapid extraction kit (Sangon Bioengineering (Shanghai) Co., LTD.) was used for DNA extraction of blood samples, according to manufacturer’s instructions.
Multiple amplification and high-throughput sequencing: A primer pool containing four SNPs was designed and synthesized, the SNP target sequences were amplified by two-step PCR, and the Illumina-compatible library was prepared. The first PCR system was as follows: DNA template (10 ng/μL) 2 μL; Upstream primer pool (10 μM) 1 μL; Downstream primer pool (10 μM) 1 μL; 2× PCR Ready Mix 15 μL (total volume 25μL; Kapa HiFi Ready Mix) After the reaction system was prepared, the following reaction cycles were performed on a PCR apparatus (Bio-RAD, T100TM): pre-denaturation at 98°C for 3 min, followed by eight cycles of denaturation at 98°C for 30 seconds, annealing at 50°C for 30 seconds, and extension at 72°C for 30 seconds. This was followed by 25 cycles of denaturation at 98°C for 30 seconds, annealing at 66°C for 30 seconds, and extension at 72°C for 30 seconds. The final 72°C extension was 5 minutes. After the reaction was complete, the temperature was 4°C. The PCR products were detected by 1% agarose gel electrophoresis to determine the correct size of the products, and the PCR products were purified and recovered using AMPure XP magnetic beads. The first PCR product was used as the template to perform the second PCR reaction to obtain a library with molecular tags. The reaction system was as follows: DNA template (10 ng/μL) 2 μL, universal P7 primer (including molecular label, 10 μM) 1 μL; Universal P5 primer (10 μM) 1 μL; PCR Ready Mix 15 μL (total volume 30 μL) After the reaction system was formulated, the following PCR program was performed: pre-denaturation at 98°C for 5 min, followed by five cycles of denaturation at 94°C for 30 seconds, annealing at 55°C for 20 seconds, extension at 72°C for 30 SEC, and finally extension at 72°C for 5 min. Temperature was maintained at 4°C after completion. The final PCR product was purified and recovered using AMPure XP magnetic beads. The PCR products were mixed in equal quantities and sequenced using HiSeqXTen (Illumina, San Diego, CA).
Data quality control and genotyping analysis: Data quality control for off-machine data was performed in two steps: 1) Cutadapt (V1.2.1) was used to remove any part of the sequence containing the connector sequence; 2) Prinseq-Lite (V0.20.3) software was used for quality control of the rest of the sequence, in order from the 3’ end to the 5’ end of the sequence. Bases with a quality threshold below 20 were deleted. The remaining sequences were considered qualified for quality control. BWA (V0.7.13-R1126) software was then used to compare the qualified sequences to the reference genome with the default parameters. Based on the comparison results, samTools software (version 0.1.18) was used to calculate the genotype results of the target loci. Finally, Annovar (2018–04-16) software was used for gene annotation of mutation sites.
Statistical Analysis
All statistical analyses were performed using SPSS 25.0. Software packages (Chicago, IL, USA) and GraphPad Prism 5.0 (GraphPad Software, San Diego, CA, USA). The genotype frequencies of rs2242385, rs155979, rs3762983, and rs3762984 were obtained by direct counting. The Hardy-Weinberg equilibrium was tested using the χ2 test. The distribution of rs2242385, rs155979, rs3762983, and rs3762984 genotypes in cases and controls was tested using the X test, and the association between the four polymorphisms and the risk of depression was assessed by odds ratio (OR) and 95% confidence interval (Cls). P < 0.05 was considered statistically significant.
Result
Genotype frequencies of the two polymorphisms in the control and depression groups are shown in Table 2. The control group did not deviate from the Hardy-Weinberg equilibrium (rs2242385:P = 0.10; rs155979: P = 0.33; rs3762983: P = 0.74; rs3762984: P = 0.06). Our eQTL results showed that rs2242385, rs155979, rs3762983 and rs3762984 polymorphic loci had no effect on the expression of LncRNA NONHSAT102891. There was a significant difference in the distribution of rs155979 GC genotype between the depression and control groups (OR = 1.53, 95% CI of 1.09–2.15, P = 0.01). The frequency of rs155979 was significantly different between the patient and control groups (OR = 1.49, 95% CI of 1.08–2.05, P = 0.01), and the G allele frequency of rs155979 in the depression group was significantly higher than that in the control group (OR = 1.28,95% CI of 1.00–1.64, P = 0.05). No significant differences were observed between the case and control groups for the remaining three polymorphisms.
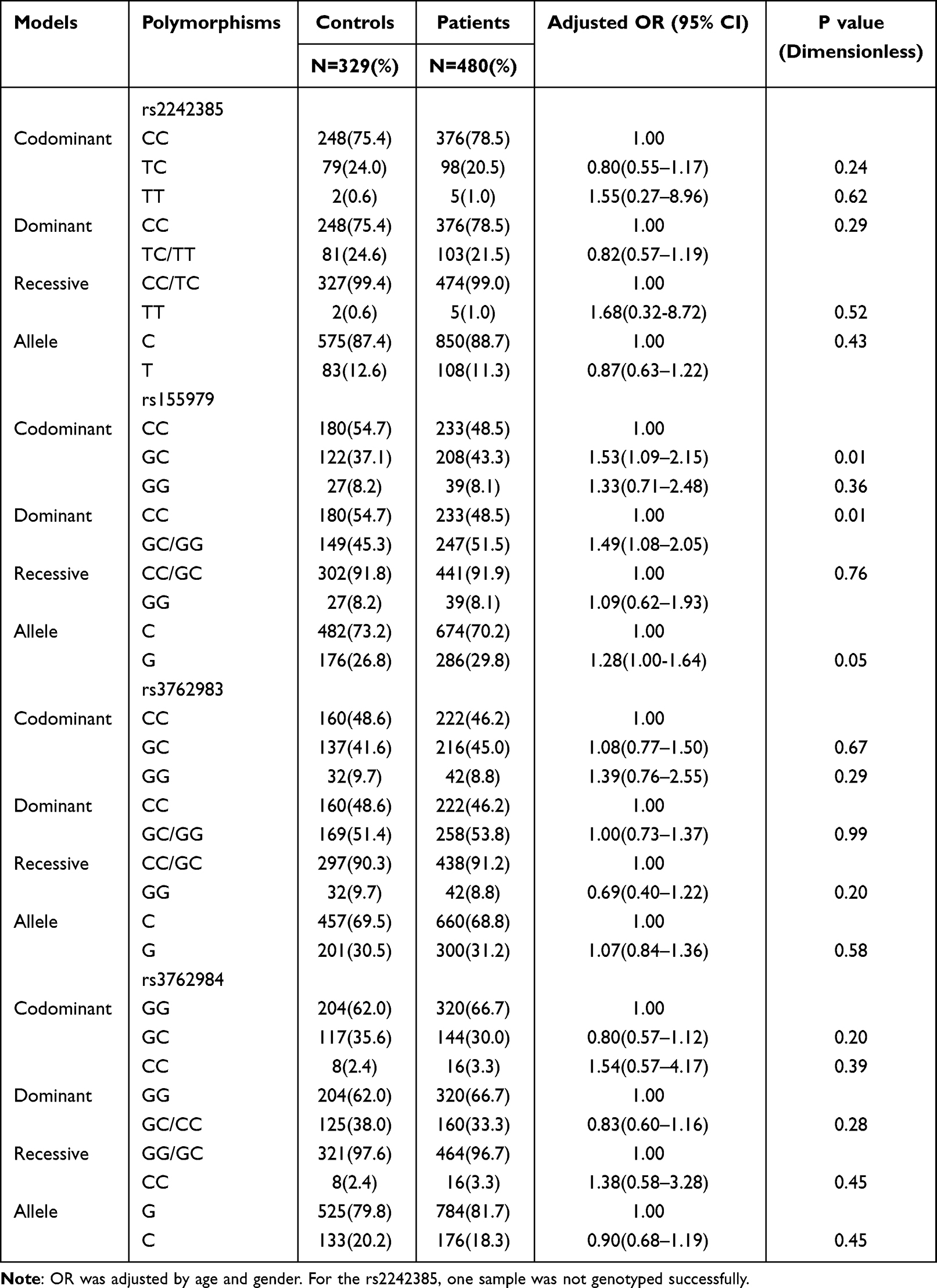 |
Table 2 Association of the rs2242385, rs155979, rs3762983 and rs3762984 Polymorphisms with Depression Risk |
Stratified analysis according to depressive episode (severe vs mild/moderate), attempted suicide (yes vs no), first-episode patients (yes vs no) showed that rs155979 polymorphism was different in patients with suicide attempt. The frequency of GC and GG genotypes of rs155979 in patients with attempted suicide was significantly higher than that in the remaining patients (C/C vs G/C-G/G: OR = 2.08, 95% CI of 1.18–3.65, P = 0.01; Table 3). However, no significant correlation was found between rs155979 polymorphism and the other two variables (P>0.05). rs2242385, rs3762983, and rs3762984 polymorphisms were not significantly correlated with these three variables (P>0.05; Table 3).
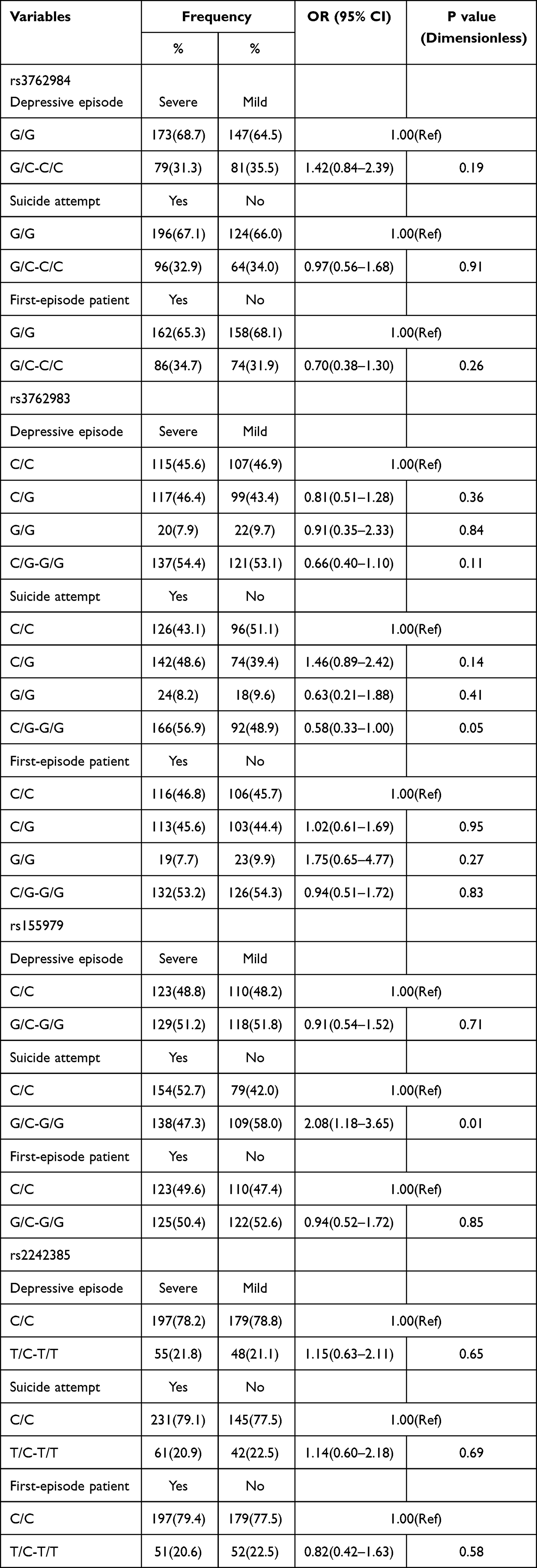 |
Table 3 Stratified Analyses of the rs2242385, rs155979, rs3762983 and rs3762984 Polymorphisms with Depression Risk |
Note: OR was adjusted by age and gender. For the rs2242385, one sample was not genotyped successfully.
Haploid analysis of rs2242385, rs155979, rs3762983, and rs3762984 polymorphic loci revealed linkage imbalance in rs155979, rs3762983 and rs3762984. The frequency of CCG haploid types was significantly higher in patients than in controls (CCG vs GCG: OR = 0.735, 95% CI of 0.554–0.975, P = 0.032; Table 4). No significant correlation was found for other haploid types.
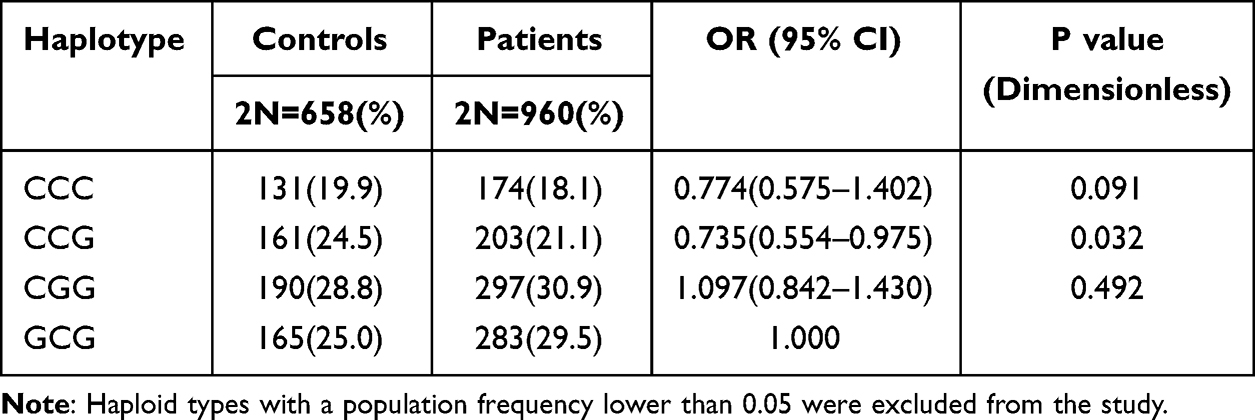 |
Table 4 Haplotype Analysis of the rs155979, rs3762983 and rs3762984 with the Depression |
rs2242385, rs155979, rs3762983 and rs3762984 polymorphic loci were used for interactive cross-validation (CV) using MDR (Multifactor Dimensionality Reduction 3.0.2) software. The results showed that rs2242385, rs155979, rs3762983 and rs3762984 interaction existed, and the best model was rs2242385-rs3762984 (CV training balanced accuracy:0.614; OR=3.101,95% CI of 2.217–4.336, P<0.001) (Table 5).
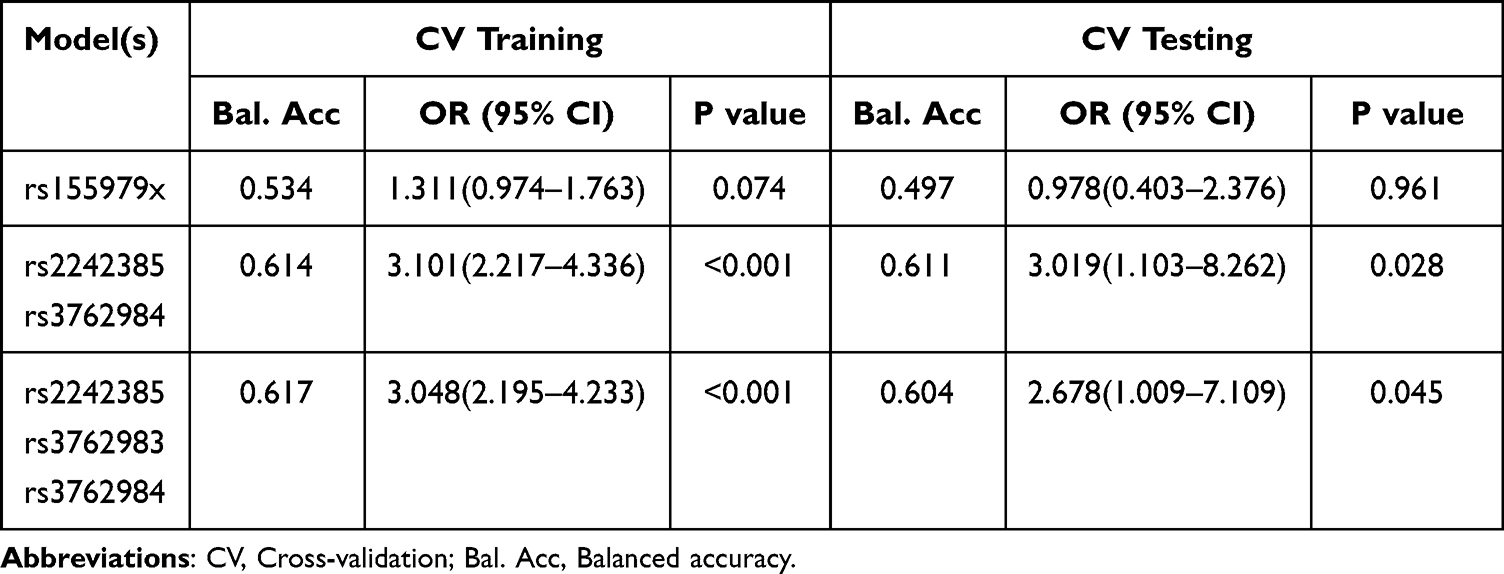 |
Table 5 The Cross-Validation Statistics of rs2242385-rs155979-rs3762983-rs3762984 Interaction |
Discussion
In this study, we investigated for the first time the contribution of genetic variation within lncRNA NONHSAT102891 gene to depression susceptibility. We found that the rs155979 GC genotype increased the risk of depression, and stratified analysis showed that the risk of suicide in patients with rs155979 GC or GG genotype was 2.08 times higher than that in the general population. Our results suggest that rs155979 polymorphism may be associated with the occurrence of depression in Chinese populations.
Depression is a common mental disorder with complex symptoms, associated with considerable morbidity, mortality, cost, and high risk of suicide. Some studies have estimated that the heritability is about 40%.17 However, until recently, genome-wide association (GWA) studies in large European sample groups failed to reliably identify genetic variations that could cause depression.18–22 Studies to investigate the prevalence of depression in different geographic regions and its association with population-specific SNPS are currently underway.23 Li Shiwu et al found that NEGR1 (regulated by TF combined with SNP rs3101339 to destroy depression risk) was dysregulated in the brains of patients with depression compared with the control group, and predicted that rs3101339 might confer depression risk by influencing NEGR1 expression.24 Parris et al found that the A allele of oxytocin receptor gene (OXTR) rs53576 was associated with a history of suicide attempts in patients with depression.25 The SNP rs10789336 located in the neuron growth regulator 1 (NEGR1) gene significantly affects the expression level of RPL31P12 in brain tissue and contributes to the risk of depression.26 Our data provide a new avenue to explore the pathophysiological mechanism of major depression.
LncRNAs are highly expressed in the central nervous system and regulate neural stem cell maintenance,27 synaptic plasticity, gliogenesis, neurogenesis,28 brain patterns, oligodendrocyte myelination, hippocampal development, and brain aging.29 Liu Zhifen et al identified 11 differentially expressed lncRNAs in patients with depression by using peripheral blood microarray analysis, which proved the correlation between lncRNA and depression.30 Kahaei et al evaluated the brain lncRNA expression patterns of a large number of patients with depression and identified 10 potential biomarkers.31 Guangyao Li et al confirmed that lncRNANONHSAT102891 was significantly down-regulated in PBMCs of patients with depression compared with the control.8 These results support our hypothesis; In this study, we investigated the pathophysiological mechanism of depression and confirmed that the rs155979 polymorphism in lncRNA NONHSAT102891 gene may be related to the occurrence of depression in Chinese population. Further research is needed to identify new susceptibility genes for depression and find precise therapeutic targets for depression. Haploid analysis results showed that the frequency of CCG haploid type in patients was significantly lower than that in healthy subjects, indicating that the CCG genotype reduced the risk of depression. Thus, the CCG haploid type might be a protective factor for depression, and the specific mechanism needs to be further explored.
Some previous studies of genetic associations have focused on the relationship between SNPs, coding genes and susceptibility to depression. Relatively little research has been done on the genetic variation of non-coding genes, such as RNA, especially lncRNA. In this study, we found that SNPS in the LncRNA region were associated with depression. This study also has some limitations. Peripheral blood was used to extract total DNA for further study. However, no other blood components or biological tissues were considered, which may provide new insights. The content of lncRNANONHSAT102891 in white blood cells was not considered in our sample collection and experiment design, and the experimental design lacking the influence of SNP on the expression of lncRNANONHSAT102891 was also a big shortcoming of us. Moreover, this study included subjects only from the central and southern parts of China. Regional differences and the relatively small sample size could affect the results, and further studies are needed on larger samples. In addition, only molecular experiments were conducted in this study, which lacked further verification by cell experiments. We will improve the above defects in the following work.
Ethics Approval and Consent to Participate
This study was conducted in accordance with the Declaration of Helsinki and approved by the ethics committee of Chengdu medical college. All patients signed an informed consent form for inclusion in the study.
Acknowledgement
This work was supported by the National Natural Science Foundation of China (No. 81901379) Natural Science Foundation of Sichuan(No.2022NSFSC0778).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Malhi GS, Mann JJ. Depression. Lancet. 2018;392(10161):2299–2312. doi:10.1016/S0140-6736(18)31948-2
2. Malhi GS, Coulston CM, Fritz K, et al. Unlocking the diagnosis of depression in primary care: which key symptoms are GPs using to determine diagnosis and severity? Aust N Z J Psychiatry. 2014;48(6):542–547. doi:10.1177/0004867413513342
3. Cai H, Xie XM, Zhang Q, et al. Prevalence of suicidality in major depressive disorder: a systematic review and meta-analysis of comparative studies. Front Psychiatry. 2021;12:690130. doi:10.3389/fpsyt.2021.690130
4. Thapa B, Torres I, Koya SF, et al. Use of Data to Understand the Social Determinants of Depression in Two Middle-Income Countries: the 3-D Commission. J Urban Health. 2021;98(Suppl 1):41–50. doi:10.1007/s11524-021-00559-6
5. Kopp F, Mendell JT. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell. 2018;172(3):393–407. doi:10.1016/j.cell.2018.01.011
6. Goyal B, Yadav SRM, Awasthee N, Gupta S, Kunnumakkara AB, Gupta SC. Diagnostic, prognostic, and therapeutic significance of long non-coding RNA MALAT1 in cancer. Biochimica Et Biophysica Acta Rev Cancer. 2021;1875(2):188502. doi:10.1016/j.bbcan.2021.188502
7. Labonté B, Engmann O, Purushothaman I, et al. Sex-specific transcriptional signatures in human depression. Nat Med. 2017;23(9):1102–1111. doi:10.1038/nm.4386
8. Li G, He M. Bioinformatics Analysis of Altered lncRNAs in Peripheral Blood Molecular Cells from Major Depressive Disorder (MDD) Patients. Blood Res Disorders. 2018;5(1):5.
9. Heidari Nia M, Sargazi S, Saravani R, Mirinejad S, Jahantigh D, Shakiba M. Relationship between GABRB2 gene polymorphisms and schizophrenia susceptibility: a case-control study and in silico analyses. Int J Neurosci. 2022;132(6):633–642. doi:10.1080/00207454.2020.1830087
10. Sargazi S, Mirani Sargazi F, Heidari Nia M, et al. Functional Variants of miR-143 Are Associated with Schizophrenia Susceptibility: a Preliminary Population-Based Study and Bioinformatics Analysis. Biochem Genet. 2022;60(3):868–881. doi:10.1007/s10528-021-10133-z
11. Sargazi S, Zahedi Abghari A, Mirinejad S, et al. Long noncoding RNA HOTAIR polymorphisms and susceptibility to bipolar disorder: a preliminary case-control study. Nucleosides Nucleotides Nucleic Acids. 2022;41(7):684–701. doi:10.1080/15257770.2022.2065017
12. Mokhtari MA, Sargazi S, Saravani R, et al. Genetic Polymorphisms in miR-137 and Its Target Genes, TCF4 and CACNA1C, Contribute to the Risk of Bipolar Disorder: a Preliminary Case-Control Study and Bioinformatics Analysis. Dis Markers. 2022;2022:1886658. doi:10.1155/2022/1886658
13. Sayad A, Badrlou E, Ghafouri-Fard S, Taheri M. Association Analysis Between the rs1899663 Polymorphism of HOTAIR and Risk of Psychiatric Conditions in an Iranian Population. J Mol Neurosci. 2020;70(6):953–958. doi:10.1007/s12031-020-01499-7
14. Liu W, Li W, Cai X, et al. Identification of a functional human-unique 351-bp Alu insertion polymorphism associated with major depressive disorder in the 1p31.1 GWAS risk loci. Neuropsychopharmacology. 2020;45(7):1196–1206. doi:10.1038/s41386-020-0659-2
15. Zeng Y, Navarro P, Shirali M, et al. Genome-wide Regional Heritability Mapping Identifies a Locus Within the TOX2 Gene Associated With Major Depressive Disorder. Biol Psychiatry. 2017;82(5):312–321. doi:10.1016/j.biopsych.2016.12.012
16. Wang L, Tang X, Liang P, et al.Correlation between variants of the CREB1 and GRM7 genes and risk of depression. BMC Psychiatry. 2023;23(1).doi:10.1186/s12888-022-04458-1
17. Corfield EC, Yang Y, Martin NG, Nyholt DR. A continuum of genetic liability for minor and major depression. Transl Psychiatry. 2017;7(5):e1131. doi:10.1038/tp.2017.99
18. Grotzinger AD, Mallard TT, Akingbuwa WA, et al. Genetic architecture of 11 major psychiatric disorders at biobehavioral, functional genomic and molecular genetic levels of analysis. Nat Genet. 2022;54(5):548–559. doi:10.1038/s41588-022-01057-4
19. Cai N, Bigdeli TB. Sparse whole-genome sequencing identifies two loci for major depressive disorder. Nature. 2015;523(7562):588–591. doi:10.1038/nature14659
20. Ripke S, Wray NR, Lewis CM, et al. A mega-analysis of genome-wide association studies for major depressive disorder. Mol Psychiatry. 2013;18(4):497–511. doi:10.1038/mp.2012.21
21. Wray NR, Ripke S, Mattheisen M, et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat Genet. 2018;50(5):668–681. doi:10.1038/s41588-018-0090-3
22. Hyde CL, Nagle MW, Tian C, et al. Identification of 15 genetic loci associated with risk of major depression in individuals of European descent. Nat Genet. 2016;48(9):1031–1036. doi:10.1038/ng.3623
23. Issler O, van der Zee YY, Ramakrishnan A, et al. Sex-Specific Role for the Long Non-coding RNA LINC00473 in Depression. Neuron. 2020;106(6):912–926.e915. doi:10.1016/j.neuron.2020.03.023
24. Li S, Li Y, Li X, et al. Regulatory mechanisms of major depressive disorder risk variants. Mol Psychiatry. 2020;25(9):1926–1945. doi:10.1038/s41380-020-0715-7
25. Parris MS, Grunebaum MF, Galfalvy HC, et al. Attempted suicide and oxytocin-related gene polymorphisms. J Affect Disord. 2018;238:62–68. doi:10.1016/j.jad.2018.05.022
26. Wang X, Cheng W, Zhu J, et al. Integrating genome-wide association study and expression quantitative trait loci data identifies NEGR1 as a causal risk gene of major depression disorder. J Affect Disord. 2020;265:679–686. doi:10.1016/j.jad.2019.11.116
27. Walker D, Walker D, Leach J. Anxiety: etiology, treatment, and Christian perspectives. J Christian Nursing. 2014;31(2):84–91. doi:10.1097/CNJ.0000000000000051
28. Huang X, Luo YL, Mao YS, Ji JL. The link between long noncoding RNAs and depression. Prog Neuropsychopharmacol Biol Psychiatry. 2017;73:73–78. doi:10.1016/j.pnpbp.2016.06.004
29. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
30. Liu Z, Li X, Sun N, et al. Microarray profiling and co-expression network analysis of circulating lncRNAs and mRNAs associated with major depressive disorder. PLoS One. 2014;9(3):e93388. doi:10.1371/journal.pone.0093388
31. Kahaei MS, Ghafouri-Fard S, Namvar A, Omrani MD, Sayad A, Taheri M. Association study of a single nucleotide polymorphism in brain cytoplasmic 200 long-noncoding RNA and psychiatric disorders. Metab Brain Dis. 2020;35(7):1095–1100. doi:10.1007/s11011-020-00582-7
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
rs217727 of lncRNA H19 is Associated with Cervical Cancer Risk in the Chinese Han Population
Dai J, Zhang S, Shi Y, Xu J, Liu W, Yang J, Shi L, Yan Z, Li C
Pharmacogenomics and Personalized Medicine 2023, 16:933-948
Published Date: 30 October 2023