Back to Journals » Journal of Inflammation Research » Volume 19
Association Between Ferritinophagy and M1 Macrophage Polarization During the Acute Phase of Childhood Allergic Asthma
Authors Li J ![]() , Wang X
, Wang X ![]() , Shi X, Jiang Y, Shen R
, Shi X, Jiang Y, Shen R ![]()
Received 29 October 2025
Accepted for publication 7 February 2026
Published 18 February 2026 Volume 2026:19 569956
DOI https://doi.org/10.2147/JIR.S569956
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Jingjing Li,1,* Xing Wang,2,* Xiaofeng Shi,3 Yelin Jiang,4 Ren Shen5
1Department of Laboratory Medicine, The People’s Hospital of Yuhuan, Yuhuan, Zhejiang, People’s Republic of China; 2Department of Laboratory Medicine, Sichuan Provincial People’s Hospital Chuandong Hospital & Dazhou First People’s Hospital, Dazhou, Sichuan, People’s Republic of China; 3Department of Laboratory Medicine, Changxing County Hospital of Traditional Chinese Medicine, Huzhou, Zhejiang, People’s Republic of China; 4Department of Laboratory Medicine, Jiaxing Hospital of Traditional Chinese Medicine, Jiaxing, Zhejiang, People’s Republic of China; 5Department of Pediatrics, The People’s Hospital of Yuhuan, Yuhuan, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ren Shen, Department of Pediatrics, The People’s Hospital of Yuhuan, No. 18 Changle Road, Yucheng Street, Yuhuan, Zhejiang, 317600, People’s Republic of China, Tel +8615967683141, Email [email protected]
Background: The pathogenesis of allergic asthma is complex, and the role of macrophage polarization and ferritinophagy in childhood allergic asthma is still unclear. In this study, we preliminarily investigated the polarization status of splenic and lung macrophages during the acute phase of allergic asthma, and further explored the role of ferritinophagy in allergic asthma.
Methods: Detection of three monocyte subpopulations in PBMC of children with acute exacerbations of asthma and healthy children by flow cytometry. Establish an acute-phase model of allergic asthma in young mice sensitized with HDM, evaluate airway inflammation, airway resistance, and airway remodeling. We assessed the polarization status of macrophages in mouse spleen and lung tissues by flow cytometry, RT-PCR. We also detected changes in the ferritinophagy-related indexes, FTH1 and LC3B, with flow cytometry, and changes in co-localization of ferritinophagy-associated proteins in wall-isolated alveolar macrophages by laser confocal microscopy.
Results: We found an increase in the proportion of classical monocytes and a decrease in the proportion of non-classical monocytes in the PBMCs of asthmatic children. Therefore, we wanted to verify the changes in monocyte-derived macrophages in a young mouse model. We found that the proportion of splenic and lung macrophages was altered in the HDM-induced mice, which also showed an M1-polarized state. LC3B expression in macrophages increased while FTH1 expression decreased. In addition, we also found NCOA4 and LC3B in alveolar macrophages, FTH1 and LC3B, The co localization of FTH1 and LAMP1 increased, indicating that ferritinophagy was also enhanced.
Conclusion: Our research results indicate that macrophages in the acute phase model of allergic asthma in young mice exhibit M1 polarization, and an increase in ferritinophagy is associated with the pathogenesis of allergic asthma. These findings establish an associative (not causal) relationship between the two processes, providing new ideas for therapeutic strategies.
Plain Language Summary: What is the context?
Allergic asthma is a common respiratory disease in children. The exact mechanisms that trigger an acute asthma attack are not fully understood. Immune cells called macrophages play an important role in lung inflammation. A cellular process called “ferritinophagy”, which involves the recycling of iron, might also be involved in this process.
What did the study investigate?
This study aimed to investigate two key questions during the acute phase of childhood allergic asthma:
Do macrophages shift towards a pro-inflammatory state (known as M1 polarization)?
Is the ferritinophagy process activated in these macrophages?
What did the researchers do?
We first analyzed immune cells in the blood of children with asthma. Then, we established a house dust mite-induced allergic asthma model in young mice to mimic the human disease. We assessed airway inflammation and examined the polarization status of macrophages in the spleen and lungs of these mice. We also measured key indicators of ferritinophagy.
What were the main findings?
Children with asthma had a higher proportion of pro-inflammatory monocytes (precursors to macrophages) in their blood.
Mice with asthma showed significant lung inflammation and airway hypersensitivity.
Macrophages in the asthmatic mice were predominantly polarized to the pro-inflammatory M1 type in both the spleen and lungs.
The ferritinophagy process was significantly enhanced in these macrophages, indicating dysregulated iron metabolism.
What do the findings mean?
Our findings indicate that both M1 macrophage polarization and enhanced ferritinophagy are key features of acute allergic asthma in children. These two processes are likely interconnected and contribute to the worsening of inflammation. Targeting either macrophage polarization or ferritinophagy could offer new therapeutic strategies for treating allergic asthma.
Keywords: childhood allergic asthma, HDM, macrophage polarization, ferritinophagy, LC3B, FTH1
Introduction
Asthma affects nearly 350 million people worldwide, and its prevalence is still on the rise, causing a serious economic burden.1 Its typical pathological characteristics are: airflow limitation, airway inflammation and remodeling.2 House dust mite (HDM) allergy is a major trigger, approximately 80% of children with asthma being sensitized to dust mites. A study on adolescents in Guangzhou demonstrated that the risk of developing asthma in individuals allergic to house dust mites is 3.18 times higher than that in non-allergic counterparts.3 While food allergens are more prevalent in infants and young children, HDM allergy gradually becomes the dominant form with increasing age.4,5 The immune mechanisms underlying this condition are closely tied to monocyte and macrophage activity, which play pivotal roles in inflammatory responses.
Upon acute lung inflammation, monocytes are recruited from the bone marrow or circulation and infiltrate into the lung.6 Human monocytes comprise distinct subpopulations—classical, intermediate, and non-classical—based on CD14 and CD16 expression, whose proportions alter under pathological conditions.7,8 Classical monocytes are considered “pro-inflammatory” and serve as the primary source of tissue macrophages. Notably, the functional transition from classical to non-classical monocytes is impaired during inflammatory processes, influencing the quantity and function of their macrophage derivatives.9–12 Macrophages, as important immune cells in the body, are closely associated with the development of asthma. Asthma encompasses distinct phenotypes and endotypes, with immune cell dysfunction including macrophage polarization serving as a key mechanistic driver of specific endotypes.13 For pediatric allergic asthma, accumulating evidence indicates that M1 macrophage-derived pro-inflammatory cytokines (eg, TNF-α, IL-6) are key mediators of airway inflammation, and their abnormal secretion correlates with disease severity.14 Meanwhile, acute exacerbations of childhood asthma are frequently linked to aberrant differentiation of monocyte-derived macrophages, which disrupts the balance of lung immune homeostasis.15 Macrophages exhibit polarization and differentiation into different phenotypes in an inflammatory environment, with classical activation (M1) and selective activation (M2) being the two main phenotypes.16–18 During an allergic reaction, M1 macrophages produce cytokines and chemokines. The pro-inflammatory factors secreted by them not only exacerbate airway hyperresponsiveness but also actively regulate the chemotactic recruitment of neutrophils, thereby amplifying the inflammatory response and causing damage to lung tissues.19 Furthermore, in cases of acute allergen exposure, both the excessive secretion of airway mucus and the production of certain inflammatory factors require the involvement of inducible nitric oxide synthase (iNOS) expression.20 As a polarization-inducing factor, Tumor Necrosis Factor α (TNFα) synergistically drives the M1 phenotype through multiple pathways. Meanwhile, M1 macrophages recruit eosinophils via TNFα secretion, which in turn exacerbates airway mucus hypersecretion.21,22 M2 macrophages contribute to anti-inflammatory responses and tissue remodeling.23 Mannose receptor, MRC1 (CD206) is not only a specific marker of M2 macrophages, but also a core molecule for their polarization regulation, functional execution, and pathological effects.24,25 M1 or M2 undergo reversible functional changes, enabling them to respond to inflammation in the body.26,27 Therefore, intervening in the regulation of macrophage phenotype by targeting some potential regulatory factors of macrophages may have therapeutic potential in allergic asthma. Our prior clinical study identified enhanced M2 polarization of peripheral blood mononuclear cell-derived macrophages in children with acute allergic asthma exacerbations,28 while the current work focuses on tissue-resident macrophages and their M1 polarization associated with ferritinophagy.
Emerging evidence suggests a link between iron metabolism, ferroptosis, and asthma pathogenesis. Ferroptosis, an iron-dependent form of cell death driven by lipid peroxidation, is implicated in airway epithelial injury and macrophage dysfunction in asthma, with its occurrence heavily reliant on intracellular free iron levels.29 Autophagy, a lysosomal degradation process for damaged organelles or proteins, is marked by Microtubule-Associated Protein 1 Light Chain 3 Beta (LC3B).30 Of particular relevance is ferritinophagy, a selective autophagy process mediated by nuclear receptor coactivator 4 (NCOA4), which acts as a selective autophagy receptor for ferritin heavy chain-1 (FTH1). This process mediates the transport of ferritin to autolysosomes, leading to free iron release.31,32 Lysosome-associated membrane protein 1 (LAMP1) serves as a lysosomal marker, maintaining lysosomal integrity and participating in membrane transport processes.33 Excessive free iron catalyzes the generation of reactive oxygen species (ROS) through the Fenton reaction, thereby promoting the occurrence of ferroptosis. Consequently, ferritinophagy plays a central role in regulating intracellular iron balance and is closely associated with asthma occurrence.34 However, it remains unclear whether macrophage ferritinophagy regulates the development of allergic asthma by altering macrophage polarization homeostasis.
Given the complex and not fully elucidated pathogenesis of allergic asthma, continuous investigation into its immune mechanisms is essential for identifying novel therapeutic targets. In this study, we established a house dust mite (HDM)-induced acute asthma model in juvenile mice. We investigated the polarization status of macrophages and changes in ferritinophagy in this model. Our findings reveal a significant M1-type polarization phenotype in macrophages as well as an increase in ferritinophagy activity during the acute onset of asthma. These results not only elucidate the critical roles of aberrantly activated ferritinophagy and M1 macrophage polarization in acute allergic asthma in children, but also provide an experimental basis and theoretical support for new therapeutic strategies targeting iron metabolism and macrophage dysfunction.
Materials and Methods
Ethical Statement
This study was conducted in accordance with the principles of the Declaration of Helsinki. The clinical component was reviewed and approved by the Ethics Committee of Yuhuan People’s Hospital (No. 2022-061). The animal experiments were performed following the National Institutes of Health’s “Guide for the Care and Use of Laboratory Animals” and were approved by the Animal Health and Welfare Committee of Wenzhou Medical University Affiliated First Hospital (NO. 2021-0248). The reporting of animal research adheres to the ARRIVE guidelines 2.0.
Patient Selection and Specimen Collection
The study cohort comprised 16 children with allergic asthma and 15 healthy controls recruited from the People’s Hospital of Yuhuan. Asthmatic children aged 5–14 years, diagnosed with allergen-induced asthma, were enrolled. Age-matched healthy children undergoing routine health checks during the same period served as controls. No significant differences in baseline characteristics (age, height, weight) were observed between groups (Table 1). After obtaining parental informed consent, peripheral blood was drawn from all participants. Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Paque density gradient centrifugation. The isolated cells were cryopreserved in serum-free, protein-free freezing medium (NCM Biotech, Suzhou, China) at −80°C for subsequent analysis.
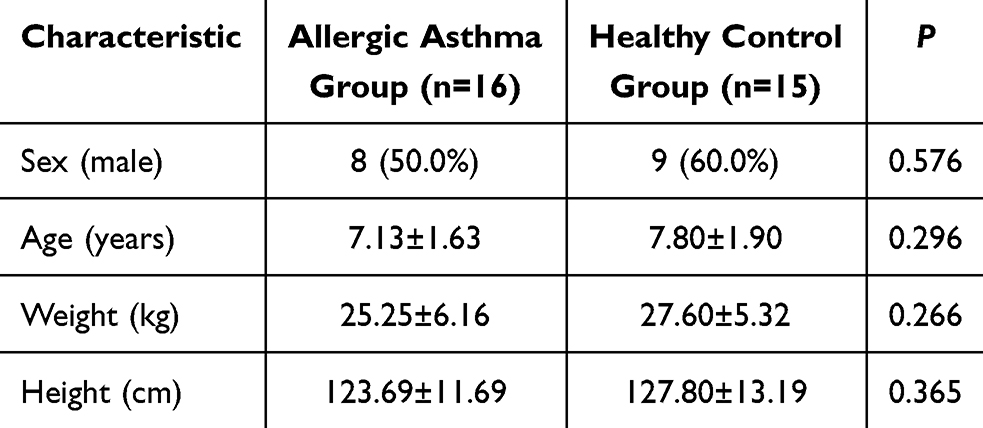 |
Table 1 Comparison of Two Sets of General Information |
Animals
Female C57BL/6J mice (five weeks old, 16 ± 2g) were procured from Zhejiang Vital River Laboratory Animal Technology Co., Ltd. All mice were housed under specific pathogen-free (SPF) conditions at the Wenzhou Medical University Experimental Animal Center, maintaining a controlled environment (temperature 22 ± 2°C, humidity 50 ± 5%) with a standard light-dark cycle and ad libitum access to food and water. Only healthy mice displaying normal behavior were included in the study. Exclusion criteria, established a priori, comprised: 1) accidental mortality during the procedure, and 2) severe respiratory distress (manifested as resting cyanosis) following HDM sensitization or challenge. Data points resulting from technical failures (eg, flow cytometer clogging, RNA degradation) were excluded during the analysis phase. Group sizes (n=5–14) were determined based on common practice in established HDM-induced allergic asthma models (eg, n=6–10 per group in prior studies),35,36 our pilot data, and resource availability.
Immunoblotting
Serum allergen-specific IgE profiling for 10 inhalant and 9 food allergens was performed on asthmatic children (n=16) using a commercial semi-quantitative immunoblotting kit (EUROIMMUN, Luebeck, Germany), following the manufacturer’s protocol. In brief, patient serum samples were incubated with allergen-coated membrane strips. After washing, alkaline phosphatase-conjugated anti-human IgE antibody was added. Following another wash cycle, a substrate solution was applied to develop colorimetric signals. The air-dried strips were then scanned and analyzed using dedicated software for semi-quantification.
Allergen-Induced Mouse Asthma Model
Before the experiment, all mice were kept for 1 week to adapt to the laboratory environment. The mice were randomly divided into two groups: Phosphate buffered saline (PBS), and House dust mite (HDM). The mice were sensitized with 100 μg HDM (Greer Labs, House Dust Mite, NC, USA) by means of transoral intubation (i.t.) on day 0, from days 14 to 19, the sensitized mice were challenged with 10 μg HDM in the same way as previously, control mice were treated with PBS, each dosing volume was 25μL, the mice were inhaled with 3% −4% concentration of isoflurane (RWD, Isoflurane, Guangdong, China) for 1 minute and rapidly intubated after anesthesia. On the 20th day, the mice were euthanized under anesthesia using well-trained professional techniques through rapid and painless cervical dislocation. To minimize potential confounders, the order of daily treatments (PBS or HDM) and the cage locations on the rack were randomized throughout the experiment.
Lung Histopathology
The lung tissues were perfused and fixed in 4% paraformaldehyde for 48 hours. Following paraffin embedding, 4 μm-thick sections were prepared. For histological assessment, sections from each group (n=6) were stained with Hematoxylin and Eosin (H&E) to evaluate inflammatory cell infiltration, Periodic Acid-Schiff (PAS) to visualize goblet cell hyperplasia and mucus, and Masson’s Trichrome to assess collagen deposition. Image-J software was utilized to quantify the area percentage of collagen fibers in Masson-stained sections.
Airway Hyperresponsiveness (AHR) Measurement
Within 24 hours of the last HDM challenge, AHR was accurately assessed using the Flexivent system (SCIREQ, FlexiVent, Montreal, QC, Canada). Mice were anesthetized, tracheotomized, and connected to the ventilator. Baseline lung function was recorded, and airway resistance (Rn) was measured in response to increasing doses of nebulized methacholine (0, 3.125, 6.25, 12.5, 25, 50 mg/mL).
Quantitative Real-Time PCR
Total RNA was extracted from mouse lung tissue using Trizol reagent. RNA quality and concentration were determined before reverse transcription into cDNA using a commercial kit (PrimeScript™ FAST RT Reagent Kit with gDNA Eraser, Takara, Japan). Quantitative PCR was performed on a CFX Connect™ system (Bio-Rad, USA) with gene-specific primers (Table 2). β-ACTIN served as the endogenous control. The relative mRNA expression levels of target genes (TNF-α, iNOS, CD206, NCOA4, LC3B) were calculated using the 2−ΔΔCt method.
 |
Table 2 Primer Sequences |
Enzyme-Linked Immunosorbent Assay (ELISA)
Blood was collected from the retro-orbital plexus of anesthetized mice. Serum was separated by centrifugation and stored at −80°C. The concentration of total IgE in mouse serum was quantified using a commercial Mouse IgE ELISA Kit (Dakewe Biotech, China) according to the manufacturer’s instructions.
Flow Cytometry
PBMCs
Living cells were selected using Zombie B550rm Fixable Viability Kit (1:200 PBS), staining on ice for 20 minutes. Then stain the cell surface with pre-mixed antibodies (Supplement Table 1) on ice for 30 minutes. After staining, fix and permeabilize the cells with eBioscience™ Fixation/Permeabilization kit (BD Biosciences, CA, USA), and then perform intracellular staining (Supplement Table 2). The above process requires avoiding light.
Mice Cells
Single-cell suspensions derived from mouse blood, spleen and lungs were collected and blocked for 15 min with anti-CD16/CD32 in staining buffer to prevent non-specific binding. For cell surface staining, cells were stained with the antibody (Supplement Table 1) for 20 min on ice. The subsequent steps are the same as PBMC (Supplement Table 2). The samples were tested on the machine (BD, LSRFortessa X-20, CA, USA), the results were analyzed using FlowJo software.
BAL Collection and Alveolar Macrophage Isolation
Following euthanasia, the trachea was exposed and cannulated. The lungs were lavaged ten times with 1 mL of ice-cold sterile PBS per wash. The collected bronchoalveolar lavage fluid (BALF) was pooled per mouse and centrifuged (500g, 5 min, 4°C). The cell pellet was resuspended in complete RPMI-1640 medium (supplemented with 10% FBS and 1% penicillin-streptomycin) and plated. After a 1-hour adhesion period at 37°C, non-adherent cells were removed by gentle washing with PBS. The adherent population, highly enriched for alveolar macrophages (>98% purity as confirmed by flow cytometry), was used for subsequent experiments.
Immunofluorescence
Adherent alveolar macrophages were fixed and permeabilized on ice using a solution of 4% paraformaldehyde containing 0.3% Triton X-100 for 20 minutes. Cells were then incubated overnight at 4°C with primary antibodies against LC3B (Proteintech, CL488-18725, 1:100), FTH1 (Thermo Fisher Scientific, 39G3, 1:100), NCOA4 (Proteintech, 10968-1-AP, 1:300), and LAMP1 (Thermo Fisher Scientific, MA1-164, 1:100). After PBS washes, appropriate fluorescently-labeled secondary antibodies (Proteintech, 1:500) were applied for 1.5 hours at room temperature in the dark. Nuclei were counterstained with DAPI. Finally, images were captured by a confocal microscope (Nikon, Japan) and analyzed using the ImageJ software.
Statistical Analysis
Data are presented as mean ± SEM. Statistical analyses were performed in GraphPad Prism 9.0. Normality and homogeneity of variance were assessed using the Shapiro–Wilk and F-tests, respectively. Two-group comparisons were made with an unpaired Student’s t-test if assumptions were met, or with the Mann–Whitney U-test otherwise. P-values < 0.05 were considered statistically significant.
Results
House Dust Mite (HDM) is Identified as the Primary Allergen in Asthmatic Children
The distribution of participants with positive allergen results is shown in (Table 3). The results indicated that house dust mite (HDM) was the most prevalent allergen among the 16 included asthmatic children, and the serum IgE levels in asthmatic children, as detected by the cobas® e 601 module (Roche, Diagnostics, USA), were significantly higher than those in the healthy children group (Figure 1D). The experimental framework for the clinical sample analysis is outlined in (Figure 1A).
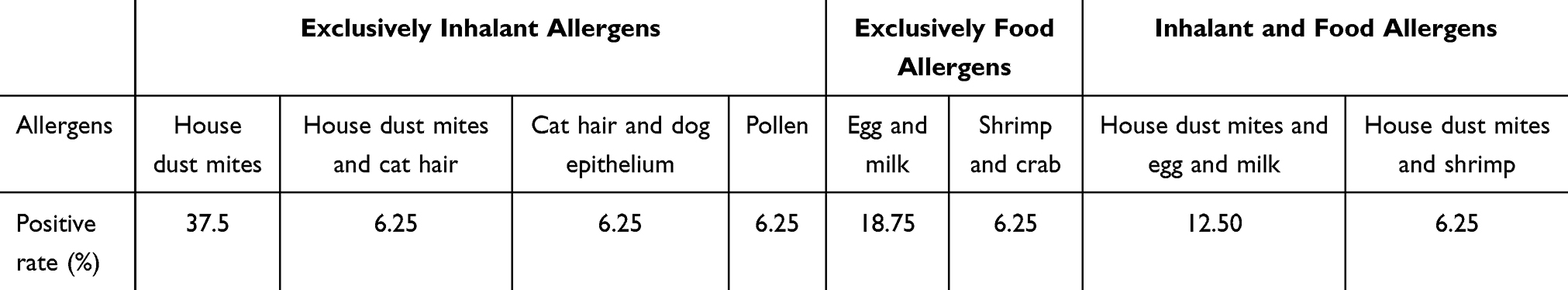 |
Table 3 Distribution of Allergen Positivity Rates |
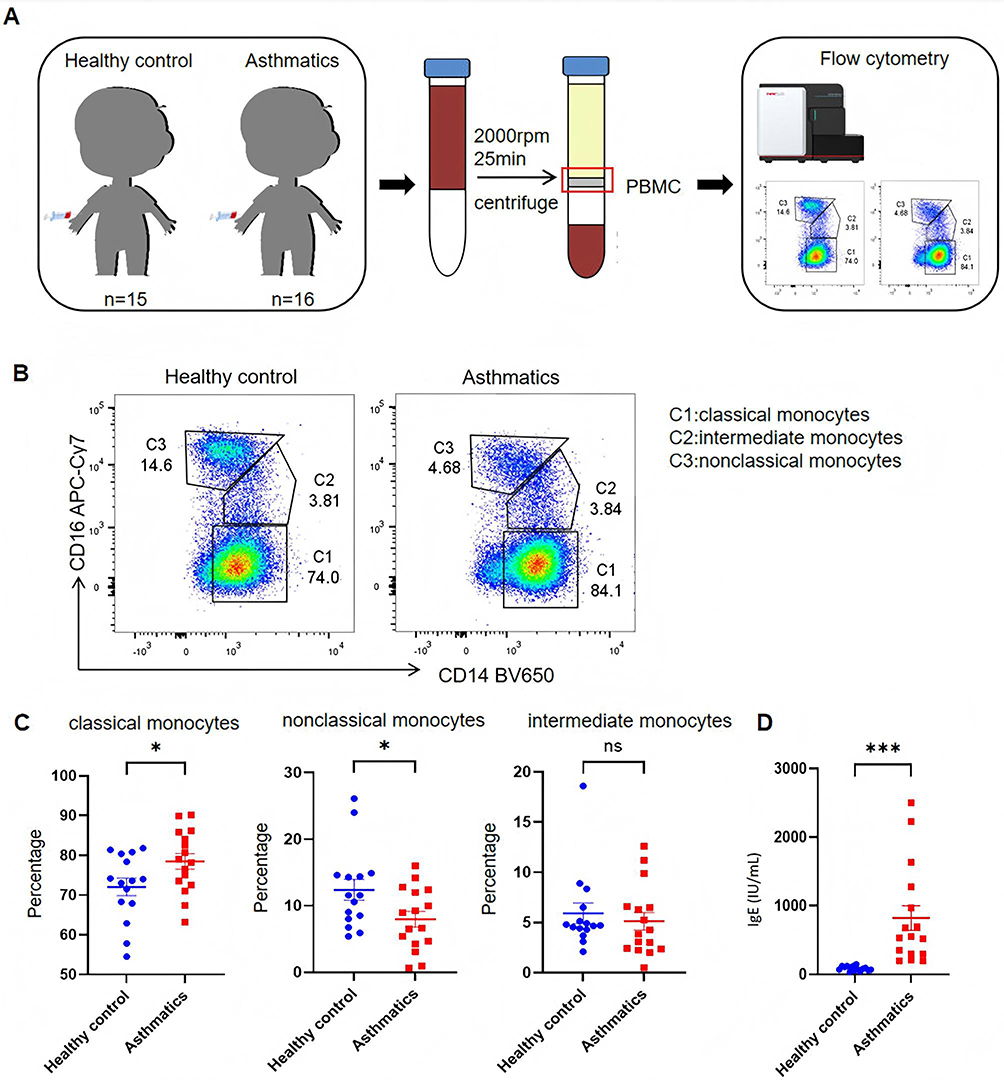 |
Figure 1 Analysis of Monocyte Subpopulations in PBMCs From Children With Allergic Asthma. (A) Schematic of the human sample workflow: Peripheral Blood Mononuclear Cells (PBMCs) were isolated from healthy controls (n = 15) and asthmatic children (n = 16) via centrifugation, followed by monocyte subpopulation analysis using flow cytometry. (B) Representative flow cytometry plots (gated on CD14/CD16) showing three monocyte subsets: C1 (classical monocytes, CD14++CD16-), C2 (intermediate monocytes, CD14++CD16+), and C3 (non-classical monocytes, CD14+CD16++). (C) Quantified proportions of each monocyte subpopulation in PBMCs from healthy controls and asthmatic children. (D) Serum IgE levels (measured by ELISA) in healthy controls and asthmatic children. *P<0.05, ***P<0.001, ns: not significant (P≥0.05). |
Elevated Levels of Classical Monocytes in PBMCs of Children with Acute Asthma
Our gating strategy (Supplement Figure 1A) recognized three subpopulations of monocytes in PBMCs: classical (CD14++CD16-), intermediate (CD14++CD16+) and nonclassical (CD14+CD16++) subpopulations. The proportion of classical monocytes was found to be elevated and the proportion of nonclassical monocytes was found to be decreased in the asthma group in PBMCs relative to the healthy control group, as detected by flow cytometry (Figure 1B and C). Classical monocytes are pro-inflammatory and are generally recognized as a major source of macrophages.37,38 Therefore, we would like to further investigate the changes in macrophages in allergic asthma.
Airway Hyperresponsiveness and Inflammation in HDM-Induced Mouse Asthma Model
To further investigate monocyte changes observed in asthmatic children, we established an HDM-induced asthma model in young mice (Figure 2A). HDM-challenged mice demonstrated significant airway hyperresponsiveness during methacholine challenge (Figure 2B) and exhibited elevated serum IgE levels compared to PBS controls (Figure 2C). Histological analysis showed that HDM-treated mice developed substantial airway inflammation, evidenced by increased inflammatory cell infiltration in H&E-stained sections (Figure 2D and E), enhanced goblet cell hyperplasia in PAS staining (Figure 2F and G), and elevated collagen fiber content in Masson trichrome staining (Figure 2H and I). The above experiments validated the successful establishment of the asthma model in young mice.
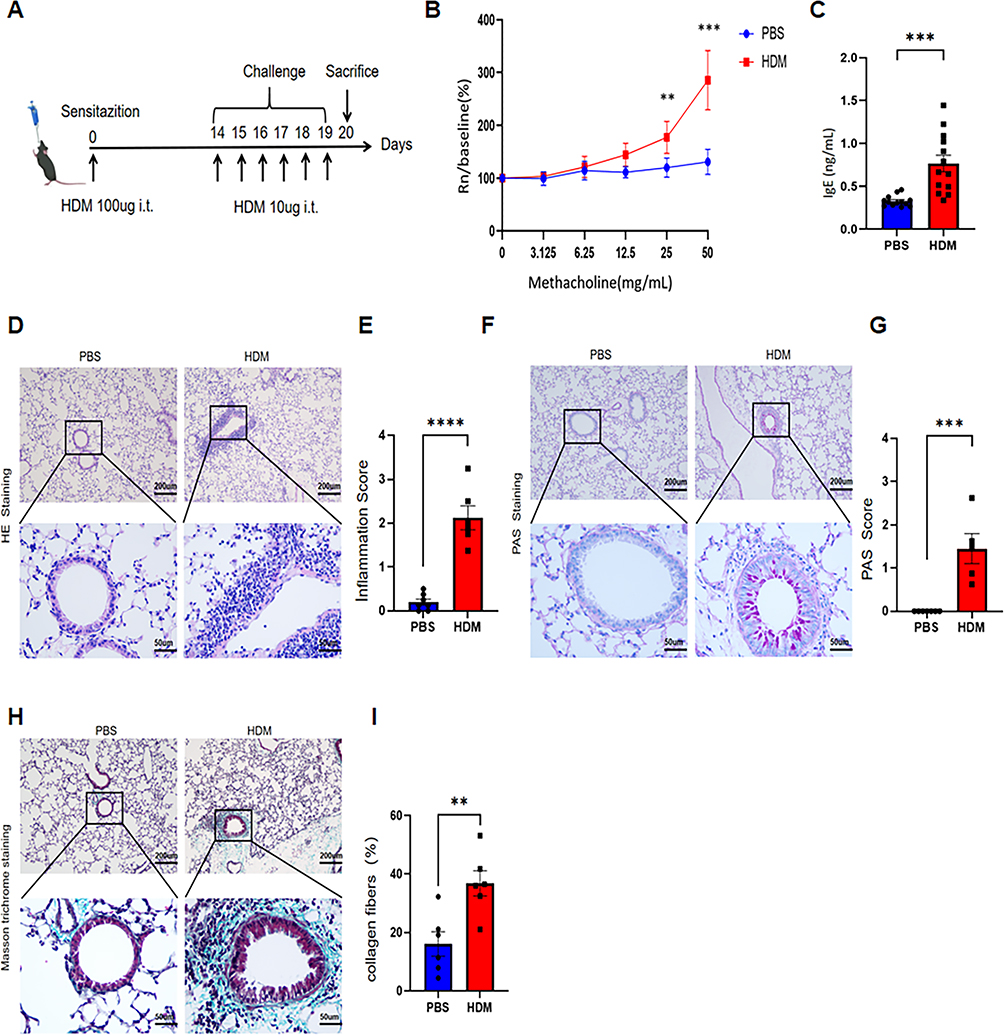 |
Figure 2 Establishment and Validation of HDM-Induced Mouse Allergic Asthma Model. (A) Schematic of the HDM-induced allergic asthma model in mice. (B) Airway responsiveness (Rn) to increasing methacholine concentrations in PBS- and HDM-treated groups (n = 5 per group). (C) Serum IgE levels by ELISA in PBS or HDM sensitized and challenged mice (n=13 per group). (D) Representative H&E-stained lung tissue sections (scale bars: 200 μm and 50 μm, respectively). (E) Quantified inflammatory scores from H&E staining. (F) Representative PAS-stained lung tissues sections (scale bars: 200 μm and 50 μm, respectively). (G) Quantified PAS scores. (H) Representative Masson trichrome-stained lung tissues sections (scale bars: 200 μm and 50 μm, respectively). (I) Percentage of collagen fiber content in airway tissues. (n = 6). **P<0.01, ***P<0.001, ****P<0.0001. |
Since inflammatory monocytes play an important role in allergic asthma,39 we developed a set-gate strategy (Supplement Figure 1B) and examined the peripheral blood cells of two groups of mice by flow cytometry, the results showed that the proportion of neutrophils and eosinophils increased in the HDM group (Figure 3A and B), and the proportion of monocytes significantly increased (Figure 3C). The above validation indicates that the mouse model has consistent characteristics with human allergic asthma, making it an ideal mouse model for our research on macrophages derived from monocytes.
 |
Figure 3 Flow Cytometric Analysis of Peripheral Blood Immune Cell Proportions in HDM-Challenged Mice. Representative flow cytometry plots and quantified proportions of (A) neutrophils, (B) eosinophils, and (C) monocytes in peripheral blood of PBS-treated mice (n=9) and HDM- (n=14). *P<0.05, **P<0.01. |
Macrophages in Spleen and Alveolar Macrophages Decreased After HDM Challenged
Through our gating strategy (Supplement Figure 1C), it was confirmed that compared to the PBS group, the HDM group showed a decrease in the percentage of RPM and Pre RPM in the spleen of mice (Figure 4A and B). Red pulp macrophages (RPMs) are thought to be tissue-resident macrophages in the spleen, and Pre-RPM has been confirmed to be an intermediate developmental stage in the transition of splenic monocytes to RPM.40 When inflammation occurs in the lungs, splenic monocytes are recruited to the lungs, resulting in a decrease in splenic macrophages differentiated from them.41–43 Therefore, we further performed flow cytometry on lung tissue, and our gating strategy is shown in (Supplement Figure 1D), the results showed that the number of alveolar macrophages decreased due to lung inflammation, which was clearly reflected in the HDM challenged mouse group. However, there was no significant difference in the changes of interstitial macrophages between the two groups (Figure 4C and D). The above results indicate that alveolar macrophages, as a crucial group of macrophages for inflammation control, tissue repair, and host defense, were consumed in the first time in allergic asthma caused by HDM attack on mouse lungs, whereas there was no significant change in the number of interstitial macrophages, as developmental intermediates.
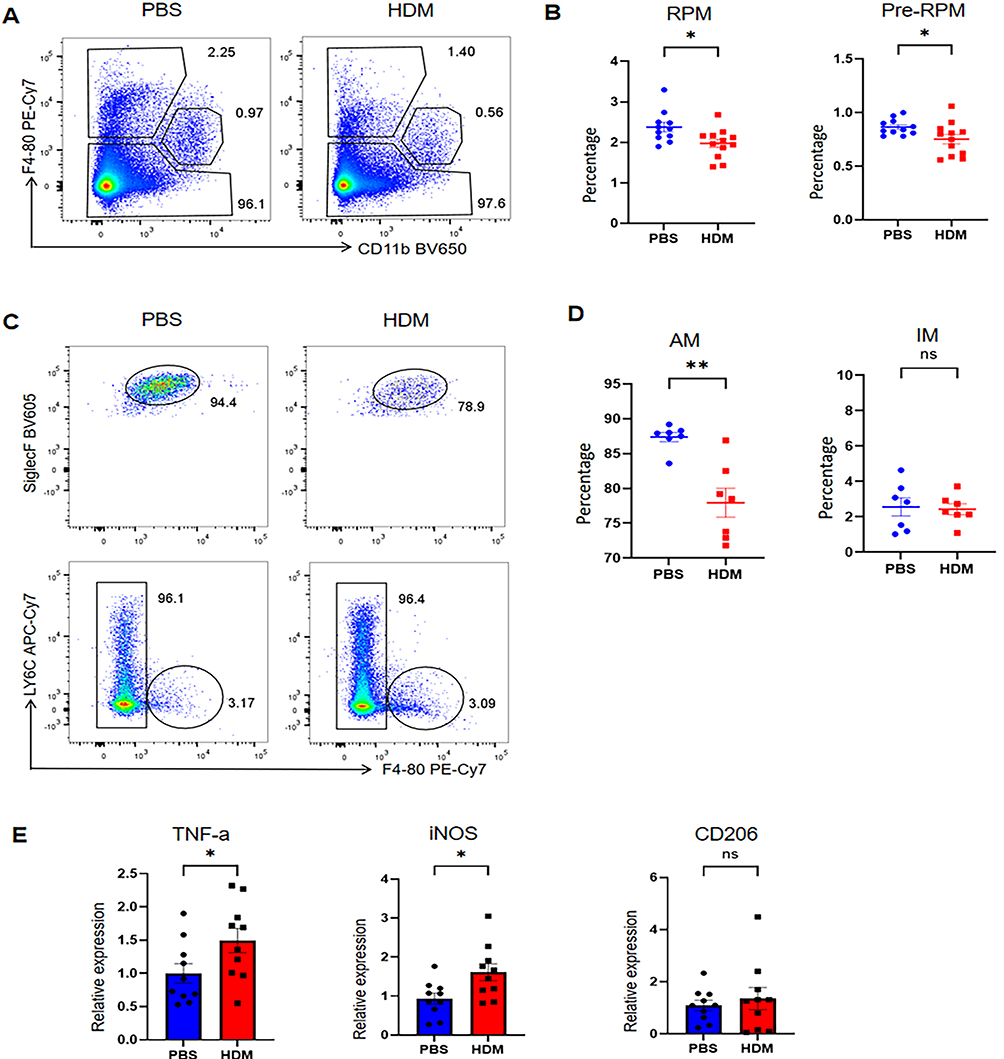 |
Figure 4 Macrophage Subset Proportions and Pro-Inflammatory Gene Expression in HDM-Treated Mice. (A) Representative flow plots and (B) quantified proportions of Red Pulp Macrophages (RPMs) and pre-RPMs in mouse spleens. (C) Representative flow plots and (D) quantified proportions of Alveolar Macrophages (AMs) and Interstitial Macrophages (IMs) in mouse lungs. (E) Relative mRNA expression of TNF-α, iNOS, and CD206 in lung tissues (measured by RT-qPCR; n=10 per group). *P<0.05, **P<0.01, ns: not significant (P≥0.05). |
Increased M1 Phenotype Macrophages in Spleen and Lung of HDM Induced Allergic Asthma Model
Studies have shown that macrophages exhibit different polarization phenotypes in the development of acute and chronic inflammatory lung disease,26 so we wanted to explore which phenotype was predominant in our asthma model. We used RT-PCR to detect the expression levels of TNF-α, iNOS, and CD206 mRNAs in lung tissue. The results showed that compared with the PBS group, the expression levels of TNF-α and iNOS mRNA in lung tissue of the HDM group increased, whereas no significant difference was observed in the expression of CD206 mRNA (Figure 4E).
Then we used UMAP dimensionality reduction analysis with multiple markers to divide red pulp macrophage (RPMs) into three categories. There were differences between the three types of macrophages in the PBS group and HDM group (Figure 5A and B). We found differential expression of iNOS and no significant difference in CD206 (Figure 5C and D). Similarly, we also performed Uniform Manifold Approximation and Projection (UMAP) dimensionality reduction analysis on pre-RPM and found the same thing (Figure 5E–H). We also performed UMAP dimensionality reduction analysis on alveolar macrophages and compared the PBS group and HDM group (Figure 6A). We found that the expression of iNOS was enhanced in the HDM group (Figure 6B), while the expression of CD206 showed a certain weakening trend (Figure 6C). The dimensionality reduction analysis of interstitial macrophages (Figure 6D) showed that the expression of iNOS was also enhanced in HDM (Figure 6E), while the expression of CD206 showed no significant difference (Figure 6F).
 |
Figure 5 UMAP Dimensionality Reduction Analysis of Splenic Macrophage Subsets in Mice. (A) UMAP plots of splenic Red Pulp Macrophage (RPM) subsets from PBS- and HDM-treated mice. (B) Proportional distribution of splenic RPM subpopulations. (C) Heatmap of marker gene expression levels in splenic RPM subpopulations. (D) UMAP plots showing iNOS and CD206 expression intensity in splenic RPM subsets. (E) UMAP plots of splenic pre-RPM subsets from PBS- and HDM-treated mice. (F) Proportional distribution of splenic pre-RPM subpopulations. (G) Heatmap of marker gene expression levels in splenic pre-RPM subpopulations. (H) UMAP plots showing iNOS and CD206 expression intensity in splenic pre-RPM subsets. |
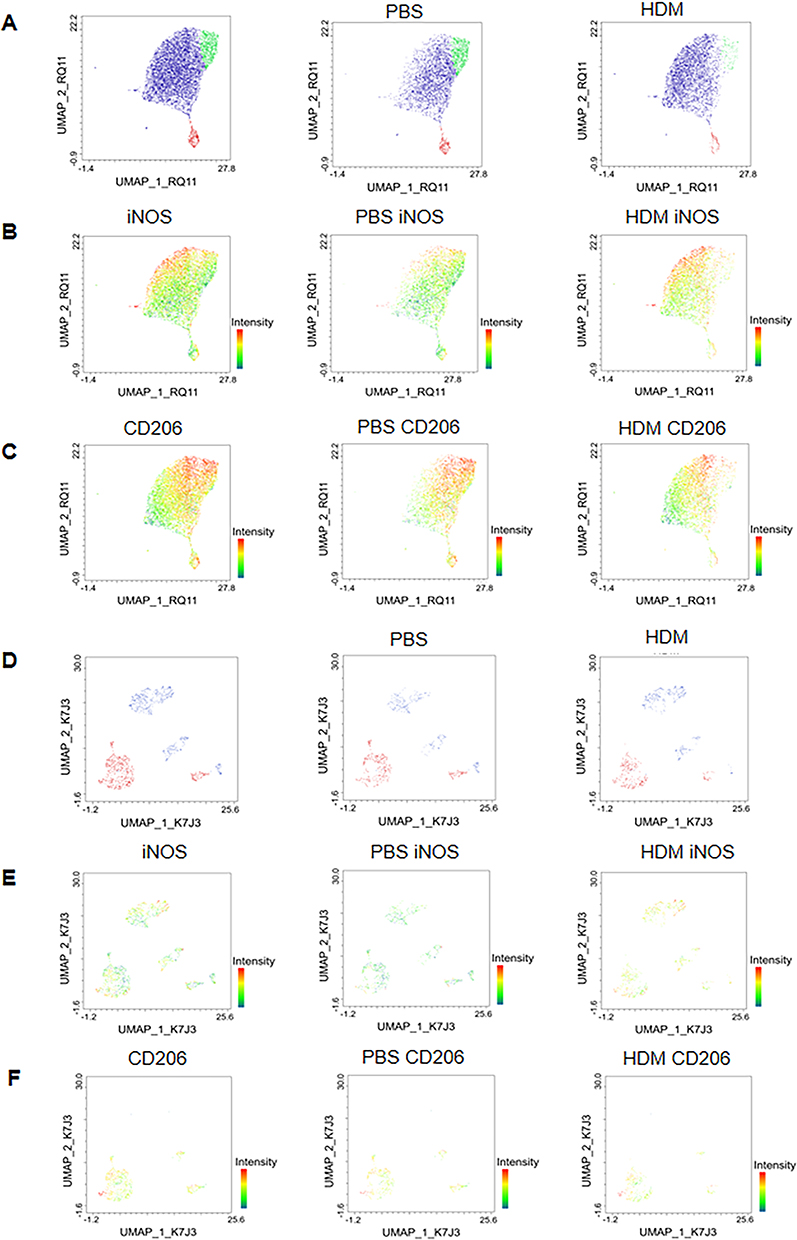 |
Figure 6 UMAP Dimensionality Reduction Analysis of pulmonary Macrophages Subsets in Mice. (A) UMAP plots of alveolar macrophage subsets from PBS- and HDM-treated mice. (B and C) UMAP plots showing iNOS and CD206 expression intensity in alveolar macrophage subsets of PBS- and HDM-treated mice, respectively. (D) UMAP plots of interstitial macrophage subsets from PBS- and HDM-treated mice. (E and F) UMAP plots showing iNOS and CD206 expression intensity in interstitial macrophage subsets of PBS- and HDM-treated mice, respectively. |
Subsequently, we used conventional flow assays to detect the polarization of macrophages in splenic and lung tissues. The results showed that the expression of iNOS in RPMs and Rp RPMs of the spleen was significantly enhanced in the HDM group (Figure 7A and B), while there was no significant difference in CD206 (Figure 7C and D). Similarly, iNOS expression was also enhanced in AMs and IMs of the lungs after HDM challenge (Figure 7E and F), while CD206 showed no trend of change (Figure 7G and H).
 |
Figure 7 M1 Polarization of Splenic and Pulmonary Macrophages in HDM-Treated Mice. (A) Representative flow plots and (B) quantified mean fluorescence intensity (MFI) of iNOS expression in splenic macrophages (Pre-RPMs, RPMs). (C) Representative flow plots and (D) quantified MFI of CD206 expression in splenic macrophages. (E) Representative flow plots and (F) quantified MFI of iNOS expression in pulmonary macrophages (AMs, IMs). (G) Representative flow plots and (H) quantified MFI of CD206 expression in pulmonary macrophages. *P<0.05, **P<0.01,***P<0.001, ns: not significant (P≥0.05). |
The above results suggest that HDM-induced allergic asthma moves towards a proinflammatory macrophage phenotype with increased M1 polarization.
Enhanced Macrophage Ferritinophagy in HDM Induced Asthma Model
We have previously confirmed that macrophages undergo M1 polarization in an acute asthma model. Therefore, we aim to further explore whether, during the process of M1 polarization, macrophages trigger enhanced ferritinophagy due to changes in metabolic demands and stress status, thereby maintaining the pro-inflammatory dominance of M1-type macrophages during the peak phase of acute asthmatic inflammation. We isolated alveolar macrophages from mouse bronchoalveolar lavage fluid (BALF) and confirmed their purity to be over 98% using flow cytometry (Supplement Figure 2A). Therefore, we used the same method to isolate alveolar macrophages and performed immunofluorescence assays to investigate the co-localization of important ferritinophagy-related proteins in cells. NCOA4 acts as a selective autophagy receptor and is an important regulator of ferritinophagy, mediating the transport of intracellular ferritin to lysosomes for degradation.31,44 LC3B is a recognized indicator of autophagy and marks the formation of autophagosomes. We found that HDM treatment increased the co-localization of the autophagic marker LC3B (green) and NCOA4 (red) in alveolar macrophages (Figure 8A and B). Meanwhile, compared with the PBS group, the HDM group also showed an increased proportion of co-localization of LC3B (green) and FTH1 (main components of ferritin, red) (Figure 8C and D). Due to the transport of ferritin to lysosomes for degradation, we also investigated the co-localization of ferritin and lysosomes. FTH1 and LAMP1 (lysosome labeled, green fluorescence) were used to fluorescently stain alveolar macrophages, and the results showed that the HDM group had more co-localization points (Figure 8E and F). In addition, we also found that the mRNA expression of NCOA4 and LC3B in lung tissue was enhanced in the HDM group (Supplement Figure 2B). These experimental data indicate that the ferritinophagy regulation of macrophages is involved in HDM-induced allergic asthma.
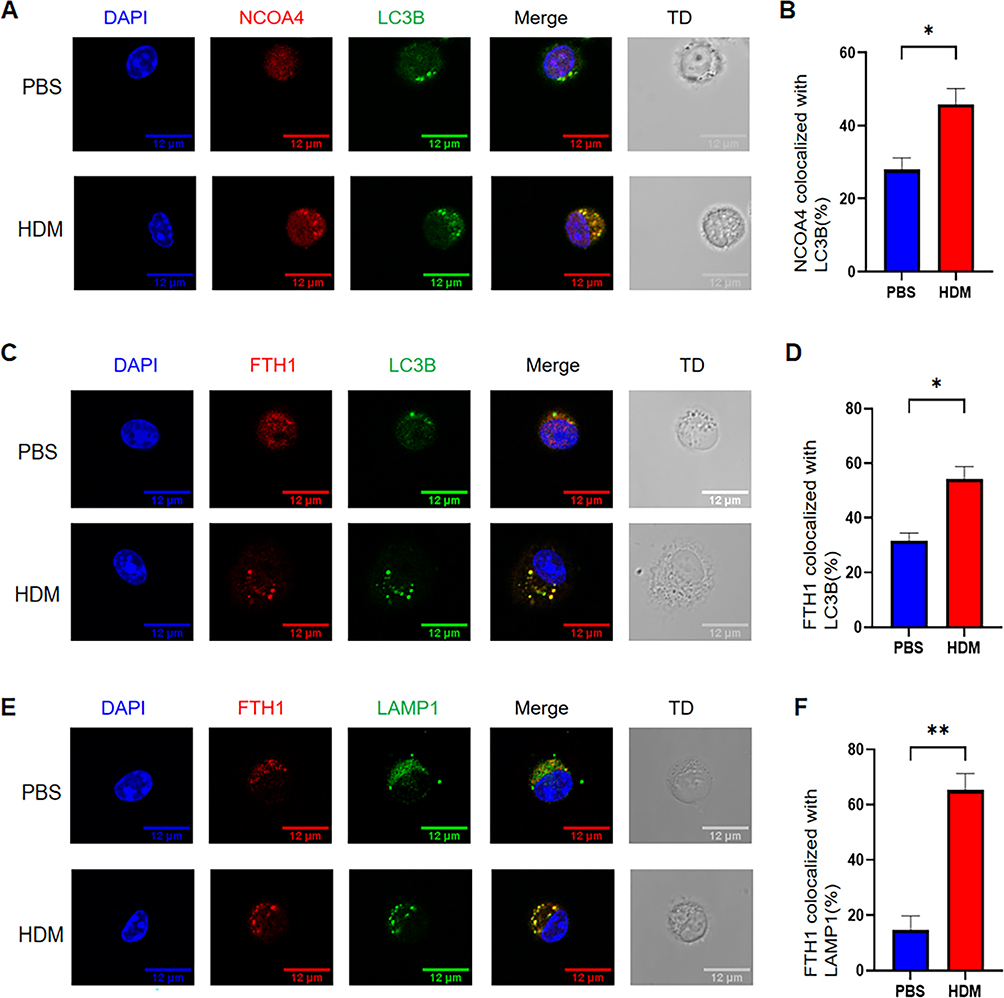 |
Figure 8 Colocalization of Ferritinophagy-Related Proteins in Alveolar Macrophages. Representative confocal images (scale bars: 12 μm) showing colocalization (merge) of (A) NCOA4 (red) and LC3B (green), (C) FTH1 (red) and LC3B (green), and (E) FTH1 (red) and LAMP1 (green) in alveolar macrophages from PBS- and HDM-treated mice; TD: transmitted light. Quantified percentages of alveolar macrophages with (B) NCOA4-LC3B colocalization, (D) FTH1-LC3B colocalization, and (F) FTH1-LAMP1 colocalization. Results are presented as mean ± SEM. *P<0.05, **P<0.01. |
To further validate the changes in ferritinophagy-related proteins in the HDM-induced asthma model, we used flow cytometry to detect alterations in LC3B and FTH1 in mouse spleen and lung macrophages. Our flow cytometry analysis found that the flow representation and frequency statistics of LC3B expression in macrophages in both the spleen and lung were enhanced after HDM induction (Figure 9A and B). Meanwhile, the expression of ferritin heavy chain 1 (FTH1), a major iron storage protein, was decreased in both spleen and lung macrophages (Figure 9C and D). Additionally, after HDM induction, macrophages showed M1 polarization, and we speculated that ferritinophagy might be involved in the process of regulating polarization homeostasis in macrophages.
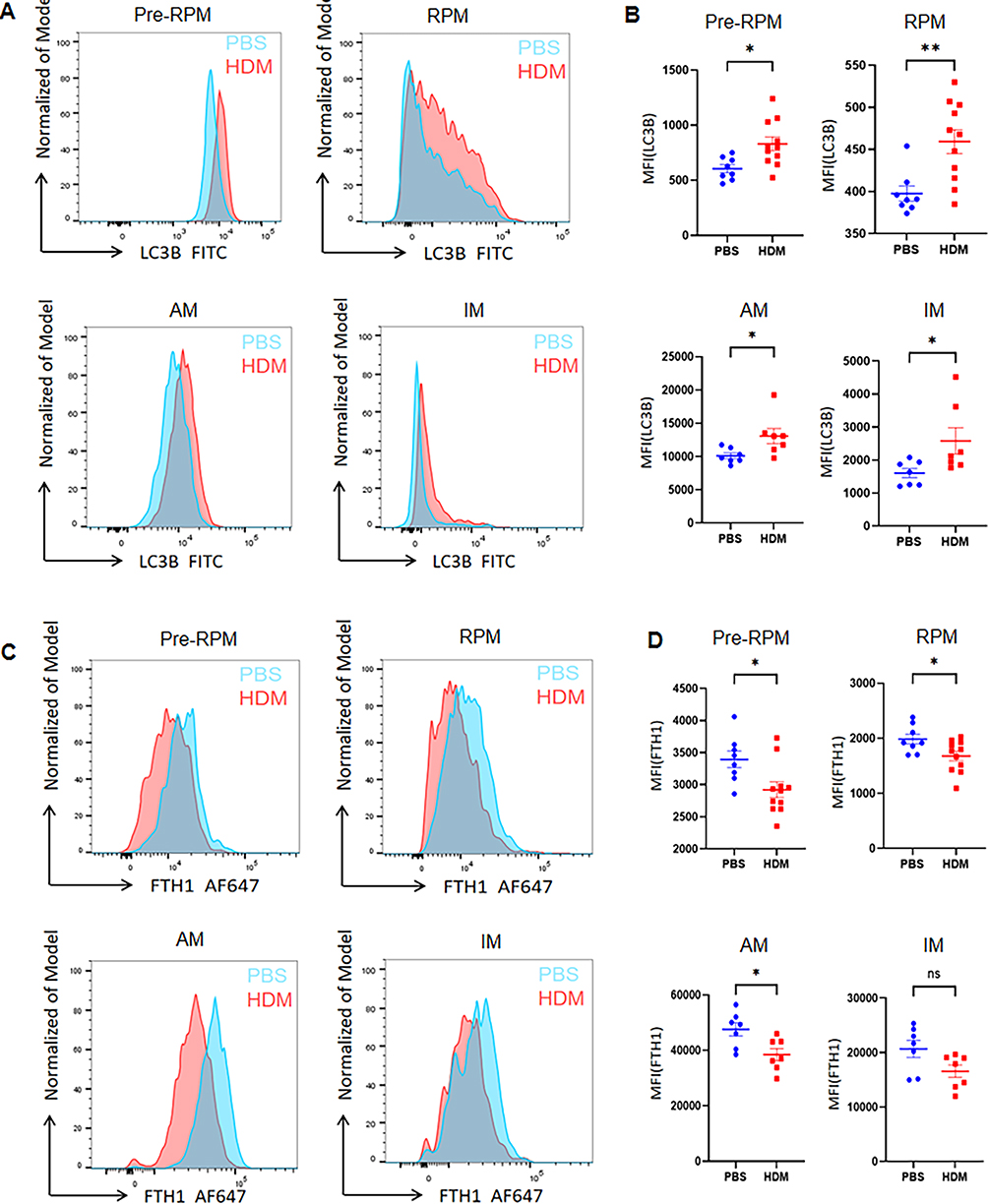 |
Figure 9 Enhanced Ferritinophagy in Splenic and Pulmonary Macrophages of HDM-Treated Mice. (A) Representative flow plots and (B) quantified mean fluorescence intensity (MFI) of LC3B expression in splenic (Pre-RPMs, RPMs) and pulmonary (AMs, IMs) macrophages. (C) Representative flow plots and (D) quantified MFI of FTH1 expression in splenic and pulmonary macrophages. *P<0.05, **P<0.01, ns: not significant (P≥0.05). |
Discussion
This study, integrating clinical observations with experimental models, reveals a potential association between ferritinophagy and M1 macrophage polarization during the acute phase of childhood allergic asthma. It is important to emphasize that our findings primarily establish a correlation, not a causal relationship, between enhanced ferritinophagy and M1 polarization, providing a new perspective for understanding asthma immunopathology.
The translational of our work is evidenced by consistent monocyte/macrophage alterations across species. We confirmed HDM as the predominant allergen in asthmatic children, consistent with epidemiological studies,45–47 and successfully established a corresponding mouse model replicating key clinical features including airway hyperresponsiveness and elevated IgE. More importantly, the increased classical monocyte proportion in children’s PBMCs paralleled monocyte changes in HDM-exposed mice. Circulating classical monocytes increase and migrate under inflammatory conditions, and are recruited to diseased tissues.38 Previous studies have shown differential expression of macrophage scavenger receptor 1 (MSR1) as a macrophage precursor in monocytes of peripheral blood mononuclear cells (PBMCs) from allergic asthma patients.48 Compared with non-asthmatic individuals, asthma patients exhibit increased proportions of classical monocytes and neutrophils, suggesting co-recruitment of monocytes and neutrophils to the airways of asthma patients.37 These findings indicate that asthma influences monocyte alterations. Our results reinforce the monocyte-macrophage axis as the core of asthma athogenesis.
Our investigation revealed coordinated alterations in macrophage distribution and polarization. In the literature that spleen reservoir monocytes readily leave the spleen and migrate to inflamed tissues (eg, lungs and heart) during acute inflammation.49,50 This results in a decrease in the number of pre-RPMs and RPMs differentiated from splenic monocytes.40 Our experimental results also indicate that spleen macrophages are reduced in asthmatic mice compared to the control group, suggesting that spleen monocytes also migrate out of the spleen tissue during acute asthma episodes. Furthermore, our findings reveal a significant reduction in alveolar macrophages (AMs) in the asthma group, while the number of interstitial macrophages (IMs) shows no significant change in the results. Fan et al showed that alveolar macrophage pyroptosis and necrotic apoptosis have important effects on the progression of lung inflammation.51 In early inflammation, recruited monocyte-derived alveolar macrophages exhibit strong accumulation, followed by a gradual decrease in their numbers.52 Whereas interstitial macrophages (IMs), which are long-lived and slow to self-renew, show slower turnover rates and are replenished by circulating monocytes.53,54 Our asthma model was created in a way that better fits the acute attacks of asthma with six consecutive excitations of the HDM, and the analytical assay performed on the seventh day; Therefore, alveolar macrophages—a key macrophage population crucial for inflammation control, tissue repair, and host defense—were significantly depleted in the initial phase, while the number of interstitial macrophages, serving as developmental intermediates, showed no significant change. Most significantly, we observed predominant M1 polarization in both splenic and lung macrophages, characterized by upregulated iNOS without corresponding M2 marker (CD206) changes. This M1 biased response may play a dominant role in the exacerbation of inflammation in the acute phase of asthma, directly exacerbating airway inflammation and tissue damage by promoting the release of pro-inflammatory cytokines and recruitment of inflammatory cells. We believe that the lack of significant changes in M2 markers may reflect the immune characteristics of asthma in the acute phase, where pro-inflammatory mechanisms dominate, while the anti-inflammatory and tissue repair functions of M2 macrophages may be more significant in the chronic or remission phase.
To further validate the biological significance and robustness of our key findings, we quantified the effect sizes (Cohen’s d) and their 95% confidence intervals for core indicators, including monocyte subsets, macrophage proportions, polarization markers, and ferritinophagy-related proteins, as summarized in Supplementary Table 3. Notably, the percentage of alveolar macrophages (AM) between HDM-challenged and PBS control mice exhibited a very large effect size (Cohen’s d=2.30), while M1 polarization markers (eg, RPM iNOS, IM iNOS) and ferritinophagy-related proteins (eg, AM FTH1, RPM LC3B) showed large to very large effect sizes. These results confirm that the observed phenotypic and functional alterations in macrophages are not only statistically significant but also biologically impactful, reinforcing the reliability of our conclusions regarding M1 polarization during acute allergic asthma.
The most innovative aspect of our study concerns ferritinophagy activation in asthmatic macrophages. We demonstrated enhanced ferritinophagy through multiple lines of evidence: increased NCOA4-LC3B colocalization, elevated LC3B expression, reduced FTH1 levels, and enhanced FTH1-LAMP1 association. The concurrent ferritinophagy enhancement and M1 polarization in the same macrophage populations suggests a potential mechanistic link, though whether ferritinophagy drives polarization or represents a parallel process remains unclear. Notably, the non-IgE-mediated immunoreactivity to HDM and other common allergens, a key feature of asthma endotypes previously classified as “intrinsic”, highlights the need to integrate iron metabolism and macrophage polarization signatures into precision medicine strategies for asthma subtyping and personalized treatment.55,56 To address this critical knowledge gap, future studies should employ definitive functional approaches including NCOA4 knockdown models and specific ferritinophagy inhibitors to establish causality and delineate the precise molecular mechanisms connecting ferritinophagy to macrophage polarization.
Notably, this study has certain limitations that merit consideration. First, the relatively small clinical cohort (16 asthmatic children vs 15 controls) may restrict statistical power and the generalizability of our findings, which we plan to address through multi-center collaboration and larger-sample validation in future research. Second, potential confounders including children’s dietary iron intake, systemic iron status, and concurrent systemic inflammation were not controlled in the current study; these factors will be rigorously accounted for in subsequent investigations to further validate the observed associations between ferritinophagy and M1 macrophage polarization. Third, while group sizes were based on established models and post hoc analysis indicated sufficient statistical power, a formal a priori power calculation was not performed. Future confirmatory studies should include prospective power calculations based on the effect sizes reported here. Fourth, this study focused on ferritinophagy markers but did not quantify their downstream effects (eg, intracellular iron levels, ROS, ferroptosis markers), and mechanistic experiments (eg, NCOA4 knockdown, inhibitor intervention) are lacking. These aspects render the findings hypothesis-generating, and future research will address these gaps to clarify the causal and regulatory relationships between ferritinophagy and M1 polarization.
Conclusion
In summary, based on clinical observations showing an increased proportion of classical monocytes in the peripheral blood of asthmatic children, we established an acute asthma model in young mice using house dust mite (HDM), the allergen with the highest sensitization rate in clinical settings. We demonstrated that during acute asthma attacks, macrophages in the spleen and lungs of model mice exhibited an M1-polarized phenotype, which was accompanied by enhanced ferritinophagy. Our study reveals a novel association between these two processes, thereby laying a foundation for further investigation into their potential causal relationship and specific regulatory mechanisms. These findings may provide new insights for the prevention and treatment of allergic asthma in children.
Abbreviations
HDM, House Dust Mite; PBMCs, Peripheral Blood Mononuclear Cells; FTH1, Ferritin Heavy Chain-1; LC3B, Microtubule-Associated Protein 1 Light Chain 3 Beta; NCOA4, Nuclear Receptor Coactivator 4; LAMP1, Lysosome-Associated Membrane Protein 1; iNOS, Inducible Nitric Oxide Synthase; TNF-α, Tumor Necrosis Factor α; AHR, Airway Hyperresponsiveness; BALF, Bronchoalveolar Lavage Fluid.
Data Sharing Statement
All data supporting the findings from this study are available from the corresponding author upon reasonable request.
Acknowledgments
We appreciate the platform support provided by Wenzhou Medical University and Yuhuan People’s Hospital, and we also thank all participants.
Author Contributions
Jingjing Li: Conceptualization, Methodology, Formal Analysis, Investigation, Data Curation, Writing – Original Draft, Visualization. Xing Wang: Methodology, Validation, Investigation, Data Curation, Writing – Review & Editing. Xiaofeng Shi: Investigation, Resources, Writing – Review & Editing, Data Curation. Yelin Jiang: Investigation, Data Curation, Writing – Review & Editing. Ren Shen: Conceptualization, Resources, Writing – Review & Editing, Supervision, Project Administration, Funding Acquisition. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Zhejiang Province Medical and Health Science and Technology Plan Project (No.: 2023KY1349).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Asher MI, García-Marcos L, Pearce NE, Strachan DP. Trends in worldwide asthma prevalence. Eur Respir J. 2020;56(6):2002094. doi:10.1183/13993003.02094-2020
2. Dong L, Wang Y, Zheng T, et al. Hypoxic hUCMSC-derived extracellular vesicles attenuate allergic airway inflammation and airway remodeling in chronic asthma mice. Stem Cell Res Ther. 2021;12(1):4. doi:10.1186/s13287-020-02072-0
3. Li J, Wang H, Chen Y, Zheng J, Wong GW, Zhong N. House dust mite sensitization is the main risk factor for the increase in prevalence of wheeze in 13- to 14-year-old schoolchildren in Guangzhou city, China. Clin Exp Allergy. 2013;43(10):1171–21. doi:10.1111/cea.12157
4. Casas L, Sunyer J, Tischer C, et al. Early-life house dust mite allergens, childhood mite sensitization, and respiratory outcomes. Allergy. 2015;70(7):820–827. doi:10.1111/all.12626
5. Ballardini N, Bergström A, Wahlgren CF, et al. IgE antibodies in relation to prevalence and multimorbidity of eczema, asthma, and rhinitis from birth to adolescence. Allergy. 2016;71(3):342–349. doi:10.1111/all.12798
6. Hou F, Xiao K, Tang L, Xie L. Diversity of macrophages in lung homeostasis and diseases. Front Immunol. 2021;12:753940. doi:10.3389/fimmu.2021.753940
7. Zawada AM, Rogacev KS, Rotter B, et al. SuperSAGE evidence for CD14++CD16+ monocytes as a third monocyte subset. Blood. 2011;118(12):e50–61. doi:10.1182/blood-2011-01-326827
8. Kapellos TS, Bonaguro L, Gemünd I, et al. Human monocyte subsets and phenotypes in major chronic inflammatory diseases. Front Immunol. 2019;10:2035. doi:10.3389/fimmu.2019.02035
9. Guilliams M, Mildner A, Yona S. Developmental and functional heterogeneity of monocytes. Immunity. 2018;49(4):595–613. doi:10.1016/j.immuni.2018.10.005
10. Chiu S, Bharat A. Role of monocytes and macrophages in regulating immune response following lung transplantation. Curr Opin Organ Transplant. 2016;21(3):239–245. doi:10.1097/mot.0000000000000313
11. Trzebanski S, Kim JS, Larossi N, et al. Classical monocyte ontogeny dictates their functions and fates as tissue macrophages. Immunity. 2024;57(6):1225–1242.e6. doi:10.1016/j.immuni.2024.04.019
12. Menezes S, Melandri D, Anselmi G, et al. The heterogeneity of Ly6C(hi) monocytes controls their differentiation into iNOS(+) macrophages or monocyte-derived dendritic cells. Immunity. 2016;45(6):1205–1218. doi:10.1016/j.immuni.2016.12.001
13. Kuruvilla ME, Lee FE, Lee GB. Understanding asthma phenotypes, endotypes, and mechanisms of disease. Clin Rev Allergy Immunol. 2019;56(2):219–233. doi:10.1007/s12016-018-8712-1
14. Kuo CH, Tsai ML, Li CH, et al. Altered pattern of macrophage polarization as a biomarker for severity of childhood asthma. J Inflamm Res. 2021;14:6011–6023. doi:10.2147/jir.S319754
15. Zhu X, Cui J, Yi L, et al. The role of T cells and macrophages in asthma pathogenesis: a new perspective on mutual crosstalk. Mediators Inflamm. 2020;2020:7835284. doi:10.1155/2020/7835284
16. Saradna A, Do DC, Kumar S, Fu QL, Gao P. Macrophage polarization and allergic asthma. Transl Res. 2018;191:1–14. doi:10.1016/j.trsl.2017.09.002
17. Shapouri-Moghaddam A, Mohammadian S, Vazini H, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. 2018;233(9):6425–6440. doi:10.1002/jcp.26429
18. Han X, Liu L, Huang S, et al. RNA m(6)A methylation modulates airway inflammation in allergic asthma via PTX3-dependent macrophage homeostasis. Nat Commun. 2023;14(1):7328. doi:10.1038/s41467-023-43219-w
19. Luo M, Zhao F, Cheng H, Su M, Wang Y. Macrophage polarization: an important role in inflammatory diseases. Front Immunol. 2024;15:1352946. doi:10.3389/fimmu.2024.1352946
20. Naura AS, Zerfaoui M, Kim H, et al. Requirement for inducible nitric oxide synthase in chronic allergen exposure-induced pulmonary fibrosis but not inflammation. J Immunol. 2010;185(5):3076–3085. doi:10.4049/jimmunol.0904214
21. Chang C, Chen G, Wu W, et al. Exogenous IL-25 ameliorates airway neutrophilia via suppressing macrophage M1 polarization and the expression of IL-12 and IL-23 in asthma. Respir Res. 2023;24(1):260. doi:10.1186/s12931-023-02557-5
22. Lee SC, Lee YJ, Choi I, Kim M, Sung JS. CXCL16/CXCR6 axis in adipocytes differentiated from human adipose derived mesenchymal stem cells regulates macrophage polarization. Cells. 2021;10(12):3410. doi:10.3390/cells10123410
23. Moreira AP, Cavassani KA, Hullinger R, et al. Serum amyloid P attenuates M2 macrophage activation and protects against fungal spore-induced allergic airway disease. J Allergy Clin Immunol. 2010;126(4):712–721.e7. doi:10.1016/j.jaci.2010.06.010
24. Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002;23(11):549–555. doi:10.1016/s1471-4906(02)02302-5
25. Zhou Y, Do DC, Ishmael FT, et al. Mannose receptor modulates macrophage polarization and allergic inflammation through miR-511-3p. J Allergy Clin Immunol. 2018;141(1):350–364.e8. doi:10.1016/j.jaci.2017.04.049
26. Lee JW, Chun W, Lee HJ, et al. The role of macrophages in the development of acute and chronic inflammatory lung diseases. Cells. 2021;10(4):897. doi:10.3390/cells10040897
27. Locati M, Curtale G, Mantovani A. Diversity, mechanisms, and significance of macrophage plasticity. Annu Rev Pathol. 2020;15:123–147. doi:10.1146/annurev-pathmechdis-012418-012718
28. Shen R, Yang S, Zhou Y, Li J. Analysis of the characteristics of blood inflammatory cytokines and their influencing factors in acute exacerbations of allergic asthma in children. Front Pediatr. 2025;13:1571556. doi:10.3389/fped.2025.1571556
29. Zeng Z, Huang H, Zhang J, et al. HDM induce airway epithelial cell ferroptosis and promote inflammation by activating ferritinophagy in asthma. FASEB J. 2022;36(6):e22359. doi:10.1096/fj.202101977RR
30. Hwang HJ, Ha H, Lee BS, Kim BH, Song HK, Kim YK. LC3B is an RNA-binding protein to trigger rapid mRNA degradation during autophagy. Nat Commun. 2022;13(1):1436. doi:10.1038/s41467-022-29139-1
31. He J, Li Z, Xia P, et al. Ferroptosis and ferritinophagy in diabetes complications. Mol Metab. 2022;60:101470. doi:10.1016/j.molmet.2022.101470
32. Santana-Codina N, Mancias JD. The role of NCOA4-mediated ferritinophagy in health and disease. Pharmaceuticals. 2018;11(4):114. doi:10.3390/ph11040114
33. Cheng XT, Xie YX, Zhou B, Huang N, Farfel-Becker T, Sheng ZH. Revisiting LAMP1 as a marker for degradative autophagy-lysosomal organelles in the nervous system. Autophagy. 2018;14(8):1472–1474. doi:10.1080/15548627.2018.1482147
34. Zhao J, Dar HH, Deng Y, et al. PEBP1 acts as a rheostat between prosurvival autophagy and ferroptotic death in asthmatic epithelial cells. Proc Natl Acad Sci U S A. 2020;117(25):14376–14385. doi:10.1073/pnas.1921618117
35. Hammad H, Plantinga M, Deswarte K, et al. Inflammatory dendritic cells--not basophils--are necessary and sufficient for induction of Th2 immunity to inhaled house dust mite allergen. J Exp Med. 2010;207(10):2097–2111. doi:10.1084/jem.20101563
36. Morris CR, Habibovic A, Dustin CM, et al. Macrophage-intrinsic DUOX1 contributes to type 2 inflammation and mucus metaplasia during allergic airway disease. Mucosal Immunol. 2022;15(5):977–989. doi:10.1038/s41385-022-00530-x
37. Niessen NM, Baines KJ, Simpson JL, et al. Neutrophilic asthma features increased airway classical monocytes. Clin Exp Allergy. 2021;51(2):305–317. doi:10.1111/cea.13811
38. Mai CL, Tan Z, Xu YN, et al. CXCL12-mediated monocyte transmigration into brain perivascular space leads to neuroinflammation and memory deficit in neuropathic pain. Theranostics. 2021;11(3):1059–1078. doi:10.7150/thno.44364
39. Huang C, Wang J, Zheng X, et al. Activation of TLR signaling in sensitization-recruited inflammatory monocytes attenuates OVA-induced allergic asthma. Front Immunol. 2018;9:2591. doi:10.3389/fimmu.2018.02591
40. Lu Y, Basatemur G, Scott IC, et al. Interleukin-33 signaling controls the development of iron-recycling macrophages. Immunity. 2020;52(5):782–793.e5. doi:10.1016/j.immuni.2020.03.006
41. Francis M, Sun R, Cervelli JA, et al. Editor’s highlight: role of spleen-derived macrophages in ozone-induced lung inflammation and injury. Toxicol Sci. 2017;155(1):182–195. doi:10.1093/toxsci/kfw192
42. Hsiao HM, Fernandez R, Tanaka S, et al. Spleen-derived classical monocytes mediate lung ischemia-reperfusion injury through IL-1β. J Clin Invest. 2018;128(7):2833–2847. doi:10.1172/jci98436
43. Mu X, Li Y, Fan GC. Tissue-resident macrophages in the control of infection and resolution of inflammation. Shock. 2021;55(1):14–23. doi:10.1097/shk.0000000000001601
44. Wang J, Zhu Q, Li R, Zhang J, Ye X, Li X. YAP1 protects against septic liver injury via ferroptosis resistance. Cell Biosci. 2022;12(1):163. doi:10.1186/s13578-022-00902-7
45. Carter PM, Peterson EL, Ownby DR, Zoratti EM, Johnson CC. Relationship of house-dust mite allergen exposure in children’s bedrooms in infancy to bronchial hyperresponsiveness and asthma diagnosis by age 6 to 7. Ann Allergy Asthma Immunol. 2003;90(1):41–44. doi:10.1016/s1081-1206(10)63612-5
46. Li Z, Zheng N, An Q, et al. Impact of environmental factors and bacterial interactions on dust mite allergens in different indoor dust. Sci Total Environ. 2022;844:157177. doi:10.1016/j.scitotenv.2022.157177
47. Jurkiewicz K, Jutel M, Smolinska S. Update on HDM allergy: principal changes over the years. Int J Mol Sci. 2025;26(12):5660. doi:10.3390/ijms26125660
48. Baos S, Cremades-Jimeno L, López-Ramos M, et al. Expression of Macrophage Scavenger Receptor (MSR1) in peripheral blood cells from patients with different respiratory diseases: beyond monocytes. J Clin Med. 2022;11(5):1439. doi:10.3390/jcm11051439
49. Swirski FK, Nahrendorf M, Etzrodt M, et al. Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science. 2009;325(5940):612–616. doi:10.1126/science.1175202
50. Sabatel C, Radermecker C, Fievez L, et al. Exposure to bacterial CpG DNA protects from airway allergic inflammation by expanding regulatory lung interstitial macrophages. Immunity. 2017;46(3):457–473. doi:10.1016/j.immuni.2017.02.016
51. Fan EKY, Fan J. Regulation of alveolar macrophage death in acute lung inflammation. Respir Res. 2018;19(1):50. doi:10.1186/s12931-018-0756-5
52. Janssen WJ, Barthel L, Muldrow A, et al. Fas determines differential fates of resident and recruited macrophages during resolution of acute lung injury. Am J Respir Crit Care Med. 2011;184(5):547–560. doi:10.1164/rccm.201011-1891OC
53. Schyns J, Bai Q, Ruscitti C, et al. Non-classical tissue monocytes and two functionally distinct populations of interstitial macrophages populate the mouse lung. Nat Commun. 2019;10(1):3964. doi:10.1038/s41467-019-11843-0
54. Gibbings SL, Thomas SM, Atif SM, et al. Three unique interstitial macrophages in the murine lung at steady state. Am J Respir Cell Mol Biol. 2017;57(1):66–76. doi:10.1165/rcmb.2016-0361OC
55. Olivier CE, Pinto DG, Teixeira APM, et al. Evaluating Non-IgE-Mediated Allergens’ immunoreactivity in patients formerly classified as “Intrinsic” asthmatics with help of the leukocyte adherence inhibition test. Eur J Clin Med. 2023;4(2):238. doi:10.24018/ejclinicmed.2023.4.2.238
56. Jutel M, Agache I, Zemelka-Wiacek M, et al. Nomenclature of allergic diseases and hypersensitivity reactions: adapted to modern needs: an EAACI position paper. Allergy. 2023;78(11):2851–2874. doi:10.1111/all.15889
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
IL-27 Aggravates Sepsis-Induced ARDS by Driving Macrophage Ferroptosis Through Activation of NCOA4-Mediated Ferritinophagy
Guo M, Xiong M, Shi J, Peng Q, Lin S, Xu F
Journal of Inflammation Research 2026, 19:575933
Published Date: 3 March 2026