Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 17
Assessment of Reference Genes Stability in Cortical Bone of Obese and Diabetic Mice
Authors Ai Y ![]() , Peng K, Li C, Zhang J
, Peng K, Li C, Zhang J ![]() , Wang G, Wang B
, Wang G, Wang B ![]() , Huang E
, Huang E
Received 6 December 2023
Accepted for publication 15 February 2024
Published 2 March 2024 Volume 2024:17 Pages 1081—1091
DOI https://doi.org/10.2147/DMSO.S453458
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Yuanli Ai,1,* Kun Peng,2,* Chunli Li,1,* Jun Zhang,1 Gang Wang,3,4 Bin Wang,1 Enyi Huang3,4
1Institute of Life Sciences, College of Basic Medicine, Chongqing Medical University, Chongqing, People’s Republic of China; 2Chongqing Medical and Pharmaceutical College, Chongqing, People’s Republic of China; 3Chongqing Key Laboratory of Oral Diseases and Biomedical Sciences, Stomatological Hospital of Chongqing Medical University, Chongqing, People’s Republic of China; 4Chongqing Municipal Key Laboratory of Oral Biomedical Engineering of Higher Education College of Stomatology, College of Stomatology, Chongqing Medical University, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Bin Wang; Enyi Huang, Email [email protected]; [email protected]
Introduction: Bone, a pivotal structural organ, is susceptible to disorders with profound health implications. The investigation of gene expression in bone tissue is imperative, particularly within the context of metabolic diseases such as obesity and diabetes that augment the susceptibility to bone fractures. The objective of this study is to identify a set of internal control genes for the analysis of gene expression.
Methods: This study employs reverse transcription-quantitative real-time polymerase chain reaction (RT-qPCR) to assess gene expression in bone tissue. We selected fourteen housekeeping genes and assessed their stability in the cortical bone of mouse models for obesity and diabetes using four well-established algorithms (GeNorm, BestKeeper, NormFinder, and the comparative Delta Ct method).
Results and Conclusion: We identified Rpl13a as the mostly stably expressed reference gene in cortical bone tissue from mouse models of obesity and diabetes (db/db), while Gapdh was found to be the most stable reference gene in another diabetes model, KKAy mice. Additionally, Ef1a, Ppia, Rplp0, and Rpl22 were identified as alternative genes suitable for normalizing gene expression in cortical bone from obesity and diabetes mouse models. These findings enhance RT-qPCR accuracy and reliability, offering a strategic guide to select reference gene for studying bone tissue gene expression in metabolic disorders.
Keywords: reference genes, Rpl13a, Gapdh, cortical bone, metabolic disorders
Introduction
Gene expression serves as the pivotal regulator in orchestration of the intricate processes involved in bone morphogenesis, homeostasis, and repair; variations in gene expression led to different bone diseases.1,2 Within the continually remodeling bone tissue, a harmonious interplay of distinct cellular cohorts, notably osteoblasts, osteoclasts, and osteocytes, mediates the synthesis and secretion of an array of extracellular matrix proteins, growth factors, and cytokines.3 This complex process of bone homeostasis is meticulously governed at the transcriptional level, guided by a multifaceted interplay, involving transcription factors, epigenetic modulators, and elaborate signaling pathways. RNA, synthesized from DNA via transcription, is key in gene expression and comes in two forms: protein-coding and noncoding. Protein-coding genes produce messenger RNAs (mRNAs) which are translated into proteins, crucial for bone modeling and remodeling.4,5 The expression of non-coding RNAs, including small, long, and circular types, is also involved in bone development and the pathogenesis of bone-related disorders.6–8 Circular RNAs affect bone formation by regulating specific gene expression.6 MicroRNAs (like miR-30, miR-124, miR-204) and long non-coding RNAs also play significant roles in bone growth and stability.7,8 Dysregulations that disrupt this genetic equilibrium within the skeletal framework can engender a spectrum of skeletal disorders, encompassing conditions such as osteoporosis, osteoarthritis, and osteogenic neoplasms.9 Hence, delving into the nuanced complexities of gene regulation in bone is crucial for the pursuit of pioneering therapeutic modalities for these incapacitating disorders. In recent years, an array of sophisticated gene expression analysis methodologies,10 including microarray profiling, RNA sequencing, droplet digital PCR11 and reverse transcription-quantitative real-time PCR (RT-qPCR), have substantially enriched our comprehension of the molecular substratum governing the dynamics of bone development, preservation, and pathological changes, offering the promise of groundbreaking strides in this field.
When the expression levels of genes are low, PCR becomes essential for their detection. Housekeeping genes such as RN18S1, GAPDH, and ACTB (β-actin) are commonly used as reference genes for gene expression analysis in published studies;12 however, their use as reference genes is not always appropriate, as their expression can vary across species, tissue types, cell lines, developmental stages, and in response to experimental treatments.13–16 Moreover, the expression of housekeeping genes can vary among different diseases.
The prevalence of obesity and diabetes, which are considered significant metabolic diseases, is increasing worldwide.17 Metabolic bone diseases encompass a diverse range of disorders characterized by disrupted skeletal homeostasis; these conditions often exhibit varying patterns of genetic inheritance, resulting in different genetic profiles converging to produce a similar phenotype.4 RT-qPCR is frequently performed to explore the role of alternative genes in these metabolic diseases, and normalization is a crucial factor in obtaining reliable qPCR results.18 Obesity and diabetes are also linked to an increased risk of bone fractures, highlighting the importance of studying gene expression in bone tissue related to these conditions; however, selecting suitable reference genes can be challenging and time-consuming, and use of a panel of reference genes is often necessary to increase the reliability of results. As rodents, such as mice, serve as models for humans in many experiments, evaluating the stability of reference genes in murine models is crucial. High-fat diet-induced obese mice, along with KKAy and db/db strains, are key models for diabetes research. These models are essential for investigating bone health and skeletal disorders, as both show notable bone quality impairments. It is necessary to evaluate the stability of reference genes in the cortical bone of three murine models of metabolic disorders.19 A previous study evaluated the stability of reference genes in human trabecular bone and provided the optimum choice for human bone disease;20 however, similar studies in cortical bone from mice fed with standard diet, or from mouse models of metabolic disorders, have not been reported. As different conclusions are obtained using different reference genes,21,22 it is necessary to screen for a set of stable internal control genes in mice for gene expression analysis.
In our study, we aimed to evaluate the stabilities of 14 selected housekeeping genes in three mouse models of general metabolic conditions and to identify a set of internal control genes for the analysis of gene expression. Our findings will provide a reference for research using these models and will save researchers time and expense.
Materials and Methods
Sample Preparation
Seven-week-old KK-Ay and wild-type mice (n = 8 per group) were purchased from Beijing HFK Bioscience Co., Ltd, and eight-week-old db/db mice and littermate controls were obtained from GemPharmatech Co., Ltd. Mice were housed in a specific pathogen-free room, and provided with standard diet and free access to water. Rooms were maintained at a suitable temperature and humidity, with a 12 h/12 h light/dark cycle. KKAy and db/db mice and their age and gender matched controls were kept for 18 weeks and 12 weeks, respectively; both of these mouse strains have a typical diabetic phenotype.23
To observe the influence of obesity in bone tissue, a control group and obese mice, fed with a high-fat diet for 9 weeks (referred to as HFD mice), were kept under the same environment described above for KKAy and db/db mice. After euthanasia, humerus bones were collected and soft tissue removed. All animal experiments were approved by the Ethics Committee of the Chongqing Medical University (IACUC-CQMU-2023-0036, approved on 16 March 2023), adhered to the Guidelines for the ethical review of laboratory animal welfare People’s Republic of China National Standard GB/T 35892–2018.
cDNA Synthesis
Cortical bones of humeri without bone marrow were acquired by centrifugation,24 after cutting off the epiphyseal plates from both ends, and homogenized in a tube containing 1 mL of TRIzol reagent (Thermo Fischer Scientific, Waltham, MA, USA). Total RNA extraction was then performed according to the standard protocol.25 The RNA was extracted by adding 200 μL of chloroform to each sample, mixing it, and incubating for 3 minutes. After centrifugation at 12,000 rpm for 15 minutes at 4°C, the supernatant was mixed with isopropanol and left to rest for 10 minutes before being centrifuged again. The resulting precipitate was washed three times with 75% ethanol and resuspended in DEPC-treated water.
RNA was quantified using a Nanophotometer (IMPLEN). The RNA quality control was followed the previous method.26 A260: A280 ratios were between 1.8 and 2.0. After extraction, 1.5 μg aliquots of total RNA were used for cDNA synthesis, following the standard protocol for the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific) in a final volume of 20 μL, by incubation for 5 min at 25°C, 60 min at 42°C, and 5 min at 70°C, followed by 5 min at 65°C.
RT-qPCR
RT-qPCR was performed using QuantStudio5 (Applied Biosystems, Thermo Fischer Scientific) with SYBR Green. Reaction mixtures contained PowerUp SYBR Green Master Mix (Thermo Fischer Scientific), 0.5 μM of forward and reverse primers (Table 1), and 0.25 μL of cDNA, in a final volume of 10 μL. Reactions were carried out in MicroAmp1 Optical 384-well plates (Thermo Fischer Scientific), using the following program: 2 min at 50°C and 10 min at 95°C, followed by 40 cycles of 15s at 95°C and 1 min at 60°C, and dissociation curves were plotted. From the fourteen candidate reference genes, Rpl22, Hprt, Atpf1, Ubc, Ppia primer pairs were obtained from the literature.27 The rest of primers were designed using Primerbank and verified using the BLAST tool, or acquired from previous publications.28,29
 |
Table 1 Primer Sequences of Candidate Genes in Cortical Bone Tissue of Humeri in Mice |
Statistical Analysis
For the physiological analyses of mice, unpaired student’s t-test was used for comparisons between two different groups. The confidence interval was 95%. Four algorithms previously described in the literature were used to select the most suitable reference genes for analysis of bone tissues from HFD, KK-Ay and db/db mice: GeNorm,29 BestKeeper,30 NormFinder,31 and the comparative Delta Ct method.32 All analyses were conducted using the web-based RefFinder tool33 (http://blooge.cn/RefFinder).
Results
To obtain suitable housekeeping genes for analysis of the expression of target genes in cortical bone tissue using RT-qPCR, we established three frequently used mouse models of metabolism: KKAy, db/db, and HFD. Primers to amplify candidate genes were chosen from published studies after a review of investigations that included reference gene selection.
Stable Reference Gene Selection in Cortical Bone from Wild-Type Mice
Mice are frequently used as animal models in medical research.19 We first analyzed the stability of candidate genes in 20–25-week-old mice fed with normal diet (Table 2), by determining the expression levels of each candidate gene in bone samples (Figure 1A). The CT value of the gray line was 15, while that of the red line was 33; values outside of these lines were considered to indicate unreliable low and high expression of candidate genes in mouse cortical bone tissue, respectively. As shown in Figure 1B, stability of housekeeping gene expression was comprehensively analyzed in wild type mice. Based on comparative delta Ct, NormFinder, and Bestkeeper data, Rplp0 was the most stably expressed gene. Analysis using the GeNorm algorithm selected Ppia and Gapdh as the best combined group. Further, Gapdh was selected as the best among three commonly used internal control genes (Gapdh, Rn18s, and ActB) by the four algorithms. ActB and B2m emerged as the least stable among the evaluated candidate genes.
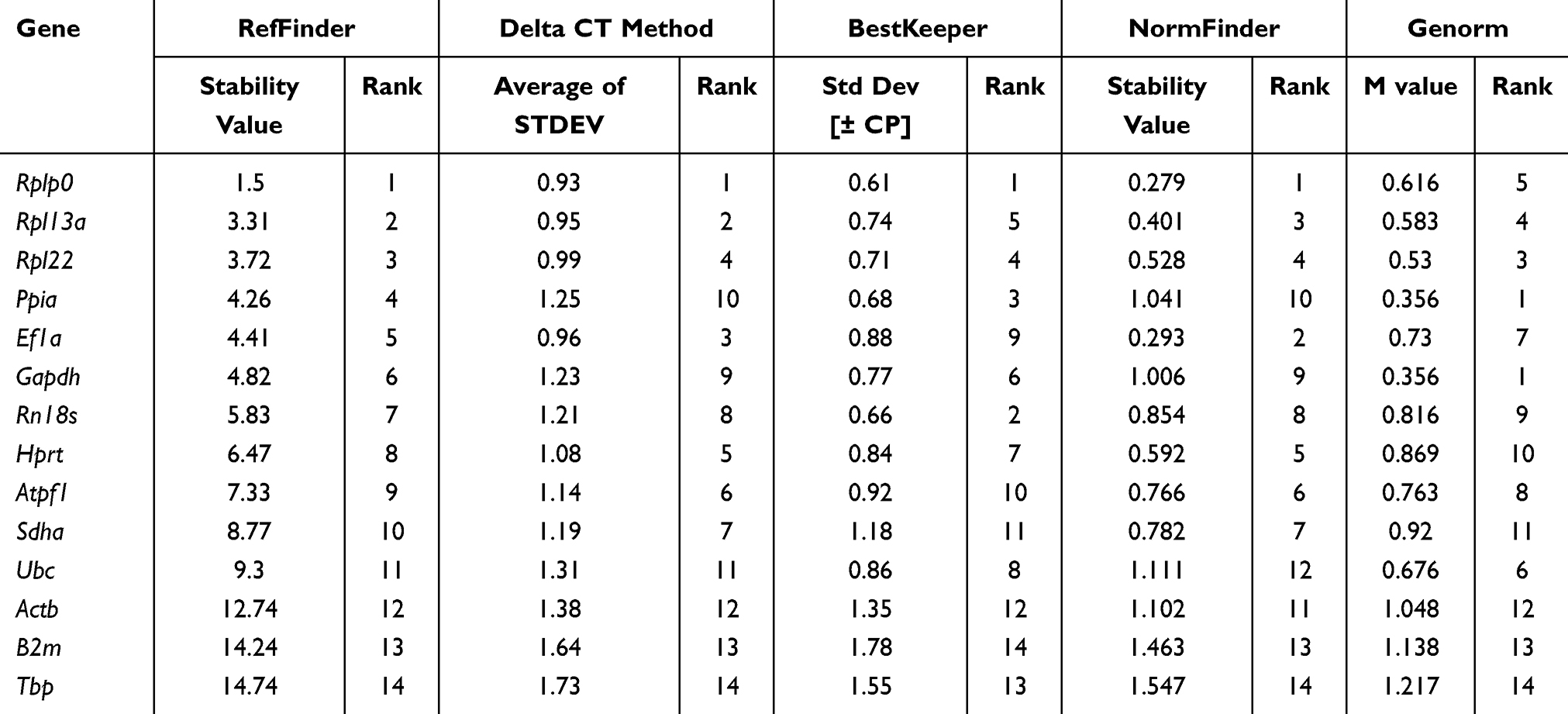 |
Table 2 Stability of Expression of Candidate Genes in Cortical Bone Tissue of C57BL/6J Mice (20–25 Weeks Old) |
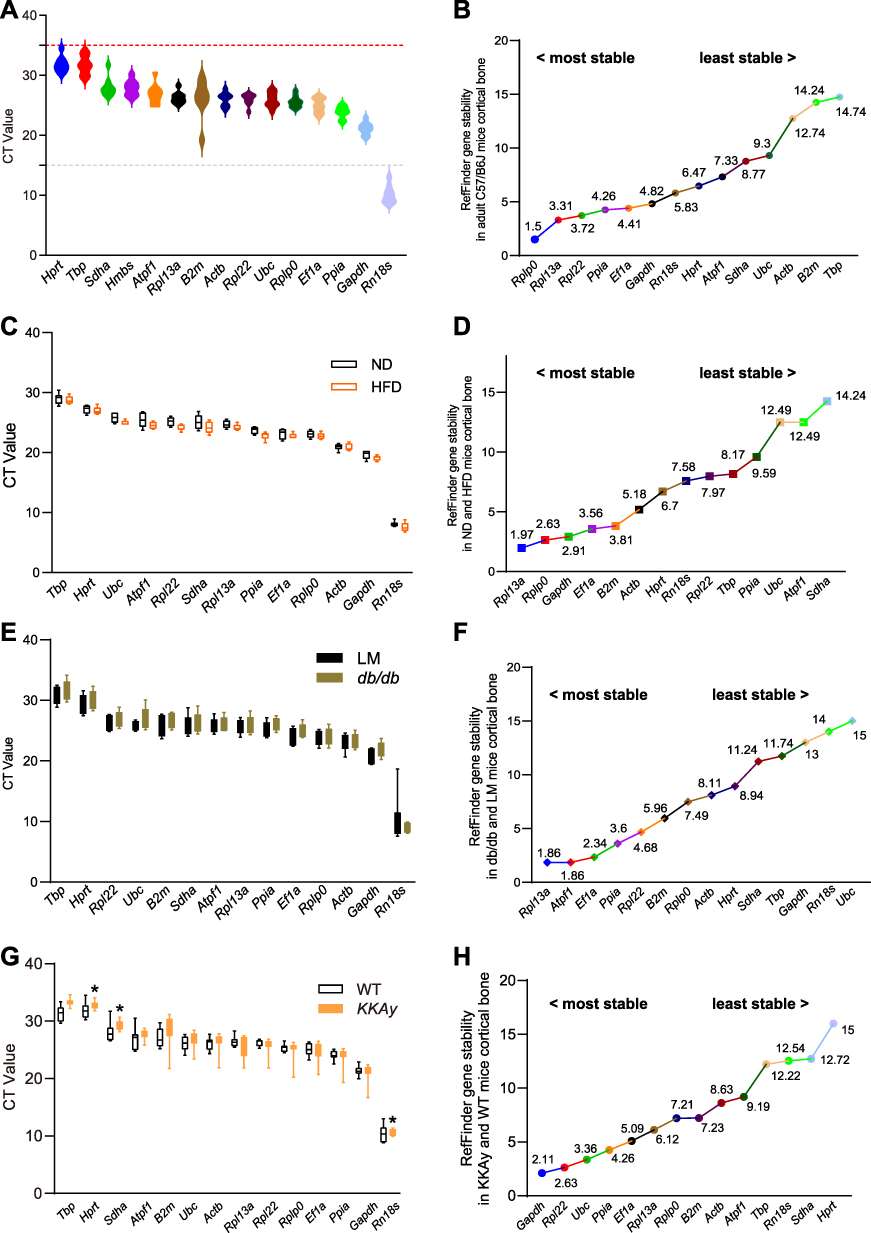 |
Figure 1 CT Values and Gene Stability Analysis Using RefFinder algorithm in Different Mouse Groups. (A) CT value of wild type (C57BL/6J) mice. (B) Gene stability in wild-type mice (RefFinder). (C) CT values in normal vs high-fat diet mice. (D) Gene stability in diet-treated mice (RefFinder). (E) CT values in db/db mice vs littermates. (F) Gene stability in db/db mice (RefFinder). (G) CT values in KKAy mice vs C57BL/6J mice. (H) Gene stability in KKAy vs C57BL/6J mice (RefFinder). |
Stable Selection of Reference Genes in Cortical Bone of HFD Mice
Increasing numbers of people are living with obesity, which is disruptive to their quality of life. Here, we assessed the suitability of housekeeping genes for analysis of control and HFD group mice, with an obese phenotype; excess energy leads to obesity, and often occurs in mice fed a high fat diet. Expression levels of the candidate genes are presented in Figure 1C. For comparisons between control and obese animals, Rpl13a was identified as having the most stable expression by RefFinder, comparative delta Ct, and GeNorm; however, NormFinder selected Ef1a as the most stable reference gene, while B2m was the most stable gene according to BestKeeper (Table 3). In general, Rpl13a was stably expressed, while Sdha was the most unstable reference gene in comparisons between control and obese animals (Figure 1D).
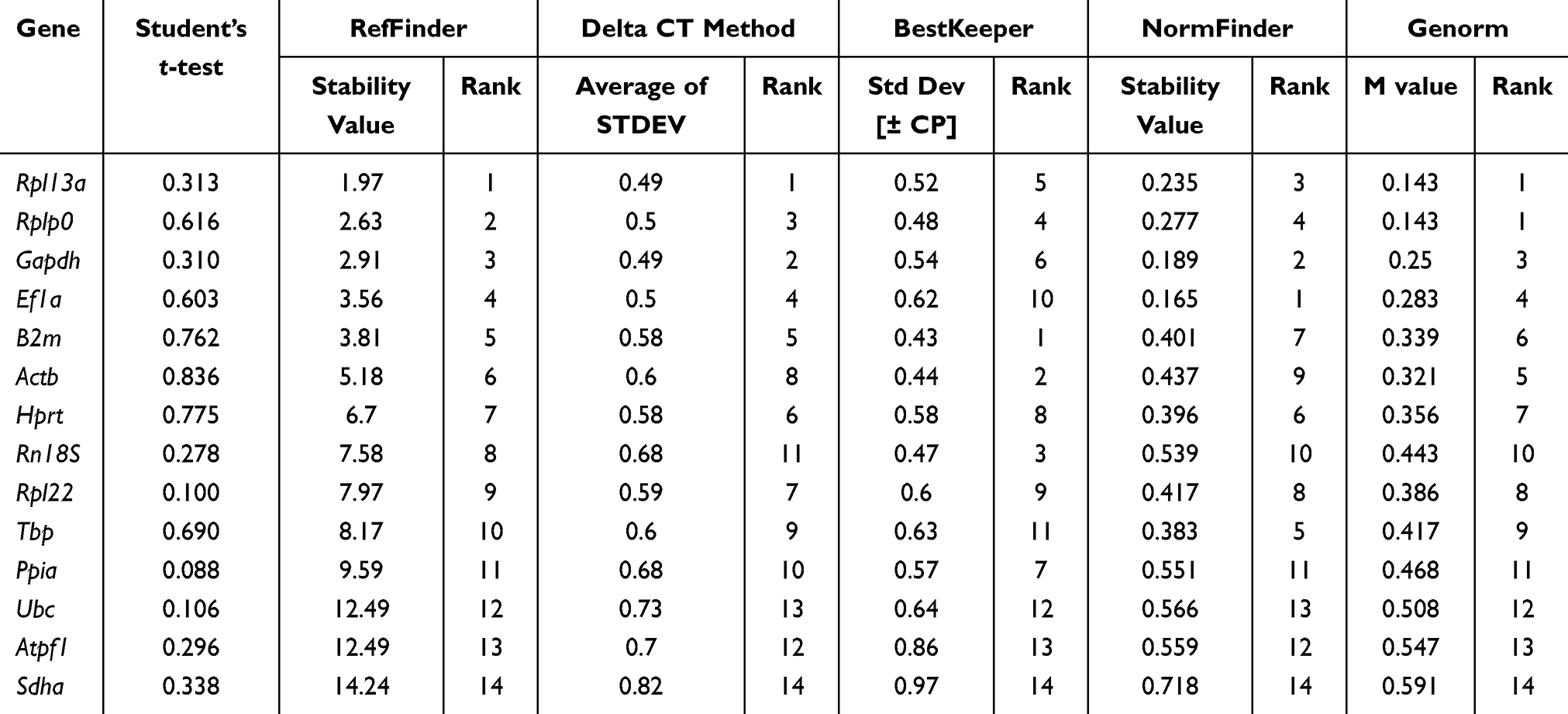 |
Table 3 Stability of Expression of Candidate Genes in Cortical Bone Tissue of Control and HFD Mice |
Stable Reference Gene Selection in Cortical Bone of db/db Mice
Diabetes is severe threat to public health.34 We analyzed gene expression in cortical bone tissue of control group and diabetic model db/db mice (Table 4). Rpl13a, Ef1a, Ppia, and Atpf1 were identified as the more stable reference genes using the four algorithms. GeNorm identified Atpf1 and Rpl13a as the best stable gene pair. B2m was expressed more stable than Actb, Gapdh, and Rn18s in control and db/db mice. Comprehensive analysis of the expression stability of internal control genes is presented in Figure 1E and F.
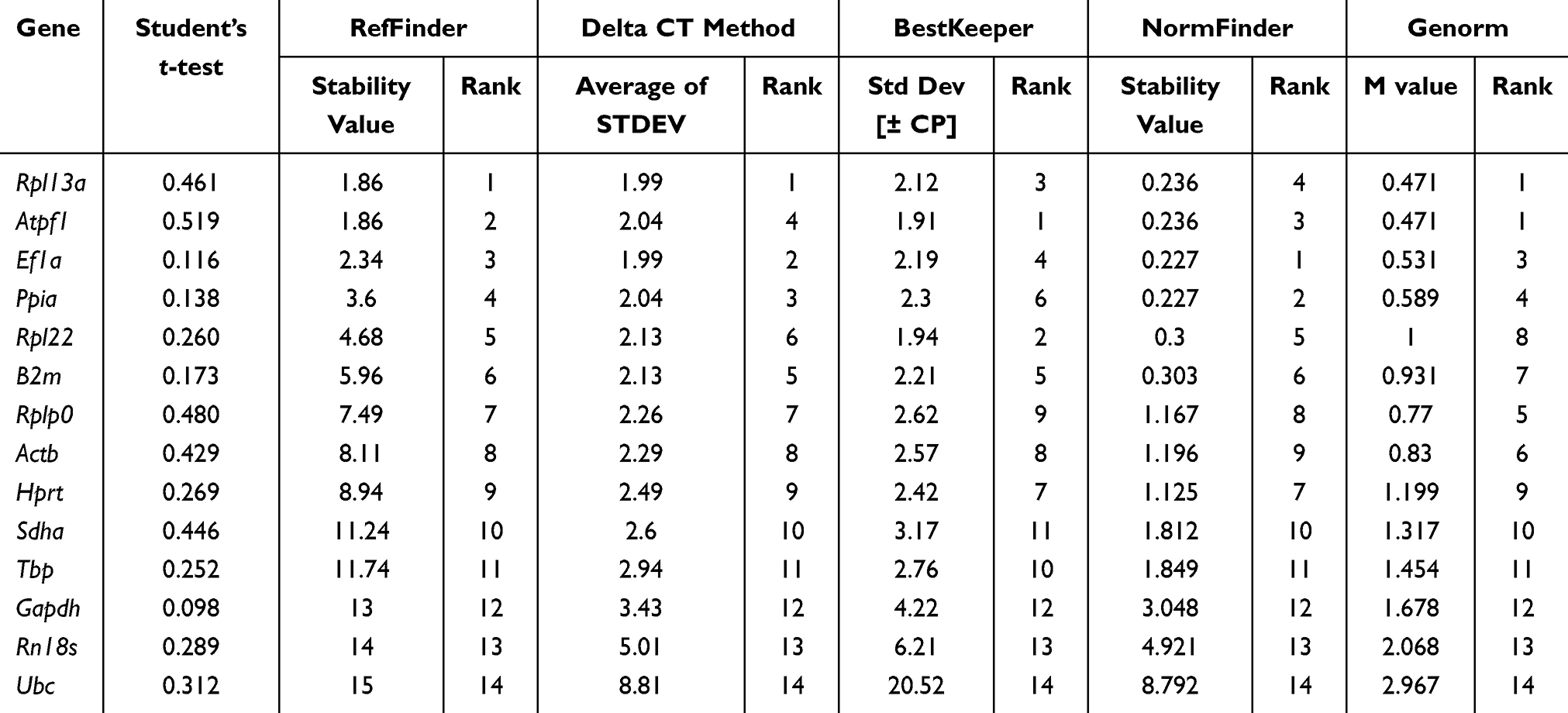 |
Table 4 Stability of Expression of Candidate Genes in Cortical Bone Tissue of Control and db/db Mice |
Stable Reference Gene Selection in Cortical Bone from KKAy Mice
We next analyzed the stability of selected housekeeping genes in the control group and another diabetic mouse model, KKAy (Table 5). Several genes were excluded from the gene expression analysis, due to the difference between the two groups, and further analysis was conducted using the remaining selected genes. Gapdh was identified as the best reference gene using comparative delta Ct and RefFinder. Rpl22 was identified as the most stable gene by BestKeeper, while Ubc was chosen as the best reference gene using NormFinder. Rpl22 and Ppia were the best gene pair for gene expression analysis, according to the GeNorm algorithm. Gene expression data in control and KKAy mice are presented in Figure 1G, and comprehensive analysis of gene stability is presented in Figure 1H.
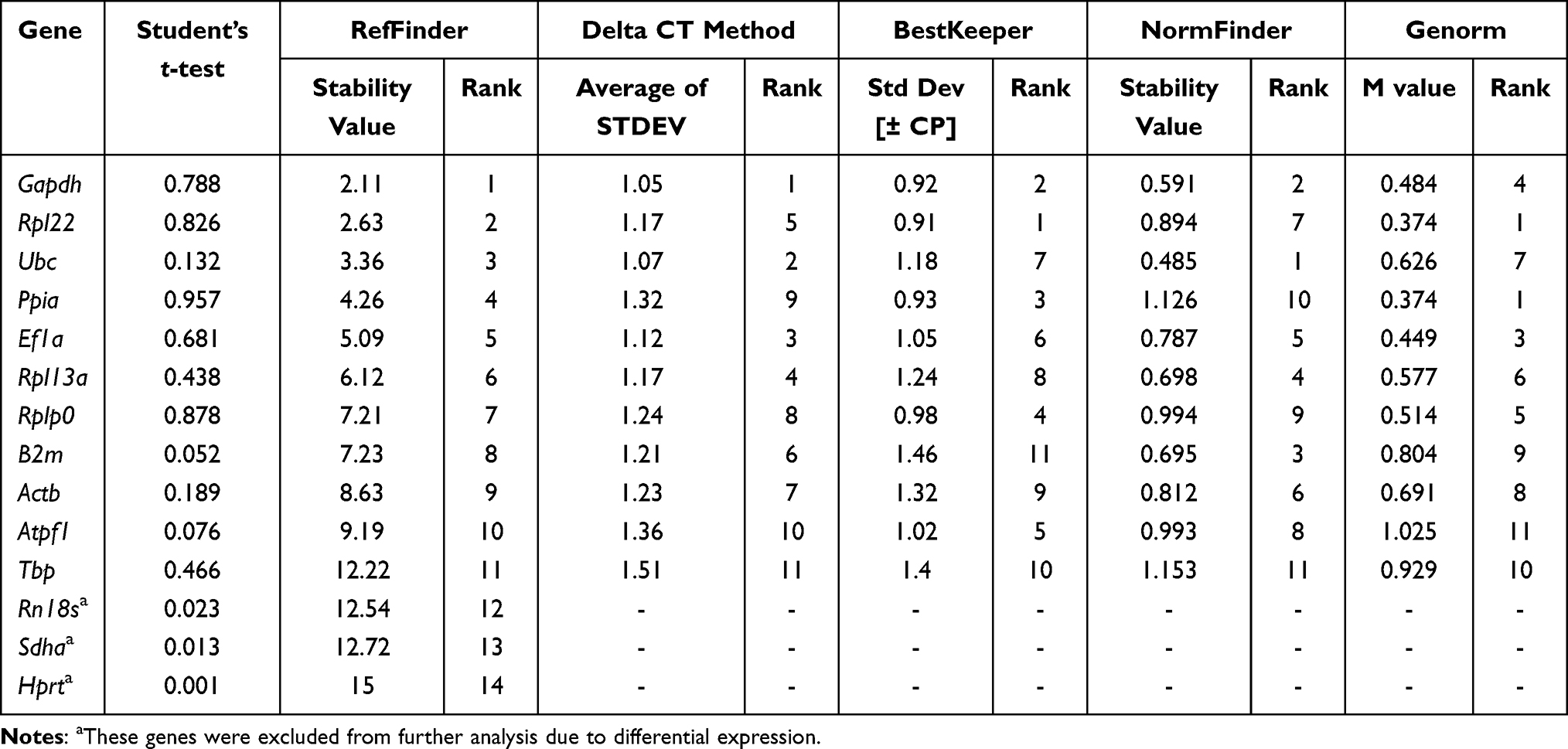 |
Table 5 Stability of Expression of Candidate Genes in Cortical Bone Tissue of C57BL/6J and KKAy Mice |
Overall, our findings show that the most frequently used reference genes (ie, Gapdh, Actb, and Rn18s) are not consistently stable in the mouse models analyzed. We found that Gapdh was more stable in mice fed with a standard or high-fat diet and in KKAy mice. The best reference gene was not the same in the two diabetic models; Rpl13a was identified as better in db/db mice, while Gapdh was selected in KKAy mice. We found that Rpl22, Rplp0, Ppia, and Ef1a were expressed relatively stable and may be better choices as candidate reference genes in analysis of mouse cortical bone tissue.
Discussion
To date, there remains no standard criterion for selection of reference genes, which are generally used generally for qPCR analyses.28 The qPCR technique is constantly being improved35 and the selection of reference genes in various samples is an important factor to consider.36 Different reference genes may have variable influence on the analysis of target genes, unlike immunohistochemical or other gold standard analysis techniques.13,15 A review reported that none of the popular reference genes, RN18S1, ACTB, B2M, GAPDH, and HPRT, were stable in more than 50% of analyses.13 Few studies have validated the stability of housekeeping genes, making the precision of results questionable. Bone tissue is comprised of bone cells (predominantly osteocytes) and abundant bone matrix, and it is difficult to obtain high-quality RNA for identification of stable reference genes.37 Some investigations have validated the stability of reference genes in osteogenic differentiation during various treatments,38–41 but only a few have focused on bone tissue,20,42 particularly in the context of metabolic disorders.
Several advancements in methods for studying metabolic disorders have been made recently. Chen et al43 employed metabolomics and machine learning to identify 26 plasma markers for early and personalized detection of metabolic syndrome. Mass spectrometry and spectroscopy, enhanced by machine learning, are advancing metabolic in vitro analysis for faster and more accurate results.44 Cao et al utilized a novel single-cell metabolomics method to explore diverse metabolic states in stem cells.45 Metabolomics, which involves the analysis of cellular metabolites, provides insights into the final products of cellular processes; however, it lacks the direct gene expression information offered by transcriptomics. Transcriptomics is gaining importance in bone research for its ability to detect real-time gene changes, aiding in early bone disease detection and management.46 Despite this, RT-qPCR remains widely used in life sciences for its cost-effectiveness and simplicity.
In this study, we aimed to validate the most stable reference genes for gene expression analysis of cortical bone tissue samples from mice fed a standard diet, as well as mouse models of obesity and diabetes.47,48 The expression stability of 14 genes was evaluated using four different algorithms. Both the comparative delta Ct and GeNorm methods rely on pairwise comparisons of gene expression. The BestKeeper algorithm estimates gene stability by considering the standard deviation of Cq values. While three of the aforementioned algorithms are based on intragroup variation, the NormFinder algorithm incorporates both intragroup and intergroup variation to estimate gene stability. The final results achieved with each algorithm often differed from one another, due to various methods of approaching the problem of calculating the expression stability used by the algorithms. The RefFinder algorithm uses the geometric mean of the results of the other algorithms to conduct a general analysis. Although this approach may not be biologically relevant in some cases, the result of RefFinder is consistently corroborated by those of the majority of other algorithms,27 indicating that the reference genes chosen using this approach are reliable.
In this study, the RefFinder algorithm validated Rpl13a as the most stable gene in the db/db and HFD models, but not in KKAy mice, possibly because ribosomal proteins are crucial components involved in basic physiological cell processes, which are key to maintaining cell development and tissue homeostasis.49,50 Ribosomal proteins are established to have essential roles in ribosome assembly and protein translation, and are also reported to have ribosome-independent functions. RPLP0 can be recruited by irradiation-induced paraspeckles, interact with NONO in the nucleus, and is involved in DNA repair.51 Eukaryotic translation elongation factor 1A (EF1A) is required for recruitment of aminoacyl tRNAs to the A-site of the ribosome during translation elongation and binding to GTP.52 GAPDH was chosen as a reference gene in most published studies, possibly because its sequence is relatively conserved and due to its established reliability as a reference gene;53 however, GAPDH is involved in various physical process in cells, and can be significantly influenced by other factors.12 In our study, Gapdh was considered the most stable housekeeping gene in KKAy mice, but found to be unstable in db/db mice. These genes are relatively stable in the three models studied. The gene that encodes the 18S rRNA subunit, Rn18s, exhibited highly variable expression in the mouse models analyzed, despite its high expression levels in mouse cortical bone tissue. The theoretical assumptions applicable to reference genes are generally unlike those for target genes, because reference gene transcripts are often present at much greater abundance than that of the studied gene.12 Moreover, while high level expression may lead to greater errors, dilution of samples before RT-qPCR may prevent detection of target genes. Previous reports concluded that Rn18s in inadequate for use in normalization of RT-qPCR analysis of osteogenic and chondrogenic differentiation54 and in mouse osteoblasts and osteoclasts.55 The Hprt1 gene encodes hypoxanthine-guanine phosphoribosyl transferase (HPRT) may regulate bone metabolism,56,57 and was also variably expressed in the models analyzed in this study. In addition, differences in levels of modification-dependent protein catabolic processes and protein ubiquitination in db/db mice may be due to variation in Ubc expression.
There are several limitations in our study. Firstly, the sample size is relatively small. Additionally, each sample in our study required the analysis of 14 genes, accompanied by a duplicate test, surpassing the capacity of a single 384-well reaction plate. Consequently, we were compelled to employ multiple plates. This experimental setup may introduce potential inconsistencies across different plates due to the limited capacity for accommodating all samples and their replicates on a single plate. Moreover, we have not yet verified the selected stable reference genes through normalization of target genes. Future work will be dedicated to validating the most suitable set of these stable genes for effective normalization of target genes.
Conclusion
Cautious selection of reference genes can contribute to the accuracy of gene expression analysis. It is recommended that RT-qPCR data are normalized using at least two validated reference genes. We identified Rpl13a as the mostly stably expressed reference gene in cortical bone tissue from mouse models of obesity and diabetes (db/db), while Gapdh was the most stable reference gene in another diabetes model, KKAy mice. Ef1a, Ppia, Rplp0, and Rpl22 are alternative genes that could be used for the normalization of gene expression in cortical bone tissue from mouse models of obesity and diabetes. In conclusion, we selected a set of housekeeping genes for use in analysis of bone from three mouse models of metabolic conditions, which will provide a convenient reference for researchers.
Acknowledgment
This research was funded by Natural Science Foundation of Chongqing (China), CSTB2022NSCQ-MSX0110, cstc2021jcyj-msxmX0320; the Future Medical Youth Innovation Program of Chongqing Medical University, W0046. The APC was funded by Chongqing Medical University.
Disclosure
The authors report no conflicts of interest in this work.
References
1. van Lierop AH, Hamdy NA, van Egmond ME, Bakker E, Dikkers FG, Papapoulos SE. Van Buchem disease: clinical, biochemical, and densitometric features of patients and disease carriers. J Bone Miner Res. 2013;28(4):848–854. doi:10.1002/jbmr.1794
2. Balemans W, Ebeling M, Patel N, et al. Increased bone density in sclerosteosis is due to the deficiency of a novel secreted protein (SOST). Hum Mol Genet. 2001;10(5):537–543. doi:10.1093/hmg/10.5.537
3. Salhotra A, Shah HN, Levi B, Longaker MT. Mechanisms of bone development and repair. Nat Rev Mol Cell Biol. 2020;21(11):696–711. doi:10.1038/s41580-020-00279-w
4. Oton-Gonzalez L, Mazziotta C, Iaquinta MR, et al. Genetics and epigenetics of bone remodeling and metabolic bone diseases. Int J Mol Sci. 2022;23(3):1500. doi:10.3390/ijms23031500
5. Raggatt LJ, Partridge NC. Cellular and molecular mechanisms of bone remodeling. J Biol Chem. 2010;285(33):25103–25108. doi:10.1074/jbc.R109.041087
6. Mazziotta C, Badiale G, Cervellera CF, Tognon M, Martini F, Rotondo JC. Regulatory mechanisms of circular RNAs during human mesenchymal stem cell osteogenic differentiation. Theranostics. 2024;14(1):143–158. doi:10.7150/thno.89066
7. Sikora M, Marycz K, Smieszek A. Small and long non-coding RNAs as functional regulators of bone homeostasis, acting alone or cooperatively. Mol Ther Nucleic Acids. 2020;21:792–803. doi:10.1016/j.omtn.2020.07.017
8. Lanzillotti C, De Mattei M, Mazziotta C, et al. Long non-coding RNAs and MicroRNAs interplay in osteogenic differentiation of mesenchymal stem cells. Front Cell Dev Biol. 2021;9:646032. doi:10.3389/fcell.2021.646032
9. Chan WCW, Tan Z, Mkt T, Chan D. Regulation and role of transcription factors in osteogenesis. Int J Mol Sci. 2021;22(11):5445. doi:10.3390/ijms22115445
10. Singh KP, Miaskowski C, Dhruva AA, Flowers E, Kober KM. Mechanisms and measurement of changes in gene expression. Biol. Res. Nurs. 2018;20(4):369–382. doi:10.1177/1099800418772161
11. Petersen JL, Tietze SM, Burrack RM, Steffen DJ. Evidence for a de novo, dominant germ-line mutation causative of osteogenesis imperfecta in two Red Angus calves. Mammalian Genome. 2019;30(3–4):81–87. doi:10.1007/s00335-019-09794-4
12. Kozera B, Rapacz M. Reference genes in real-time PCR. J Appl Genet. 2013;54(4):391–406. doi:10.1007/s13353-013-0173-x
13. Chapman JR, Waldenström J, Pena I Subirà RN. With reference to reference genes: a systematic review of endogenous controls in gene expression studies. PLoS One. 2015;10(11):e0141853. doi:10.1371/journal.pone.0141853
14. Nazet U, Schröder A, Grässel S, Muschter D, Proff P, Kirschneck C. Housekeeping gene validation for RT-qPCR studies on synovial fibroblasts derived from healthy and osteoarthritic patients with focus on mechanical loading. PLoS One. 2019;14(12):e0225790. doi:10.1371/journal.pone.0225790
15. Schwarz AP, Malygina DA, Kovalenko AA, Trofimov AN, Zaitsev AV. Multiplex qPCR assay for assessment of reference gene expression stability in rat tissues/samples. Mol Cell Probes. 2020;53:101611. doi:10.1016/j.mcp.2020.101611
16. Wang S, Wang B, He H, Sun A, Guo C. A new set of reference housekeeping genes for the normalization RT-qPCR data from the intestine of piglets during weaning. PLoS One. 2018;13(9):e0204583 doi:10.1371/journal.pone.0204583
17. Ng ACT, Delgado V, Borlaug BA, Bax JJ. Diabesity: the combined burden of obesity and diabetes on heart disease and the role of imaging. Nat Rev Cardiol. 2021;18(4):291–304. doi:10.1038/s41569-020-00465-5
18. Bustin SA, Benes V, Garson JA, et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 2009;55(4):611–622. doi:10.1373/clinchem.2008.112797
19. Robinson NB, Krieger K, Khan FM, et al. The current state of animal models in research: a review. Int J Surg. 2019;72:9–13 doi:10.1016/j.ijsu.2019.10.015
20. Cepollaro S, Della Bella E, de Biase D, Visani M, Fini M. Evaluation of RNA from human trabecular bone and identification of stable reference genes. J Cell Physiol. 2018;233(6):4401–4407. doi:10.1002/jcp.26319
21. Zhang W-X, Fan J, Ma J, Rao Y-S, Zhang L, Yan Y-E. Selection of suitable reference genes for quantitative real-time PCR normalization in three types of rat adipose tissue. Int J Mol Sci. 2016;17(6). doi:10.3390/ijms17060968
22. Liu L, Han H, Li Q, et al. Selection and validation of the optimal panel of reference genes for RT-qPCR analysis in the developing rat cartilage. Front Genet. 2020;11:590124. doi:10.3389/fgene.2020.590124
23. Fajardo RJ, Karim L, Calley VI, Bouxsein ML. A review of rodent models of type 2 diabetic skeletal fragility. J Bone Miner Res. 2014;29(5):1025–1040. doi:10.1002/jbmr.2210
24. Amend SR, Valkenburg KC, Pienta KJ. Murine hind limb long bone dissection and bone marrow isolation. J Vis Exp. 2016;110:53936 doi:10.3791/53936
25. Faraldi M, Mangiavini L, Conte C, Banfi G, Napoli N, Lombardi G. A novel methodological approach to simultaneously extract high-quality total RNA and proteins from cortical and trabecular bone. Open Biol. 2022;12(5):210387. doi:10.1098/rsob.210387
26. Becker C, Hammerle-Fickinger A, Riedmaier I, Pfaffl MW. mRNA and microRNA quality control for RT-qPCR analysis. Methods San Diego Calif. 2010;50(4):237–243. doi:10.1016/j.ymeth.2010.01.010
27. Almeida-Oliveira F, Leandro JGB, Ausina P, Sola-Penna M, Majerowicz D. Reference genes for quantitative PCR in the adipose tissue of mice with metabolic disease. Biomed Pharmacothe. 2017;88:948–955. doi:10.1016/j.biopha.2017.01.091
28. Joshi CJ, Ke W, Drangowska-Way A, O’Rourke EJ, Lewis NE. What are housekeeping genes? PLoS Comput Biol. 2022;18(7):e1010295. doi:10.1371/journal.pcbi.1010295
29. Vandesompele J, De Preter K, Pattyn F, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3(7). doi:10.1186/gb-2002-3-7-research0034
30. Pfaffl MW, Tichopad A, Prgomet C, Neuvians TP. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--excel-based tool using pair-wise correlations. Biotechnol Lett. 2004;26(6):509–515. doi:10.1023/B:BILE.0000019559.84305.47
31. Andersen CL, Jensen JL, Ørntoft TF. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004;64(15):5245–5250. doi:10.1158/0008-5472.CAN-04-0496
32. Silver N, Best S, Jiang J, Thein SL. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol Biol. 2006;7:33. doi:10.1186/1471-2199-7-33
33. Xie F, Xiao P, Chen D, Xu L, Zhang B. miRDeepFinder: a miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol Biol. 2012;80(1):75–84. doi:10.1007/s11103-012-9885-2
34. Hu C, Jia W. Diabetes in China: epidemiology and genetic risk factors and their clinical utility in personalized medication. Diabetes. 2018;67(1):3–11. doi:10.2337/dbi17-0013
35. Gadkar V, Filion M. New developments in quantitative real-time polymerase chain reaction technology. Curr Issues Mol Biol. 2014;16:1–6 doi:10.21775/cimb.016.001
36. Gachon C, Mingam A, Charrier B. Real-time PCR: what relevance to plant studies? J Exp Bot. 2004;55(402):1445–1454. doi:10.1093/jxb/erh181
37. Carter LE, Kilroy G, Gimble JM, Floyd ZE. An improved method for isolation of RNA from bone. BMC Biotechnol. 2012;12(1):5. doi:10.1186/1472-6750-12-5
38. Okamura K, Inagaki Y, Matsui TK, et al. RT-qPCR analyses on the osteogenic differentiation from human iPS cells: an investigation of reference genes. Sci Rep. 2020;10(1):11748. doi:10.1038/s41598-020-68752-2
39. Jacobi A, Rauh J, Bernstein P, Liebers C, Zou X, Stiehler M. Comparative analysis of reference gene stability in human mesenchymal stromal cells during osteogenic differentiation. Biotechnol Prog. 2013;29(4):1034–1042. doi:10.1002/btpr.1747
40. Hasler J, Hatt LP, Stoddart MJ, Armiento AR. Stable reference genes for qPCR analysis in BM-MSCs undergoing osteogenic differentiation within 3D hyaluronan-based hydrogels. Int J Mol Sci. 2020;21(23):9195. doi:10.3390/ijms21239195
41. Franko N, Vrščaj LA, Zore T, Ostanek B, Marc J, Lojk J. TBP, PPIA, YWHAZ and EF1A1 are the most stably expressed genes during osteogenic differentiation. Int J Mol Sci. 2022;23(8):4257. doi:10.3390/ijms23084257
42. Otto E, Köhli P, Appelt J, et al. Validation of reference genes for expression analysis in a murine trauma model combining traumatic brain injury and femoral fracture. Sci Rep. 2020;10(1):15057. doi:10.1038/s41598-020-71895-x
43. Chen Y, Xu W, Zhang W, et al. Plasma metabolic fingerprints for large-scale screening and personalized risk stratification of metabolic syndrome. Cell Rep Med. 2023;4(7):101109. doi:10.1016/j.xcrm.2023.101109
44. Chen X, Shu W, Zhao L, Wan J. Advanced mass spectrometric and spectroscopic methods coupled with machine learning for in vitro diagnosis. View. 2023;4(1):20220038 doi:10.1002/VIW.20220038
45. Cao J, Yao QJ, Wu J, et al. Deciphering the metabolic heterogeneity of hematopoietic stem cells with single-cell resolution. Cell Metab. 2024;36(1):209–221.e206. doi:10.1016/j.cmet.2023.12.005
46. Reppe S, Datta HK, Gautvik KM. Omics analysis of human bone to identify genes and molecular networks regulating skeletal remodeling in health and disease. Bone. 2017;101:88–95. doi:10.1016/j.bone.2017.04.012
47. Kleinert M, Clemmensen C, Hofmann SM, et al. Animal models of obesity and diabetes mellitus. Nat Rev Endocrinol. 2018;14(3):140–162. doi:10.1038/nrendo.2017.161
48. Takagi S, Miura T, Yamashita T, et al. Characteristics of diabetic osteopenia in KK-Ay diabetic mice. Biol Pharm Bull. 2012;35(3):438–443. doi:10.1248/bpb.35.438
49. Robledo S, Idol RA, Crimmins DL, Ladenson JH, Mason PJ, Bessler M. The role of human ribosomal proteins in the maturation of rRNA and ribosome production. RNA. 2008;14(9):1918–1929. doi:10.1261/rna.1132008
50. Pillet B, García-Gómez JJ, Pausch P, et al. The dedicated chaperone Acl4 escorts ribosomal protein Rpl4 to its nuclear Pre-60S assembly site. PLoS Genet. 2015;11(10):e1005565. doi:10.1371/journal.pgen.1005565
51. Wang Y-L, Zhao -W-W, Bai S-M, et al. DNA damage-induced paraspeckle formation enhances DNA repair and tumor radioresistance by recruiting ribosomal protein P0. Cell Death Dis. 2022;13(8):709. doi:10.1038/s41419-022-05092-1
52. Mishra AK, Gangwani L, Davis RJ, Lambright DG. Structural insights into the interaction of the evolutionarily conserved ZPR1 domain tandem with eukaryotic EF1A, receptors, and SMN complexes. Proc Natl Acad Sci U S A. 2007;104(35):13930–13935. doi:10.1073/pnas.0704915104
53. De Keyser E, Desmet L, Van Bockstaele E, De Riek J. How to perform RT-qPCR accurately in plant species? A case study on flower colour gene expression in an azalea (Rhododendron simsii hybrids) mapping population. BMC Mol Biol. 2013;14:13. doi:10.1186/1471-2199-14-13
54. Ragni E, Viganò M, Rebulla P, Giordano R, Lazzari L. What is beyond a qRT-PCR study on mesenchymal stem cell differentiation properties: how to choose the most reliable housekeeping genes. J Cell Mol Med. 2013;17(1):168–180. doi:10.1111/j.1582-4934.2012.01660.x
55. Stephens AS, Stephens SR, Morrison NA. Internal control genes for quantitative RT-PCR expression analysis in mouse osteoblasts, osteoclasts and macrophages. BMC Res Notes. 2011;4:410. doi:10.1186/1756-0500-4-410
56. Yan B, Li J, Zhang L. Identification of B cells participated in the mechanism of postmenopausal women osteoporosis using microarray analysis. Int J Clin Exp Med. 2015;8(1):1027–1034.
57. De lima CAD, de Lima SC, Barbosa AD, et al. Postmenopausal osteoporosis reference genes for qPCR expression assays. Sci Rep. 2019;9(1):16533. doi:10.1038/s41598-019-52612-9
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.