Back to Journals » Neuropsychiatric Disease and Treatment » Volume 16
Association Between Glutathione Peroxidase-1 (GPx-1) Polymorphisms and Schizophrenia in the Chinese Han Population
Authors Shao X, Yan C ![]() , Sun D, Fu C
, Sun D, Fu C ![]() , Tian C, Duan L, Zhu G
, Tian C, Duan L, Zhu G ![]()
Received 15 July 2020
Accepted for publication 7 September 2020
Published 6 October 2020 Volume 2020:16 Pages 2297—2305
DOI https://doi.org/10.2147/NDT.S272278
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Yu-Ping Ning
Xiaojun Shao1 *,* Ci Yan1 *,* Dongxue Sun,1 Chunfeng Fu,1 Chunsheng Tian,1 Li Duan,1 Gang Zhu1,2
1Department of Psychiatry, The First Affiliated Hospital of China Medical University, Shenyang 110001, People’s Republic of China; 2Central Laboratory, The First Affiliated Hospital of China Medical University, Shenyang 110001, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Gang Zhu
Department of Psychiatry, Tthe First Affiliated Hospital of China Medical University, Shenyang 110001, People’s Republic of China
Tel/ Fax +86-24-83282184
Email [email protected]
Objective: The dopamine and oxidative stress hypotheses are leading theories of the pathoetiology of schizophrenia (SCZ). Glutathione Peroxidase 1 (GPx-1), a major antioxidant enzyme, and the most abundantly expressed member of the GPx family, plays an important role in metabolic dopamine changes, which are closely related to neurological and psychiatric disorders. The impact of GPx-1 polymorphisms has rarely been explored in the field of SCZ. Here, we explored the possible relationship between GPx-1 gene polymorphisms and SCZ in Chinese Han subjects by using the polymerase chain reaction-restriction fragment length polymorphism method.
Methods: DNA from 786 patients (360 patients with schizophrenia and 426 healthy controls) was genotyped for the single-nucleotide polymorphisms rs1800668 C/T and rs1050450 C/T in GPx-1 using polymerase chain reaction-restriction fragment length polymorphism analysis. Analysis of the association between GPx-1 and SCZ was performed using SPSS 22.0, while Haploview 4.2 software and SHEsis software were used to perform linkage disequilibrium analysis and haplotype analysis.
Results: The results indicated that the GPx-1 polymorphisms rs1050450 and rs1800668 were associated with SCZ. We found that the C-allele of rs1800668 C/T may be a protection factor against SCZ in general, but in particular, for males. Furthermore, the CT and TC (GPx-1 rs1800668 C/T and rs1050450 C/T) haplotypes may be susceptible to SCZ in the population. Finally, no significant differences in allelic or genotypic frequencies of rs1050450 were detected between cases and controls from whole or stratification analyses by gender.
Conclusion: GPx-1 polymorphisms are related to SCZ in Chinese Han subjects. Our results suggested that GPx-1 may be a potential gene that influences SCZ.
Keywords: case-control study, Chinese Han population, GPx1, polymorphism, schizophrenia
Introduction
Schizophrenia (SCZ) is a disabling and severe mental illness, which usually manifests in early adulthood and is clinically characterized by sensory, mental, emotional and behavioral disorders. Studies have shown that the lifetime prevalence of SCZ is about 1%, although the incidence of SCZ is increasing year by year.1–3 The etiology and pathophysiology of SCZ, however, are not clear, and it is acknowledged that they are almost certainly multifactorial and comprehensive. A multitude of evidence has shown that genetic factors,4 disturbances in various neurotransmitters (eg, dopamine [DA], glutamine, serotonin),5 neuroanatomical structure changes,6 and psychosocial factors7 all increase the risk of SCZ.
The DA hypothesis,8 which proposes excessive dopaminergic activity in the mesolimbic pathway results in the increased likelihood of SCZ, has been illustrated by the onset of psychosis induced by excessive amphetamine (which causes the release of DA).9 As a result, pharmacologists have been developing DA receptor blockade use for use as antipsychotic drugs.10 Several studies have used meta-analysis, a powerful tool to summarize contradicting results from different studies, to show that genetic variants of the dopamine D2 receptor gene and dopamine D4 receptor gene play major roles in etiology of SCZ.11–13
Glutathione peroxidase (GPx) is a key reactive oxygen species (ROS) scavenger14 that combines with catalase (CAT) and superoxide dismutase (SOD) to form an antioxidant defense system.15 GPx utilizes a reducing agent to eliminate hydrogen peroxide and lipid peroxides, preventing peroxidative damage to the cell membrane and other organelles by regulating glutathione (GSH).16,17 GPx is responsible for recycling GSH18 as well as regulating GSH regeneration.19 GSH is an electron donor in the redox process of GPx.20 GPx-1 is the most widely distributed enzyme of the five GPx homologues, and is especially abundant in vascular endothelial cells.21 The GPx-1 locus is located on chromosome 3p21.322 and contains two exons and one intron.23
There is considerable evidence that GPx has a strong association with some diseases, many with links to the DA system.24–28 In fact, DA is a major source of ROS in the central nervous system, as it can produce ROS and DA‐quinone during its autooxidation.29 GPx is expressed in neurons and glial cells and is involved in the endogenous response to central nervous system oxidative stress as a key enzyme in the antioxidant system18 in the brain. Mice deficient in GPx-1 were particularly susceptible to 6-hydroxydopamine injury, which is a hydroxylated derivative of dopamine.30 Meanwhile, both in vitro and in vivo, GPx-1 can protect against and attenuate 6-hydroxydopamine toxicity in dopaminergic neurons.31 GPx is also closely related to neurological and psychiatric disorders. Studies have found that in Alzheimer’s disease (AD), degeneration of the central cholinergic system and free radical metabolism are gradually aggravated, and that SOD and GPx activities in the hippocampus, which are closely related to the DA system, are significantly decreased over time.32 It has also been reported in animal experiments that the activities of GPx and reductase in AD-related APP/PS-1 mice treated with antioxidants before induced oxidation were higher than those in controls. Antioxidant treatment was able to significantly reduce oxidative stress and improve the cognitive impairment AD model mice.33 Studies have shown that in Parkinson’s disease (PD) patients, GSH loss occurs in the cell bodies and dendritic processes of dopaminergic nigral neurons,34 and total PD GSH is selectively exhausted in the early stages of PD.35,36
Many studies have shown that GPx plays an important role in the pathophysiology of SCZ. One study found that GPx activity in patients with SCZ was lower than in healthy controls.37 Vincent et al reported that low GSH and ascorbic acid levels in the brain are associated with dopaminergic system disorders, which are implicated in the development and progression of cognitive impairment in SCZ.38 Buckman et al found that brain atrophy measured after head computerized tomography scans in patients with chronic SCZ was negatively correlated with GSH-px activity in platelets, which was more significant in patients with SCZ with negative symptoms as the core clinical manifestation.39
Currently, a tremendous amount of domestic and foreign scientific research have proven that GPx-1 polymorphisms are associated with different diseases, such as coronary artery disease,22 cancer,40–45 diabetes and its vascular complications,46 rheumatoid arthritis,47 Huntington’s disease,48 PD,26,49 autism,50 and postpartum hemorrhage.51 However, research into the involvement of GPx-1 in SCZ is lacking.
This study detected GPx-1 polymorphisms by the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method, and explored whether there was a correlation between GPx-1 polymorphisms and SCZ in the Chinese Han population.
Materials and Methods
Patients and Controls
SCZ was confirmed in 360 subjects (male = 196, female = 164) with a mean age of 36.81 ± 10.75 years according to Diagnostic and Statistical Manual for Mental Disorders (DSM-IV) (American Psychiatric Association, 2004) criteria and confirmed independently by two senior psychiatrists based on a structured interview and medical records. All subjects were recruited from six hospitals in Liaoning Province, China. We also recruited 426 healthy Chinese Han subjects (male = 203, female = 223) with a mean age of 36.16 ± 8.89 years, who were without any mental disorders or neurological or physical diseases as controls. All subjects underwent a complete interview to exclude psychiatric disorders.
Principles of Ethical Review
This study was based on the Declaration of Helsinki. All volunteers were informed of the experimental method, objective, experimental benefits, risks, and their rights prior to signing informed consent. This study was approved by the Ethics Committee of China Medical University (Shenyang, Liaoning, China).
Polymorphism Genotyping
In this case-control study, rs1800668 and rs1050450 were selected based on a vast multitude of literature. For genomic DNA isolation, 2 mL of EDTA-treated blood from each volunteer was taken for DNA isolation using a DNA extraction kit (Wizard Genomic DNA purification kit, Thermo Fisher Scientific, Waltham, MA, USA). Detection of two single nucleotide polymorphisms, including rs1800668 and rs1050450, were performed by a PCR-based amplification strategy. RFLP analysis was carried out using the restriction enzymes HpyCH4III and BlpI (New England Biolabs, Ipswich, MA, USA). PCR primers were assembled by a local representative of Takara Biotechnology (Dalian, China). The forward primer 5ʹ-CAGCCTCCTATGCCAAACC-3ʹ and reverse primer 5ʹ-GCTCGTTCATCTGGGTGTAGTC-3ʹ were used to amplify rs1800668; the forward primer 5ʹ-CTCCCTTGTTTGTGGTTAG-3ʹ and reverse primer 5ʹ-CCTCCCTCGTAGGTTTAG-3ʹ were used to amplify rs1050450. PCR reaction conditions included 5 min degeneration (94 °C), 30 s degeneration (94 °C, 30 cycles), 30 s annealing (62.4 °C for 1800668 and 55 °C for rs1050450), 30 cycles of elongation for 1 min (72 °C), and a 10 min elongation (72 °C). The sizes of PCR products were 561 bp (rs1800668) and 529 bp (rs1050450). A 2% agarose gel was used to resolve PCR products and digested products. The lengths of restriction digest products were 330/231 bp for rs1800668 C/T, and 419/110 bp for rs1050450 C/T polymorphisms. The 561 bp PCR product for rs1800668 was cleaved into two fragments of 330 bp and 231 bp for the CC homozygote and three fragments of 561 bp, 330 and 231 bp were generated for the CT heterozygote, while an uncleaved PCR product of 561 bp was generated for the TT homozygote. For rs1050450, two fragments of 419 bp and 110 bp were generated for the CC homozygote and three fragments of 529 bp, 419 and 110 bp were generated for the CT heterozygote, while an uncleaved PCR product of 529 bp was generated for the TT homozygote.
Statistical Analysis
The research data involved in this experiment are expressed using frequency, percentage, and mean ± standard deviation (SD). The chi-square test or Fisher’s exact test were used to analyze the frequency of allele and genotype in patients and controls. All tests, including the odds ratio (OR) and 95% confidence interval (CI) were performed using SPSS v23.0 for windows (SPSS Inc., Chicago, IL, USA). Haploview 4.2 software (https://www.broadinstitute.org/haploview/haploview) and SHEsis software (http://analysis.bio-x.cn) were used to perform linkage disequilibrium and haplotype analysis. P-values < 0.05 were defined as statistically significant.
Results
Genotype Analysis
Rs1800668 and rs1050450 were genotyped by PCR-RFLP in this study. Agarose gels demonstrating homozygotes and heterozygotes for both SNPs are shown in Supplementary Figure S1 and Supplementary Figure S2.
Hardy–Weinberg Equilibrium
Table 1 describes the results for Hardy–Weinberg equilibrium estimations along with genotype and allele frequencies. These data indicated that the two selected single nucleotide polymorphisms (SNPs; rs1800668 and rs1050450) were in accordance with the genetic balance of Hardy–Weinberg equilibrium; thus, the selected samples were representative of the population.
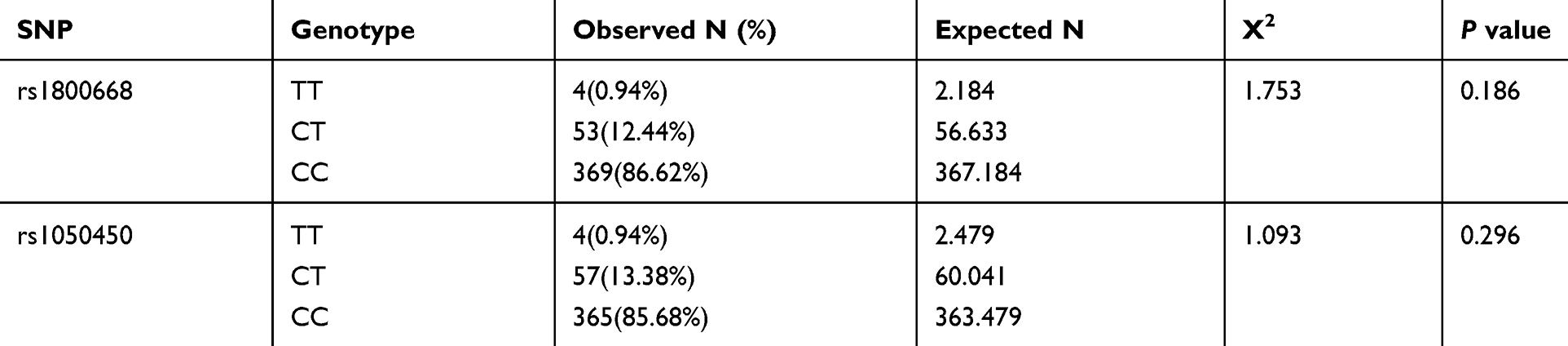 |
Table 1 Genotype Frequencies of GPx-1 Rs1800668 and Rs1050450 |
Association Between GPx-1 Polymorphisms and SCZ
Table 2 describes the genotype and allele distributions of the GPx-1 polymorphism (rs1800668) in the SCZ group and controls. There were significant associations in rs1800668 polymorphism genotype between the SCZ group and controls (X2 = 6.120, P = 0.013). There were significant differences in the frequency of the allele (X2 = 4.065, P = 0.044). We found that the C-allele of rs1800668 C/T may be a protective factor for SCZ (OR = 0.716, CI: 0.517–0.992). The frequency of GPx-1 rs1800668 in dominant and recessive models was then analyzed. The results showed that there was a significant difference in GPx1 rs1800668 in dominant models between the SCZ group and controls (X2 = 5.296, P = 0.021), but no significant difference was obtained using the recessive model frequency (X2 = 0.379, P = 0.693). Therefore, no significance was observed between the rs1050450 genotype and SCZ (see Supplementary Table S1). When analyzed separately by sex, we found a significant difference in rs1800668 among the frequency of different genotypic groups, alleles, and dominant models in males. Meanwhile, no difference was obtained in the recessive model. There was no difference between the females in the SCZ group and controls for rs1800668 (Table 3). Therefore, no sex effect was observed between the rs1050450 genotype and SCZ (see also the Supplementary Table S2).
 |
Table 2 Genotype and Allele Distributions of Rs1800668 in the SCZ Group and Controls |
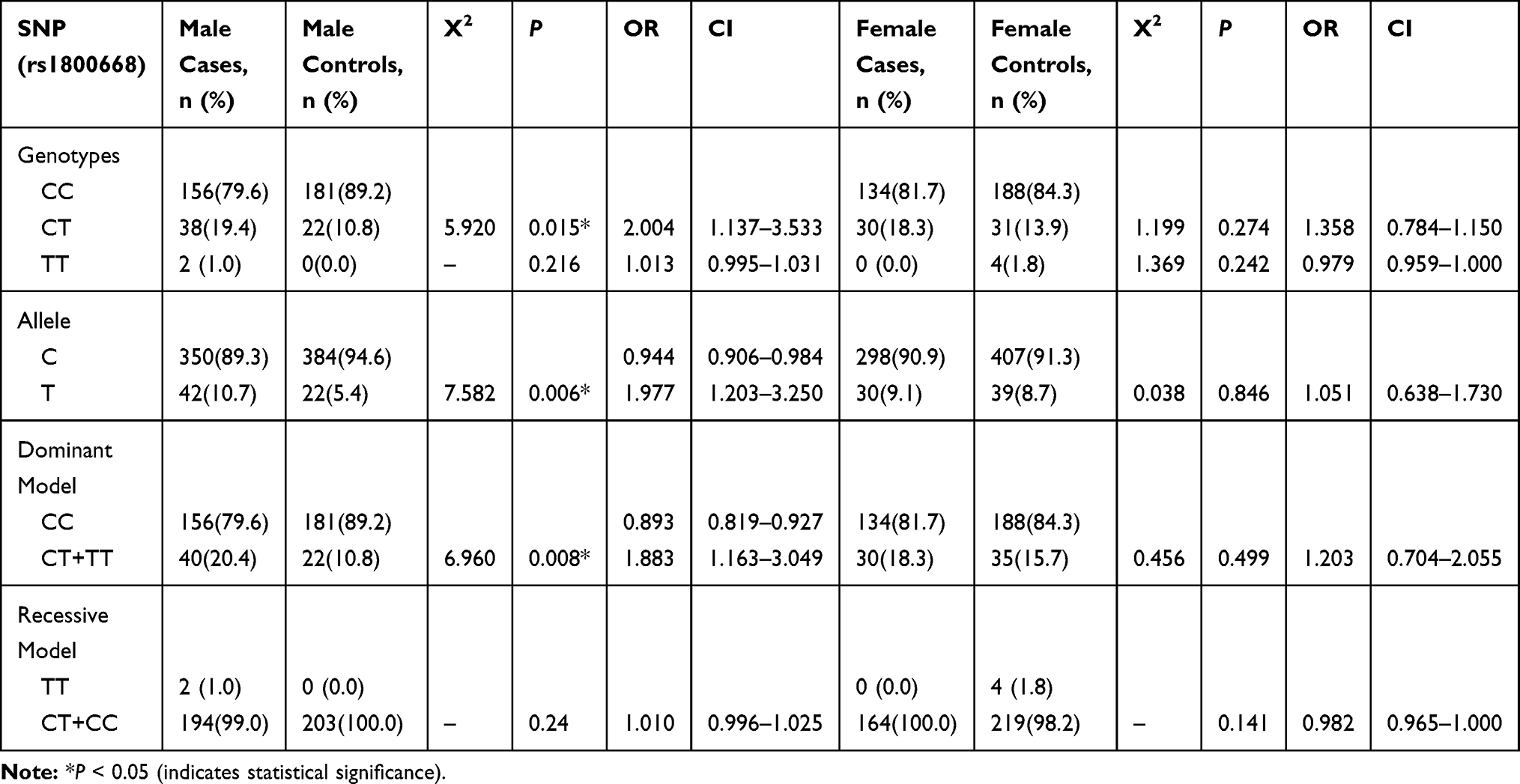 |
Table 3 Genotype and Allele Distributions of GPx-1 (Rs1800668) in the SCZ Group and Controls Separated by Gender |
Linkage Disequilibrium Analysis
Linkage disequilibrium, described by D´ and r2, refers to the occurrence of alleles belonging to two or more gene seats on a chromosome at the same time, which is higher than the random occurrence frequency, that is, the phenomenon of non-random distribution of haplotype genes. Here, D´= 0.600 and r 2 = 0.349; the linkage disequilibrium between rs1800668 and rs1050450 is shown in Figure 1.
 |
Figure 1 Linkage disequilibrium of rs1800668 and rs1050450. |
Haplotype Analysis
Haplotype analysis of rs1800668 and rs1050450 in the SCZ group and controls showed that the frequencies of CC, CT, TC, and TT were significantly different; as shown in Table 4.
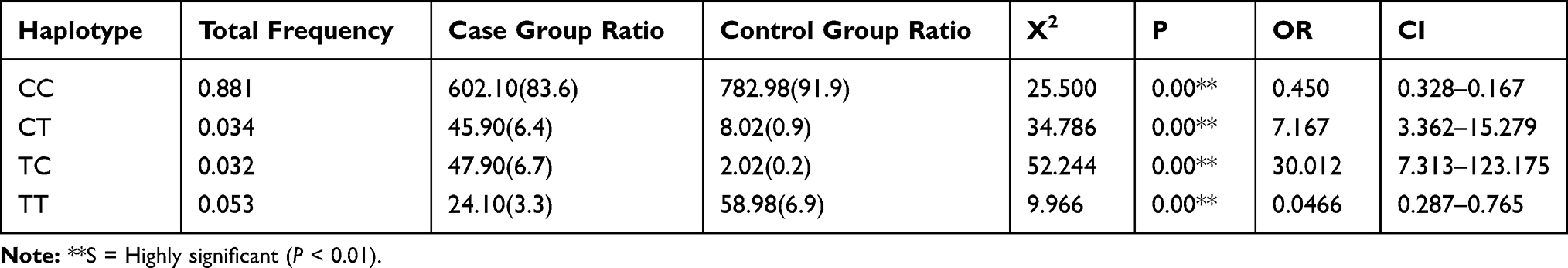 |
Table 4 Haplotype Distribution of Rs1800668 and Rs1050450 Between the SCZ Group and Controls |
Discussion
Various GPx-1 SNPs are evident in the general population, including Prol98Leu (rs1050450) in Exon 2, −602A/G, 2T/C in the promoter region, and Ala5/Ala6/Ala7 repeat polymorphisms in Exon 1.46 Additionally, there are 592G/A,52 and −46C/T (rs1800668)53 polymorphisms, among others. In this study, rs1050450 and rs1800668 were selected to evaluate associations between selected SNPs and SCZ in Chinese Han subjects as well as to establish the effect of sex. The principal findings of this study were that the GPx-1 polymorphisms rs1050450 and rs1800668 were significantly associated with SCZ in the Chinese Han population, and that, in particular, the CT and TC (rs1800668 and rs1050450) haplotypes may be risk factors for SCZ. Furthermore, the C allele may be a protective factor for SCZ in general, and in particular for males patients with the GPx-1 rs1800668 polymorphism.
Although a pathological mechanism of SCZ and a therapeutic strategy still remain unknown, the theory that oxidative stress plays an important role in SCZ pathophysiological mechanisms by triggering neuronal inflammation has gained increased attention.54 Oxidative stress is defined as the imbalance between ROS and antioxidant protection systems. When the brain’s demand for oxygen increases and the body fails to provide this oxygen in a timely manner, cerebral oxidative stress will occur,55 which subsequently causes damage to lipids, DNA, and proteins, all of which are related to the pathogenesis of SCZ.56 In cells exposed to ROS, GPx-1 showed superior protection against SOD and CAT.57 Studies have shown that GPx-1 polymorphisms, especially rs1800668, can affect the activity of the GPx-1 promoter, and that the C allele is related to increased GPx activity,40,58 which may play a role in improving antioxidant activity. Here, we found that the genotype and dominant model frequency of GPx-1 rs1800668 C/T showed statistically significant differences between the SCZ group and controls, which indicated that rs1800668 likely play a major role in the pathogenesis of SCZ in the Chinese Han population and significant differences were found between the dominant model and the recessive model in the two groups. To the best our knowledge, this study is the first to thoroughly investigate associations between GPx-1 rs1800668 and SCZ. GPx-1 rs1800668 increases the antioxidant capacity of patients with SCZ and protects proteins, lipids, and DNA from oxidative stress, and also helps to maintain normal signal transmission between neurons and normal transmission of neurotransmitters; all of which may be may be due to C-allele-mediated increases in GPx-1 activity.59 The GPx-1 polymorphism rs1800668 affects GPx-1 activity, and studies have shown that GPx-1 activity is strongly negatively correlated with brain atrophy and the ventricle/brain ratio in patients with SCZ, that is, the higher the GPx-1 activity the smaller the area of brain atrophy.60 The results of this study, meanwhile, showed that there was a statistical difference in the GPx-1 rs1800668 C/T alleles between the SCZ group and controls. Specifically, the C-allele may reduce the odds of SCZ, a result consistent with the conclusion that the C-allele may increase GPx-1 activity, and thus, increase antioxidant capacity and reduce the incidence of SCZ. Regarding sex stratification, there was a significant difference in the allelic and genotypic frequencies, as well as the dominant model in SCZ among men for rs1800668, with the C-allele having a protective effect. There was no difference detected between SCZ patients and controls in our study for rs1800668 in females. Indeed, many studies have found that men and women have different levels of oxidative stress, which are mainly the result of women having higher levels of oxidative stress in their lifetime,61–63 and also of genetic factors, which may impact the levels of oxidative stress. Taken together, the results indicate that the rs1800668 polymorphism is significantly related to SCZ, and this relation is likely due to fact that rs1800668 can reduce GPx-1 activity.
Among the existing reports of GPx-1 SNPs, rs1050450 has been the most extensively studied. This variation occurs at nucleotide 593 (rs1050450), creating a C ≥ T allele in Exon 2 that leads to a proline to leucine52 substitution at codon 198, which results decreased GPx-1 activity.64 We found, in agreement with a previous study, that no statistical difference was obtained in allelic or genotypic frequencies of rs1050450 between the SCZ group and controls.65 However, larger-scale studies from different regions are necessary to confirm these findings.
Although our results indicated that there was no association between GPx-1 gene polymorphisms and SCZ in women, we are not able to completely rule out any association between the two. We only detected the GPx-1 rs1800668 C/T and rs1050450 C/T polymorphisms. There are likely more polymorphisms on other loci that have yet to be discovered, several of which may shed new light on the associations between SCZ and sex. We also cannot rule out an important role of oxidative stress in the pathophysiology of SCZ in women, because antioxidants other than GPx-1, including SOD and CAT, act collectively in the response against oxidative stress, and each has an important role at different stages of free radical metabolism.66,67
Based on linkage disequilibrium and haplotype analyses, we observed a linkage disequilibrium in rs1800668 C/T and rs1050450 C/T between the SCZ group and controls. The CT and TC haplotype were found to increase the risk of SCZ, whereas the CC and TT haplotypes were found to have a protective effect against disease development. Since the haplotype of the locus involved in this experiment has not been explored in other studies, the results found here are not comparable. Generally, the effect of a single SNP is not as effective as that of multiple sites, and the results of this experiment suggest that we should further carry out relevant site research in the future, and in particular, explore as many sites as possible for a gene to generate more accurate experimental results.
It should be noted that there were a few limitations to this study. First, the sample size was relatively small. We will expand the size in future studies to further explore relationships between GPx-1 and SCZ. Second, we could not determine GPx-1, SOD or CAT enzyme activities, or indeed any oxidative stress markers, such as GSH. Further research may draw reliable conclusions by detecting these enzyme activities in SCZ patients with different genotypes. Therefore, other factors, including cross-sectional data and different recruitment centers, should be considered when conducting future studies.
Conclusions
GPx-1 polymorphisms are related to SCZ in the Chinese Han population. This is the first study to observe that the GPx-1 polymorphisms rs1050450 and rs1800668 were related to SCZ in the Chinese Han population. The results showed the CT and TC (rs1800668 and rs1050450) haplotypes may be risk factors for SCZ. Our results also suggest, that the C allele in the polymorphism rs1800668 may be a protective factor against SCZ in general, and in particular in males.
Acknowledgments
This work was supported by grants from the Major Project of the Department of Science & Technology of Liaoning Province (201JH8/10300019). We thank all the volunteers for participating in this study.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gysin R, Kraftsik R, Sandell J, et al. Impaired glutathione synthesis in schizophrenia: convergent genetic and functional evidence. Proc Natl Acad Sci U S A. 2007;104(42):16621–16626. doi:10.1073/pnas.0706778104
2. Freedman R, Wood AJJ. Schizophrenia. N Engl J Med. 2003;349(18):1738–1749. doi:10.1056/NEJMra035458
3. Guloksuz S, van Os J. The slow death of the concept of schizophrenia and the painful birth of the psychosis spectrum. Psychol Med. 2018;48(2):229–244.
4. Harrison PJ, Owen MJ. Genes for schizophrenia? Recent findings and their pathophysiological implications. Lancet. 2003;361(9355):417–419.
5. Harrison PJ. Neurochemical alterations in schizophrenia affecting the putative receptor targets of atypical antipsychotics. Focus on dopamine (D1, D3, D4) and 5-HT2a receptors. Br J Psychiatry Suppl. 1999;38:12–22.
6. Shenton ME, Dickey CC, Frumin M, McCarley RW. A review of MRI findings in schizophrenia. Schizophr Res. 2001;49(1–2):1–52.
7. Degnan A, Baker S, Edge D, et al. The nature and efficacy of culturally-adapted psychosocial interventions for schizophrenia: A systematic review and meta-analysis. Psychol Med. 2017;48:1–14.
8. Kubota M, Nagashima T, Takano H, et al. Affinity States of Striatal Dopamine D2 Receptors in Antipsychotic-Free Patients with Schizophrenia. Int J Neuropsychopharmacol. 2017;20(11):928–935.
9. Lieberman JA, Kane JM, Alvir J. Provocative tests with psychostimulant drugs in schizophrenia. Psychopharmacology. 1987;91(4):415–433.
10. Howes O, McCutcheon R, Stone J. Glutamate and dopamine in schizophrenia: an update for the 21 st century. J Psychopharmacol. 2015;29(2):97–115. doi:10.1177/0269881114563634
11. González-Castro TB, Hernández-Díaz Y, Juárez-Rojop IE, et al. The role of C957T, TaqI and Ser311Cys polymorphisms of the DRD2 gene in schizophrenia: systematic review and meta-analysis. Behav Brain Funct. 2016;12(1):29. doi:10.1186/s12993-016-0114-z
12. Schwab SG, Wildenauer DB. Genetics of psychiatric disorders in the GWAS era: an update on schizophrenia. Eur Arch Psychiatry Clin Neurosci. 2013;263(Suppl S2):S147–154. doi:10.1007/s00406-013-0450-z
13. Gejman PV, Sanders AR, Duan J. The role of genetics in the etiology of schizophrenia. Psychiatr Clin North Am. 2010;33(1):35–66. doi:10.1016/j.psc.2009.12.003
14. Do TD, Thi Mai N, Duy Khoa TN, et al. Molecular Characterization and Gene Expression of Glutathione Peroxidase 1 in Tor tambroides Exposed to Temperature Stress. Evol Bioinform Online. 2019;15:1176934319853580.
15. Tolomeo AM, Carraro A, Bakiu R, et al. Peroxiredoxin 6 from the Antarctic emerald rockcod: molecular characterization of its response to warming. J Comp Physiol B. 2016;186(1):59–71.
16. Hansen JM, Go Y-M, Jones DP. Nuclear and mitochondrial compartmentation of oxidative stress and redox signaling. Annu Rev Pharmacol Toxicol. 2006;46(1):215–234. doi:10.1146/annurev.pharmtox.46.120604.141122
17. Sies H. Glutathione and its role in cellular functions. Free Radic Biol Med. 1999;27(9–10):916–921. doi:10.1016/S0891-5849(99)00177-X
18. Salminen LE, Paul RH. Oxidative stress and genetic markers of suboptimal antioxidant defense in the aging brain: a theoretical review. Rev Neurosci. 2014;25(6):805–819.
19. Chance B, Sies H, Boveris A. Hydroperoxide metabolism in mammalian organs. Physiol Rev. 1979;59(3):527–605. doi:10.1152/physrev.1979.59.3.527
20. Dringen R. Metabolism and functions of glutathione in brain. Prog Neurobiol. 2000;62(6):649–671. doi:10.1016/S0301-0082(99)00060-X
21. Forgione MA, Weiss N, Heydrick S, et al. Cellular glutathione peroxidase deficiency and endothelial dysfunction. American Journal of Physiology-Heart and Circulatory Physiology. 2002;282(4):H1255–H1261. doi:10.1152/ajpheart.00598.2001
22. Kiss C, Li J, Szeles A, et al. Assignment of the ARHA and GPX1 genes to human chromosome bands 3p21.3 by in situ hybridization and with somatic cell hybrids. Cytogenetic and Genome Research. 1997;79(3–4):228–230. doi:10.1159/000134729
23. Ishida K, Morino T, Takagi K, Sukenaga Y. Nucleotide sequence of a human gene for glutathione peroxidase. Nucleic Acids Res. 1987;15(23):10051. doi:10.1093/nar/15.23.10051
24. Bai X, Wey MC-Y, Martinez PA, Shi C, Fernandez E, Strong R. Neurochemical and motor changes in mice with combined mutations linked to Parkinson’s disease. Pathobiology of Aging & Age-Related Diseases. 2017;7(1):1267855. doi:10.1080/20010001.2017.1267855
25. Cardoso BR, Ong TP, Jacob-Filho W, et al. Glutathione Peroxidase 1 Pro198Leu Polymorphism in Brazilian Alzheimer’s Disease Patients: relations to the Enzyme Activity and to Selenium Status. J Nutrigenet Nutrigenomics. 2015;29(2):72–80. doi:10.1159/000338682
26. Gardaneh M, Gholami M, Maghsoudi N. Synergy between glutathione peroxidase-1 and astrocytic growth factors suppresses free radical generation and protects dopaminergic neurons against 6-hydroxydopamine. Rejuvenation Res. 2013;263(S2):195–204. doi:10.1089/rej.2010.1080
27. Paz-y-Mino C, López-Cortés A, Muñoz MJ, et al. Genetic polymorphisms in apolipoprotein E and glutathione peroxidase 1 genes in the Ecuadorian population affected with Alzheimer’s disease. Am J Med Sci. 2006;46(1):373–377. doi:10.1097/MAJ.0b013e3181e93475
28. Power JHT, Blumbergs PC. Cellular glutathione peroxidase in human brain: cellular distribution, and its potential role in the degradation of Lewy bodies in Parkinson’s disease and dementia with Lewy bodies. Acta Neuropathol. 2009;117(1):63–73. doi:10.1007/s00401-008-0438-3
29. Zucca FA, Basso E, Cupaioli FA, et al. Neuromelanin of the human substantia nigra: an update. Neurotox Res. 2014;25(1):13–23. doi:10.1007/s12640-013-9435-y
30. Klivenyi P, Andreassen OA, Ferrante RJ, et al. Mice deficient in cellular glutathione peroxidase show increased vulnerability to malonate, 3-nitropropionic acid, and 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine. J Neurosci. 2000;20(1):1–7.
31. Ridet JL, Bensadoun JC, Déglon N, Aebischer P, Zurn AD. Lentivirus-mediated expression of glutathione peroxidase: neuroprotection in murine models of Parkinson’s disease. Neurobiol Dis. 2006;21(1):29–34.
32. Pocernich CB, Butterfield DA. Elevation of glutathione as a therapeutic strategy in Alzheimer disease. Biochim Biophys Acta. 2012;1822(5):625–630.
33. Huang Q, Aluise CD, Joshi G, et al. Potential in vivo amelioration by N-acetyl-L-cysteine of oxidative stress in brain in human double mutant APP/PS-1 knock-in mice: toward therapeutic modulation of mild cognitive impairment. J Neurosci Res. 2010;88(12):2618–2629.
34. Pearce RK, Owen A, Daniel S, Jenner P, Marsden CD. Alterations in the distribution of glutathione in the substantia nigra in Parkinson’s disease. J Neural Transmission. 1997;104(6–7):661–677.
35. Bannon MJ, Goedert M, Williams B. The possible relation of glutathione, melanin and 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine (MPTP) to Parkinson’s disease. Biochem Pharmacol. 1984;33(17):2697–2698.
36. Sian J, Dexter DT, Lees AJ, et al. Alterations in glutathione levels in Parkinson’s disease and other neurodegenerative disorders affecting basal ganglia. Ann Neurol. 1994;36(3):348–355.
37. Bošković M, Vovk T, Saje M, et al. Association of SOD2, GPX1, CAT, and TNF genetic polymorphisms with oxidative stress, neurochemistry, psychopathology, and extrapyramidal symptoms in schizophrenia. Neurochem Res. 2013;38(2):433–442.
38. Castagne V, Rougemont M, Cuenod M, Do KQ. Low brain glutathione and ascorbic acid associated with dopamine uptake inhibition during rat’s development induce long-term cognitive deficit: relevance to schizophrenia. Neurobiol Dis. 2004;15(1):93–105.
39. Buckman TD, Kling A, Sutphin MS, Steinberg A, Eiduson S. Platelet glutathione peroxidase and monoamine oxidase activity in schizophrenics with CT scan abnormalities: relation to psychosocial variables. Psychiatry Res. 1990;31(1):1–14.
40. Edith L, Joseph L, Handy DE. Glutathione peroxidase-1 in health and disease: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal. 2011;15(7):1957.
41. Hu YJ, Diamond AM. Role of glutathione peroxidase 1 in breast cancer: loss of heterozygosity and allelic differences in the response to selenium. Cancer Res. 2003;63(12):3347–3351.
42. Hu YJ, Dolan ME, Bae R, et al. Allelic loss at the GPx-1 locus in cancer of the head and neck. Biol Trace Elem Res. 2004;101(2):97–106.
43. Lee CH, Lee KY, Choe KH, et al. [Effects of oxidative DNA damage and genetic polymorphism of the glutathione peroxidase 1 (GPX1) and 8-oxoguanine glycosylase 1 (hOGG1) on lung cancer]. J Prevent Med Public Health. 2006;39(2):130–134.Korean.
44. Ravn-Haren G, Olsen A, Tjonneland A, et al. Associations between GPX1 Pro198Leu polymorphism, erythrocyte GPX activity, alcohol consumption and breast cancer risk in a prospective cohort study. Carcinogenesis. 2006;27(4):820–825.
45. Vogel U, Olsen A, Wallin H, Overvad K, Tjonneland A, Nexo BA. No association between GPX Pro198Leu and risk of basal cell carcinoma. Cancer Epidemiol Biomarkers Prevent. 2004;13(8):1412–1413.
46. Hamanishi T, Furuta H, Kato H, et al. Functional variants in the glutathione peroxidase-1 (GPx-1) gene are associated with increased intima-media thickness of carotid arteries and risk of macrovascular diseases in japanese type 2 diabetic patients. Diabetes. 2004;53(9):2455–2460.
47. Irfan S, Rani A, Sameem M, Nawaz SK, Liaqat I, Arshad M. Association of rs1800668 polymorphism in glutathione peroxidase- 1 gene and risk of rheumatoid arthritis in Pakistani population. Pakistan j Med Sci. 2016;32(5):1204–1207.
48. Zheng J, Winderickx J, Franssens V, Liu B. A Mitochondria-Associated Oxidative Stress Perspective on Huntington’s Disease. Front Mol Neurosci. 2018;11:329.
49. Redensek S, Flisar D, Kojovic M, et al. Genetic variability of inflammation and oxidative stress genes does not play a major role in the occurrence of adverse events of dopaminergic treatment in Parkinson’s disease. J neuroinflammation. 2019;16(1):50.
50. Ming X, Johnson WG, Stenroos ES, Mars A, Lambert GH, Buyske S. Genetic variant of glutathione peroxidase 1 in autism. Brain Dev. 2010;32(2):105–109.
51. Endler M, Saltvedt S, Eweida M, Akerud H. Oxidative stress and inflammation in retained placenta: a pilot study of protein and gene expression of GPX1 and NFkappaB. BMC Pregnancy Childbirth. 2016;16(1):384.
52. Ratnasinghe D, Tangrea JA, Andersen MR, et al. Glutathione peroxidase codon 198 polymorphism variant increases lung cancer risk. Cancer Res. 2000;60(22):6381–6383.
53. Peters U, Hayes RB, Chatterjee N, et al. Serum selenium and genetic variation in the selenoprotein GPX1 and risk of colorectal adenoma. Cancer Res. 2004;64.
54. Yao JK, Keshavan MS. Antioxidants, redox signaling, and pathophysiology in schizophrenia: an integrative view. Antioxid Redox Signal. 2011;15(7):2011–2035.
55. Liu C, Song S, Zhang J, Li X, Gao H. Effects of GSTA1 and GPX3 Polymorphisms on the Risk of Schizophrenia in Chinese Han Population. Neuropsychiatr Dis Treat. 2020;16:113–118.
56. Lohr JB. Oxygen radicals and neuropsychiatric illness. Some speculations. Arch Gen Psychiatry. 1991;48(12):1097–1106.
57. Takeshita S, Inoue N, Ueyama T, Kawashima S, Yokoyama M. Shear stress enhances glutathione peroxidase expression in endothelial cells. Biochem Biophys Res Commun. 2000;273(1):66–71.
58. Najafi M, Ghasemi H, Roustazadeh A, Alipoor B. Phenotype and genotype relationship of glutathione peroxidase1 (GPx1) and rs 1800668 variant: the homozygote effect on kinetic parameters. Gene. 2012;505(1):19–22.
59. Vecchio M, Curro M. The Oxidative Stress Response in Elite Water Polo Players: effects of Genetic Background. BioMed Res Int. 2017;2017:7019694.
60. Buckman TD, Kling AS, Eiduson S, Sutphin MS, Steinberg A. Glutathione peroxidase and CT scan abnormalities in schizophrenia. Biol Psychiatry. 1987;22(11):1349–1356.
61. Bartolini Gritti B, Binder CJ. Oxidation-specific epitopes are major targets of innate immunity in atherothrombosis. Hamostaseologie. 2016;36(2):89–96.
62. Jablonska E, Gromadzinska J, Reszka E, et al. Association between GPx1 Pro198Leu polymorphism, GPx1 activity and plasma selenium concentration in humans. Eur J Nutr. 2009;48(6):383–386.
63. Khadir A, Tiss A. Gender-specific association of oxidative stress and inflammation with cardiovascular risk factors in Arab population. BioMed Res Int. 2015;2015:512603.
64. Hong Z, Tian C, Zhang X. GPX1 gene Pro200Leu polymorphism, erythrocyte GPX activity, and cancer;risk. Mol Biol Rep. 2013;40(2):1801–1812.
65. Gao H, Liu C, Song S, et al. GPX1 Pro198Leu polymorphism and GSTP1 Ile105Val polymorphisms are not associated with the risk of schizophrenia in the Chinese Hanpopulation.Neuroreport.2017;28(15):969–972.doi:10.1097/WNR.0000000000000870
66. Balu DT. The NMDA Receptor and Schizophrenia: from Pathophysiology to Treatment. Adva Pharmacol. 2016;76:351–382.
67. Raffa M, Atig F, Mhalla A, Kerkeni A, Mechri A. Decreased glutathione levels and impaired antioxidant enzyme activities in drug-naive first-episode schizophrenic patients. BMC Psychiatry. 2011;11:124.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.