Back to Journals » ImmunoTargets and Therapy » Volume 14
Cytokine Couture: Designer IL2 Molecules for the Treatment of Disease
Authors Dashwood A, Ghodsinia AA, Dooley J, Liston A
Received 25 January 2025
Accepted for publication 20 March 2025
Published 4 April 2025 Volume 2025:14 Pages 403—431
DOI https://doi.org/10.2147/ITT.S500229
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Shurin
Amy Dashwood,1,2 Arman Ali Ghodsinia,1 James Dooley,1 Adrian Liston1
1Department of Pathology, University of Cambridge, Cambridge, UK; 2Immunology Programme, Babraham Institute, Cambridge, UK
Correspondence: Amy Dashwood; Email [email protected] Adrian Liston, Email [email protected]
Abstract: Interleukin 2 (IL2) is a dual-acting cytokine, playing important roles in both immune activation and regulation. The role IL2 plays as a potent activator of CD8 T cells saw IL2 become one of the earliest immunotherapies, used for the treatment of cancer. In more recent years refined understanding of IL2, and the potent capacity it has for Treg stimulation, has seen low-dose IL2 therapy trialled for the treatment of auto-immune and inflammatory conditions. However, despite clinical successes, IL2 therapy is not without its caveats. The complicated receptor biology of IL2 gives rise to a narrow therapeutic window, made problematic by its short half-life. Armed with a better understanding of the structure of IL2 in complex with its receptors, many attempts have been made to create designer IL2 molecules which overcome these problems. A wide range of approaches have been used, resulting in > 100 designer IL2 molecules. These include antibody complexes, fusion proteins, mutant IL2 molecules and PEGylation, each uniquely modifying the biological activity in an effort to enhance its therapeutic potential. Collectively, designer IL2 molecules form a blueprint outlining modification pathways available to other immunotherapeutics, paving the way for the next generation of immunotherapy.
Keywords: cytokine, protein engineering, T cell, treg, interleukin 2
Introduction
Since the discovery of IL2 as T cell growth factor in 1978 by Kendal Smith and team,1 IL2 has been an alluring target for therapeutic use. Based on the early understanding of IL2 in the promotion of effector T cell responses, initial drug development was based around stimulating anti-tumour responses. Indeed, the approval of Proleukin (Aldesleukin) for the treatment of renal carcinoma in 1992,2 was arguably the first cancer immunotherapeutic. Although IL2 therapy provided promising results in cancer treatment, with 15–17% of patients experiencing an objective clinical regression in disease,3 dosing was, and remains, difficult. Patients receive large bolus injections daily to reach therapeutic concentrations due to the short half-life of IL2.4,5 In addition, off target effects of high dose IL2 can lead to serious side effects, such as vascular leak syndrome (VLS).6
Many of the early disappointments in IL2 therapeutics can, in retrospect, be attributed to the unappreciated role of IL2 as the key cytokine in regulatory T cell (Treg) homeostasis,7,8 making the biology of this molecule more complicated than initially appreciated. The renaissance of IL2 as an anti-inflammatory biologic lead to reinvigorated therapeutic testing of IL2, with refined dosing to stimulate and promote Treg survival in a range of inflammatory diseases, such as GVHD and SLE.9–13 Recently, with enhanced understanding of the structure of IL2 binding to its receptors,14,15 the opportunity has arisen to modify this complicated cytokine to enhance the biological properties needed for therapeutic use in both inflammatory and anti-inflammatory contexts. Whilst designer IL2 molecules have been well reviewed in the context of cancer immunotherapy and inflammatory disease,16,17 here we aim to create an extensive resource which brings together the <100 designer IL2 molecules in the clinic or in development as therapeutics and mouse analogues developed as key research tools, their design strategies, known biological features, and their potential to finally unlock the power of IL2 therapy.
A Hierarchy of Binding: IL2 and Its Receptors
The relationship between IL2 and its receptors lies at the heart of its dualistic behaviour, playing a critical role in both immune activation and regulation. Cellular selectivity of the IL2 molecule is driven through differential receptor expression on cellular targets which dictate the affinity and downstream signalling pathways. The IL2 receptor exists in three conformations, characterised by their relative affinity, each made up of a different assembly of the protein compartments IL2Rα (CD25), IL2Rβ (CD122) and IL2Rγ (CD132) (Figure 1). The highest affinity receptor is a trimer comprised of all three components.18 At baseline, the high affinity receptor is expressed primarily on the surface of Tregs, with stimulation leading to increased Treg fitness and survival.19,20 Upon IL2 engagement, the receptor complex activates downstream JAK-STAT signalling, particularly the STAT5 pathway, leading to enhanced Treg proliferation, survival, and suppressive function. This mechanism is crucial for maintaining immune tolerance and preventing autoimmunity. In contrast, the intermediate affinity receptor is a dimer comprised of an IL2Rβ and an IL2Rγ subunit which is primarily expressed on the surface of CD8 T and NK cells. This receptor configuration has an affinity ~100 fold lower than its trimeric counterpart.18 Consequently, under normal physiological conditions, IL2 availability is limited for CD8+ T and NK cells, instead favouring Treg expansion. However, during immune activation and inflammatory responses, IL2 production can surge, leading to transient stimulation of effector cells through the intermediate-affinity receptor. This dynamic regulation allows IL2 to promote cytotoxic T cell and NK cell proliferation, differentiation, and effector function, which are essential for pathogen clearance and tumour immunity.21 Additionally, IL2Rα (CD25) can function independently as a low-affinity receptor, binding IL2 without initiating downstream signalling. It can also be shed into the extracellular environment as a soluble form (sCD25), which has been proposed to act as a decoy receptor, sequestering excess IL2 and modulating its bioavailability.22 While the exact physiological role of sCD25 remains incompletely understood, it is thought to contribute to immune regulation by limiting IL2-driven activation of effector cells. Elevated levels of sCD25 have been associated with various inflammatory and autoimmune diseases, suggesting its potential as a biomarker for immune dysregulation.22–25 As IL2 binding to the IL2RA and IL2RB subunits is mediated through different interaction surfaces, the processes are largely, although not entirely, independent. This understanding has enabled the bioengineering of IL2 into potential therapeutics with altered affinity to one or both receptors. Alterations in this interface, or modifications to prolong the short half-life of the molecule, are the primary modifications present in the >100 altered IL2 molecules currently characterised.
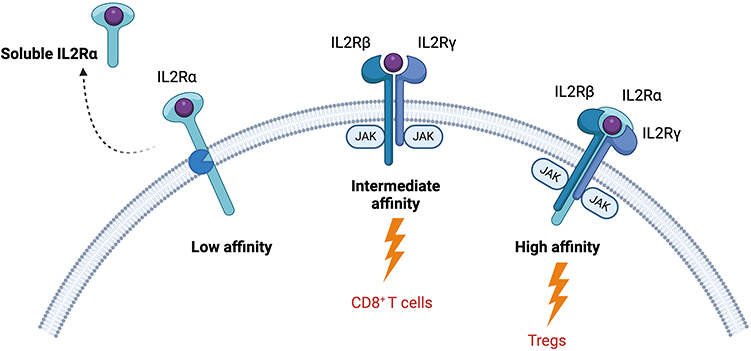 |
Figure 1 IL2 receptor biology. IL2 is a pleiotropic cytokine; its dualistic behaviour arises from its relationship with its differentially expressed receptors. The low affinity receptor is comprised of only the IL2Rα subunit and can be cleaved to become a soluble cytokine receptor. It is thought to act as a decoy to quench excess IL2. The intermediate affinity receptor is primarily expressed by inflammatory cells such as CD8 T cells and in made up of the IL2Rβ and IL2Rγ subunits. Finally, the high affinity receptor is highly expressed on regulatory T cells and is comprised of all three subunits. Created in BioRender. Dashwood, A. (2025) https://BioRender.com/ h38f115. |
Anti-Inflammatory IL2Rα-Biased Agents
With the high therapeutic potential of harnessing the suppressive capacity of Tregs for treating inflammatory conditions, extensive work has gone into developing strategies to elevate the capacity of IL2 to work through the IL2Rα component. As Tregs have the highest basal expression of IL2Rα, these therapeutics shift the response towards Treg survival and fitness, creating a more anti-inflammatory environment. The strategies used to create these α-biased IL2 molecules are diverse, including antibody complexes, Fc fusions, point mutations, directed PEGylation, and de novo design (Table 1). Each strategy has had varying success with a several candidates making it into clinical trials.
 |
Table 1 IL2Rα-Biased (Treg-Promoting) IL2 Muteins |
Antibody Complexes
The use of anti-IL2 antibodies to create immune complexes were the first inadvertent steps towards producing receptor-biased IL2 products. Initially generated to neutralise IL2, the conflicting effects of different IL2 antibodies were eventually identified to relate to the site of IL2 binding, with antibodies that bound the IL2Rα interface giving a relative boost to IL2Rβ signals, and vice versa.107 An example of this being used to create IL2Rα bias is IL2/F5111, which is a complex between IL2 and the F5111 anti-IL2 antibody.26,27 F5111 binds to IL2 in a manner that sterically blocks the binding site to the IL2Rβ receptor, preventing responses from cells that rely exclusively on this interface for signalling. Antibody complexes have the additional benefit that they can extend protein half-life by increasing its molecular weight and altering its biophysical properties, therefore reducing clearance. This beneficial effect has potential issues when transferring to clinic, ie how long are these complexes stable for and can the complex dissociate within the body resulting in off target effects. For these reasons, it may be beneficial to covalently link the IL2 molecule, via the IL2Rβ-interaction face, to a fusion protein carrier, to achieve similar effects. Additionally, antibody complexes can bear further production costs over fusion proteins or muteins as two separate proteins need to be expressed and purified.
Decreased IL2Rβ Binding
With insights delivered on IL2 receptor binding by antibody neutralisation and structural studies, new mutated proteins, or muteins, could be designed that interfered with the capacity of IL2 to bind one of its receptors without the need for antibody blockade (Figure 2). In the case of Treg-biased muteins, the simplest approach is to mutate residues in IL2 which are important to binding the IL2Rβ interface, thereby preventing activation of the inflammatory cells that exclusively rely on IL2Rβ-IL2Rγ for signalling. An example of this approach is the IL2 mutant N88D, where mutating the asparagine at position 88 to an aspartic acid, decreased IL2 affinity to the IL2Rβ subunit.37 The authors report a 30–80-fold reduced ability to activate receptors present on effector T cells and NK cells whilst having only a minimal reduction (6-fold) in the ability to activate the high affinity receptor on Tregs. This marginal decrease in overall bioactivity of IL2 is often seen in IL2Rβ-impeded muteins.32,39,40,55 A probable explanation for this decreased bioactivity is the requirement to form the trimer to allow signalling, as the IL2Rα subunit cannot signal alone. However, despite this, N88D entered phase 1 clinical trials in the form of an Fc fusion protein (PT101), in ulcerative colitis.71 Results of this clinical trial will help shed light on whether this decrease in bioactivity will affect overall performance in vivo.
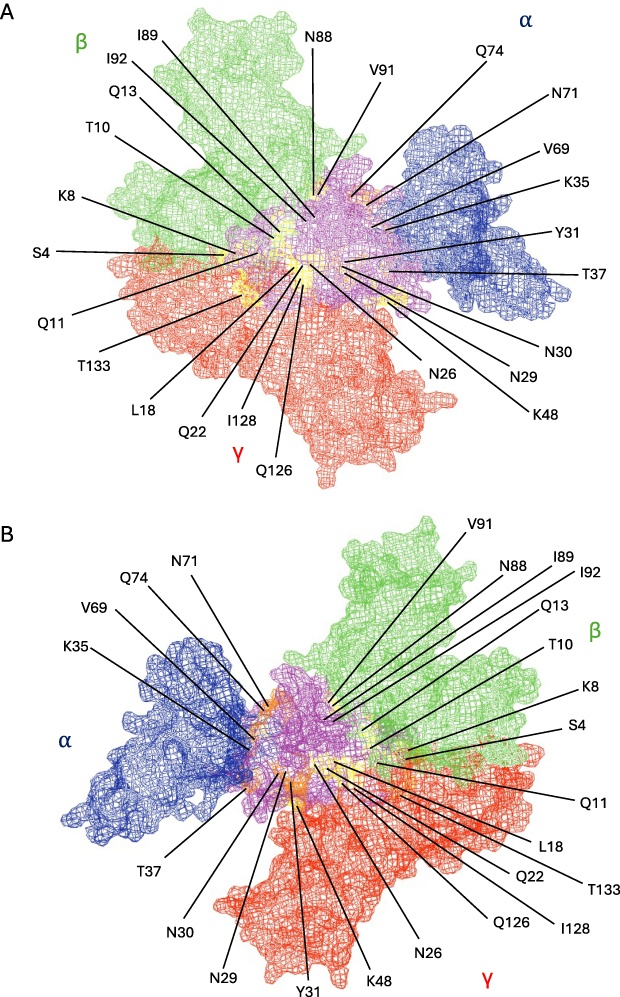 |
Figure 2 IL2Rα-biased point muteins. IL2 (purple) in complex with receptor subunits IL2Rα (blue), IL2Rβ (green), and IL2Rγ (red), based on 2ERJ,15 visualised from above the membrane (A) and from below the membrane (B). Residues of IL2 which have previously been targeted for mutation are highlighted in Orange (disrupt IL2Rα binding) and yellow (enhance IL2Rβγ binding). (Figure created in PyMol). |
Increased IL2Rα Binding
An alternate way to mutate IL2 to enhance its anti-inflammatory effects is to increase affinity to the IL2Rα subunit. Rao et al performed this type of mutation with M1 and M6.44 Both muteins possess point mutations, Q74P and V69A which lie at the interface with the IL2Rα subunit. M6 has one further mutation I128T which lies close to the predicted IL2Rβ interface. Both mutants were shown to have an increased affinity for the alpha subunit. However, when M1 and M6 were evaluated in proliferation assays using KIT-225 T cell line, M1 failed to perform better than WT IL2, suggesting that the increased binding to the IL2Rα subunit alone was not enough to confer increased biological potency. On the other hand, M6 performed better than WT IL2, increasing proliferation by 50–60%.44 Neither mutein has yet progressed to pre-clinical or clinical studies.
Pro-Inflammatory IL2Rβ/IL2Rγ-Biased Agents
For enhanced therapeutic use of IL2 in its original purpose, oncology, the reciprocal changes are required. IL2Rβ-biased IL2 modifications are expected to have enhanced or maintained expansion of CD8 T cell and NK cell anti-tumour responses, without inducing a suppressive Treg response through IL2Rα (Table 2). While the engineering strategies used closely parallel those of IL2Rα-biased agents, many more candidates have progressed to the clinic, likely due to the simpler structural problem of excluding a non-essential signalling component (IL2Rα).
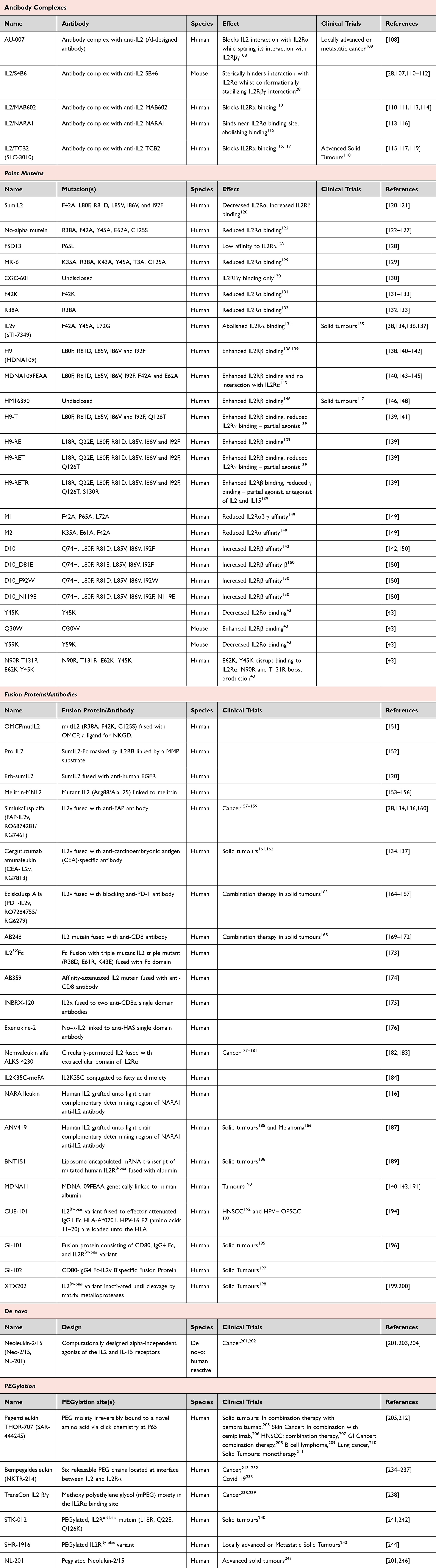 |
Table 2 IL2Rβ/IL2Rγ-Biased (CD8 T Cell-Promoting), Inflammatory Agents |
Antibody Complexes
Similar to the earliest IL2Rα-biased IL2 agents, antibody complexing was among the earliest approaches used to alter the biophysical properties and bioactivity of IL2 to bias it towards the dimeric receptor. These complexes result in pro-inflammatory responses. Several examples have made it into clinical trials in various types of cancer. Amongst these is IL2/TCB2, a complex formed of IL2 and an anti-IL2 which blocks binding to the α receptor subunit.115,117 In preclinical studies the complex, which was prepared daily before IP injection, resulted in inhibition of tumour growth in three independent models. This effect was further enhanced when used in conjunction with anti-PD1, resulting in 100% tumour rejection of MC38 colon cancer cells.115 IL2/TCB2 has since moved into phase 1/2 clinical trials, which are actively recruiting patients with solid tumours, as both a monotherapy and in combination with chemo-agent gemcitabine.118
Decreased IL2Rα Binding
In the same way that designer IL2 muteins can be made with decreased IL2Rβ binding, to enhance Treg selectivity, muteins can be generated with decreased IL2Rα binding, to enhance inflammatory cell selectivity (Figure 3). One example of this is the no-alpha IL2 mutein where point mutations (R38A, F42A, Y45A, E62A, C125S) have been introduced at the IL2 interface with the IL2Rα subunit.123 The resulting effect of this is a 1000-fold decrease in the EC50 of the mutein on Tregs, whilst its ability to stimulate CD8 T cells is maintained. In preclinical models, treatment with this mutein translated to a reduction in tumour size which was later shown to be a direct effect of altering the CD8:Treg balance.124 This provides a proof-of-concept for the utility of this class of agents in a clinical setting.
 |
Figure 3 IL2Rβγ-biased point muteins. IL2 (purple) in complex with receptor subunits IL2Rα (blue), IL2Rβ (green), and IL2Rγ (red), based on 2ERJ,15 visualised from above the membrane (A) and from below the membrane (B). Residues of IL2 which have previously been targeted for mutation are highlighted in Orange (enhance IL2Rα binding) and yellow (disrupt IL2Rβγ binding). (Figure created in PyMol). |
Increased IL2Rβ Binding
IL2 can also be engineered to have an increased binding to the IL2Rβ subunit. This is the case for D10. D10 harbours a number of point mutations (Q74H, L80F, R81D, L85V, I86V, I92F) which increase the ability of IL2 to stimulate cells expressing the intermediate affinity receptor by strengthening the binding to IL2Rβ and signalling through the IL2Rβ-IL2Rγ dimer.150 In particular, the mutation at position 74 (Q74H) caused a conformational change which allows D10 to create stronger hydrophobic interactions in the receptor pocket whilst mutations R81D, I92F contributed to a higher binding energy.150 These gain-of-function mutations provide a theoretical biotechnological benefit compared to the loss-of-function strategies, as the predicted protein production required to achieve functional thresholds should be reduced.
Ortho IL2 – Designer IL2 to Match Designer T Cells
Another exciting concept in the IL2 mutein space are Ortho-IL2 cytokines. Ortho IL2 cytokines are IL2 replacement molecules designed to bind in pairs with a replacement (orthogonal IL2) extracellular domain fused to the IL2 receptor. This creates classical IL2 signals, through the interaction of entire new ligand-receptor pairs. In practice, this requires engineering T cells to express the orthogonal IL2 receptor, transplanting them into the patient and then exposing to Ortho-IL2. As Ortho-IL2 has no affinity for the native IL2 receptor, it will specifically activate the ortho-IL2 receptor on the engineered (transplanted) T cells. There are many different orthogonal pairs which have been developed, which are now being tested in different pre-clinical models, for example, in cancer,247,248 in transplantation models249 and in GVHD.250 This elegant method bypasses issues relating to the IL2 molecule itself, and overcomes the issue that even with strong receptor biasing, off-target effects may still occur due to receptor co-expression, for example, expression of IL2Rα on activated CD8 T cells. However, Ortho-IL2 is only suitable for approaches that utilise transplanted and engineerable T cells, limiting its utility and making it a costly therapeutic approach.
IL2 Engineering Designed to Increase Half-Life
Distinct from the problem of biasing the specificity of IL2 towards pro- or anti-inflammatory functions, additional engineering has been performed to increase the half-life (Table 3). The serum half-life of IL2 is only ~12 minutes,251 making it more difficult to use as a therapeutic without using large bolus doses to sustain therapeutic concentrations. Unfortunately, the bolus dose approach influences the hierarchy of reactivity, by supporting higher affinity reactions, and can trigger adverse events through triggering atypical IL2 pathways.252,253 This narrowing of the therapeutic window limits utility, making it desirable to create IL2 molecules with a longer half-life.
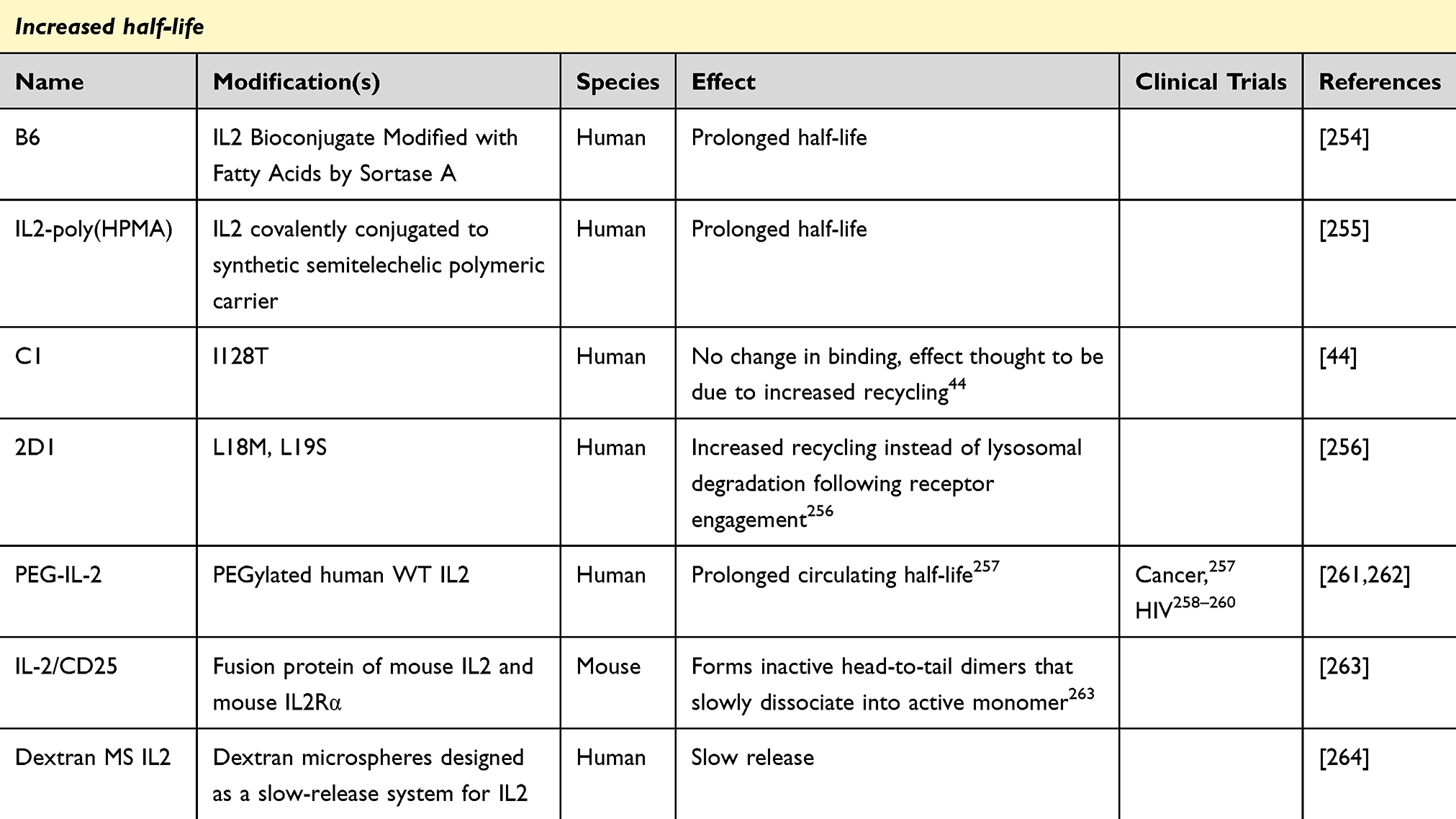 |
Table 3 IL2 Agents Designed to Have an Increased Half-Life |
While the function of antibody complexes described above focused on altering the specificity of IL2 by shielding one of its two receptor interfaces, antibody binding also substantially lengthens the half-life of IL2, allowing lower doses to be used with greater spacing. The effect of antibody complexing on half-life is likely brought about by both slower degradation and decreased consumption by off-target cells. For example, Boyman and team showed IL2/mAbCD122 complexes, using SB46 (mouse) and MAB602 (human), relied on increased serum half-life brought about by FcRn to enhance IL2s biological function.110 In addition, they showed IL2/mAbCD25 complexes using JES6-1 had a further increased half-life, suggesting differing consumption rates may also play a factor.110 Another interesting case was reported by Ward et al who, when designing CD8-biased fusion proteins, instead, discovered that an IL2-sCD25 fusion protein forms inactive head-to-tail dimers. These dimers which slowly dissociate in vivo, act as a slow-release system for IL2, boosting Treg responses.263 A similar effect on half-life-extension can be generated through engineering a fusion protein combining IL2 with a protein with improved pharmacodynamics. These fusion proteins can even be based off IL2 muteins, to incorporate multiple improvements in the same molecule. For example, HSA-IL2m is a fusion protein made up of a mutated IL2 (for IL2Rα-biased specificity) and serum albumin,55 to extend half-life.265 The HSA-IL2 mutein even has an extended half-life over the already augmented HSA-IL2 (wildtype) molecule, potentially due to restricted consumption from IL2Rβ binding.55
Half-life extension can also be generated through fusion to non-protein carriers, chiefly polyethylene glycol (PEG). This was first explored in the context of IL2 as early as 1989 by Zimmerman et al, who showed PEG-IL2 to prolong the half-life of IL2 in mouse261 and again, shown by Yang et al in 1991, who reported PEG-IL2 to have a half-life 25 times longer than WT.262 Pegylated-IL2 progressed into clinical trials, with historical trials in cancer and HIV having limited success.258,259,262 Like antibody binding, PEGylation of a molecule can serve as dual purpose, as targeted PEGylation can be used to interfere with a protein-receptor binding interface through the addition of a bulky group to key binding residues. This approach was used by Zhang et al, where they used targeted PEGylation to add 20kDa PEG molecules to tyrosine 31 and threonine 51 to create dual-31/51-20K.106 These amino acid residues are close to the binding interface with the IL2Rβ subunit of the receptor. As a result, the selectivity of dual-31/51-20K, shown as EC50 ratio of CD8 T cell/Treg cells in pSTAT5 assays, was increased almost 3-fold, creating a Treg bias response,106 albeit at the cost of a ~30-fold decrease in total bioreactivity.106 The reverse bias can also be introduced through PEGylation, for IL2 molecules moving into the clinical space for the treatment of cancer. Many of them use PEGylation at the IL2Rα binding interface to change binding affinity. THOR-707 is the most progressed mutein, reporting success in multiple clinical trials.205,266 THOR-707 utilises click chemistry to create a site-specific PEGylated IL2 at P65, resulting in a 10-fold decrease in EC50 on Tregs whilst maintaining CD8 responses.212 Clinical trials in a range of cancer subtypes have revealed THOR-707 to have a large clinical safety window, whilst being successful in producing minor-partial responses.
Finally, there are a few other non-protein carriers that have been used to extend IL2 half-life, without introducing a receptor usage bias. B6 is a bioconjugate of IL2 modified with fatty acid molecules using sortase A.254 The addition of fatty-acid moieties helps increase half-life by non-covalent bonding to serum albumin. This technique has been used in a number of FDA-approved drugs including long-acting insulin Levemir where fatty acid groups have been added to lysine.267 Due to the presence of many Lysine residues in IL2, the technique was altered using sortase A to create a single modification site. Clearance of B6 from the serum is 15-fold slower than WT IL2.254 In addition to the beneficial increase in half life, fatty acid molecules also increased the bioavailability of IL2 by increasing the hydrophilicity, giving an added advantage over antibody fusions or pegylated IL2 molecules.254 Using these techniques can make IL2 more favourable for therapeutic use, decreasing the number of injections needed whilst also generating more stable pharmacodynamics.
Engineering of Targeted IL2 Delivery
An additional class of IL2 engineering covers those modifications which seek to overcome off target systemic effects of IL2 therapy by directing IL2 localisation to target tissues, fine-tuning the dose and reducing off-target detrimental effects (Table 4). The classical approach to targeting IL2 to a particular tissue is through antibody fusion, where the antibody provides the localisation signal. For example, Simlukafusp alfa is a fusion between a IL2Rβ-selective IL2v and anti-fibroblast activation protein (FAP) antibody.136 Coupling the IL2v to anti-FAP gives a double pronged approach. The IL2v ensures CD8 selective stimulation whilst the anti-FAP targets this to the tumour specifically. In principle, this should reduce effects in off-target tissues and reduce the amount of agent needed by increasing the local concentration in the tumour only. Simlukafusp has achieved some success in a clinical trial in metastatic cervical cancer patients. When treated in combination with atezolizumab (anti-PD-L1) 44 out of 47 patients had an observed response.160
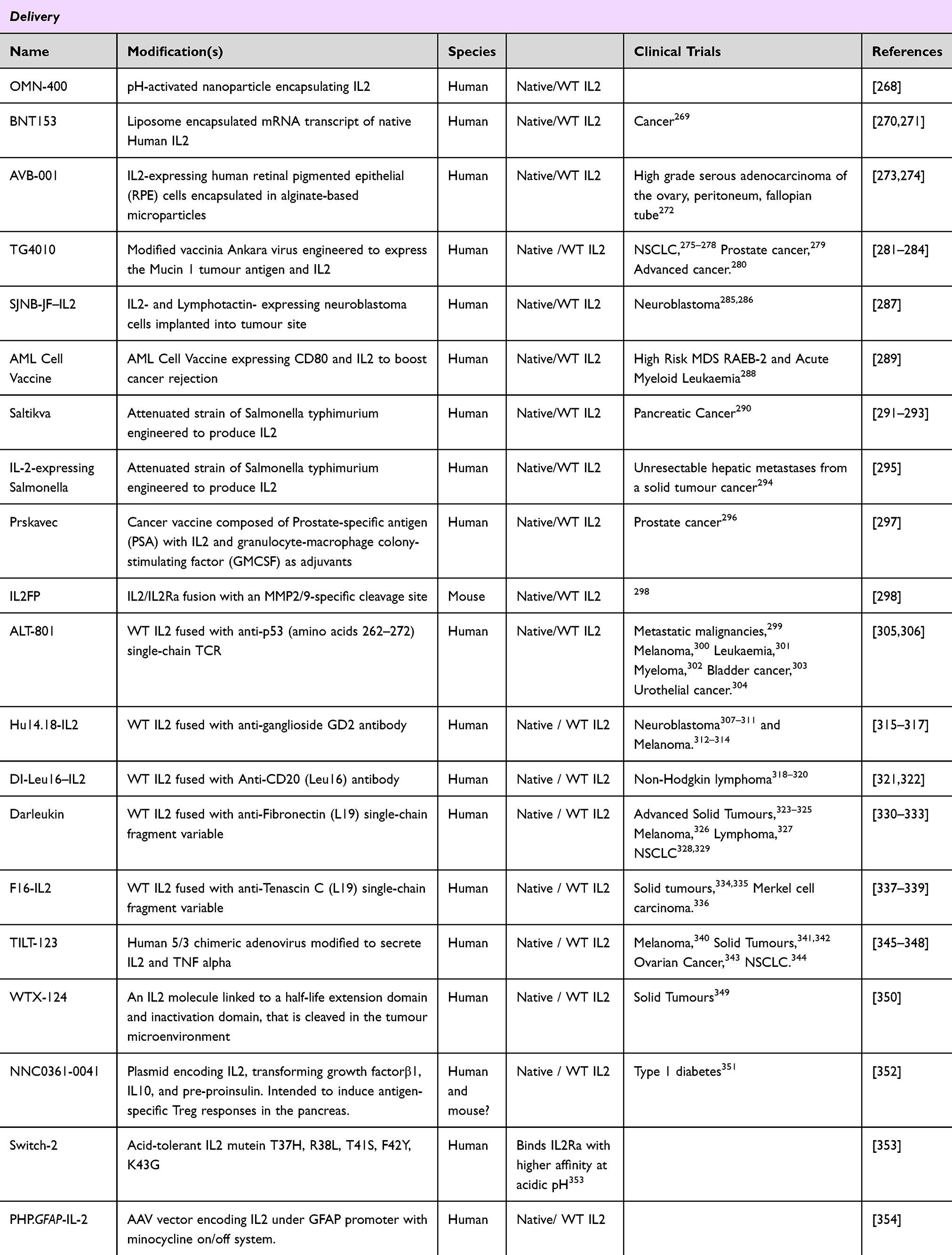 |
Table 4 Delivery Systems for Optimised IL2 Dosing |
An alternative way to target IL2 to tumours is to create fusion proteins which are activated only within the tumour. IL2FP is a fusion protein between mouse IL2 and IL2Rα linked via an MMP2/9 cleavage site. The cleavage site is designed to exploit the dysregulated protease activity in the tumour microenvironment, releasing IL2 from its bound receptor only in the tumour.298 In mouse tumour models, IL2FP was able to be cleaved within the tumour site, resulting in a change in immune composition in the tumour, inducing IFNγ secretion and leading to a reduction in tumour burden and an increase in survival.298
Directed delivery can also take advantage of microenvironmental changes. OMN-400 utilises pH-activated nanoparticles, with the metabolic acidosis of the tumour microenvironment triggering delivery of the IL2 cargo to the tumour.268 At normal physiological pH, the IL2 is held inactive, encapsulated within the nanoparticle. However, in the acidic environment inside the tumour, the nano-particle is denatured, and the IL2 is released. Encapsulation resulted in reduced renal clearance and increased tumour retention of IL2, resulting in a decrease in tumour burden when compared to un-encapsulated IL2.268 In a similar vein, IL2 itself can be engineered to activate in an acidic environment. The Switch-2 IL2 mutein is engineered to have enhanced potency under acidic conditions, aiding tumour rejection with fewer effects in off-target tissues.353
IL2 Muteins with Increased Production
Finally, moving away from modifications, which alter specificity and efficacy of IL2, are muteins which boost production (Table 5). Biological therapeutics are more expensive to produce than their small molecule competitors, making them less accessible to the wider patient population. This is due in part to the expensive processes used in their production, with biological limits on the amount of protein able to be produced in reactors. These limitations arise due to folding constraints that do not exist under the physiological protein concentrations at which these proteins have evolved to be produced—concentrations much lower than those achieved in bioreactors. Often proteins are maladapted to increased production, quickly reaching concentrations where solubility is affected and aggregates form. It is possible to alter production efficiency by mutating IL2. This was first described by Rojas et al, who screened a phage display library of IL2 muteins for those with increased display levels.355 They found a single mutation, K35E, resulted in increased secretion of IL2-containing fusion proteins up to 20-fold.355 Taking a more directed approach, we have recently used SoluBIS, a computational algorithm-based method to predict aggregation-prone linear segments of IL2.43 Human IL2 was found to possess two aggregation-prone regions. Mutations N90R and T131R were predicted to disrupt these regions and help boost production.43 Whilst alone, this was not significant, these mutations did provide rescue to a poorly produced CD8 bias mutein E62K Y45K. By increasing production efficiency using these muteins, the cost of protein production will be proportionally reduced.
 |
Table 5 IL2 Muteins with Increased Production |
Clinical Status
Despite credible advances in the field, no designer IL2 molecules have yet been approved by regulatory authorities for routine clinical use. Several designer molecules have, however, reached clinical trials, largely in the oncology space with IL2Rβγ biased-molecules. Bempegaldesleukin (NKTR-214), a pegylated IL2 variant designed to stimulate inflammatory cells, initially appeared promising, advancing to melanoma and renal carcinoma Phase 3 trials in combination with the anti-PD-1 antibody pembrolizumab. However, after failing to meet primary endpoints for response rates and survival, its development was suspended.357 This may reflect the tumour’s ability to suppress immune responses rather than an inherent failure of the molecule itself, highlighting the broader challenge of improving immune checkpoint inhibitor therapies. Similarly, development of BAY 50–4798, an IL2 variant with an N88R mutation, appears to have been discontinued after Phase 1 trials revealed a short half-life (2 hours) and limited advantages over wild-type IL2.48 The need for frequent dosing and lack of improvements in bioactivity and pharmacokinetics made further development unviable.
Despite limited success of the few designer molecules to complete clinical trials, new IL2 candidates continue to advance into clinical trials. XTX202, an IL-2Rβγ-biased variant, remains inactive until cleaved by matrix metalloproteases in the tumour microenvironment, allowing for localized activation.358 In preclinical non-human primate studies, XTX202 was well tolerated at doses >42-fold higher than IL2 surrogates, suggesting a significantly improved therapeutic index due to its targeted activation.358 As of January 2024, XTX202 is actively progressing through clinical trials, with initial results showing a favourable safety profile and localized tumour activation, making it a strong candidate for combination therapy.359 By integrating localized activation and IL2βγ biasing, XTX202 offers a superior safety profile compared to high-dose IL2 while potentially enhancing anti-tumour immune responses. A plethora of IL2Rα-biased molecules are also available for clinical progression although with a different balance of safety profiles and regulatory barriers compared to the development of IL2Rβγ-based molecules in oncology. While the development of designer IL2 molecules has been a complex and challenging process with several setbacks for first-generation molecules, new multi-strategy approaches and more sophisticated design features offer renewed hope for success.
Future Directions
While classical point mutations and protein complexes have allowed for the discovery of many interesting designer IL2 molecules, new technologies and design strategies continue to build on this to create more sophisticated candidates. A novel approach to creating receptor bias in IL2 is to start from scratch and build de novo synthetic mimetics, using the receptor binding sites as a starting point. NEO-TRA1 is a synthetic molecule developed by Neoleukin Therapeutics, a company which specializes in de novo protein development using computational, artificial intelligence (AI) methods. NEO-TRA1 is designed as an IL2 signalling mimetic, without intrinsic receptor binding capacity, instead being coupled to an anti-IL2Rα antibody for high selectivity for Tregs cells.90 Such de novo design allows for fine tuning of the protein, adding and removing custom binding sites. However, as the protein is non-native, the risk of immunogenicity is increased. NEO-TRA1 may prove an interesting test-case of this trade-off. Taking this approach a step further, Neoleukin has developed a de novo mimetic with enhanced specificity to IL2Rβ. Neoleukin-2/15 is a computationally designed IL2Rα-independent agonist of the IL2 and IL-15 receptors.203 It has a binding site for the IL2Rβ-IL2Rγ heterodimer but not for the IL2Rα subunit. It has four helices – three which are involved in binding and a fourth which holds the first three in place. In vitro, Neoleukin-2/15 stimulates IL2Rα− cells more potently that WT IL2, whilst some potency for IL2Rα+ cells is lost but not completely abolished.203 In vivo, treatment with Neoleukin-2/15 results in an increase in the CD8:Treg ratio, driving a decrease in tumour burden and increase in survival in mouse models of melanoma.203 Importantly, in these preclinical models, there are no signs of immunogenicity, despite the non-native structure, and while the clinical development has been suspended, a lack of immunogenicity was also observed in the Phase 1 clinical trial,360 suggesting the use of de novo proteins may be safer than initially thought. As AI technology improves, it is likely that in silico design of cytokine mimetics will become more common with increased success rates.
Beyond protein engineering, significant advancements are also being made in cytokine delivery systems, including viral vectors and lipid encapsulation. One example of improving the therapeutic properties of IL2 through delivery developed is the use of AAV delivery systems. IL2 can be encoded within an AAV vector, with expression determined by tissue trophism of the capsid. Depending on the capsid and route of delivery, this can cause enrichment within particular tissues, allowing a tissue-biased expansion of Tregs.361,362 The specificity of delivery can be further enhanced by the use of a tissue-specific promoter. We previously utilised this technique to expand Tregs, specifically in the brain. In this example, IL2 is delivered to the mouse brain using the PHB.B capsid and an astrocyte-specific promoter, GFAP.354 An additional benefit of these AAV-delivery approaches is the sustained production of the cytokine, allowing steady levels of IL2 to be achieved without a bolus injection or multiple dosing as well as the inclusion of a druggable switch. As yet, these systems have not reached the clinic. An alternative delivery approach is to administer mRNA encoding proteins rather than the protein itself. BNT151 is a lipid encapsulated mRNA which encodes mutated human IL2Rβ-bias fused with albumin being developed by BioNTech.189 It is currently in phase 1/2a clinical trials in solid tumours, however no outcomes have yet been published. This method provides several advantages over recombinant protein delivery, including controlled cytokine expression, localized activation, and the potential for combination with cancer vaccines and immunotherapies. Additionally, the ability to produce cytokines in vivo rather than relying on large-scale recombinant protein manufacturing offers significant cost and scalability benefits.
The future of IL2-based immunotherapy is likely to be shaped by continued advancements in AI-driven cytokine engineering and next-generation delivery systems, both of which aim to enhance therapeutic efficacy while minimizing toxicity. As novel de novo cytokines and innovative gene delivery technologies continue to emerge, these approaches may overcome many of the limitations associated with traditional cytokine therapies, paving the way for more precise and effective immunomodulatory treatments.
Conclusion
IL2 has proven to be fertile territory for biomodifications. Over a hundred altered molecules, mimics or fusions have been made, with five major design classes targeting different aspects of biology: IL2Rα-selective, IL2Rβ-selective, half-life extenders, tissue-specific delivery agents, and super secreters. Many approaches have been tried in both mouse and human, with mouse analogues being crucial for use as research tools and to explore in vivo effects in cases where human agents do not elicit equivalent cross-reactivity in pre-clinical studies. To achieve these respective changes in biology a multitude of different protein engineering approaches have been applied, including antibody complexing, fusion proteins, point mutations, PEGylation and novel delivery systems, with varying success.
Protein engineering of IL2 has proven to be tricky balancing act, with the native IL2 still the gold standard to beat for clinical efficacy. As seen with many of the aforementioned examples, engineered alterations in specificity often come at a cost of reduced overall bioactivity. As a result, larger quantities of designer IL2 may be needed to reach therapeutic range, increasing cost and the likelihood of off-target effects, mitigating the advantages gained by the enhanced specificity. In addition, the more sophisticated and synthetic approaches such as de novo mimetics may end up enhancing immunogenicity and thus increasing the potential for harmful anti-drug reactions. As more designer IL2 molecules progress further into the clinic, it will be informative to see which molecules provoke such problems.
Currently, although there are many candidates in clinical trials, none of the above designer IL2 molecules have been approved for wide-scale clinical use. The reasons for this are likely to be multifactorial, with a low therapeutic window, improved utility of the native IL2 shifting the goal-posts, and new technologies and approaches rendering first-generation molecules obsolete before they can complete the clinical pipeline. The next generation of IL2 engineering is likely to incorporate multiple design features in a single molecule, improving selectivity and specificity while prolonging half-life and targeting the delivery to the site of action. Such multi-dimensional approaches, harnessing the progress made by different groups, are likely needed to tackle issues of tissue specificity, cellular selectivity, dose titration, production costs, and bio-availability. However, despite the slow progress at the clinical level, the many lessons learned from the incredible protein engineering of IL2 provide a rationale path forward for the modification of other cytokines and immunotherapies.
Disclosure
Dr Amy Dashwood and Professor Adrian Liston report that The University of Cambridge is joint owner of a patent for AAV-based delivery of IL2 and a pending patent application for IL2 muteins, with the authors being potential financial beneficiaries of commercialization. AL and JD are founders of Aila Biotech Ltd. Amy Dashwood reports a patent GB2412771.4 pending to Cambridge Enterprise Limited; Katholieke Universiteit Leuven; VIB VZW; Babraham Institute. The authors report no other conflicts in this work.
References
1. Gillis S, Ferm MM, Ou W, et al. T cell growth factor: parameters of production and a quantitative microassay for activity. J Immunol. 1978;120(6):2027–2032. doi:10.4049/jimmunol.120.6.2027
2. Thayer A. Interleukin-2 wins FDA market clearance. Chem Eng News Archive. 1992;70(19):5.
3. Schwartzentruber DJ. Guidelines for the safe administration of high-dose interleukin-2. J Immunother. 2001;24(4):287–293. doi:10.1097/00002371-200107000-00004
4. Dutcher JP, Schwartzentruber DJ, Kaufman HL, et al. High dose interleukin-2 (Aldesleukin) - expert consensus on best management practices-2014. J ImmunoTher Cancer. 2014;2(1):26. doi:10.1186/s40425-014-0026-0
5. Anderson PM, Sorenson MA. Effects of route and formulation on clinical pharmacokinetics of interleukin-2. Clin Pharmacokinet. 1994;27(1):19–31. doi:10.2165/00003088-199427010-00003
6. Baluna R, Vitetta ES. Vascular leak syndrome: a side effect of immunotherapy. Immunopharmacology. 1997;37(2–3):117–132. doi:10.1016/S0162-3109(97)00041-6
7. Sakaguchi S, Sakaguchi N, Asano M, et al. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol. 1995;155(3):1151–1164. doi:10.4049/jimmunol.155.3.1151
8. Setoguchi R, Hori S, Takahashi T, et al. Homeostatic maintenance of natural Foxp3(+) CD25(+) CD4(+) regulatory T cells by interleukin (IL)-2 and induction of autoimmune disease by IL-2 neutralization. J Exp Med. 2005;201(5):723–735. doi:10.1084/jem.20041982
9. He J, Zhang R, Shao M, et al. Efficacy and safety of low-dose IL-2 in the treatment of systemic lupus erythematosus: a randomised, double-blind, placebo-controlled trial. Ann Rheumatic Dis. 2020;79(1):141–149. doi:10.1136/annrheumdis-2019-215396
10. Humrich JY, Cacoub P, Rosenzwajg M, et al. Low-dose interleukin-2 therapy in active systemic lupus erythematosus (LUPIL-2): a multicentre, double-blind, randomised and placebo-controlled Phase II trial. Ann Rheumatic Dis. 2022;81(12):1685–1694. doi:10.1136/ard-2022-222501
11. Whangbo JS, Kim HT, Mirkovic N, et al. Dose-escalated interleukin-2 therapy for refractory chronic graft-versus-host disease in adults and children. Blood Adv. 2019;3(17):2550–2561. doi:10.1182/bloodadvances.2019000631
12. Matsuoka K, Koreth J, Kim HT, et al. Low-dose interleukin-2 therapy restores regulatory T cell homeostasis in patients with chronic graft-versus-host disease. Sci Transl Med. 2013;5(179):179ra43. doi:10.1126/scitranslmed.3005265
13. Rosenzwajg M, Lorenzon R, Cacoub P, et al. Immunological and clinical effects of low-dose interleukin-2 across 11 autoimmune diseases in a single, open clinical trial. Ann Rheum Dis. 2019;78(2):209–217. doi:10.1136/annrheumdis-2018-214229
14. Wang X, Rickert M, Garcia KC. Structure of the QUATERNARY COMPLEX OF INTERLEUKIN-2 WITH ITS Α, ß, AND ΓC RECEptors. Science. 2005;310(5751):1159–1163. doi:10.1126/science.1117893
15. Stauber DJ, Debler EW, Horton PA, et al. Crystal structure of the IL-2 signaling complex: paradigm for a heterotrimeric cytokine receptor. Proc Natl Acad Sci. 2006;103(8):2788–2793. doi:10.1073/pnas.0511161103
16. Hernandez R, Põder J, LaPorte KM, et al. Engineering IL-2 for immunotherapy of autoimmunity and cancer. Nat Rev Immunol. 2022;22(10):614–628. doi:10.1038/s41577-022-00680-w
17. Raeber ME, Sahin D, Karakus U, Boyman O. A systematic review of interleukin-2-based immunotherapies in clinical trials for cancer and autoimmune diseases. eBioMedicine. 2023;90:1.
18. Spangler JB, Moraga I, Mendoza JL, et al. Insights into cytokine–receptor interactions from cytokine engineering. Ann Rev Immunol. 2015;33(1):139–167. doi:10.1146/annurev-immunol-032713-120211
19. Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol. 2003;4(4):330–336. doi:10.1038/ni904
20. Pierson W, Cauwe B, Policheni A, et al. Antiapoptotic Mcl-1 is critical for the survival and niche-filling capacity of Foxp3+ regulatory T cells. Nat Immunol. 2013;14(9):959–965. doi:10.1038/ni.2649
21. Liston A, Gray DH. Homeostatic control of regulatory T cell diversity. Nat Rev Immunol. 2014;14(3):154–165. doi:10.1038/nri3605
22. Damoiseaux J. The IL-2 – IL-2 receptor pathway in health and disease: the role of the soluble IL-2 receptor. Clin Immunol. 2020;218:108515. doi:10.1016/j.clim.2020.108515
23. Vanmaris RM, Rijkers GT. Biological role of the soluble interleukin-2 receptor in sarcoidosis. Sarcoidosis. Sarcoidosis Vasc Diffuse Lung Dis. 2017;34(2):122–129.
24. Janik JE, Morris JC, Pittaluga S, et al. Elevated serum-soluble interleukin-2 receptor levels in patients with anaplastic large cell lymphoma. Blood. 2004;104(10):3355–3357. doi:10.1182/blood-2003-11-3922
25. Kaya H, Kaji M, Usuda D. Soluble interleukin-2 receptor levels on admission associated with mortality in coronavirus disease 2019. Int J Infect Dis. 2021;105:522–524. doi:10.1016/j.ijid.2021.03.011
26. Trotta E, Bessette PH, Silveria SL, et al. A human anti-IL-2 antibody that potentiates regulatory T cells by a structure-based mechanism. Nat Med. 2018;24(7):1005–1014. doi:10.1038/s41591-018-0070-2
27. VanDyke D, Iglesias M, Tomala J, et al. Engineered human cytokine/antibody fusion proteins expand regulatory T cells and confer autoimmune disease protection. Cell Rep. 2022;41(3):111478. doi:10.1016/j.celrep.2022.111478
28. Spangler JB, Tomala J, Luca V, et al. Antibodies to interleukin-2 elicit selective T cell subset potentiation through distinct conformational mechanisms. Immunity. 2015;42(5):815–825. doi:10.1016/j.immuni.2015.04.015
29. Tomala J, Weberova P, Tomalova B, et al. IL-2/JES6-1 mAb complexes dramatically increase sensitivity to LPS through IFN-γ production by CD25+Foxp3- T cells. eLife. 2021;10:e62432. doi:10.7554/eLife.62432
30. Wilson MS, Pesce JT, Ramalingam TR, et al. Suppression of murine allergic airway disease by IL-2: anti-IL-2 monoclonal antibody-induced regulatory T cells. J Immunol. 2008;181(10):6942–6954. doi:10.4049/jimmunol.181.10.6942
31. Lin Y, Wang X, Qin Y, et al. A single-agent fusion of human IL-2 and anti-IL-2 antibody that selectively expands regulatory T cells. Commun Biol. 2024;7(1):299. doi:10.1038/s42003-024-05987-z
32. Khoryati L, Pham MN, Sherve M, et al. An IL-2 mutein engineered to promote expansion of regulatory T cells arrests ongoing autoimmunity in mice. Sci Immunol. 2020;5(50). doi:10.1126/sciimmunol.aba5264.
33. Chen AC, Cai X, Li C, et al. A treg-selective IL-2 mutein prevents the formation of factor VIII inhibitors in hemophilia mice treated with factor VIII gene therapy. Front Immunol. 2020;11:638. doi:10.3389/fimmu.2020.00638
34. Khoryati L, Pham MN, Sherve M, et al. Regulatory T cell expansion by a highly CD25-dependent IL-2 mutein arrests ongoing autoimmunity. bioRxiv. 2020;2019:862789.
35. Kishimoto TK, Fournier M, Michaud A, et al. Synergistic activity of IL-2 mutein with tolerogenic ImmTOR nanoparticles leads to massive expansion of antigen-specific Tregs and protection against autoimmune disease. bioRxiv. 2023;2023:202–205.
36. Jamison BL, Campbell DJ. An IL-2 mutein promotes Foxp3+ Treg-mediated suppression of dendritic cell activation in response to inflammatory stimuli. J Immunol. 2022;208(1_Supplement):
37. Peterson LB, Bell CJM, Howlett SK, et al. A long-lived IL-2 mutein that selectively activates and expands regulatory T cells as a therapy for autoimmune disease. J Autoimmun. 2018;95:1–14. doi:10.1016/j.jaut.2018.10.017
38. Wu W, Chia T, Lu J, et al. IL-2Rα-biased agonist enhances antitumor immunity by invigorating tumor-infiltrating CD25+CD8+ T cells. Nat Cancer. 2023;4(9):1309–1325. doi:10.1038/s43018-023-00612-0
39. Lu DR, Wu H, Driver I, et al. Dynamic changes in the regulatory T-cell heterogeneity and function by murine IL-2 mutein. Life Sci Alliance. 2020;3(5):e201900520. doi:10.26508/lsa.201900520
40. Pham MN, Khoryati L, Jamison BL, et al. In vivo expansion of antigen-specific regulatory T cells through staggered Fc.IL-2 mutein dosing and antigen-specific immunotherapy. ImmunoHorizons. 2021;5(9):782–791. doi:10.4049/immunohorizons.2100051
41. Rao BM, Driver I, Lauffenburger DA, et al. High-affinity CD25-binding IL-2 mutants potently stimulate persistent T cell growth. Biochemistry. 2005;44(31):10696–10701. doi:10.1021/bi050436x
42. Liu DV, Maier LM, Hafler DA, et al. Engineered interleukin-2 antagonists for the inhibition of regulatory T cells. J Immunother. 2009;32(9):887–894. doi:10.1097/CJI.0b013e3181b528da
43. Dashwood A, Makuyana N, van der Kant R, et al. Directed disruption of IL2 aggregation and receptor binding sites produces designer biologics with enhanced specificity and improved production capacity. Comput Struct Biotechnol J. 2025;27:1112–1123. doi:10.1016/j.csbj.2025.03.002
44. Rao BM, Girvin AT, Ciardelli T, et al. Interleukin-2 mutants with enhanced alpha-receptor subunit binding affinity. Protein Eng. 2003;16(12):1081–1087. doi:10.1093/protein/gzg111
45. Glassman CR, Su L, Majri-Morrison SS, et al. Calibration of cell-intrinsic interleukin-2 response thresholds guides design of a regulatory T cell biased agonist. Elife. 2021;10:e65777.
46. Shanafelt AB, Lin Y, Shanafelt M-C, et al. A T-cell-selective interleukin 2 mutein exhibits potent antitumor activity and is well tolerated in vivo. Nat Biotechnol. 2000;18(11):1197–1202. doi:10.1038/81199
47. Safety and efficacy of BAY 50-4798 in patients with HIV infection. Available from: https://classic.clinicaltrials.gov/show/NCT00059462.
48. Margolin K, Atkins MB, Dutcher JP, et al. Phase I Trial of BAY 50-4798, an interleukin-2–specific agonist in advanced melanoma and renal cancer. Clin Cancer Res. 2007;13(11):3312–3319. doi:10.1158/1078-0432.CCR-06-1341
49. Matthews L, Chapman S, Ramchandani MS, et al. BAY 50-4798, a novel, high-affinity receptor-specific recombinant interleukin-2 analog, induces dose-dependent increases in CD25 expression and proliferation among unstimulated, human peripheral blood mononuclear cells in vitro. Clin Immunol. 2004;113(3):248–255. doi:10.1016/j.clim.2004.07.009
50. Steppan S, Eckart MR, Bajsarowicz K, et al. Reduced secondary cytokine induction by BAY 50-4798, a high-affinity receptor-specific interleukin-2 analog. J Interferon Cytokine Res. 2006;26(3):171–178. doi:10.1089/jir.2006.26.171
51. Steppan S, Kupfer K, Mayer A, et al. Genome wide expression profiling of human peripheral blood mononuclear cells stimulated with BAY 50-4798, a novel T cell selective interleukin-2 analog. J Immunother. 2007;30(2):150–168. doi:10.1097/01.cji.0000211320.07654.f1
52. Andreas W, Paulsen D, Werner S, et al. The T cell-selective IL-2 mutant AIC284 mediates protection in a rat model of multiple sclerosis. J Neuroimmunol. 2015;282:63–72. doi:10.1016/j.jneuroim.2015.03.020
53. Davey RT, Pertel PE, Benson A. Safety, tolerability, pharmacokinetics, and efficacy of an interleukin-2 agonist among HIV-infected patients receiving highly active antiretroviral therapy. J Interferon Cytokine Res. 2008;28(2):89–100. doi:10.1089/jir.2007.0064
54. ModernaTX I. A study to evaluate the safety, tolerability, pharmacokinetics, and pharmacodynamics of mRNA-6231 in healthy adults. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04916431.
55. de Picciotto S, DeVita N, Hsiao CJ, et al. Selective activation and expansion of regulatory T cells using lipid encapsulated mRNA encoding a long-acting IL-2 mutein. Nat Commun. 2022;13(1):3866. doi:10.1038/s41467-022-31130-9
56. Celgene. A safety study of CC-92252 in healthy adult subjects and adult subjects with psoriasis. 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03971825.
57. Padutsch T, Wang Z, Peng Y, et al. Superior treg-expanding properties of a novel dual-acting cytokine fusion protein. Front Pharmacol. 2019;10:10. doi:10.3389/fphar.2019.00010
58. Xencor I, I.C. Research. Safety, tolerability and PK of a single subcutaneous injection of XmAb27564 in healthy volunteers. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04857866.
59. Xencor I. Multiple ascending-dose study of XmAb®27564 in patients with psoriasis or atopic dermatitis. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT06005792.
60. Amgen. Study of efavaleukin alfa in healthy Chinese, Japanese, and Caucasian participants. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04987333.
61. Amgen. Safety, tolerability, pharmacokinetics, pharmacodynamics, and immunogenicity of efavaleukin alfa in participants with systemic lupus erythematosus. 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03451422.
62. Amgen. Efficacy and safety of efavaleukin alfa in subjects with active systemic lupus erythematosus. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04680637.
63. Amgen. Safety and efficacy of efavaleukin alfa in subjects with steroid refractory chronic graft versus host disease. 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03422627.
64. Amgen. Safety and efficacy of efavaleukin alfa in subjects with active rheumatoid arthritis. 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03410056.
65. Amgen. Safety and efficacy of efavaleukin alfa in participants with moderately to severely active ulcerative colitis. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT04987307.
66. Amgen. Long-term safety and efficacy of efavaleukin alfa in participants with moderately to severely active ulcerative colitis. 2023. Available from: https://classic.clinicaltrials.gov/show/NCT05672199.
67. Sarkar N, Hu X, Tchao N, et al. OP0140 Regulatory T cell defects in sle and therapy with a novel Il-2 mutein: phase 1 clinical results with efavaleukin alfa. Ann Rheumatic Dis. 2023;82(Suppl 1):92–93. doi:10.1136/annrheumdis-2023-eular.369
68. Tchao N, Sarkar N, Hu X, et al. AB0432 Efavaleukin Alfa, a novel il-2 mutein, selectively expands regulatory t cells in patients with sle: final results of a phase 1b multiple ascending dose study. Ann Rheumatic Dis. 2022;81(Suppl 1):1343–1344. doi:10.1136/annrheumdis-2022-eular.2244
69. Gorski KS, Stern J, Hsu Y-H, et al. THU0031 Phenotype of foxp3+ regulatory t-cells expanded by the il-2 mutein, amg 592 in healthy subjects in phase 1, first-in-human study. Ann Rheumatic Dis. 2018;77(Suppl 2):243. doi:10.1136/annrheumdis-2018-eular.7212
70. Sundy JS, Otipoby KL, Higginson-Scott N, et al. AB0282 safety, tolerability and selective expansion of regulatory t cells by a single dose of the novel IL-2 mutein PT101 in a phase 1 study in healthy volunteers. Ann Rheumatic Dis. 2021;80(Suppl 1):1167. doi:10.1136/annrheumdis-2021-eular.1200
71. Sharp M, LLC D. A study of MK-6194 (PT101) in participants with active Ulcerative Colitis (UC) (MK-6194-002). 2021. Available from:https://classic.clinicaltrials.gov/show/NCT04924114.
72. Sharp M, LLC D. Multiple rising dose study of MK-6194 in participants with atopic dermatitis (MK-6194-008). 2022. Available from: https://classic.clinicaltrials.gov/show/NCT05450198.
73. Sharp M, LLC D. A clinical study of MK-6194 for the treatment of vitiligo (MK-6194-007). 2023. Available from: https://classic.clinicaltrials.gov/show/NCT06113328.
74. Sharp M, LLC D. Efficacy and safety of MK-6194 in adult participants with systemic lupus erythematosus (MK-6194-006). 2023. Available from: https://classic.clinicaltrials.gov/show/NCT06161116.
75. Higginson-Scott N, Otipoby KL, Viney J. PT101: a treg selective agonist IL-2 mutein therapy for autoimmunity. J Immunol. 2020;204(1_Supplement):
76. Visweswaraiah J, Sampson E, Petaipimol P, et al. OP0023 generation of PT101, A highly selective Il-2 mutein for treatment of autoimmune diseases. Ann Rheumatic Dis. 2021;80(Suppl 1):13.
77. Ltd. I.B.C. Safety, tolerability and preliminary efficacy of IBI363 in subjects with advanced solid tumors or lymphoma. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT05460767.
78. Ltd. I.B.C. A phase 1, open-label, multicenter, dose escalation study of IBI363 (PD1-IL2m) in subjects with advanced solid malignancies or lymphomas. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT05290597.
79. Hospital, H.P.T. and X.H.o.C.S. University. The efficacy and safety of IBI363 in solid tumors. 2023. Available from: https://classic.clinicaltrials.gov/show/NCT06081907.
80. Ltd. I.B.C. A study of IBI363 in subjects with advanced melanoma. 2023. Available from: https://classic.clinicaltrials.gov/show/NCT06081920.
81. Ltd. I.B.C. A study of IBI363 in subjects with advanced solid malignancies. 2024. Available from: https://classic.clinicaltrials.gov/show/NCT06281678.
82. Roche H-L. Safety, tolerability, pharmacokinetics, and pharmacodynamics of RO7049665 in healthy volunteers. 2017. Available from: https://classic.clinicaltrials.gov/show/NCT03221179.
83. Roche H-L. Effect of RO7049665 on the time to relapse following steroid tapering in participants with Autoimmune Hepatitis (AIH). 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04790916.
84. Roche H-L. Double-blinded, placebo-controlled phase 1b study for safety, PK, efficacy, PD of RO7049665 in patients with Ulcerative Colitis (UC). 2019. Available from: https://classic.clinicaltrials.gov/show/NCT03943550.
85. Merck KGaA D, Germany. Available from: EMD 521873 in advanced solid tumors, MTD finding. 2006. https://classic.clinicaltrials.gov/show/NCT01032681.
86. Merck KGaA D, Germany. EMD 521873 plus radiotherapy in Non Small Cell Lung Cancer (NSCLC). 2009. Available from: https://classic.clinicaltrials.gov/show/NCT00879866.
87. Laurent J, Touvrey C, Gillessen S, et al. T-cell activation by treatment of cancer patients with EMD 521873 (Selectikine), an IL-2/anti-DNA fusion protein. J Transl Med. 2013;11(1):5. doi:10.1186/1479-5876-11-5
88. Gillies SD, Lan Y, Hettmann T, et al. A low-toxicity IL-2–based immunocytokine retains antitumor activity despite its high degree of il-2 receptor selectivity. Clin Cancer Res. 2011;17(11):3673–3685. doi:10.1158/1078-0432.CCR-10-2921
89. Ltd, K. Kymab to present update on lead immunocytokine program KY1043 at two European scientific meetings in November 2019. 2019.
90. Chen A, Tampella G, Quijano-Rubio A, et al. NEO-TRA1: a CD25-targeted de novo non-alpha agonist of the il-2 receptor selectively expands regulatory T cells. Blood. 2022;140(Supplement 1):1652–1653. doi:10.1182/blood-2022-159254
91. Ikeda M, Yamaguchi S, Murakami M, et al. OP0008 A novel site-specific pegylated IL-2 with potent and treg-selective activity in vivo. Ann Rheumatic Dis. 2022;81(Suppl 1):5. doi:10.1136/annrheumdis-2022-eular.369
92. Masahiro I, Murakamim YS, Hideyuki O, et al. IL-2 VARIANT. In: PubChem. European Patent Office: KYOWA KIRIN CO LTD; 2017.
93. Company ELA and N. Therapeutics. A study of NKTR-358 (LY3471851) in healthy participants. 2017. Available from: https://classic.clinicaltrials.gov/show/NCT04380324.
94. Company, E.L.a. and N. Therapeutics. A phase I study of LY3471851 in healthy participants. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT05565729.
95. Company, E.L.a. and N. Therapeutics. A study of LY3471851 in healthy participants. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT04133116.
96. Company, E.L.a. and N. Therapeutics. A study of NKTR-358 (LY3471851) in participants with Systemic Lupus Erythematosus (SLE). 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03556007.
97. Company, E.L.a. and N. Therapeutics. A study of LY3471851 in adults with Systemic Lupus Erythematosus (SLE). 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04433585.
98. Company, E.L.a. and N. Therapeutics. A study of LY3471851 in Participants With Psoriasis. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT04119557.
99. Company, E.L.a. and N. Therapeutics. A study of LY3471851 in Participants With Eczema. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT04081350.
100. Company, E.L.a. and N. Therapeutics. A Study of Ly3471851 in adult participants with moderately to severely active Ulcerative Colitis (UC). 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04677179.
101. Therapeutics, N. A Phase 2b study to evaluate rezpegaldesleukin (rezpeg) in the treatment of severe to very severe alopecia areata in adult patients (Rezolve AA). 2024. Available from: https://classic.clinicaltrials.gov/show/NCT06340360.
102. Therapeutics, N. A phase 2b study to evaluate rezpegaldesleukin (rezpeg) in the treatment of adult patients with moderate-to-severe atopic dermatitis. 2023. Available from: https://classic.clinicaltrials.gov/show/NCT06136741.
103. Dixit N, Fanton C, Langowski JL, et al. NKTR-358: a novel regulatory T-cell stimulator that selectively stimulates expansion and suppressive function of regulatory T cells for the treatment of autoimmune and inflammatory diseases. J Trans Autoimmun. 2021;4:100103. doi:10.1016/j.jtauto.2021.100103
104. NEKTAR®. Rezpegaldesleukin (REZPEG/NKTR-358). [cited 2023 04/09/2023]; Available from: https://www.nektar.com/pipeline/rd-pipeline/nktr-358.
105. Christie F, Furie R, Chindalore V, et al. Selective expansion of regulatory T cells by NKTR-358 in healthy volunteers and patients with systemic lupus erythematosus. J Trans Autoimmun. 2022;5:100152. doi:10.1016/j.jtauto.2022.100152
106. Zhang B, Sun J, Wang Y, et al. Site-specific PEGylation of interleukin-2 enhances immunosuppression via the sustained activation of regulatory T cells. Nat Biomed Eng. 2021;5(11):1288–1305. doi:10.1038/s41551-021-00797-8
107. Boyman O, Kovar M, Rubinstein MP, et al. Selective stimulation of T cell subsets with antibody-cytokine immune complexes. Science. 2006;311(5769):1924–1927. doi:10.1126/science.1122927
108. Amit I, Levitin N, Gadrich M, et al. Negative feedback expansion of tregs caused by endogenous IL-2 limits the activity of IL-2-based therapies. J Cancer Immunol. 2023;5(Issue 1):29–39. doi:10.33696/cancerimmunol.5.074
109. A phase 1/2, first-in-human, open label, dose escalation and expansion study of AU-007, a monoclonal antibody that binds to IL-2 and inhibits IL-2Rα binding, in patients with unresectable locally advanced or metastatic cancer. 2022. Available from: https://clinicaltrials.gov/study/NCT05267626.
110. Létourneau S, Van leeuwen EMM, Krieg C, et al. IL-2/anti-IL-2 antibody complexes show strong biological activity by avoiding interaction with IL-2 receptor alpha subunit CD25. Proc Natl Acad Sci U S A. 2010;107(5):2171–2176. doi:10.1073/pnas.0909384107
111. Krieg C, Létourneau S, Pantaleo G, et al. Improved IL-2 immunotherapy by selective stimulation of IL-2 receptors on lymphocytes and endothelial cells. Proc Natl Acad Sci U S A. 2010;107(26):11906–11911. doi:10.1073/pnas.1002569107
112. Kamimura D, Sawa Y, Sato M, et al. IL-2 in vivo activities and antitumor efficacy enhanced by an anti-IL-2 mAb. J Immunol. 2006;177(1):306–314. doi:10.4049/jimmunol.177.1.306
113. Arenas-Ramirez N, Zou C, Popp S, et al. Improved cancer immunotherapy by a CD25-mimobody conferring selectivity to human interleukin-2. Sci Transl Med. 2016;8(367):367ra166. doi:10.1126/scitranslmed.aag3187
114. Caudana P, Núñez NG, De la Rochere P, et al. IL2/anti-IL2 complex combined with CTLA-4, but not PD-1, blockade rescues antitumor NK cell function by regulatory T-cell modulation. Cancer Immunol Res. 2019;7(3):443–457. doi:10.1158/2326-6066.CIR-18-0697
115. Lee JY, Lee E, Hong S-W, et al. TCB2, a new anti-human interleukin-2 antibody, facilitates heterodimeric IL-2 receptor signaling and improves anti-tumor immunity. Oncoimmunology. 2020;9(1):1681869. doi:10.1080/2162402X.2019.1681869
116. Sahin D, Arenas-Ramirez N, Rath M, et al. An IL-2-grafted antibody immunotherapy with potent efficacy against metastatic cancer. Nat Commun. 2020;11(1):6440. doi:10.1038/s41467-020-20220-1
117. Kim J, Lee J-Y, Park S-Y, et al. Crystal structure of human interleukin-2 in complex with TCB2, a new antibody-drug candidate with antitumor activity. Oncoimmunology. 2021;10(1):1899671. doi:10.1080/2162402X.2021.1899671
118. Selecxine. A phase 1/2, open-label, multicenter, dose escalation and expansion study of SLC-3010 monotherapy and in combination. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT05525247.
119. Kim G-A, Kim D-E, Kim Y-J, et al. Abstract 689: strong anti-tumor activity and stability of IL-2/anti IL-2 conjugate SLC-3010 in preclinical experiments. Cancer Res. 2021;81(13_Supplement):689. doi:10.1158/1538-7445.AM2021-689
120. Sun Z, Ren Z, Yang K, et al. A next-generation tumor-targeting IL-2 preferentially promotes tumor-infiltrating CD8+ T-cell response and effective tumor control. Nat Commun. 2019;10(1):3874. doi:10.1038/s41467-019-11782-w
121. Bae J, Liu L, Moore C, et al. IL-2 delivery by engineered mesenchymal stem cells re-invigorates CD8+ T cells to overcome immunotherapy resistance in cancer. Nat Cell Biol. 2022;24(12):1754–1765. doi:10.1038/s41556-022-01024-5
122. Carmenate T, Montalvo G, Lozada SL, et al. The antitumor effect induced by an IL-2 ‘no-alpha’ mutein depends on changes in the CD8(+) T lymphocyte/Treg cell balance. Front Immunol. 2022;13:974188. doi:10.3389/fimmu.2022.974188
123. Carmenate T, Montalvo G, Lozada SL, et al. The antitumor effect induced by an IL-2 ‘no-alpha’ mutein depends on changes in the CD8+ T lymphocyte/Treg cell balance. Front Immunol. 2022;13:974188.
124. Casadesús AV, Cruz BM, Díaz W, et al. Potent immunomodulatory and antitumor effect of anti-CD20-IL2no-alpha tri-functional immunocytokine for cancer therapy. Front Immunol. 2022;13:1021828. doi:10.3389/fimmu.2022.1021828
125. Carmenate T, Pacios A, Enamorado M, et al. Human IL-2 mutein with higher antitumor efficacy than wild type IL-2. J Immunol. 2013;190(12):6230–6238. doi:10.4049/jimmunol.1201895
126. Casadesús AV, Deligne C, Diallo BK, et al. A rationally-engineered IL-2 improves the antitumor effect of anti-CD20 therapy. Oncoimmunology. 2020;9(1):1770565. doi:10.1080/2162402X.2020.1770565
127. Rojas G, Carmenate T, Leon K. Molecular dissection of the interactions of an antitumor interleukin-2-derived mutein on a phage display-based platform. J mol Recog. 2015;28(4):261–268. doi:10.1002/jmr.2440
128. Chen X, Ai X, Wu C, et al. A novel human IL-2 mutein with minimal systemic toxicity exerts greater antitumor efficacy than wild-type IL-2. Cell Death Dis. 2018;9(10):989. doi:10.1038/s41419-018-1047-2
129. Kobayashi M, Kojima K, Murayama K, et al. MK-6, a novel not-α IL-2, elicits a potent antitumor activity by improving the effector to regulatory T cell balance. Cancer Sci. 2021;112(11):4478–4489. doi:10.1111/cas.15127
130. Zhao L, Wang R, Feng C, et al. 1091 CGC-601, a novel βγ-only IL-2 variant, enhances moderate immune activation without Treg expansion, and exhibits a superior safety evidence in vivo. J ImmunoTher Cancer. 2022;10(Suppl 2):A1133–A1134.
131. Sim GC, Liu C, Wang E, et al. IL2 Variant circumvents ICOS+ regulatory T-cell expansion and promotes NK cell activation. Cancer Immunol Res. 2016;4(11):983–994. doi:10.1158/2326-6066.CIR-15-0195
132. Heaton KM, Ju G, Grimm EA. Induction of lymphokine-activated killing with reduced secretion of interleukin-1 beta, tumor necrosis factor-alpha, and interferon-gamma by interleukin-2 analogs. Ann Surg Oncol. 1994;1(3):198–203. doi:10.1007/BF02303524
133. Heaton KM, Ju G, Grimm EA. Human interleukin 2 analogues that preferentially bind the intermediate-affinity interleukin 2 receptor lead to reduced secondary cytokine secretion: implications for the use of these interleukin 2 analogues in cancer immunotherapy. Cancer Res. 1993;53(11):2597–2602.
134. Klein C, Waldhauer I, Nicolini V, et al. Novel Tumor-targeted, engineered IL-2 variant (IL2v)-based immunocytokines for immunotherapy of cancer. Blood. 2013;122(21):2278. doi:10.1182/blood.V122.21.2278.2278
135. An Open-label, Dose Escalation and Dose Expansion Clinical Study to Evaluate the Safety, Tolerability, Pharmacokinetics and Efficacy of STI-7349 in Subjects With Advanced Solid Tumors. 2023 Available from: https://clinicaltrials.gov/study/NCT05978102?term=STI-7349&rank=1
136. Waldhauer I, Gonzalez-Nicolini V, Freimoser-Grundschober A, et al. Simlukafusp alfa (FAP-IL2v) immunocytokine is a versatile combination partner for cancer immunotherapy. mAbs. 2021;13(1):1913791. doi:10.1080/19420862.2021.1913791
137. Klein C, Waldhauer I, Nicolini VG, et al. Cergutuzumab amunaleukin (CEA-IL2v), a CEA-targeted IL-2 variant-based immunocytokine for combination cancer immunotherapy: overcoming limitations of aldesleukin and conventional IL-2-based immunocytokines. OncoImmunology. 2017;6(3):e1277306. doi:10.1080/2162402X.2016.1277306
138. Rafei M, Fidai S, Merchant R, et al. MDNA109: effect of an interleukin-2 superkine on CD8 T-cell properties in the tumor microenvironment. J Clin Oncol. 2019;37(15_suppl):e14220–e14220. doi:10.1200/JCO.2019.37.15_suppl.e14220
139. Suman M, Ring A, Amarnath S, et al. Interleukin-2 activity can be fine tuned with engineered receptor signaling clamps. Immunity. 2015;42(5):826–838. doi:10.1016/j.immuni.2015.04.018
140. Merchant F, To M, Merchant R. Emergence of novel long-acting mono- and bi-specific IL-2/IL-13 superkines as potent immune modulators. Eur J Cancer. 2020;138:S57.
141. Mo F, Yu Z, Li P, et al. An engineered IL-2 partial agonist promotes CD8(+) T cell stemness. Nature. 2021;597(7877):544–548. doi:10.1038/s41586-021-03861-0
142. Levin AM, Bates DL, Ring AM, et al. Exploiting a natural conformational switch to engineer an interleukin-2 ‘superkine’. Nature. 2012;484(7395):529–533. doi:10.1038/nature10975
143. Merchant R, Galligan C, Munegowda MA, et al. Fine-tuned long-acting interleukin-2 superkine potentiates durable immune responses in mice and non-human primate. J Immunother Cancer. 2022;10(1):e003155. doi:10.1136/jitc-2021-003155
144. To MD, Munegowda MA, Merchant R, et al. Abstract 5532: an ‘anti-PD1-IL2 beta-only super-agonist’ displays potent anti-tumor efficacy. Cancer Res. 2022;82(12_Supplement):5532. doi:10.1158/1538-7445.AM2022-5532
145. Sharma A, To MD, Kataria H, et al. Abstract A005: synergistically engaging a b-selective IL-2 agonist with PD1/PDL-1 blockade in a bifunctional superkine, MDNA223. Cancer Immunol Res. 2023;11(12_Supplement):A005–A005. doi:10.1158/2326-6074.TUMIMM23-A005
146. Kim J, Choi J, Kim YY, et al. Abstract 1814: a long-acting and CD122-enhanced IL-2 analog, HM16390, shows a potent and durable anti-tumor effect in both syngeneic B16F10 or CT26 mouse models. Cancer Res. 2023;83(7_Supplement):1814. doi:10.1158/1538-7445.AM2023-1814
147. A phase I, open-label, multicenter, dose escalation and expansion study of HM16390 in patients with advanced or metastatic solid tumors. 2024 Available from: https://clinicaltrials.gov/study/NCT06724016?term=HM16390&rank=1.
148. Kim J, Choi J, Kim Y, et al. 1052 durable anti-tumor effect induced by a long-acting and ‘beta-intensified’ IL-2 mutein, HM16390, in various immunological conditions. J ImmunoTher Cancer. 2023;11(Suppl 1):A1159–A1159.
149. Beig Parikhani A, Bagherzadeh K, Dehghan R, et al. Human IL-2Rɑ subunit binding modulation of IL-2 through a decline in electrostatic interactions: a computational and experimental approach. PLoS One. 2022;17(2):e0264353. doi:10.1371/journal.pone.0264353
150. Mao Z, Fu X, Dong Z, et al. Study of the molecular mechanism of interleukin-2 mutein D10 binding to IL-2 receptors by molecular simulations. mol Simulat. 2018;44(12):973–980. doi:10.1080/08927022.2018.1467013
151. Banerjee A, Li D, Guo Y, et al. A reengineered common chain cytokine augments CD8+ T cell-dependent immunotherapy. JCI Insight. 2022;7(10). doi:10.1172/jci.insight.158889.
152. Hsu EJ, Cao X, Moon B, et al. A cytokine receptor-masked IL2 prodrug selectively activates tumor-infiltrating lymphocytes for potent antitumor therapy. Nat Commun. 2021;12(1):2768. doi:10.1038/s41467-021-22980-w
153. Liu M, Zong J, Liu Z, et al. A novel melittin-MhIL-2 fusion protein inhibits the growth of human ovarian cancer SKOV3 cells in vitro and in vivo tumor growth. Cancer Immunol Immunother. 2013;62(5):889–895. doi:10.1007/s00262-013-1401-2
154. Shao G, QIAN D, WANG H, et al. Construction of the plasmid coding for the expression of the EGFP-M-IL-2(88Arg, 125Ala) fusion protein and the anti‑tumor effects exerted by the fusion protein in HeLa-60 cells Corrigendum in /10.3892/ol.2021.12623. Oncol Lett. 2015;9(6):2729–2735. doi:10.3892/ol.2015.3125
155. Liu M, Wang H, Liu L, et al. Melittin-MIL-2 fusion protein as a candidate for cancer immunotherapy. J Transl Med. 2016;14(1):155. doi:10.1186/s12967-016-0910-0
156. Li L, Qian D, Shao G, et al. High-level expression, purification and study of bioactivity of fusion protein M-IL-2(88Arg, 125Ala) in Pichia pastoris. Protein Expression Purification. 2014;101:99–105.
157. Roche H-L. A study evaluating safety, pharmacokinetics, and therapeutic activity of RO6874281 as a single agent (Part A) or in combination with trastuzumab or cetuximab (Part B or C). 2015. Available from: https://classic.clinicaltrials.gov/show/NCT02627274.
158. Roche H-L. Study to evaluate safety, pharmacokinetics and therapeutic activity of RO6874281 as a combination therapy in participants with unresectable advanced and/or metastatic Renal Cell Carcinoma (RCC). 2017. Available from: https://classic.clinicaltrials.gov/show/NCT03063762.
159. Roche H-L. A study to evaluate safety and therapeutic activity of RO6874281 in combination with pembrolizumab, in participants with advanced or metastatic melanoma. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT03875079.
160. Italiano A, Verlingue L, Prenen H, et al. Clinical activity and safety of simlukafusp alfa, an engineered interleukin-2 variant targeted to fibroblast activation protein-α, combined with atezolizumab in patients with recurrent or metastatic cervical cancer. J Clin Oncol. 2021;39(15_suppl):5510. doi:10.1200/JCO.2021.39.15_suppl.5510
161. Roche H-L. A study of Intravenous (IV) cergutuzumab amunaleukin and atezolizumab in combination in participants with locally advanced and/or metastatic solid tumors. 2015. Available from: https://classic.clinicaltrials.gov/show/NCT02350673.
162. An open-label, multi-center, dose-escalation, phase i study with an expansion phase, to evaluate safety, pharmacokinetics and therapeutic activity of RO6895882, an immunocytokine, consisting of a variant of interleukin-2 (IL-2v) targeting Carcinoembryonic Antigen (CEA) administered intravenously, in patients with advanced and/or metastatic solid tumors. 2013 Available from: https://clinicaltrials.gov/study/NCT02004106.
163. Roche H-L. A study to evaluate safety and anti-tumor activity of eciskafusp alfa (RO7284755) alone or in combination with atezolizumab in participants with advanced and/or metastatic solid tumors. 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04303858.
164. Codarri Deak L, Hashimoto M, Umaña P, et al. Beyond checkpoint inhibition: PD-1 cis-targeting of an IL-2Rβγ-biased interleukin-2 variant as a novel approach to build on checkpoint inhibition. Oncoimmunology. 2023;12(1):2197360. doi:10.1080/2162402X.2023.2197360
165. Tichet M, Wullschleger S, Chryplewicz A, et al. Bispecific PD1-IL2v and anti-PD-L1 break tumor immunity resistance by enhancing stem-like tumor-reactive CD8(+) T cells and reprogramming macrophages. Immunity. 2023;56(1):162–179.e6. doi:10.1016/j.immuni.2022.12.006
166. Umana P. Abstract ND03: PD1-IL2v: a next generation, PD-1-targeted cytokine. Cancer Res. 2021;81(13_Supplement):ND03–ND03. doi:10.1158/1538-7445.AM2021-ND03
167. Codarri Deak L, Nicolini V, Hashimoto M, et al. PD-1-cis IL-2R agonism yields better effectors from stem-like CD8+ T cells. Nature. 2022;610(7930):161–172. doi:10.1038/s41586-022-05192-0
168. Asher Biotherapeutics, I. Sharp M, LLC D. A study evaluating AB248 alone or in combination with pembrolizumab in adult patients with solid tumors. 2023. Available from: https://classic.clinicaltrials.gov/show/NCT05653882.
169. Bio A. AB248: a CD8-TARGETED IL-2 IMMUNOTHERAPY. 2023. Available from: https://asherbio.com/pipeline/ab248/.
170. Moynihan K, Pappas D, Sultan H, et al. 1092 The CD8+ T cell selectivity of AB248 is essential for optimal anti-tumor activity and safety in nonclinical models. J ImmunoTher Cancer. 2022;10(Suppl 2):A1135–A1135.
171. Moynihan KD, Pappas D, Park T, et al. Abstract 3518: AB248 is a CD8+ T cell selective IL-2 designed for superior safety and anti-tumor efficacy. Cancer Res. 2022;82(12_Supplement):3518. doi:10.1158/1538-7445.AM2022-3518
172. Buchbinder EI, Spigel DR, Albany C, et al. Abstract CT250: an open-label, phase 1a/b study of AB248, a CD8+ selective IL-2 mutein fusion protein, alone or in combination with pembrolizumab in patients with advanced solid tumors. Cancer Res. 2023;83(8_Supplement):CT250–CT250. doi:10.1158/1538-7445.AM2023-CT250
173. Vazquez-Lombardi R, Loetsch C, Zinkl D, et al. Potent antitumour activity of interleukin-2-Fc fusion proteins requires Fc-mediated depletion of regulatory T-cells. Nat Commun. 2017;8(1):15373. doi:10.1038/ncomms15373
174. Bio A. AB359: a CD8-Targeted Interleukin-2 (IL-2) immunotherapy for the treatment of chronic viral infections. Available from: https://asherbio.com/pipeline/ab359/.
175. Sulzmaier FJ, Kern N, Ahn SJ, et al. INBRX-120, a CD8α-targeted detuned IL-2 that selectively expands and activates tumoricidal effector cells for safe and durable in vivo responses. J Immunother Cancer. 2023;11(1):e006116.
176. Cheng L, Ye F, Chen S, et al. 1081 exenokine-2: a half-life extended no-α-IL-2 with improved preclinical pharmacological properties supports first-in-human clinical development. J ImmunoTher Cancer. 2022;10(Suppl 2):A1123–A1123.
177. Alkermes I. A study of the effects of ALKS 4230 (nemvaleukin alfa) on subjects with solid tumors. 2016. Available from: https://classic.clinicaltrials.gov/show/NCT02799095.
178. Alkermes I. A dose escalation and cohort expansion study of subcutaneously-administered cytokine ALKS 4230 (Nemvaleukin Alfa) as a single agent and in combination with anti-PD-1 antibody (pembrolizumab) in subjects with select advanced or metastatic solid tumors (ARTISTRY-2). 2019. Available from: https://classic.clinicaltrials.gov/show/NCT03861793.
179. Alkermes I. Less frequent IV dosing & Tumor Microenvironment (TME) study of nemvaleukin alfa (ALKS 4230) monotherapy and in combination with pembrolizumab (ARTISTRY-3). 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04592653.
180. Alkermes I, Network IO. A study of ALKS 4230 (nemvaleukin alfa) with pembrolizumab in head and neck cancer. 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04144517.
181. Alkermes I. Nemvaleukin Alfa (ALKS 4230) monotherapy in patients with advanced cutaneous melanoma or advanced mucosal melanoma - ARTISTRY-6. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04830124.
182. Lopes JE, Fisher JL, Flick HL, et al. ALKS 4230: a novel engineered IL-2 fusion protein with an improved cellular selectivity profile for cancer immunotherapy. J Immunother Cancer. 2020;8(1):e000673.
183. Vaishampayan UN, Tomczak P, Muzaffar J, et al. Nemvaleukin alfa monotherapy and in combination with pembrolizumab in patients (pts) with advanced solid tumors: ARTISTRY-1. J Clin Oncol. 2022;40(16_suppl):2500. doi:10.1200/JCO.2022.40.16_suppl.2500
184. Wang X, Chen G, Nie L, et al. IL-2K35C-moFA, a long-acting engineered cytokine with decreased interleukin 2 receptor α binding, improved the cellular selectivity profile and antitumor efficacy in a mouse tumor model. Cancers. 2022;14(19):4742.
185. ANV419 first in human study phase 1: open-label, dose escalation study of ANV419 as single agent and in combination with ipilimumab in patients with relapsed/refractory advanced solid tumors. 2021 Available from: https://clinicaltrials.gov/study/NCT04855929.
186. A phase 1/2 study of ANV419 as monotherapy or in combination with Anti-PD-1 or Anti-CTLA-4 antibody following anti-PD-1/Anti-PD-L1 antibody treatment in patients with unresectable or metastatic cutaneous melanoma (OMNIA-1). 2022 Available from: https://clinicaltrials.gov/study/NCT05578872?term=ANV419&rank=4.
187. Murer P, Brannetti B, Rondeau J-M, et al. Discovery and development of ANV419, an IL-2/anti-IL-2 antibody fusion protein with potent CD8+ T and natural killer cell-stimulating capacity for cancer immunotherapy. mAbs. 2024;16. doi:10.1080/19420862.2024.2381891
188. Phase I/IIa, first-in-human, open-label, dose escalation trial with expansion cohorts to evaluate safety, pharmacokinetics, pharmacodynamics, and preliminary efficacy of BNT151 as a monotherapy and in combination with other anti-cancer agents in patients with solid tumors. 2020 Available from: https://clinicaltrials.gov/study/NCT04455620?term=%20BNT151&rank=1.
189. Witzel S, B.t. FG, Muik A, et al. Il2 Agonists
190. Medicenna Therapeutics, I. Sharp M, LLC D. A Beta-only IL-2 ImmunoTherapY Study. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT05086692.
191. To M, Merchant N, Coello M, et al. 743 Pharmacokinetic and pharmacodynamic profile of a first-in-human study with MDNA11, an engineered long-acting beta-only IL-2 agonist. J ImmunoTher Cancer. 2022;10(Suppl 2):A776–A776.
192. A Phase1, First-in-Human, Open-Label, Dose Escalation and Expansion Study of CUE-101 Monotherapy in second Line or CUE-101 Combination Therapy With Pembrolizumab in First Line Patients With HPV16+ Recurrent/Metastatic HNSCC. 2019 Available from: https://clinicaltrials.gov/study/NCT03978689?term=CUE-101&rank=2.
193. Non-randomized phase 2 trial of three schedules of CUE-101 administered before surgery or definitive chemoradiation therapy in HLA-A*0201 positive patients with locally advanced, HPV16-positive oropharyngeal squamous-cell carcinoma. 2021 Available from: https://clinicaltrials.gov/study/NCT04852328?term=CUE-101&rank=1.
194. Quayle SN, Girgis N, Thapa DR, et al. CUE-101, a novel E7-pHLA-IL2-Fc fusion protein, enhances tumor antigen-specific T-cell activation for the treatment of HPV16-driven malignancies. Clin Cancer Res. 2020;26(8):1953–1964. doi:10.1158/1078-0432.CCR-19-3354
195. A Phase 1/2, Open-Label, Dose-Escalation, and Expansion Study to Evaluate Safety, Tolerability, Pharmacokinetics, and Therapeutic Activity of GI-101 as a Single Agent and in Combination With Pembrolizumab, Lenvatinib or Local Radiotherapy in Patients With Advanced or Metastatic Solid Tumors (Keynote B59). 2021 Availble from: https://clinicaltrials.gov/study/NCT04977453?term=GI-101&rank=1.
196. Pyo K-H, Koh YJ, Synn C-B, et al. Abstract 1826: comprehensive preclinical study on GI-101, a novel CD80-IgG4-IL2 variant protein, as a therapeutic antibody candidate with bispecific immuno-oncology target. Cancer Res. 2021;81(13_Supplement):1826. doi:10.1158/1538-7445.AM2021-1826
197. An open-label, multicenter, dose escalation and expansion phase 1/2 study to evaluate the safety, tolerability and pharmacokinetics, and anti-tumor activity of GI-102, a CD80-IgG4 Fc-IL-2v bispecific fusion protein, as a single agent and in combination with conventional anti-cancer drugs, pembrolizumab or Trastuzumab Deruxtecan(T-DXd) in patients with advanced or Metastatic Solid Tumors (KEYNOTE-G08). 2023 Available from: https://clinicaltrials.gov/study/NCT05824975?term=GI-102&rank=1.
198. A first-in-human, multicenter, phase 1/2, open-label study of XTX202 in Patients With Advanced Solid Tumors. 2021 Available from: https://clinicaltrials.gov/study/NCT05052268?term=XTX202&rank=1.
199. O’Neil J, Guzman W, Yerov O, et al. Tumor-selective activity of XTX202, a protein-engineered IL-2, in mice without peripheral toxicities in nonhuman primates. J Clin Oncol. 2021;39(15_suppl):1.
200. McKean M, Powderly JD, Duncan M, et al. A first-in-human, multicenter, phase 1/2, open-label study of XTX202, a masked and tumor-selective recombinant human interleukin-2 (IL-2) protein, in patients with advanced solid tumors. J Clin Oncol. 40(16_suppl):1.
201. Huard J, Tatalick L, Walkey C, Swanson R. NL-201, a de novo agonist of IL-2 and IL-15 receptors, demonstrates synergistic antitumor activity with anti-PD-1 checkpoint inhibitor therapy in a preclinical non-Hodgkin lymphoma model. Blood. 2021;138(Supplement 1):4560.
202. Neoleukin Therapeutics, I. Sharp M, LLC D. NL-201 monotherapy and in combination with pembrolizumab in patients with relapsed or refractory cancer. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04659629.
203. Silva D-A, Yu S, Ulge UY, et al. De novo design of potent and selective mimics of IL-2 and IL-15. Nature. 2019;565(7738):186–191. doi:10.1038/s41586-018-0830-7
204. Quijano-Rubio A, Bhuiyan AM, Yang H, et al. A split, conditionally active mimetic of IL-2 reduces the toxicity of systemic cytokine therapy. Nature Biotechnol. 2023;41(4):532–540. doi:10.1038/s41587-022-01510-z
205. Janku F, Abdul-Karim R, Azad A, et al. Abstract LB041: THOR-707 (SAR444245), a novel not-alpha IL-2 as monotherapy and in combination with pembrolizumab in advanced/metastatic solid tumors: interim results from HAMMER, an open-label, multicenter phase 1/2 Study. Cancer Res. 2021;81(13_Supplement):LB041–LB041. doi:10.1158/1538-7445.AM2021-LB041
206. Sanofi. A study of SAR444245 combined with cemiplimab for the treatment of participants with various advanced skin cancers (pegathor skin 201). 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04913220.
207. A Study of SAR444245 Combined With Other Anticancer Therapies for the Treatment of Participants With HNSCC (Master Protocol) (Pegathor Head and Neck 204). 2021. Available from: https://classic.clinicaltrials.gov/show/NCT05061420.
208. Sanofi MS, LLC D. A study of SAR444245 combined with other anticancer therapies for the treatment of participants with gastrointestinal cancer (master protocol) (pegathor gastrointestinal 203). 2021. Available from: https://classic.clinicaltrials.gov/show/NCT05104567.
209. Sanofi, Sharp M, LLC D. A study of SAR444245 with or without other anticancer therapies for the treatment of adults and adolescents with relapsed or refractory B cell lymphoma (master protocol) pegathor lymphoma 205. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT05179603.
210. Sanofi MS, LLC D. A study of SAR444245 combined with other anticancer therapies for the treatment of participants with lung cancer or mesothelioma (pegathor lung 202). 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04914897.
211. Synthorx I. A sanofi company, A study evaluating safety and therapeutic activity of THOR-707 in adult subjects with advanced or metastatic solid tumors (THOR-707-101). 2019. Available from: https://classic.clinicaltrials.gov/show/NCT04009681.
212. Ptacin JL, Caffaro CE, Ma L, et al. An engineered IL-2 reprogrammed for anti-tumor therapy using a semi-synthetic organism. Nat Commun. 2021;12(1):4785. doi:10.1038/s41467-021-24987-9
213. University of Wisconsin, M. and N. Therapeutics. Bempegaldesleukin (NKTR-214) with radiation and anti-PD-1 immunotherapy for head and neck squamous cell carcinoma. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04936841.
214. Therapeutics, N. and Squibb B-M. A single-arm study of bempegaldesleukin (NKTR-214) plus nivolumab in cisplatin ineligible patients who have locally advanced or metastatic urothelial cancer. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT03785925.
215. Therapeutics, N. and Squibb B-M. A study of bempegaldesleukin (NKTR-214: BEMPEG) in combination with nivolumab compared with the investigator’s choice of a Tyrosine Kinase Inhibitor (TKI) therapy (either sunitinib or cabozantinib monotherapy) for advanced metastatic Renal Cell Carcinoma (RCC). 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03729245.
216. Therapeutics, N. and Squibb B-M. A dose escalation and cohort expansion study of NKTR-214 in combination with nivolumab and other anti-cancer therapies in patients with select advanced solid tumors. 2016. Available from: https://classic.clinicaltrials.gov/show/NCT02983045.
217. Therapeutics, N. and ImaginAb I. A CD8 positron emission tomography with computed tomography (PET/CT) study using 89Zr Df-IAB22M2C in patients with metastatic melanoma receiving bempegaldesleukin (NKTR-214) and nivolumab. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT04955262.
218. Therapeutics, N. REVEAL study of NKTR-262 in combination with NKTR-214 and nivolumab in patients with locally advanced metastatic solid tumor malignancies. 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03435640.
219. Therapeutics, N. A phase 1/2 multicenter dose escalation and expansion study of NKTR-214 in subjects with locally advanced or metastatic solid tumors. 2015. Available from: https://classic.clinicaltrials.gov/show/NCT02869295.
220. Squibb B-M, N. Therapeutics, and O.P.C. Ltd. An investigational study of NKTR-214 combined with nivolumab in Japanese participants with advanced solid tumors. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT03745807.
221. Squibb B-M and N. Therapeutics. A study of bempegaldesleukin (BEMPEG: NKTR-214) in combination with nivolumab in children, adolescents and young adults with recurrent or treatment-resistant cancer. 2021. Available from: https://classic.clinicaltrials.gov/show/NCT04730349.
222. Squibb B-M and N. Therapeutics. A study to compare bempegaldesleukin (BEMPEG: NKTR-214) combined with nivolumab and Tyrosine Kinase Inhibitor (TKI) to nivolumab and TKI alone in participants with previously untreated kidney cancer that is advanced or has spread. 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04540705.
223. Squibb B-M and N. Therapeutics. A study of nivolumab plus bempegaldesleukin (bempeg/NKTR-214) vs nivolumab alone vs standard of care in participants with bladder cancer that may have invaded The muscle wall of the bladder and who cannot get cisplatin, a type of medicine given to treat bladder cancer. 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04209114.
224. Squibb B-M and N. Therapeutics. A study of NKTR-214 combined with nivolumab vs nivolumab alone in participants with previously untreated inoperable or metastatic melanoma. 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03635983.
225. Center, M.S.K.C. A Study of NKTR-214 in combination with nivolumab in patients with metastatic and/or locally advanced sarcoma. 2017. Available from: https://classic.clinicaltrials.gov/show/NCT03282344.
226. Calithera Biosciences, I. and N. Therapeutics. A study of TAK-659 in combination with NKTR-214 in participants with advanced non-Hodgkin Lymphoma (NHL). 2019. Available from: https://classic.clinicaltrials.gov/show/NCT03772288.
227. ASA, N.T. N. Therapeutics, and AS V. A study to evaluate safety and efficacy of multiple dosing with VB10.NEO and bempegaldesleukin (NKTR-214) immunotherapy in patients with locally advanced or metastatic cancer. 2018. Available from: https://classic.clinicaltrials.gov/show/NCT03548467.
228. Immunotherapy, P.I.f.C. Platform study for prostate researching translational endpoints correlated to response to inform use of novel combinations. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT03835533.
229. Pfizer. Avelumab with bempegaldesleukin with or without talazoparib or enzalutamide in advanced or metastatic solid tumors. 2019. Available from: https://classic.clinicaltrials.gov/show/NCT04052204.
230. Therapeutics, N. Bempegaldesleukin and pembrolizumab with or without chemotherapy in locally advanced or metastatic solid tumors. 2017. Available from: https://classic.clinicaltrials.gov/show/NCT03138889.
231. Therapeutics, N. BEMPEG with pembrolizumab vs pembrolizumab alone in patients with metastatic or recurrent HNSCC (PROPEL-36). 2022. Available from: https://classic.clinicaltrials.gov/show/NCT04969861.
232. Therapeutics, N and Squibb B-M. Study to compare adjuvant immunotherapy of bempegaldesleukin combined with nivolumab versus nivolumab after complete resection of melanoma in patients at high risk for recurrence. 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04410445.
233. Therapeutics, N. A placebo controlled trial of Bempegaldesleukin (BEMPEG; NKTR-214) with standard of care in patients with mild COVID-19. 2020. Available from: https://classic.clinicaltrials.gov/show/NCT04646044.
234. Charych D, Khalili S, Dixit V, et al. Modeling the receptor pharmacology, pharmacokinetics, and pharmacodynamics of NKTR-214, a kinetically-controlled interleukin-2 (IL2) receptor agonist for cancer immunotherapy. PLoS One. 2017;12(7):e0179431. doi:10.1371/journal.pone.0179431
235. Charych DH, Hoch U, Langowski JL, et al. NKTR-214, an engineered cytokine with biased IL2 receptor binding, increased tumor exposure, and marked efficacy in mouse tumor models. Clin Cancer Res. 2016;22(3):680–690. doi:10.1158/1078-0432.CCR-15-1631
236. Parisi G, Saco JD, Salazar FB, et al. Persistence of adoptively transferred T cells with a kinetically engineered IL-2 receptor agonist. Nat Commun. 2020;11(1):660. doi:10.1038/s41467-019-12901-3
237. Bristol Myers Squibb and nektar announce update on phase 3 PIVOT IO-001 trial evaluating Bempegaldesleukin (BEMPEG) in combination with opdivo (nivolumab) in previously untreated unresectable or metastatic melanoma Bristol Myers Squibb. 2022 Available from: https://news.bms.com/news/details/2022/Bristol-Myers-Squibb-and-Nektar-Announce-Update-on-Phase-3-PIVOT-IO-001-Trial-Evaluating-Bempegaldesleukin-BEMPEG-in-Combination-with-Opdivo-nivolumab-in-Previously-Untreated-Unresectable-or-Metastatic-Melanoma/default.aspx.
238. Rosen DB, Kvarnhammar AM, Laufer B, et al. TransCon IL-2 β/γ: a novel long-acting prodrug with sustained release of an IL-2Rβ/γ-selective IL-2 variant with improved pharmacokinetics and potent activation of cytotoxic immune cells for the treatment of cancer. J Immunother Cancer. 2022;10(7):e004991.
239. A/S, A.P.O.D. and A.P. A/S. A study to investigate safety and tolerability of TransCon IL-2 β/γ alone or in combination with pembrolizumab and/or chemotherapy or TransCon TLR7/8 agonist in adult participants with locally advanced or metastatic solid tumor malignancies. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT05081609.
240. Synthekine, Phase 1a/1b study of STK-012 monotherapy and in combination with pembrolizumab in patients with solid tumors. 2022. Available from: https://classic.clinicaltrials.gov/show/NCT05098132.
241. Emmerich J, Bauer M, Semana M, et al. Abstract 1744: STK-012, an alpha/beta selective IL-2 mutein for the activation of the antigen-activated T cells in solid tumor. Cancer Res. 2021;81(13_Supplement):1744.
242. Spigel D, Spira A, Zamarin D, et al. Abstract CT244: a phase 1a/1b study of STK-012, an α/β IL-2 receptor selective partial agonist as monotherapy and in combination with pembrolizumab in advanced solid tumors (NCT05098132). Cancer Res. 2022;82(12_Supplement):CT244–CT244. doi:10.1158/1538-7445.AM2022-CT244
243. An open-label, multicenter, dose escalation and expansion study of SHR-1916 in subjects with locally advanced or metastatic solid tumor malignancies. 2021 Available from: https://clinicaltrials.gov/study/NCT04842630?term=SHR-1916&rank=1.
244. Jiangsu HengRui Medicine Co. L. A phase 1 dose escalation and expansion study of SHR-1916 in subjects with locally advanced or metastatic solid tumors. cited 2024 https://clinicaltrials.gov/study/NCT04842630?term=shr-1916&rank=1. Available from 2024.
245. A First-in-Human Phase 1 Study of NL-201 Monotherapy and in Combination With Pembrolizumab in Patients With Relapsed or Refractory Cancer. 2020 Available from: https://clinicaltrials.gov/study/NCT04659629?term=NL-201&rank=1.
246. Naing A, Callahan M, Costello B, et al. 509 A first-in-human phase 1 study of NL-201 in patients with relapsed or refractory cancer. Reg Young Invest Award Abstracts. 2021;9(Suppl 2):A540.
247. Sockolosky JT, Trotta E, Parisi G, et al. Selective targeting of engineered T cells using orthogonal IL-2 cytokine-receptor complexes. Science. 2018;359(6379):1037–1042. doi:10.1126/science.aar3246
248. Zhang Q, Hresko ME, Picton LK, et al. A human orthogonal IL-2 and IL-2Rβ system enhances CAR T cell expansion and antitumor activity in a murine model of leukemia. Sci, Trans Med. 2021;13(625):eabg6986. doi:10.1126/scitranslmed.abg6986
249. Hirai T, Ramos TL, Lin P-Y, et al. Selective expansion of regulatory T cells using an orthogonal IL-2/IL-2 receptor system facilitates transplantation tolerance. J Clin Invest. 2021;131(8). doi:10.1172/JCI139991.
250. Ramos TL, Bolivar-Wagers S, Jin S, et al. Prevention of acute GVHD using an orthogonal IL-2/IL-2Rβ system to selectively expand regulatory T cells in vivo. Blood. 2023;141(11):1337–1352. doi:10.1182/blood.2022018440
251. Konrad MW, Hemstreet G, Hersh EM, et al. Pharmacokinetics of recombinant interleukin 2 in humans. Cancer Res. 1990;50(7):2009–2017.
252. Whyte CE, Singh K, Burton OT, et al. Context-dependent effects of IL-2 rewire immunity into distinct cellular circuits. J Exp Med. 2022;219(7):1.
253. Van Gool F, Molofsky AB, Morar MM, et al. Interleukin-5-producing group 2 innate lymphoid cells control eosinophilia induced by interleukin-2 therapy. Blood. 2014;124(24):3572–3576. doi:10.1182/blood-2014-07-587493
254. Qian M, Zhang Q, Lu J, et al. Long-acting human interleukin 2 bioconjugate modified with fatty acids by sortase A. Bioconjugate Chem. 2021;32(3):615–625. doi:10.1021/acs.bioconjchem.1c00062
255. Votavova P, Tomala J, Subr V, et al. Novel IL-2-Poly(HPMA)nanoconjugate based immunotherapy. J Biomed Nanotechnol. 2015;11(9):1662–1673. doi:10.1166/jbn.2015.2114
256. Fallon EM, Liparoto SF, Lee KJ, et al. Increased endosomal sorting of ligand to recycling enhances potency of an interleukin-2 analog. J Biol Chem. 2000;275(10):6790–6797. doi:10.1074/jbc.275.10.6790
257. Yang JC, Topalian SL, Schwartzentruber DJ, et al. The use of polyethylene glycol-modified interleukin-2 (PEG-IL-2) in the treatment of patients with metastatic renal cell carcinoma and melanoma. Cancer. 1995;76(4):687–694. doi:10.1002/1097-0142(19950815)76:4<687::aid-cncr2820760424>3.0.co;2-m
258. Immunomodulation of HIV-1 infected individuals With PEG-Interleukin-2. 1999 Available from: https://clinicaltrials.gov/study/NCT00002017?term=PEG-Interleukin-2&rank=1.
259. Safety and Efficacy of Polyethylene Glycolated IL-2 (PEG IL-2) plus zidovudine in HIV positive, asymptomatic and symptomatic individuals. 1999 Available from: https://clinicaltrials.gov/study/NCT00000664?term=PEG-Interleukin-2&rank=3.
260. Safety and efficacy of Polyethylene Glycolated IL-2 (PEG IL-2) plus zidovudine or dideoxyinosine in HIV positive, asymptomatic and symptomatic individuals. 1999 Available from: https://clinicaltrials.gov/study/NCT00001997?term=PEG-Interleukin-2&rank=4.
261. Zimmerman RJ, Aukerman SL, Katre NV, et al. Schedule dependency of the antitumor activity and toxicity of polyethylene glycol-modified interleukin 2 in murine tumor models. Cancer Res. 1989;49(23):6521–6528.
262. Yang JC, Schwarz SL, Perry-Lalley DM, et al. Murine studies using polyethylene glycol-modified recombinant human interleukin 2 (PEG-IL-2): antitumor effects of PEG-IL2 alone and in combination with adoptive cellular transfer. Lymphokine Cytokine Res. 1991;10(6):475–480.
263. Ward NC, Yu A, Moro A, et al. IL-2/CD25: a long-acting fusion protein that promotes immune tolerance by selectively targeting the IL-2 receptor on regulatory T cells. J Immunol. 2018;201(9):2579–2592. doi:10.4049/jimmunol.1800907
264. Koten JW, Van Luyn MJA, Cadée JA, et al. IL-2 loaded dextran microspheres with attractive histocompatibility properties for local IL-2 cancer therapy. Cytokine. 2003;24(3):57–66. doi:10.1016/S1043-4666(03)00267-9
265. Rogers B, Dong D, Li Z, et al. Recombinant human serum albumin fusion proteins and novel applications in drug delivery and therapy. Curr Pharm Des. 2015;21(14):1899–1907. doi:10.2174/1381612821666150302120047
266. Falchook G, Gan H, Fu S, et al. 481 Phase 1/2 study of THOR-707 (SAR444245), a pegylated recombinant non-alpha IL-2, as monotherapy and in combination with pembrolizumab or cetuximab in patients (pts) with advanced solid tumors. J ImmunoTher Cancer. 2021;9(Suppl 2):A511–A511.
267. Chapman TM, Perry CM. Insulin detemir: a review of its use in the management of type 1 and 2 diabetes mellitus. Drugs. 2004;64(22):2577–2595. doi:10.2165/00003495-200464220-00008
268. Ding X, Miller J, Su Q, et al. 574 ONM-400, a novel approach for interleukin-2 therapy using a ph-activated nanoparticle targeting metabolic acidosis in solid cancers. J ImmunoTher Cancer. 2020;8(Suppl 3):A345–A345.
269. Phase I, first-in-human, open-label, dose escalation trial to evaluate safety, pharmacokinetics, pharmacodynamics, and anti-tumor activity of BNT152+153 in patients with solid tumors. 2021 Available from: https://clinicaltrials.gov/study/NCT04710043?term=BNT152%2B153%20&rank=1.
270. Sahin U, L.k. MV, Diken M, Kreiter S, Tillman B. Treatment using cytokine encoding RNA
271. Beck JD, Diken M, Suchan M, et al. Long-lasting mRNA-encoded interleukin-2 restores CD8+ T cell neoantigen immunity in MHC class I-deficient cancers. Cancer Cell. 2024;42(4):568–582.e11. doi:10.1016/j.ccell.2024.02.013
272. A phase 1/2 open-label, multicenter, dose escalation and expansion study of AVB-001, an intraperitoneally administered, cell-generated, human IL-2 immunotherapy in patients with platinum-resistant, high-grade, serous adenocarcinoma of the ovary, primary peritoneum, or fallopian tube. 2022 Available from: https://clinicaltrials.gov/study/NCT05538624?term=AVB-001&rank=1.
273. Nash AM. Clinically translatable cytokine delivery platform for eradication of intraperitoneal tumors. Sci Adv. 2022-03;8(9):1.
274. Jazaeri A, Carmona G, Nash A, et al. Favorable preclinical efficacy and safety profile of AVB-001, a novel IL-2 cell-based immunotherapy that eradicates ovarian cancer in mouse tumor models and supports first-in-human clinical development (1271). Gynecologic Oncol. 2023;176:S170–S171. doi:10.1016/j.ygyno.2023.06.178
275. A Phase II Study Evaluating the Efficacy and the Safety of First-Line Chemotherapy Combined With TG4010 and Nivolumab in Patients With Advanced Non-Squamous Non-Small Cell Lung Cancer (NSCLC). 2017 Available from: https://clinicaltrials.gov/study/NCT03353675?term=TG4010&rank=5.
276. Phase II Trial of TG4010 Plus Nivolumab in Previously Treated Patients With Metastatic Non-Small Cell Lung Cancer (NSCLC), I. National Cancer, Transgene. 2016 Available from: https://clinicaltrials.gov/study/NCT02823990?term=TG4010&rank=1
277. A Phase IIB/III randomized, double-blind, placebo controlled study comparing first line therapy with or without TG4010 immunotherapy product in patients with stage IV Non-Small Cell Lung Cancer (NSCLC). 2011 Available from: https://clinicaltrials.gov/study/NCT01383148?term=TG4010&rank=2.
278. A phase IIb multicentric controlled study evaluating the therapeutic vaccine TG4010(MVA-MUC1-IL2) as an adjunct to standard chemotherapy in advanced non small cell lung cancer. 2006 Available from: https://clinicaltrials.gov/study/NCT03353675?term=TG4010&rank=5
279. Randomized multicenter phase II study evaluating two dosing schedules of TG4010(MVA-MUC1-IL2) in patients with adenocarcinoma of the prostate, Available from: https://clinicaltrials.gov/study/NCT00040170?term=TG4010&rank=3.
280. Phase I Bridging Trial of TG4010 as Antigen-Specific Immunotherapy in Patients With MUC-1 Positive Advanced Cancer. 2000 Available from: https://clinicaltrials.gov/study/NCT00004881?term=TG4010&rank=6.
281. Tosch C, Bastien B, Barraud L, et al. Viral based vaccine TG4010 induces broadening of specific immune response and improves outcome in advanced NSCLC. J ImmunoTher Cancer. 2017;5(1). doi:10.1186/s40425-017-0274-x.
282. Quoix E, Ramlau R, Westeel V, et al. Therapeutic vaccination with TG4010 and first-line chemotherapy in advanced non-small-cell lung cancer: a controlled phase 2B trial. Lancet Oncol. 2011;12(12):1125–1133. doi:10.1016/S1470-2045(11)70259-5
283. Dreicer R, Stadler WM, Ahmann FR, et al. MVA-MUC1-IL2 vaccine immunotherapy (TG4010) improves PSA doubling time in patients with prostate cancer with biochemical failure. Invest New Drugs. 2008;27(4):379–386. doi:10.1007/s10637-008-9187-3
284. Rochlitz C, Figlin R, Squiban P, et al. Phase I immunotherapy with a modified vaccinia virus (MVA) expressing human MUC1 as antigen‐specific immunotherapy in patients with MUC1‐positive advanced cancer. J Gene Med. 2003;5(8):690–699. doi:10.1002/jgm.397
285. A phase I/II study using allogeneic tumor cell vaccination with oral metronomic cytoxan in patients with high-risk neuroblastoma (ATOMIC), C. center for and B.C.o.M. 2010 Available from: https://clinicaltrials.gov/study/NCT01192555.
286. A phase I/II study of immunization with lymphotactin and interleukin 2 gene modified neuroblastoma tumor cells after high-dose chemotherapy and autologous stem cell rescue in patients with high risk neuroblastoma, C. Center for and B.C.o.M. Gene Therapy. 2008 https://clinicaltrials.gov/study/NCT00703222.
287. Haight AE, Bowman LC, Ng CY, et al. Humoral response to vaccination with interleukin‐2‐expressing allogeneic neuroblastoma cells after primary therapy. Med Pediatr Oncol. 2000;35(6):712–715. doi:10.1002/1096-911x(20001201)35:6<712::aid-mpo50>3.0.co;2-7
288. N.H.S.T. King’s College Hospital. A Phase I Study of B7.1 (CD80)/IL-2 Immune Gene Therapy for High Risk MDS RAEB-2 and Acute Myeloid Leukaemia Patients Unsuitable for Allogeneic Haematological Stem Cell Transplant. ; 2015 Available from: https://clinicaltrials.gov/study/NCT02493829?term=B7.1%20(CD80)%2FIL-2%20&rank=2.
289. Chan L, Hardwick N, Darling D, et al. IL-2/B7.1 (CD80) fusagene transduction of AML blasts by a self-inactivating lentiviral vector stimulates T cell responses in vitro: a strategy to generate whole cell vaccines for AML. mol Ther. 2005;11(1):120–131. doi:10.1016/j.ymthe.2004.09.006
290. A phase 2 study of saltikva (attenuated salmonella typhimurium containing the human gene for interleukin-2) in patients with metastatic pancreatic cancer. 2020 Available from: https://clinicaltrials.gov/study/NCT04589234?term=saltikva&rank=1.
291. Gniadek TJ, Augustin L, Schottel J, et al. A phase I, dose escalation, single dose trial of oral attenuated salmonella typhimurium containing human IL-2 in patients with metastatic gastrointestinal cancers. J Immunother. 2020;43(7):217–221. doi:10.1097/CJI.0000000000000325
292. Saltzman D. Abstract LB161: microbial based immunotherapy: saltikva as a novel therapeutic for solid tumors. Cancer Res. 2021;81(13_Supplement):LB161–LB161. doi:10.1158/1538-7445.AM2021-LB161
293. Kavan P, Saltzman DA, Muegge J, et al. Abstract CT035: addition of Salmonella-IL2 to FOLFIRINOX for metastatic stage 4 pancreatic cancer nearly doubles median survival. Cancer Res. 2023;83(8_Supplement):CT035–CT035. doi:10.1158/1538-7445.AM2023-CT035
294. A phase 1 study of an il-2 expressing, attenuated salmonella typhimurium in patients with unresectable hepatic spread from any non-hematologic primary. 2010 Available from: https://www.clinicaltrials.gov/study/NCT01099631.
295. Masonic Cancer Center, U.o.M. IL-2 expressing, attenuated salmonella typhimurium in unresectable hepatic spread. 2020. Available from: https://www.clinicaltrials.gov/study/NCT01099631#study-record-dates.
296. A phase 2, randomized study of proscavax, a PSA/IL-2/GM-CSF vaccine, in treatment-naive patients with clinically localized prostate cancer versus an active surveillance strategy. 2018 Available from: https://www.clinicaltrials.gov/study/NCT03579654?term=PSA%2FIL-2%2FGM-CSF&rank=2.
297. OncBioMune. ProscaVax for prostate cancer. 2024; Available from: https://oncbiomune.com/proscavax/.
298. Norville K, Skrombolas D, Ferry SL, et al. A protease activatable interleukin-2 fusion protein engenders antitumor immune responses by interferon gamma-dependent and interferon gamma-independent mechanisms. J Interferon Cytokine Res. 2022;42(7):316–328. doi:10.1089/jir.2022.0043
299. Phase I study of ALT-801 in patients with progressive metastatic malignancies. 2007 Available from: https://www.clinicaltrials.gov/study/NCT00496860?term=ALT-801&rank=4.
300. I. National Cancer. Phase Ib/II Study of ALT-801 With Cisplatin in Patients With Metastatic Melanoma. ; 2009 Available from: https://www.clinicaltrials.gov/study/NCT01029873?term=ALT-801&rank=5.
301. M.D.A.C. Center. A Single-Center Open-Label Phase I Study of ALT-801 for Ex Vivo Maturation and in Vivo Retargeting of Haploidentical Natural Killer Cells Delivered Following Fludarabine, Cytarabine, and G-CSF in Patients With Relapsed/Refractory Acute Myeloid Leukemia. 2011 Available from: https://www.clinicaltrials.gov/study/NCT01478074?term=ALT-801&rank=3
302. I. National Cancer. A Study of ALT-801 in Patients With Relapsed or Refractory Multiple Myeloma. 2012 Available from: https://www.clinicaltrials.gov/study/NCT01670994?term=ALT-801&rank=8
303. James and P. Esther King Biomedical Research. A Phase Ib/II Study of ALT-801 in Patients With Bacillus Calmette-Guerin (BCG) Failure Non-Muscle Invasive Bladder Cancer. ; 2012 Available from: https://www.clinicaltrials.gov/study/NCT01625260?term=ALT-801&rank=7.
304. Cancer IN. A Phase Ib/II Trial of ALT-801 in Combination With Cisplatin and Gemcitabine in Muscle Invasive or Metastatic Urothelial Cancer. 2011 Available from: https://www.clinicaltrials.gov/study/NCT01326871?term=ALT-801&rank=9.
305. Card KF, et al. A soluble single-chain T-cell receptor IL-2 fusion protein retains MHC-restricted peptide specificity and IL-2 bioactivity. Cancer Immunol Immunother. 2004;53:1.
306. Fishman MN, Thompson JA, Pennock GK, et al. Phase I trial of ALT-801, an interleukin-2/T-cell receptor fusion protein targeting p53 (aa264–272)/HLA-A*0201 complex, in patients with advanced malignancies. Clin Cancer Res. 2011;17(24):7765–7775. doi:10.1158/1078-0432.CCR-11-1817
307. Cancer IN. A Phase I/IB Intergroup Trial of the HU14.18-IL2 Fusion Protein in Children With Refractory Neuroblastoma and Other GD2 Positive Tumors. 1999 Available from: https://www.clinicaltrials.gov/study/NCT00003750?term=HU14.18-IL2%20&rank=5.
308. Cancer IN. A Phase II Study of hu14.18-IL2 in Children With Recurrent or Refractory Neuroblastoma. 2004 Available from: https://www.clinicaltrials.gov/study/NCT00082758?term=HU14.18-IL2%20&rank=7.
309. Feasibility/Phase II Study of hu14.18-IL2 Immunocytokine + GM-CSF and Isotretinoin in Patients With Relapsed or Refractory Neuroblastoma. ; 2011 Available from: https://clinicaltrials.gov/study/NCT01334515
310. A Safety/Feasibility Trial of the Addition of the Humanized Anti-GD2 Antibody (hu14.18K322A) With and Without Natural Killer Cells to Chemotherapy in Children and Adolescents With Recurrent/Refractory Neuroblastoma. ; 2012 Available from: https://clinicaltrials.gov/study/NCT01576692?term=hu14.18K322A&rank=5.
311. Treatment of Relapsed or Refractory Neuroblastoma and Osteosarcoma With Ex-Vivo Expanded and Activated Haploidentical NK Cells and Hu14.18-IL2. ; 2017 Available from: https://clinicaltrials.gov/study/NCT03209869?term=Hu14.18-IL2&rank=3.
312. Phase II Trial of Hu14.18-IL2 (EMD 273063) in Subjects With Advanced Melanoma.; 2005 Available from: https://clinicaltrials.gov/study/NCT00109863?term=Hu14.18-IL2&rank=4.
313. A Pilot Trial of HU14.18-IL2 (EMD273063) in Subjects With Completely Resectable Recurrent Stage III or Stage IV Melanoma.2007 Available from: https://clinicaltrials.gov/study/NCT00590824?term=Hu14.18-IL2&rank=1.
314. Phase I/II Intratumoral Administration of Hu14.18-IL2, With Local Radiation, Nivolumab and Ipilimumab in Subjects With Advanced Melanoma; 2019 Available from: https://clinicaltrials.gov/study/NCT03958383?term=Hu14.18-IL2&rank=2.
315. Hank JA, J.e.s. JG, Jaeger P, Gillies SD, Sondel PM. Activation of human effector cells by a tumor reactive recombinant anti-ganglioside GD2 interleukin-2 fusion protein (ch14.18-IL2). Clin Cancer Res. 1996;2(12):1951–1959.
316. Shusterman S, London WB, Gillies SD, et al. Antitumor activity of Hu14.18-IL2 in patients with relapsed/refractory neuroblastoma: a Children’s Oncology Group (COG) phase II study. J Clin Oncol. 2010;28(33):4969–4975. doi:10.1200/JCO.2009.27.8861
317. Osenga KL, Hank JA, Albertini MR, et al. A phase i clinical trial of the hu14.18-IL2 (EMD 273063) as a treatment for children with refractory or recurrent neuroblastoma and melanoma: a study of the children’s oncology group. Clin Cancer Res. 2006;12(6):1750–1759. doi:10.1158/1078-0432.CCR-05-2000
318. A Phase I Study of De-Immunized DI-Leu16-IL2 Immunocytokine in Patients With B-Cell Non-Hodgkin Lymphoma. 2008 Available from: https://clinicaltrials.gov/study/NCT00720135?term=di-leu16-il2&rank=3.
319. A phase I/II study of de-immunized di-leu16-il2 immunocytokine administered subcutaneously in patients with B-cell Non-Hodgkin Lymphoma (NHL). 2013 Available from: https://clinicaltrials.gov/study/NCT01874288?term=di-leu16-il2&rank=1.
320. An open-label extension study of de-immunized di-leu16-il2 immunocytokine administered in patients with B-cell Non-Hodgkin Lymphoma (NHL). 2014 Available from: https://clinicaltrials.gov/study/NCT02151903?term=di-leu16-il2&rank=2.
321. Gillies SD, Lan Y, Williams S, et al. An anti-CD20–IL-2 immunocytokine is highly efficacious in a SCID mouse model of established human B lymphoma. Blood. 2005;105(10):3972–3978. doi:10.1182/blood-2004-09-3533
322. Lansigan F, Nakamura R, Quick DP, et al. DI-Leu16-IL2, an anti-CD20-interleukin-2 immunocytokine, is safe and active in patients with relapsed and refractory B-cell lymphoma: a report of maximum tolerated dose, optimal biologic dose, and recommended phase 2 dose. Blood. 2016;128(22):620. doi:10.1182/blood.V128.22.620.620
323. Editor. A Dose Finding Pharmacokinetic Study of the Tumour-Targeting Human L19IL2 Monoclonal Antibody-Cytokine Fusion Protein in Patients With Advanced Solid Tumours. 2010 Available from: https://clinicaltrials.gov/study/NCT01058538?term=L19-IL2&rank=6.
324. Phase I clinical study combining L19-IL2 with stereotactic ablative body radiotherapy in patients with oligometastatic solid tumor. 2013 Available from: https://clinicaltrials.gov/study/NCT02086721?term=L19-IL2&rank=1.
325. A phase I study to evaluate the safety and preliminary signs of efficacy of [177Lu]lu-oncofap-23 alone or in combination with L19-IL2 as a treatment of metastatic FAP-positive solid tumors. 2024 Available from: https://clinicaltrials.gov/study/NCT06640413?term=L19-IL2&rank=7
326. A phase I/II dose escalation study of the tumor-targeting human L19-IL2 monoclonal antibody-cytokine fusion protein in combination with dacarbazine for patients with metastatic melanoma. 2014 Available from: https://clinicaltrials.gov/study/NCT02076646?term=L19-IL2&rank=5
327. A phase I/II study of the tumor-targeting human L19-IL2 monoclonal antibody-cytokine fusion protein in combination with rituximab in relapsed or Refractory diffuse large B-cell lymphoma (DLBCL). 2016 Available from: https://clinicaltrials.gov/study/NCT02957019?term=L19-IL2&rank=3.
328. A randomized phase ii trial of the combination of SBRT with L19-IL2 in patients with limited metastatic Non-small Cell Lung Cancer (NSCLC),2016 Available from: https://clinicaltrials.gov/study/NCT02735850?term=L19-IL2&rank=2.
329. Stereotactic Ablative Body Radiotherapy (SABR) Combined With Immunotherapy (L19-IL2) in Stage IV NSCLC Patients, ImmunoSABR: A Multicentre, Randomised Controlled Open-Label Phase II Trial.2018 Available from: https://clinicaltrials.gov/study/NCT03705403?term=L19-IL2&rank=4.
330. Carnemolla B, Borsi L, Balza E, et al. Enhancement of the antitumor properties of interleukin-2 by its targeted delivery to the tumor blood vessel extracellular matrix. Blood. 2002;99(5):1659–1665. doi:10.1182/blood.V99.5.1659
331. Weide B, et al. A phase II study of the L19IL2 immunocytokine in combination with dacarbazine in advanced metastatic melanoma patients. Cancer Immunology. Immunotherapy. 2019;68(9):2019.
332. Johannsen M, Spitaleri G, Curigliano G, et al. The tumour-targeting human L19-IL2 immunocytokine: preclinical safety studies, phase I clinical trial in patients with solid tumours and expansion into patients with advanced renal cell carcinoma. Eur J Cancer. 2010;46(16):2926–2935. doi:10.1016/j.ejca.2010.07.033
333. Eigentler TK, Weide B, de Braud F, et al. A dose-escalation and signal-generating study of the immunocytokine L19-IL2 in combination with dacarbazine for the therapy of patients with metastatic melanoma. Clin Cancer Res. 2011;17(24):7732–7742. doi:10.1158/1078-0432.CCR-11-1203
334. A Dose-Finding, Pharmacokinetic. Phase Ib/II study of the tumour-targeting human F16IL2 monoclonal antibody-cytokine fusion protein in combination with doxorubicin in patients with advanced solid tumours. 2010 Available from: https://clinicaltrials.gov/study/NCT01131364?term=F16IL2%20&rank=1.
335. Phase Ib/II study of the tumour-targeting human F16IL2 monoclonal antibody-cytokine fusion protein in combination with paclitaxel in patients with advanced solid tumours. 2010 Available from: https://clinicaltrials.gov/study/NCT01134250?term=F16IL2%20&rank=5
336. Gmb HIB. Phase II Study of the Tumor-Targeting Human F16IL2 Monoclonal Antibody-Cytokine Fusion Protein in Combination With Paclitaxel in Patients With Metastatic Merkel Cell Carcinoma. ; 2014 Available from: https://clinicaltrials.gov/study/NCT02054884?term=F16IL2%20&rank=3.
337. Mårlind J, Kaspar M, Trachsel E, et al. Antibody-mediated delivery of interleukin-2 to the stroma of breast cancer strongly enhances the potency of chemotherapy. Clin Cancer Res. 2008;14(20):6515–6524. doi:10.1158/1078-0432.CCR-07-5041
338. Catania C, Maur M, Berardi R, et al. The tumor-targeting immunocytokine F16-IL2 in combination with doxorubicin: dose escalation in patients with advanced solid tumors and expansion into patients with metastatic breast cancer. Cell Adh Migr, 2015.
339. Berdel AF, et al. A phase I trial of the antibody-cytokine fusion protein F16IL2 IN combination with anti-CD33 immunotherapy for posttransplant AML relapse. Blood. 2021;138(Supplement 1).
340. A phase 1, open-label, dose-escalation clinical trial of tumor necrosis factor alpha and interleukin 2 coding oncolytic adenovirus TILT-123 in melanoma patients receiving adoptive cell therapy with tumor infiltrating lymphocytes. 2019 Available from: https://clinicaltrials.gov/study/NCT04217473?term=TILT-123&rank=3.
341. A phase 1, open-label, dose-escalation clinical trial of tumor necrosis factor alpha and interleukin-2 coding oncolytic adenovirus (TILT-123) in patients with injectable solid tumors. 2021 Available from: https://www.clinicaltrials.gov/study/NCT04695327?term=TILT-123&rank=1.
342. A phase I open-label, dose-escalation clinical trial of tumor necrosis factor alpha and IL-2 coding oncolytic adenovirus TILT-123 and avelumab in solid tumor patients (Melanoma and SCCHN) refractory to or progressing after anti-PD(L)1. 2022 Available from: https://www.clinicaltrials.gov/study/NCT05222932?term=TILT-123&rank=4.
343. Merck S, Dohme LLC, Editors. A Two-Part, Phase I/Ib, Open-Label, Dose-Escalation Trial of Tumor Necrosis Factor Alpha and Interleukin-2 Coding Oncolytic Adenovirus (TILT-123) in Combination With Pembrolizumab (Phase I Part) and Pembrolizumab and Pegylated Liposomal Doxorubicin (Phase Ib Part) in Patients With Platinum Resistant or Refractory Ovarian Cancer. 2022.
344. A Phase I Open-Label, Dose-Escalation Trial of Tumor Necrosis Factor Alpha and Interleukin-2 Coding Oncolytic Adenovirus (TILT-123) in Combination With Pembrolizumab in Patients With Immune Checkpoint Inhibitor Refractory Non-Small Cell Lung Cancer. 2023 Available from: https://www.clinicaltrials.gov/study/NCT06125197?term=TILT-123&rank=2.
345. Havunen R, Kalliokoski R, Siurala M, et al. Cytokine-coding oncolytic adenovirus TILT-123 is safe, selective, and effective as a single agent and in combination with immune checkpoint inhibitor anti-PD-1. Cells. 2021;10(2):246. doi:10.3390/cells10020246
346. Jirovec E, et al. Intravenous delivery of oncolytic adenovirus TILT-123 results in systemic tumor transduction and accumulation of lymphocytes. J Clin Oncol. 2024;42(16_suppl).
347. Jirovec E, Quixabeira DCA, Clubb JHA, et al. Single intravenous administration of oncolytic adenovirus TILT-123 results in systemic tumor transduction and immune response in patients with advanced solid tumors. J Exp Clin Cancer Res. 2024;43(1). doi:10.1186/s13046-024-03219-0.
348. Pakola SA, Peltola KJ, Clubb JHA, et al. Safety, efficacy, and biological data of T-cell–enabling oncolytic adenovirus TILT-123 in advanced solid cancers from the TUNIMO monotherapy phase I trial. Clin Cancer Res. 2024;30(17):3715–3725. doi:10.1158/1078-0432.CCR-23-3874
349. Editors. A Multicenter Phase I/Ib Dose Escalation and Expansion Study of WTX-124 as Monotherapy and in Combination With Pembrolizumab in Patients With Selected Advanced or Metastatic Solid Tumors. 2022. Available from: https://www.clinicaltrials.gov/study/NCT05479812?term=WTX-124%20&rank=1.
350. Nirschl CJ, Brodkin HR, Hicklin DJ, et al. Discovery of a conditionally activated IL-2 that promotes antitumor immunity and induces tumor regression. Cancer Immunol Res. 2022;10(5).
351. Nordisk ASN. A Multiple Ascending Dose Trial Investigating Safety, Tolerability and Pharmacokinetics of NNC0361-0041 Administered Subcutaneously to Patients With Type 1 Diabetes Mellitus. 2020. Available from: https://www.clinicaltrials.gov/study/NCT04279613?term=NNC0361-0041&rank=1.
352. PP P, Chaplin J, Wijaranakula M, et al. Multicomponent plasmid protects mice from spontaneous autoimmune diabetes - pubmed. Diabetes. 71(1).
353. Gaggero S, Martinez-Fabregas J, Cozzani A, et al. IL-2 is inactivated by the acidic pH environment of tumors enabling engineering of a pH-selective mutein. Sci Immunol. 2022;7(78):eade5686. doi:10.1126/sciimmunol.ade5686
354. Yshii L, Pasciuto E, Bielefeld P, et al. Astrocyte-targeted gene delivery of interleukin 2 specifically increases brain-resident regulatory T cell numbers and protects against pathological neuroinflammation. Nat Immunol. 2022;23(6):878–891. doi:10.1038/s41590-022-01208-z
355. Rojas G, Carmenate T, Santo-Tomás JF, et al. Directed evolution of super-secreted variants from phage-displayed human interleukin-2. Sci Rep. 2019;9(1):800. doi:10.1038/s41598-018-37280-5
356. Rojas G, Relova-Hernández E, Pérez-Riverón A, et al. Molecular reshaping of phage-displayed interleukin-2 at beta chain receptor interface to obtain potent super-agonists with improved developability profiles. Commun Biol. 2023;6(1):828. doi:10.1038/s42003-023-05188-0
357. Diab A, Gogas H, Sandhu S, et al. Bempegaldesleukin plus nivolumab in untreated advanced melanoma: the open-label, phase III PIVOT IO 001 trial results. J Clin Oncol. 2023;41(30):4756–4767. doi:10.1200/JCO.23.00172
358. O’Neil J, Guzman W, Yerov O, et al. Tumor-selective activity of XTX202, a protein-engineered IL-2, in mice without peripheral toxicities in nonhuman primates. J Clin Oncol. 2021;39(15_suppl):2563. doi:10.1200/JCO.2021.39.15_suppl.2563
359. Chahoud J, Zakharia Y, McKean M, et al. Phase 1/2 study of XTX202, a tumor-activated IL-2βγ, in advanced solid tumors. J Clin Oncol. 2024;42(16_suppl):2595. doi:10.1200/JCO.2024.42.16_suppl.2595
360. Neoleukin Therapeutics, I. Neoleukin therapeutics announces third quarter 2022 financial results and corporate update. 2022. Available from: https://investor.neoleukin.com/news-releases/news-release-details/neoleukin-therapeutics-announces-third-quarter-2022-financial.
361. Alves S, Churlaud G, Audrain M, et al. Interleukin-2 improves amyloid pathology, synaptic failure and memory in alzheimer’s disease mice. Brain. 2017;140(3):826–842. doi:10.1093/brain/aww330
362. Flores RR, Zhou L, Robbins PD. Expression of IL-2 in β cells by AAV8 gene transfer in pre-diabetic NOD mice prevents diabetes through activation of FoxP3-positive regulatory T cells. Gene Ther. 2014;21(8):715–722. doi:10.1038/gt.2014.45
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.