Back to Journals » Journal of Inflammation Research » Volume 19
Artemin Serves as a Novel Predictive Biomarker and Exerts a Protective Effect Against Preterm Birth
Authors Zhang J, Mei S, Li Y, Yu H, Yi X, Jin J, Zhang L, Wang Y, Mai Z, Liu G
Received 19 March 2026
Accepted for publication 21 June 2026
Published 10 July 2026 Volume 2026:19 608346
DOI https://doi.org/10.2147/JIR.S608346
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Adrian Lee
Jie Zhang,1,* Shiwei Mei,2,* Yingying Li,3 Huiwen Yu,3 Xiaochun Yi,4 Jing Jin,3 Li Zhang,3 Yijia Wang,5 Zixia Mai,3 Guocheng Liu3
1Department of Rehabilitation, Women and Children’s Hospital, Southern University of Science and Technology, Guangzhou, People’s Republic of China; 2Department of Radiation, Women and Children’s Hospital, Southern University of Science and Technology, Guangzhou, People’s Republic of China; 3Department of Obstetrics, Women and Children’s Hospital, Southern University of Science and Technology, Guangzhou, People’s Republic of China; 4Department of Urology, Yueyang People’s Hospital, Hunan Normal University, Yueyang, People’s Republic of China; 5Department of Neonatology, Guangzhou Women and Children’s Medical Center, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zixia Mai, Email [email protected] Guocheng Liu, Email [email protected]
Background: Preterm birth (PTB) is a major cause of neonatal morbidity and mortality and is closely associated with dysregulated inflammatory responses at the maternal–fetal interface. Excessive activation of inflammatory signaling pathways often represents a terminal event in PTB, highlighting the need to identify upstream immunoregulatory factors involved in disease pathogenesis. This study investigated Artemin (ARTN), a member of the glial cell line-derived neurotrophic factor family, as a potential regulator of inflammation-associated PTB.
Methods: A total of 56 pregnant women, including cases of infectious preterm birth (IPB), non-infectious preterm birth (NIPB), and term birth (TB), were enrolled. Maternal plasma samples were analyzed using OLINK proteomics, and placental tissues were examined by immunohistochemistry and Western blotting. Differentially expressed inflammatory proteins were further evaluated using Gene Ontology and Kyoto Encyclopedia of Genes and Genomes pathway analyses. The diagnostic performance of ARTN was assessed using receiver operating characteristic curve analysis. In addition, an LPS-induced mouse model of inflammation-driven PTB was used to evaluate the potential protective effect of exogenous ARTN supplementation.
Results: ARTN expression was significantly reduced in maternal circulation and placental tissues in IPB and showed a strong positive correlation with gestational age. Enrichment analyses indicated that ARTN-associated inflammatory proteins were involved in cytokine–cytokine receptor interactions and JAK–STAT signaling. Receiver operating characteristic analysis demonstrated that ARTN exhibited strong discriminatory power for IPB, with an area under the curve of 0.864, and the predictive performance was further enhanced when ARTN was combined with IL-6. In the LPS-induced mouse model, ARTN supplementation markedly reduced preterm delivery and fetal mortality.
Conclusion: These findings suggest that ARTN may be involved in inflammation-associated PTB and may have potential value as a candidate predictive biomarker. However, because the human plasma samples were collected across a broad gestational age range and largely in a peripartum clinical context, the present data do not establish ultra-early predictive utility. Given the relatively small sample size and lack of independent validation, these results should be interpreted as preliminary and require confirmation in larger prospective cohorts.
Keywords: preterm birth, artemin, biomarker, immune dysregulation, inflammatory response, therapeutic target
Introduction
Preterm birth, defined as delivery before 37 completed weeks of gestation, remains a leading cause of neonatal mortality and long-term neurodevelopmental impairment worldwide. Recent global estimates indicate that approximately 13.4 million newborns were born preterm in 2020, accounting for 9.9% of all live births worldwide.1 Neonatal morbidities associated with preterm birth include respiratory complications such as respiratory distress syndrome, which can progress to bronchopulmonary dysplasia, neurological issues including intraventricular hemorrhage and periventricular leukomalacia, retinopathy of prematurity, early and late-onset sepsis, necrotizing enterocolitis, and others.2 Despite advances in perinatal and neonatal care, the global preterm birth rate showed no measurable reduction between 2010 and 2020, highlighting the need for a deeper understanding of its underlying pathophysiological mechanisms. Preterm birth is not a single disease but is typically defined as a “syndrome” with significant etiological heterogeneity, involving multiple risk factors such as infection, uteroplacental dysfunction, maternal stress, and cervical insufficiency.3,4 However, these diverse etiologies ultimately converge on several common terminal pathways that trigger the premature onset of labor.5,6
Among these pathways, immune system dysregulation has been identified as a core effector mechanism.7,8 Successful pregnancy depends on the establishment of a unique immune tolerance microenvironment at the maternal-fetal interface, a process akin to a finely regulated “semi-allogeneic transplantation”.6,9,10 Disruption of this dynamic balance, particularly when it shifts toward a pro-inflammatory phenotype, can initiate pathological processes leading to preterm birth.11 Infection is one of the most well-established risk factors for preterm birth.12 Pathogen-associated molecular patterns (eg, lipopolysaccharide, LPS) can activate the innate immune system via Toll-like receptors (TLRs), triggering the massive release of pro-inflammatory cytokines such as tumor necrosis factor α (TNF-α), interleukin 1β (IL-1β), and interleukin 6 (IL-6).13,14 These cytokines subsequently upregulate the expression of prostaglandins and matrix metalloproteinases by activating key signaling pathways, including nuclear factor κB (NF-κB) and Janus kinase-signal transducer and activator of transcription (JAK-STAT), which collectively drive the pathological processes of uterine contraction, cervical ripening, and fetal membrane rupture.15–18 Notably, in non-infectious preterm birth, “sterile inflammation” triggered by endogenous danger signals such as tissue damage and stress also plays a critical role.19 Clinical studies have confirmed that pro-inflammatory cytokine levels are generally elevated in cervicovaginal secretions, amniotic fluid, and peripheral blood of pregnant women with preterm birth, while anti-inflammatory cytokines such as IL-10 are relatively insufficient or show compensatory increases.20,21 This suggests that an excessive inflammatory response is a common pathological mechanism underlying preterm birth across various etiologies.3
Currently, the clinical assessment of preterm birth risk primarily relies on prior obstetric history and transvaginal ultrasound measurement of cervical length.22 In the field of biomarkers, fetal fibronectin (fFN) and phosphorylated insulin-like growth factor binding protein-1 (phIGFBP-1) are commonly used for auxiliary diagnosis.23 However, the sensitivity and specificity of these methods remain suboptimal, and their core utility is limited to short-term prediction (eg, the risk of preterm birth within the next 7–14 days), offering minimal benefit for true early warning and preventive intervention.23 More importantly, many extensively studied biomarkers, such as IL-6, largely reflect downstream inflammatory activation.3 Their levels may increase after the inflammatory cascade has already been initiated, which limits their utility for early risk stratification and preventive intervention.3,24 Therefore, identifying upstream factors that reflect disruption of immune homeostasis before overt inflammatory amplification may provide additional value for understanding inflammation-associated preterm birth and for improving risk assessment in appropriate clinical contexts. Such biomarkers should focus on reflecting the imbalance in immune homeostasis rather than merely indicating inflammatory response output.
Artemin (ARTN), also known as Enovin or Neublastin, is a member of the glial cell line-derived neurotrophic factor (GDNF) family.25 ARTN signals mainly through GDNF family receptor α3 and RET receptor complexes and has traditionally been studied for its roles in neuronal survival, development, and tissue repair.26–28 More recent evidence indicates that ARTN also has non-neuronal biological functions, including anti-inflammatory and tissue-protective effects in experimental inflammatory injury models.29–32 These findings suggest that ARTN may participate in the regulation of inflammatory homeostasis rather than functioning solely as a neurotrophic factor. However, its role in pregnancy-related immune regulation and inflammation-associated preterm birth remains poorly understood.
However, a growing body of evidence demonstrates that ARTN plays pivotal roles in non-neuronal tissues. In the reproductive system, studies have confirmed that ARTN is expressed in preimplantation embryos and early gestational reproductive tracts in both mice and humans.33 As an estrogen-responsive gene, ARTN expression is markedly upregulated during pregnancy.34 In vitro experiments have shown that ARTN promotes embryonic development from the zygote to the blastocyst stage in a dose-dependent manner, enhancing cell proliferation and reducing apoptosis. Notably, these regulatory effects are specifically blocked by ARTN-specific neutralizing antibodies.35 These findings suggest that ARTN may play a critical role in maintaining pregnancy. However, the systemic regulatory functions of ARTN in term pregnancy maintenance and preterm birth pathogenesis remain largely unknown. Based on the aforementioned research background, this study aims to determine whether ARTN can serve as a novel preterm birth predictive biomarker, overcoming the limitations of traditional inflammatory markers, and to explore whether ARTN-targeted interventions could provide a new therapeutic strategy for preterm birth prevention.
Materials and Methods
Participants
The study was conducted in accordance with the ethical principles outlined in the Declaration of Helsinki, and the protocol for human sample collection was approved by the Ethics Committee of Women and Children’s Hospital, Southern University of Science and Technology, Guangzhou, China (20211022). Informed consent was obtained from all participants prior to the collection of maternal plasma samples. All participants were assured of their confidentiality and the voluntary nature of their participation. Personal identifiers were removed from the samples to maintain anonymity. The collected samples were used exclusively for the purposes of this study, and participants were free to withdraw at any time without consequence.
We collected maternal plasma samples from 56 pregnant women at Guangdong Women and Children Hospital, with gestational ages ranging from 24 to 41 weeks. The cohort included 23 women with non-infectious preterm birth (NIPB), 20 women with clinically suspected infection-associated preterm birth (IPB), and 13 women with term births (TB) as controls. Preterm birth was defined as delivery before 37 completed weeks of gestation according to WHO criteria. In this study, IPB was defined clinically as preterm birth accompanied by maternal signs suggestive of infection or systemic inflammatory response within 24–72 hours before delivery, including fever (temperature > 37.5°C) and an elevated white blood cell count above the normal reference range (4.3–11.3 × 10^9/L). NIPB was defined as preterm birth occurring before 37 completed weeks of gestation without these clinical signs during the same pre-delivery period. TB controls were defined as deliveries occurring at or after 37 completed weeks of gestation without clinical evidence of infection. Because microbiological confirmation was not available for all participants, the IPB group should be interpreted as clinically suspected infection-associated preterm birth rather than microbiologically confirmed infection or sepsis. Exclusion criteria included abortion, stillbirth, and other severe acute illnesses.
Given the exploratory nature of this biomarker discovery study and the limited availability of well-characterized maternal plasma and matched placental samples, no formal a priori power calculation was performed. The sample size was determined based on the availability of eligible participants during the study period. Therefore, the findings from the human cohort were considered preliminary and were further supported by placental validation experiments and an LPS-induced mouse model.
Sample Collection
Approximately 2 mL of peripheral blood was collected from each patient into EDTA tubes. Plasma was then isolated by centrifugation at 3000 rpm for 15 minutes at 4°C and stored at −80°C until further analysis. Placental tissue was collected following delivery. One part of the tissue was frozen at −80°C for protein extraction, while the other part was fixed in 10% phosphate-buffered formalin, embedded in paraffin, and sectioned at 4 μm thickness for subsequent examination.
Inflammation-Related Biomarkers Analysis and Bioinformatics Analysis
Plasma samples were analyzed using the OLINK 92-panel (OLINK Proteomics AB, Uppsala, Sweden) according to the manufacturer’s instructions to ensure high sensitivity and specificity. First, the sample was added, and oligonucleotides bound to antibody pairs linked to complementary oligonucleotides, forming a pairwise binding of matching probes. These probe pairs were then amplified using universal primers and digested by uracil-DNA glycosylase. Specific primers and quantitative real-time PCR were subsequently used to amplify and quantify the oligonucleotides in the sample. For bioinformatics analysis, we employed the clusterProfiler package in R to conduct Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses. Differentially expressed proteins were mapped to relevant terms or pathways in the GO and KEGG databases to identify significantly enriched GO terms and KEGG pathways. The ggplot2 R package was used to visualize the results of the GO and KEGG enrichment analyses, with the top 20 GO terms and KEGG pathways displayed as bubble charts.15
Mice and Preterm Induction
The animal experimental protocols were approved by the Southern University of Science and Technology Animal Care and Use Committee (G2024111) and were conducted in accordance with the guidelines of the National Institutes of Health on the care and ethical treatment of animals. Experiments were conducted using female C57BL/6 mice obtained from the Institute of Experimental Animals at Guangzhou Medical University. The mice were housed in an approved animal facility with ad libitum access to food and water. Pregnancy was confirmed by visual inspection of the vaginal plug, which was designated as day 0 of pregnancy. Controlled conditions of humidity (50–60%) and temperature (21 ± 2°C) were maintained, and the animals were exposed to a 12-hour light/dark cycle. Virgin female mice were mated with fertile males of the same strain. The average gestational length for C57BL/6 mice is typically 18–20 days. Animals were euthanized in a CO2 chamber, and all efforts were made to minimize suffering.
Preterm birth in mice was induced by administering lipopolysaccharide (LPS, Escherichia coli 0111; Biochemist) dissolved in saline. On day 15 of pregnancy (gestational day 15, GD15), pregnant mice were assigned to one of three groups: (1) blank control group, which received two intraperitoneal injections of 0.2 mL saline at 3-hour intervals on GD15; (2) LPS group, which received two intraperitoneal injections of 0.2 mL saline at 3-hour intervals, followed by two injections of 50 μg/kg LPS, with the first LPS injection administered 1 hour after the first saline injection and the second LPS injection administered 3 hours later; and (3) LPS + ARTN group, which received two intraperitoneal injections of recombinant ARTN at 3-hour intervals on GD15, followed by two injections of 50 μg/kg LPS using the same schedule as the LPS group. The ARTN dose was selected as an exploratory intervention dose based on preliminary feasibility considerations and prior evidence supporting the biological activity of recombinant ARTN in experimental inflammatory injury models. No dose–response experiment was performed in the present study. In addition, an ARTN-only group without LPS stimulation was not included because the primary objective of this experiment was to determine whether ARTN supplementation could attenuate LPS-induced inflammation-associated preterm birth. These design features were considered when interpreting the animal findings.
All experimental protocols were approved by the Guangzhou Medical University Animal Care and Use Committee and were conducted following the guidelines of the National Institutes of Health for the care and ethical treatment of animals.
Immunohistochemistry
Immunohistochemical staining was performed on paraffin-embedded sections of target tissues. Briefly, human or mouse placental tissues were dewaxed and subjected to antigen retrieval. The slides were baked at 60°C for over 1 hour, followed by dewaxing using a dewaxing solution and a gradient of alcohol. The slides were then immersed in 1× sodium citrate antigen retrieval solution, and steam was released from the pressure cooker’s vent hole. Afterward, the slides were treated with 3% H2O2 and incubated at room temperature for 10 minutes, followed by rinsing with PBS. The slides were permeabilized for 30 minutes in PBS containing 0.1% Triton X-100 and 1% BSA at room temperature, then incubated overnight at 4°C with primary antibodies specific to ARTN (Abmart, Shanghai, China), IL-6 (Boster, Wuhan, China), and IL-10 (Boster, Wuhan, China). After washing, the slides were incubated with secondary antibodies (Cambridge, MA, USA) for 45 minutes at 37°C. DAB solution was then applied for DAB staining, followed by hematoxylin staining of the nuclei. The sections were mounted and observed under a light microscope. Quantification was performed using ImageJ.
Western Blot
Placental tissue from human or mouse samples was collected, and proteins were extracted using a lysis buffer containing phosphatase and protease inhibitors. Protein concentration was determined using a BCA assay kit. The protein samples were diluted to 3 ng/μL with RIPA buffer, and the final concentration was adjusted to 2.4 ng/μL by adding 5× loading buffer. Equal amounts of protein were loaded onto SDS-PAGE gels, subjected to electrophoresis, and transferred to PVDF membranes. After blocking with skim milk for 2 hours, the membranes were incubated overnight at 4°C with primary antibodies specific to ARTN (Abmart, Shanghai, China). After washing, the membranes were incubated with appropriate secondary antibodies for 2 hours at room temperature. Protein bands were detected using chemiluminescent reagents and visualized by image density analysis (Bio-Rad). Data were analyzed using ImageJ. For figure preparation, Western blot images were cropped from the original uncropped membrane images for display. Any brightness or contrast adjustment was applied linearly and uniformly to the entire image, without selective modification of individual lanes or bands. Uncropped raw blot images were provided as supplementary source data.
ELISA Validation
Plasma samples from 56 pregnant women, including 20 with infectious preterm birth (IPB), 23 with non-infectious preterm birth (NIPB), and 13 with term births, were analyzed using ELISA kits for ARTN (Ruixinbio, Quanzhou, China) and IL-6. Assays were performed according to the manufacturer’s protocol. Regression analysis was applied to generate standard curves from the ELISA data, and cytokine concentrations in the samples were determined by correlating absorbance readings with the corresponding standard curves.
Statistical Analysis
Normally distributed data are presented as mean ± standard deviation, whereas non-normally distributed data are presented as median with interquartile range. Categorical variables are presented as numbers and percentages. Demographic and clinical characteristics were compared using the Chi-square test, Fisher’s exact test, Mann–Whitney U-test, or Kruskal–Wallis test, as appropriate. Logistic regression analyses were performed to explore the association between inflammatory protein levels and the risk of IPB compared with the TB or NIPB groups. To reduce potential confounding, maternal age at delivery, prepregnancy body mass index, parity, and pregnancy type were considered as covariates in sensitivity analyses where model convergence allowed. Diagnostic performance was evaluated by receiver operating characteristic (ROC) curve analysis, and the area under the curve (AUC) was calculated. The optimal cutoff value was determined using the Youden index. Because of the limited sample size and small subgroup numbers, ROC analyses and gestational age-stratified analyses were interpreted as exploratory, and no definitive diagnostic cutoff was proposed. Statistical analyses were conducted using R statistical software, GraphPad Prism 5.0, and SPSS 23.0. A p-value of < 0.05 was considered statistically significant.
Results
Characteristics of the Participants
According to the predefined clinical criteria for gestational age and infection-related signs, stratification of the 56 pregnancies revealed the following distributions: 13 cases (23%) were term births (TB), 23 cases (41%) were non-infectious preterm births (NIPB), and 20 cases (35%) were clinically suspected infection-associated preterm births (IPB). Compared to the TB group, both preterm groups had significantly lower birth weights (p < 0.0001), as well as lower one-minute (p < 0.01) and five-minute (p < 0.05) Apgar scores. No significant differences were observed in other health outcomes, including maternal age at delivery, prepregnancy BMI, antenatal complications, or other factors (Table 1).
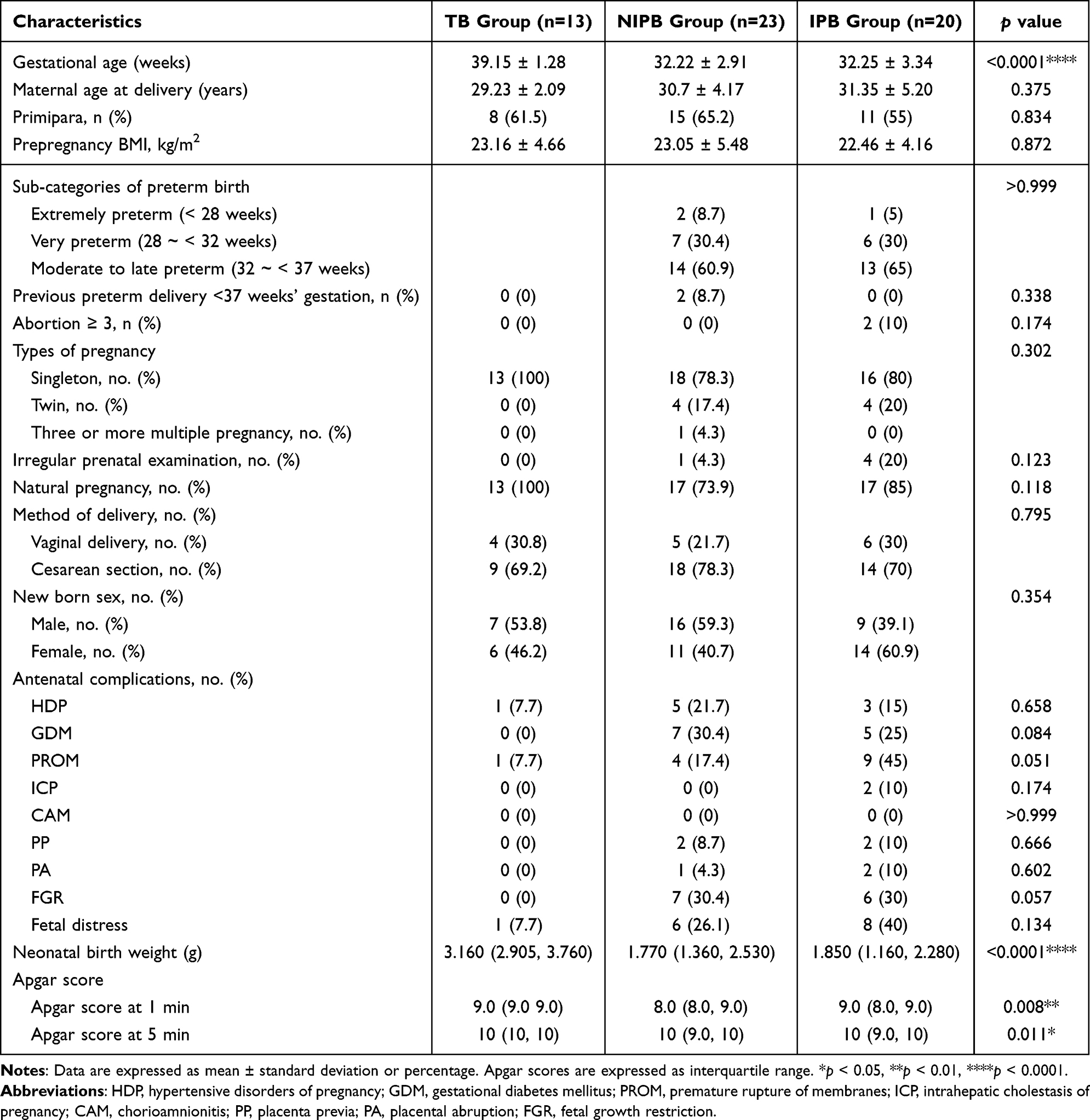 |
Table 1 Population Characteristics |
Differentially Expressed Proteins Identified by Olink Analysis Among IPB, NIPB, and TB
Plasma samples were collected from 56 pregnant women between 24 and 41 weeks of gestation and categorized into three groups: TB, IPB, and NIPB. Olink proteomic analysis identified all 92 plasma proteins across the groups. Thirteen inflammation-associated proteins were differentially expressed between the TB and NIPB groups, while sixteen proteins showed differential expression between the NIPB and IPB groups. Of these, seven proteins exhibited significant differences (Figure 1).
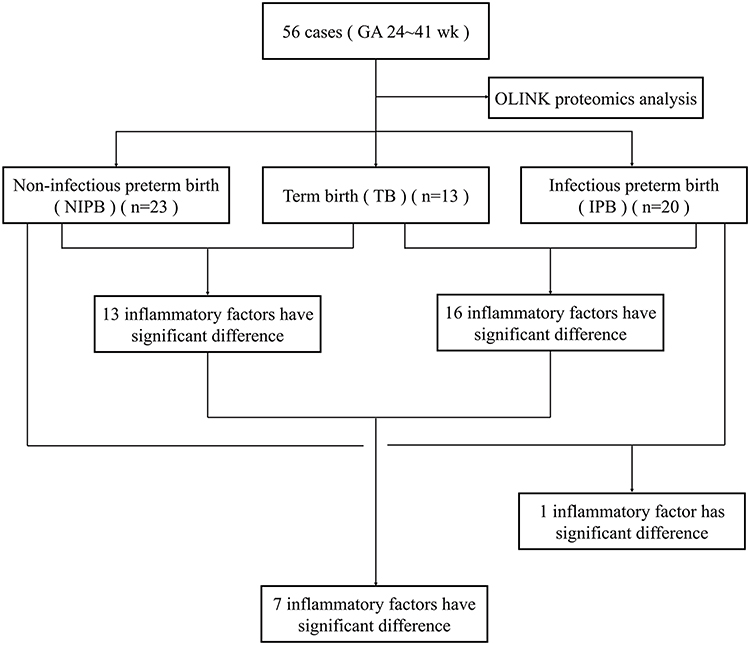 |
Figure 1 The flow chart of the study cohort. |
Between the TB and NIPB groups, IL-6, MMP-1, CCL28, and ARTN exhibited the most significant differences (p < 0.01). Most inflammatory factors, particularly IL-6, showed an upregulation trend in the NIPB group, consistent with previous studies (Figure 2A and B). The expression levels of IL-6, ARTN, IL-10, IL-24, CX3CL1, and CCL28 were significantly different between the TB and IPB groups (p < 0.01). The difference in IL-6 and ARTN expression was the most significant (p < 0.001). Similar to the NIPB group, IL-6 was significantly upregulated in the IPB group, whereas ARTN was highly expressed in the TB group and significantly decreased in the IPB group (Figure 2C and D). Furthermore, IL-6 and IL-10 exhibited higher degree scores in the protein-protein interaction (PPI) network, highlighting the important role of inflammation in preterm birth (Figure S1A–C).
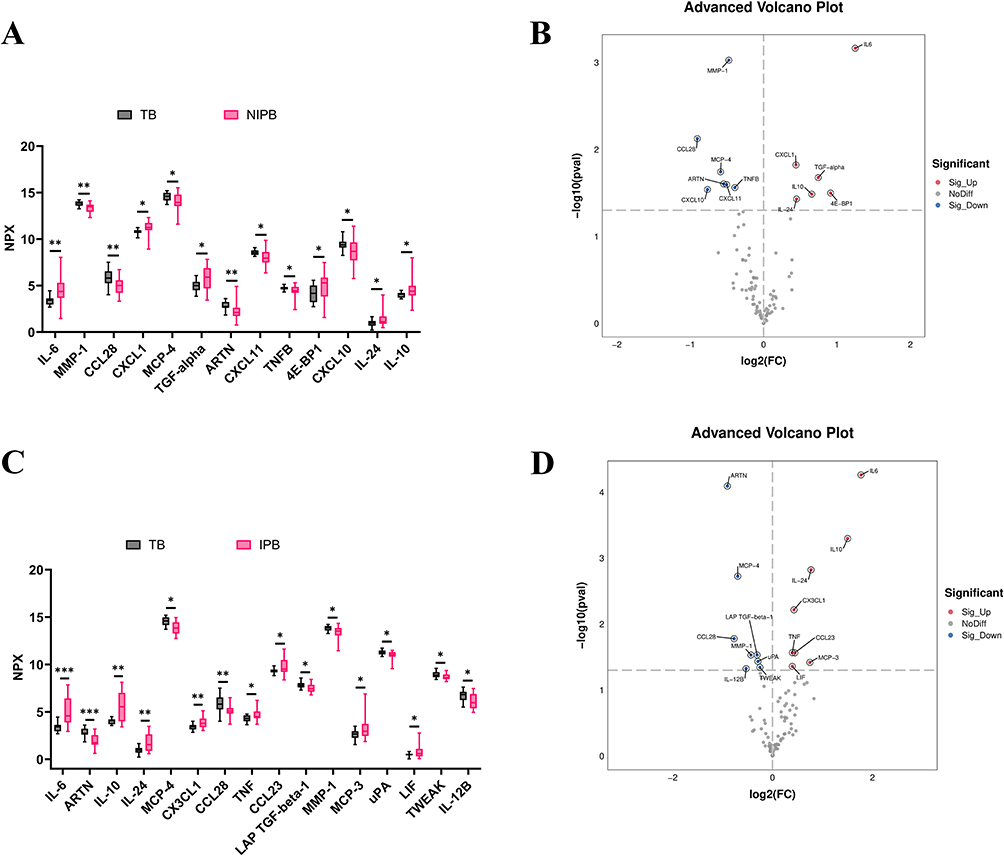 |
Figure 2 All differentially expressed inflammation-related biomarkers between the TB and NIPB groups, TB and IPB groups, and NIPB and IPB groups. (A) Box plot of the 13 inflammation-related biomarkers differentially expressed between the TB and NIPB groups. NPX, normalized protein expression, is OLINK’s arbitrary unit in Log2 scale. It is calculated from Ct values, and data preprocessing and normalization were performed to minimize both intra- and inter-assay variation. (B) Volcano plot showing the 13 inflammation-related biomarkers differentially expressed between the TB and NIPB groups. (C) Box plot of the 16 inflammation-related biomarkers differentially expressed between the TB and IPB groups. (D) Volcano plot showing the 16 inflammation-related biomarkers differentially expressed between the TB and IPB groups. Statistical significance is indicated as follows: *p < 0.05, **p < 0.01, and ***p < 0.001. |
To further explore the role of differentially expressed proteins in preterm birth, we conducted Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses. The results revealed significant enrichment in several GO terms, including positive regulation of receptor signaling via JAK, inflammatory response, and cellular response to lipopolysaccharide (LPS). Additionally, these proteins were involved in key pathways such as cytokine-cytokine receptor interactions and the JAK-STAT pathway. The cytokine-cytokine receptor pathway, commonly associated with both spontaneous abortion and preterm birth, typically exhibits increased gene expression in preterm birth.16,17 IL-6 is known to activate the JAK-STAT3 pathway, potentially inducing apoptosis in placental trophoblasts during bacterial infections or sterile inflammation. This response can be mitigated by the suppressor of cytokine signaling 3 (SOCS3) (Figure 3A and B).18,19 GO and KEGG analyses showed similar results for IPB and NIPB compared to TB, with the cytokine-cytokine receptor pathway showing the most significant differences between TB and IPB (Figure S1D-I).
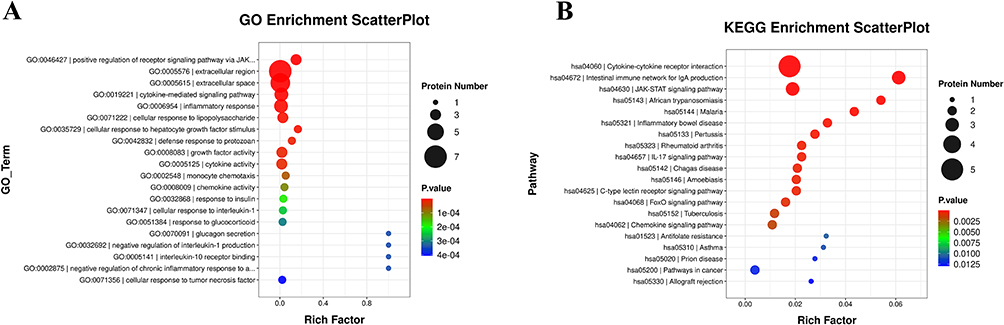 |
Figure 3 Gene Ontology (GO) (A and B). Kyoto Encyclopedia of Genes and Genomes (KEGG) (B) enrichment analysis of the differentially expressed inflammation-related proteins among the 3 groups. |
Decreased ARTN Was Associated with Preterm Birth and Showed Candidate Predictive Value
To identify biomarkers with strong predictive value for preterm birth, we analyzed the expression levels of key proteins across the groups. A Venn analysis revealed seven proteins that were differentially expressed in the comparisons between TB and both preterm groups (IPB and NIPB), indicating substantial differences between term and preterm births (Figure 4A). Among these seven proteins, IL-6, IL-10, and IL-24 were significantly elevated, whereas CCL28, ARTN, MMP-1, and MCP-4 were markedly reduced in the IPB group. Notably, IL-6 and ARTN exhibited the most significant differences between TB and IPB (p < 0.0001) (Figure 4B and Table 2).
 |
Table 2 Comparison of Differentially Expressed Proteins in the TB vs NIPB and TB vs IPB Groups |
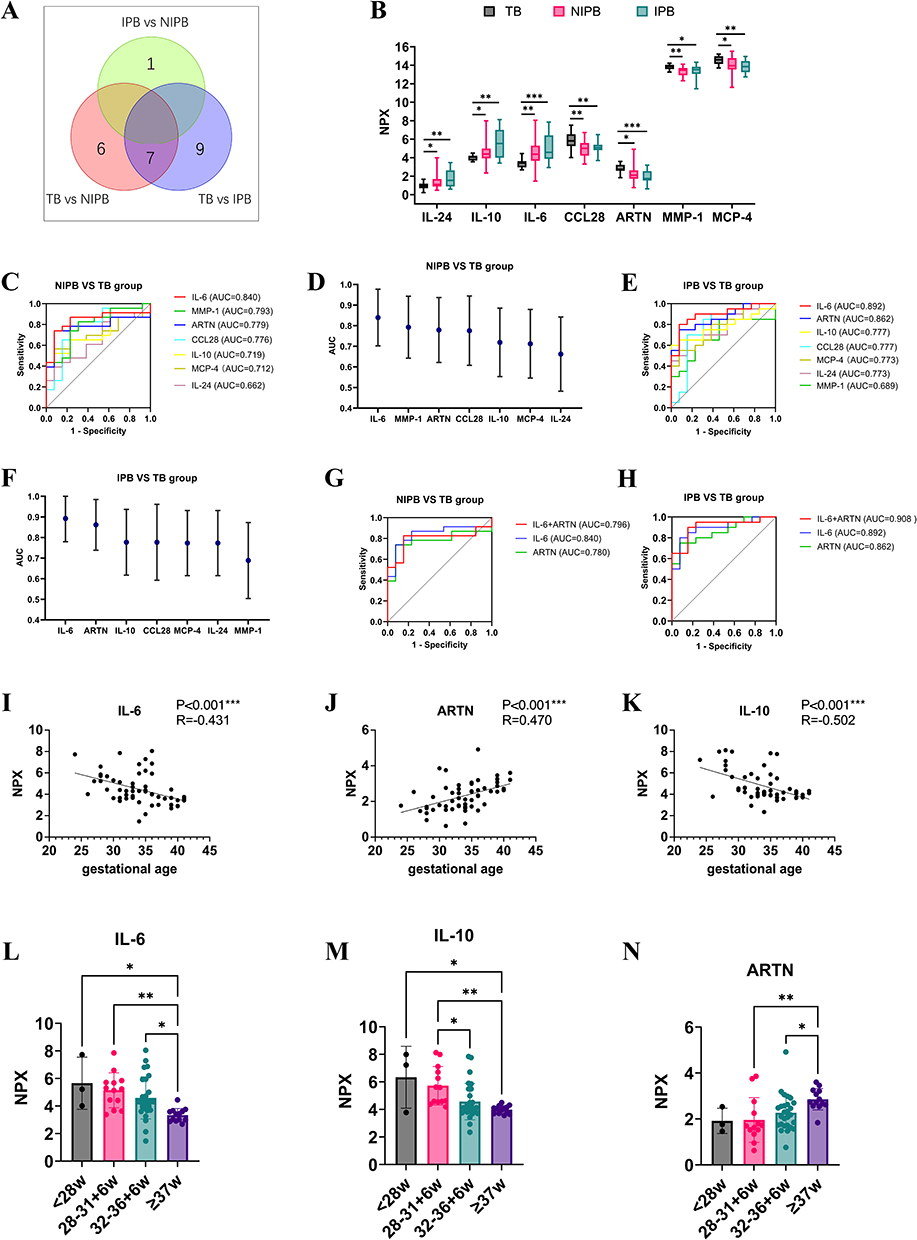 |
Figure 4 The characteristics and ROC curve of co-differential proteins detected by OLINK in TB, NIPB and IPB groups. (A) Wayne diagram showing the common and unique proteins in TB, NIPB and IPB groups. (B) Box plot of the 7 co-differential protein expressions among TB, NIPB and IPB groups (*p < 0.05, **p< 0.01, ***p< 0.001). (C and D) ROC curve analysis and characteristics of AUC of the 7 co-differential proteins in TB and NIPB group. AUC: Area under the curve. (E and F) ROC curve analysis and characteristics of AUC of the 7 co-differential proteins in TB and IPB group. (G) ROC curve analysis of the combination of IL-6 and ARTN in TB and NIPB group. (H) ROC curve analysis of the combination of IL-6 and ARTN in TB and IPB group. (I–K) Correlation analysis between IL-6, IL-10, ARTN and their gestational age at birth in 56 cases. (L–N) The expression levels of IL-6, IL-10, ARTN correspond to gestational age. |
To assess the predictive potential of these biomarkers for preterm birth, we constructed receiver operating characteristic (ROC) curves. In NIPB maternal plasma, IL-6 showed the strongest performance, with an AUC of 0.840 (95% CI: 0.702–0.977, p < 0.001). CXCL1 (AUC = 0.773), MMP-1 (AUC = 0.794), ARTN (AUC = 0.776), and CCL28 (AUC = 0.776) also showed moderate predictive value, though none achieved optimal accuracy (Figure 4C and D, Table 3). In contrast, in IPB maternal plasma, IL-6 and ARTN showed promising discriminatory performance. IL-6 had an AUC of 0.894 (95% CI: 0.741–0.986, p < 0.001), and ARTN achieved an AUC of 0.864 (95% CI: 0.741–0.986, p < 0.001), ranking second only to IL-6 (Figure 4E and F, Table 4). Because of the relatively small subgroup size, these ROC findings should be regarded as exploratory and require validation in larger independent cohorts.
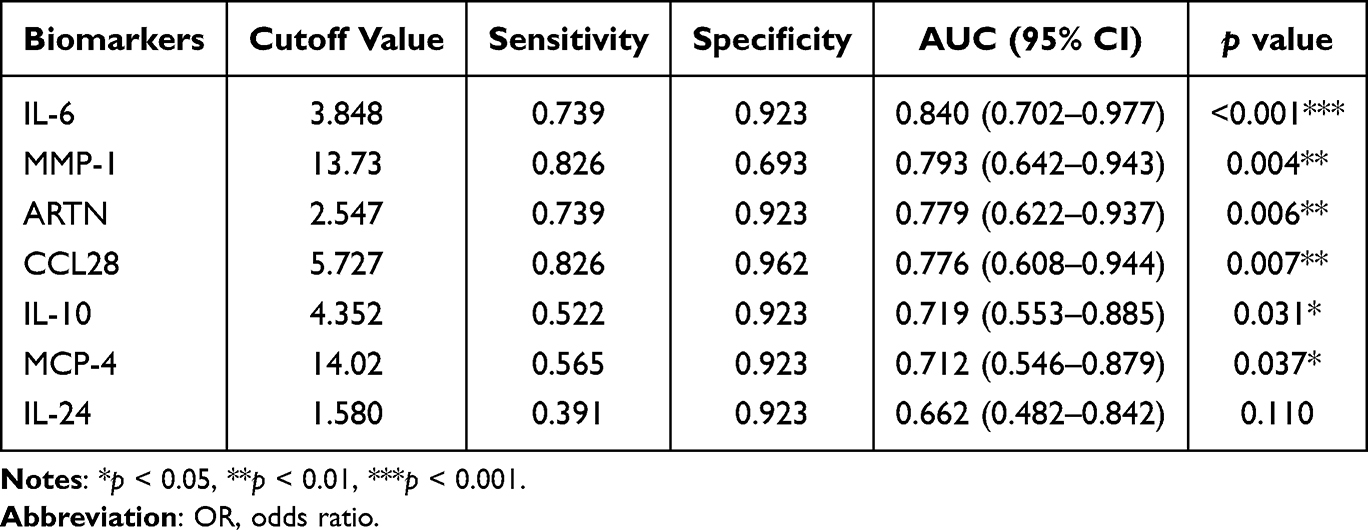 |
Table 3 Diagnostic Efficacy of Co-Differential Proteins in Distinguishing NIPB from TB |
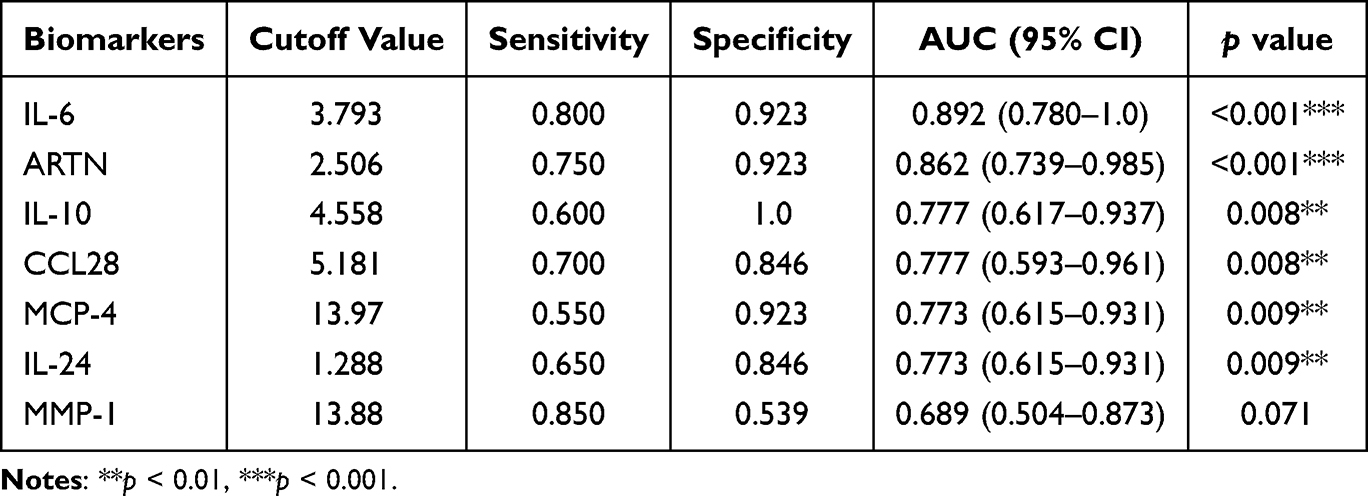 |
Table 4 Diagnostic Efficacy of Co-Differential Proteins in Distinguishing IPB from TB |
ARTN, a candidate immunomodulatory factor of peripheral nerve development and a factor implicated in tumor progression, has not been extensively examined in the context of preterm birth.12,13 IL-6, a central inflammatory mediator, contributes to preterm labor by promoting acute-phase responses, hematopoiesis, and immune activation,20,21 and influences pregnancy outcomes by regulating immune and placental cell activity.22 Our findings indicate that both IL-6 and ARTN showed promising discriminatory value for preterm birth, particularly in clinically suspected infection-associated preterm birth. Although IL-6 has been widely studied, ARTN’s function remains largely underexplored. We therefore evaluated the combined discriminatory performance of IL-6 and ARTN. While the combination did not improve discrimination in NIPB, it enhanced performance in the IPB group, surpassing either biomarker alone (Figure 4G and H, Table 5 and Table 6). These human data indicate an association between reduced ARTN levels and preterm birth rather than a causal relationship. Because the plasma samples were collected between 24 and 41 weeks of gestation and largely in a peripartum clinical context, these findings should not be interpreted as evidence of ultra-early prediction. ARTN should instead be considered a candidate biomarker requiring validation in larger prospective cohorts with earlier longitudinal sampling.
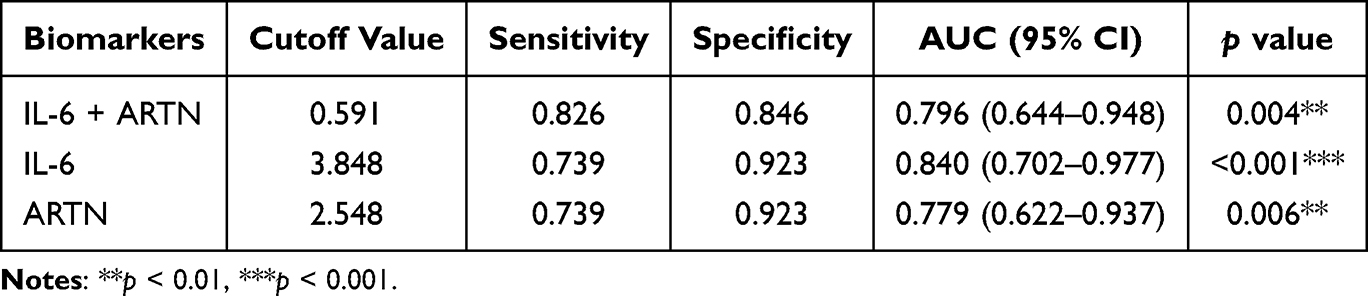 |
Table 5 ROC Curve Analysis of the Combination of IL-6+ARTN in Distinguishing NIPB from TB |
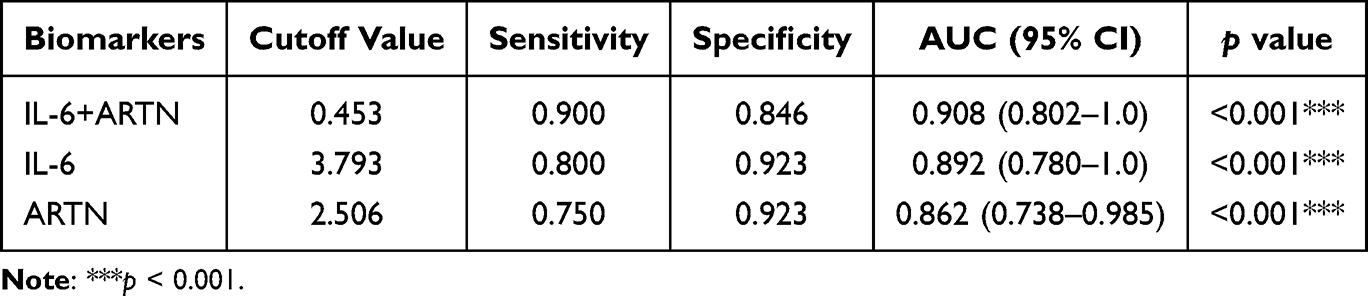 |
Table 6 ROC Curve Analysis of the Combination of IL-6+ARTN in Distinguishing IPB from TB |
We further classified infants into four categories: extremely preterm (<28 weeks), moderate preterm (28–<32 weeks), late preterm (32–<36 weeks), and term (>37 weeks). ARTN levels displayed a strong positive correlation with gestational age (Figure 4I–K). ARTN concentrations in term infants were significantly higher than those in extremely preterm infants (p < 0.001), with a similar elevation observed relative to moderate preterm infants (p < 0.05). Given ARTN’s known role in neuronal survival and peripheral nerve homeostasis, reduced ARTN levels may help explain neurosensory, motor, cognitive, and behavioral delays commonly observed in early-gestation preterm infants.23 In contrast, IL-6 levels were negatively correlated with gestational age, showing significantly higher concentrations in extremely preterm infants compared with term infants (p < 0.001), and a similar trend in late preterm infants (p < 0.01) (Figure 4L–N).24 These results further support IL-6 as both a predictor of preterm birth and a potential indicator for estimating gestational age. Other differentially expressed proteins also varied with gestational age; for instance, IL-10 levels were significantly elevated in extremely preterm infants (p < 0.05), while CCL28 levels were reduced in very preterm infants (p < 0.05) (Figure S2). Because the number of participants within some gestational age strata was limited, the gestational age-stratified analyses were intended to describe trends rather than to establish definitive gestational age-specific reference ranges or diagnostic thresholds.
ARTN Was Significantly Decreased in IPB in Both Humans and Mice
Our study identified significant differences in ARTN, IL-6, and IL-10 levels between the IPB and TB groups. While previous research has documented elevated IL-6 levels in preterm birth groups, investigations into ARTN remain limited, leaving its predictive and diagnostic value largely unexplored. Consequently, we selected these three biomarkers for further validation in clinical samples. Histochemical analysis of human placental tissue samples revealed that ARTN levels were significantly reduced (p < 0.05), while IL-6 and IL-10 levels were significantly elevated (p < 0.001, p < 0.05) in clinically suspected infection-associated preterm birth cases (Figure 5A–F). Similarly, in a mouse model, placental tissue analysis confirmed these findings: ARTN was notably decreased (p < 0.05), and IL-6 was significantly increased in preterm mice, mirroring the results observed in human samples (Figure 5G–J).
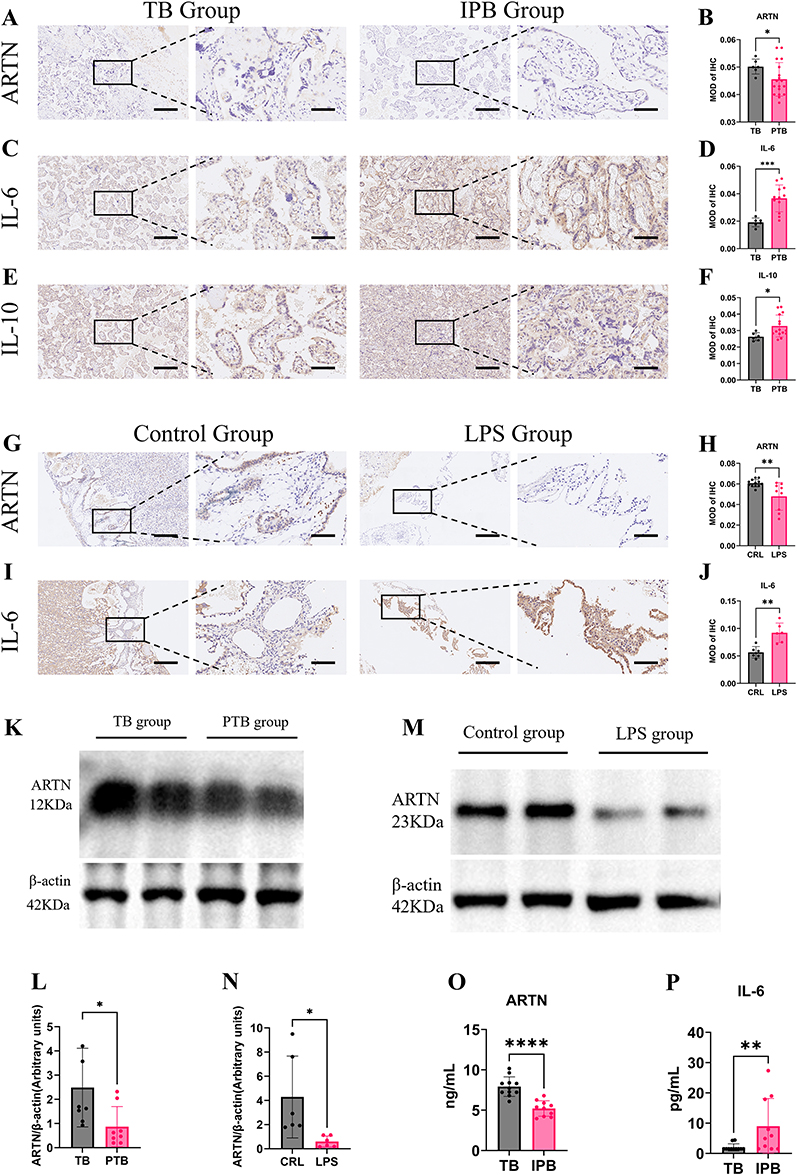 |
Figure 5 Comparison of ARTN, IL-6, and IL-10 expression in human and mouse placental tissues. (A–F) Immunohistochemical staining of ARTN, IL-6, and IL-10 in placental tissues from the TB and clinically suspected IPB groups. Placental tissues were collected after delivery, and the levels of ARTN, IL-6, and IL-10 were determined by immunohistochemistry. Original magnifications were ×100 and ×400. Scale bars represent 200 μm for ×100 images and 50 μm for ×400 images. (G–J) Immunohistochemical staining of ARTN and IL-6 in placental tissues from control and LPS-treated mice. Placental tissues were collected 12 hours after the first LPS or vehicle injection on GD15, and ARTN and IL-6 levels were determined by immunohistochemistry. Scale bars represent 200 μm for ×100 images and 50 μm for ×400 images. MOD, mean optical density. (K and L) ARTN expression levels in placental tissues from the TB and clinically suspected IPB groups, as determined by Western blotting. (M and N) ARTN expression levels in placental tissues from control and LPS-treated mice, as determined by Western blotting. (O and P) ELISA validation of maternal plasma ARTN and IL-6 levels in the TB and clinically suspected IPB groups. Data are presented as mean ± standard deviation. Statistical significance is indicated as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. |
Protein analysis of human placental tissue also revealed a significant decrease in ARTN levels in the IPB group (p < 0.05) (Figures 5K, L and S3). Previous studies have shown that maternal-derived IL-6 levels are significantly elevated in preterm placentas, activating the JAK/STAT pathway in the placental fetal compartment and initiating a cascade of inflammatory responses.25 Inhibiting IL-6 signaling can mitigate maternal and fetal responses to LPS during pregnancy, which is consistent with our findings.26 However, the role of ARTN in placental pathways remains unclear.
We also performed similar analyses in a mouse model of LPS-induced preterm birth, where ARTN levels exhibited the same trend, showing significantly decreased protein content in mouse placental tissue (p < 0.05) (Figure 5M and N). In addition to placental tissue expression analysis, we investigated differential protein expression in serum. ELISA results indicated that ARTN levels were significantly lower in the IPB group compared to the TB group (p < 0.0001), while IL-6 levels were significantly higher (p < 0.01) (Figure 5O and P). These findings are consistent with the trends observed in the Olink analysis. Because these human plasma and placental samples were collected in a peripartum cross-sectional setting, the observed ARTN reduction should be interpreted as an association with preterm birth status and gestational age, rather than evidence that ARTN downregulation causally initiates human preterm birth.
ARTN Supplementation Was Associated with Improved Pregnancy Outcomes in the LPS-Induced Preterm Birth Mouse Model
To explore whether ARTN supplementation could influence inflammation-associated preterm birth, we established an LPS-induced mouse model and administered recombinant ARTN before LPS challenge. The experimental design is shown in Figure 6A. The main difference between the LPS and LPS + ARTN groups was the administration of recombinant ARTN before LPS exposure. LPS treatment resulted in preterm delivery before GD19 in all pregnant mice (Figure 6B). In contrast, the preterm delivery rate was lower in LPS-treated mice that received ARTN supplementation. ARTN supplementation was also associated with reduced fetal mortality compared with the LPS group (Figure 6C). Pups in the LPS + ARTN group showed no significant difference in body weight compared with the blank control group (Figure 6D). These exploratory animal findings support a possible protective role of ARTN under LPS-induced inflammatory challenge. However, because no dose–response testing or ARTN-only control group was included, and because the human data were cross-sectional, the animal results should not be interpreted as definitive evidence that ARTN causally prevents human preterm birth.
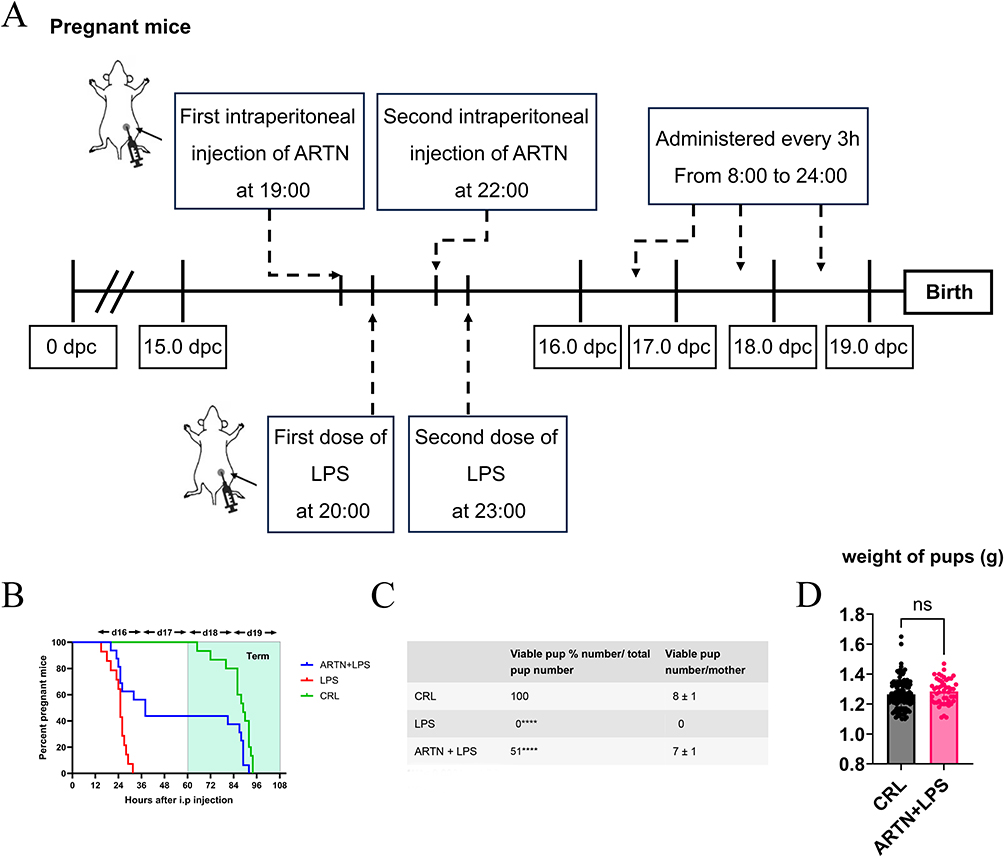 |
Figure 6 Effects of ARTN supplementation on the LPS-induced preterm birth model in mice. (A) Experimental protocol for the three groups. On gestational day 15 (GD15), pregnant C57BL/6 mice in the control, LPS, and LPS + ARTN groups received two intraperitoneal injections at 3-hour intervals. Mice in the control and LPS groups received 0.2 mL normal saline, whereas mice in the LPS + ARTN group received recombinant ARTN at 50 μg/kg. One hour after the first saline or ARTN injection, mice in the LPS and LPS + ARTN groups received two injections of LPS at 50 μg/kg, administered at a 3-hour interval. From GD16 onward, the mice were monitored every 3 hours from 8:00 AM to midnight until GD19. (B) Percentage of pregnant mice remaining undelivered following LPS administration. (C) Fetal survival rate following LPS administration. The numbers of pregnant mice were n = 15 in the control group, n = 14 in the LPS group, and n = 7 in the LPS + ARTN group. (D) Neonatal birth weight following LPS administration. The numbers of neonates were n = 116 in the control group and n = 49 in the LPS + ARTN group. Continuous data are presented as mean ± standard deviation. ****p < 0.0001. |
Discussion
In this study, we observed a consistent reduction of ARTN expression in maternal circulation and placental tissues of women with preterm birth, particularly clinically suspected infection-associated preterm birth. This ARTN reduction was associated with gestational age and showed candidate discriminatory value, especially when combined with IL-6. Because the human samples were collected in a peripartum cross-sectional setting, these findings should be interpreted as associative rather than causal. In the LPS-induced mouse model, exogenous ARTN supplementation was associated with reduced preterm delivery and improved fetal survival, providing experimental support for a possible protective role under inflammatory challenge. Collectively, these findings support ARTN as a promising candidate biomarker and potential therapeutic research direction, while further prospective and mechanistic studies are required to determine causality and define the underlying immune mechanisms.
We also identified ARTN as a candidate biomarker with promising discriminatory performance, particularly in clinically suspected infection-associated preterm birth. Conventional biomarkers, such as IL-6, primarily reflect activated inflammatory cascades and may therefore represent downstream inflammatory activity.3,36,37 In contrast, the reduction of ARTN observed in this study raises the possibility that ARTN may reflect an upstream disturbance in immune homeostasis. However, because the human samples were collected across 24–41 weeks of gestation and largely around the time of delivery, the present study cannot determine whether ARTN changes precede the onset of preterm labor or provide ultra-early prediction. Prospective longitudinal studies with samples collected earlier in pregnancy are needed to clarify the temporal relationship between ARTN decline, inflammatory activation, and subsequent preterm birth.
Our findings also suggest that ARTN may have biological relevance beyond its role as a candidate predictive biomarker. The positive correlation between ARTN expression and gestational progression indicates a possible association with pregnancy maintenance. In the LPS-induced mouse model, ARTN supplementation was associated with reduced preterm delivery and improved fetal survival, suggesting that ARTN may contribute to protection against inflammation-associated preterm birth under experimental conditions. However, the current human data do not establish causality, and the present study did not directly define the cellular targets, receptor dependence, or downstream signaling mechanisms of ARTN at the maternal–fetal interface. Therefore, ARTN-based intervention should be regarded as a potential therapeutic research direction that requires further mechanistic and preclinical validation.
The protective effect observed in the LPS-induced preterm birth model may be related to the reported immunomodulatory and anti-inflammatory properties of ARTN. Previous studies have suggested that ARTN can influence inflammatory responses in a context-dependent manner and may signal through GFRα3/RET-associated pathways in specific cell types.26,29,30 These observations provide a plausible biological basis for the effects observed in our model. Nevertheless, the present study did not directly test whether these pathways mediate the protective effect of ARTN in placental or decidual tissues. Further studies using cell-specific approaches and pathway inhibition experiments are needed to clarify the mechanism.
Based on these findings, we hypothesize that exogenous ARTN supplementation may attenuate LPS-induced inflammatory activation at the maternal–fetal interface and thereby reduce the risk of inflammation-associated preterm delivery. This hypothesis is consistent with our enrichment analysis, which implicated cytokine–cytokine receptor interactions and JAK–STAT signaling, as well as with the observed changes in inflammatory mediators such as IL-6 and IL-10. However, these data do not establish a direct causal link between ARTN and specific immune pathways. Future studies should determine whether ARTN modulates defined immune cell populations, cytokine networks, or placental signaling pathways during inflammation-associated preterm birth.
However, the evaluation of ARTN’s therapeutic potential must consider the context-dependent nature of ARTN signaling.25 The “pro-inflammatory” and “anti-inflammatory” effects of ARTN are not mutually exclusive but are jointly influenced by receptor type and target cell. When ARTN binds to the GFRα3/Ret complex on sensory neurons, it mediates pro-inflammatory effects through the ERK pathway.38 Conversely, when ARTN binds to Ret receptors on macrophages, it exerts anti-inflammatory effects via pathways including PI3K/Akt, p38/MAPK, and JNK.30
While this study provides biologically consistent evidence supporting the involvement of ARTN in inflammation-associated preterm birth, several limitations must be objectively acknowledged. First, the human samples were collected in a peripartum cross-sectional setting across a broad gestational age range of 24–41 weeks; therefore, the observed decrease in ARTN demonstrates an association with preterm birth status and gestational age but does not establish causality or ultra-early predictive utility in humans.38 Second, the clinical sample size was relatively small, particularly in subgroup and gestational age-stratified analyses, and no formal a priori power calculation was performed because this was an exploratory biomarker discovery study based on available clinical samples. Therefore, the ROC analyses may be vulnerable to overfitting, and the proposed discriminatory performance of ARTN should be interpreted as preliminary rather than definitive.39 Third, the classification of IPB was based on clinical signs suggesting infection or systemic inflammatory response, including fever and leukocytosis, rather than uniform microbiological confirmation. These criteria are nonspecific and may overlap with sterile inflammation; therefore, misclassification between infection-associated and non-infectious inflammatory preterm birth cannot be completely excluded. Fourth, although several maternal and pregnancy-related variables were considered in sensitivity analyses, the limited sample size restricted the ability to perform fully adjusted multivariable modeling without overfitting. Fifth, the animal experiment was exploratory and did not include dose–response testing or an ARTN-only control group without LPS exposure. Future studies should incorporate prospective longitudinal sampling earlier in pregnancy, larger independent cohorts, narrower gestational age strata, microbiological or histopathological confirmation of infection status, external validation cohorts, and more comprehensive preclinical designs to confirm the biomarker and therapeutic relevance of ARTN.27,40
This study provides preliminary evidence that ARTN may be associated with immune homeostasis dysregulation in inflammation-associated preterm birth and may have potential as a candidate predictive biomarker and therapeutic research direction. Future research will focus on the following aspects: First, using specific gene knockout animal models to clarify the relative contributions of ARTN in different cell types, such as trophoblasts and immune cells. Conditional knockout models, in particular, will help elucidate the function of ARTN in specific cell types. Second, in-depth investigations into the interaction mechanisms between ARTN and key signaling pathways, including the PI3K/Akt, MAPK, and JAK-STAT pathways. These studies will contribute to understanding the pathogenesis of preterm birth and may provide new potential therapeutic research direction. Third, evaluating whether ARTN-based intervention is feasible and safe in preclinical models, including systematic assessment of administration timing, dosage, delivery route, and potential side effects. Fourth, the systematic evaluation of ARTN’s potential long-term effects on offspring neurodevelopment through comprehensive behavioral, morphological, and molecular biological assessments at multiple levels and time points. Finally, conducting large-scale prospective cohort studies to validate the clinical value of the ARTN prediction model and explore its applicability in different populations and preterm birth subtypes.
Conclusions
In conclusion, this study suggests that reduced ARTN is associated with immune homeostasis dysregulation in inflammation-associated preterm birth and may have potential value as a candidate predictive biomarker. ARTN was significantly downregulated in maternal plasma and placental tissues of patients with clinically suspected infection-associated preterm birth, showed a positive correlation with gestational age, and demonstrated promising discriminatory performance, particularly when combined with IL-6. In the LPS-induced mouse model, exogenous ARTN supplementation was associated with reduced preterm delivery and fetal mortality, suggesting a possible protective role under inflammatory challenge. However, the human data were cross-sectional and largely peripartum, and therefore do not establish causality or ultra-early predictive utility. Given the relatively small sample size, broad gestational age range, and lack of independent validation, these findings should be interpreted as preliminary. Further prospective cohort studies with earlier longitudinal sampling and mechanistic experiments are required to validate the biomarker potential of ARTN and to determine whether ARTN-based intervention could represent a feasible therapeutic strategy.
ARRIVE Guidelines Statement
This study is reported in accordance with the ARRIVE guidelines. A completed ARRIVE 2.0 checklist has been uploaded as supplementary material.
Data Sharing Statement
The data presented in this study are available on request from the corresponding author.
Ethics Statement
All the animals’ experimental protocols were approved by Southern University of Science and Technology Animal Care and Use Committee and were performed following the guidelines of the National Institutes of Health on the care and ethical treatment of animals (G2024111). The protocol for human samples was approved by the ethics committee of Women and Children’s Hospital, Southern University of Science and Technology, Guangzhou, China (20211022).
Author Contributions
Jie Zhang: Conceptualization, Methodology, Formal analysis, Supervision, Writing – original draft. Shiwei Mei: Investigation, Resources, Validation, Writing – original draft. Yingying Li: Data curation, Methodology, Writing – original draft. Huiwen Yu: Methodology, Validation, Writing – original draft. Xiaochun Yi: Data curation, Writing – original draft. Jing Jin: Data curation, Writing – original draft. Li Zhang: Project administration, Resources, Writing – original draft. Yijia Wang: Data curation, Methodology, Writing – original draft. Zixia Mai: Data curation, Formal analysis, Methodology, Software, Writing – original draft. Guocheng Liu: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
We acknowledge the support from the National Natural Science Foundation of China (Grant No. 82101823) and the Natural Science Foundation of Hunan Province of China (NO.2022JJ40459).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Ohuma EO, Moller AB, Bradley E, et al. National, regional, and global estimates of preterm birth in 2020, with trends from 2010: a systematic analysis. Lancet. 2023;402:1261–18. doi:10.1016/S0140-6736(23)00878-4
2. Stoll BJ, Hansen NI, Bell EF, Higgins RD. Neonatal outcomes of extremely preterm infants from the NICHD neonatal research network. Pediatrics. 2010;126:443–456. doi:10.1542/peds.2009-2959
3. Romero R, Dey SK, Fisher SJ. Preterm labor: one syndrome, many causes. Science. 2014;345:760–765. doi:10.1126/science.1251816
4. Louis J, Muglia, Michael, Katz. The enigma of spontaneous preterm birth. New Engl J Med. 2010;362:529–535. doi:10.1056/NEJMra0904308
5. Vilotic A, Nacka-Aleksic M, Pirkovic A, et al. IL-6 and IL-8: an overview of their roles in healthy and pathological pregnancies. Int J Mol Sci. 2022;23. doi:10.3390/ijms232314574
6. Erlebacher A. Immunology of the maternal-fetal interface. Ann Rev Immunol. 2013;31:387–411. doi:10.1146/annurev-immunol-032712-100003
7. Mor G, Cardenas I. The immune system in pregnancy: a unique complexity. Am J Reprod Immunol. 2010;63:425–433. doi:10.1111/j.1600-0897.2010.00836.x
8. Sullivan G, Galdi P, Borbye-Lorenzen N, Stoye DQ, Boardman JP. Preterm birth is associated with immune dysregulation which persists in infants exposed to histologic chorioamnionitis. Front Immunol. 2021;12:722489. doi:10.3389/fimmu.2021.722489
9. Hong JY, Joo JS, Lee D. Multi-layered mechanisms of immunological tolerance at the maternal-fetal interface. Immune Network. 2024;24:e30. doi:10.4110/in.2024.24.e30
10. Li J, Wang X, Davis MM. Regulatory KIR +CD8 + T cells are elevated during human pregnancy. Sci Transl Med. 2025;17. 10.1126/scitranslmed.adm7697.
11. Cappelletti M, Della bella S, Ferrazzi E, Mavilio D, Divanovic S. Inflammation and preterm birth. J Leukoc Biol. 2016;99:67–78. doi:10.1189/jlb.3MR0615-272RR
12. Goldenberg RL, Culhane JF, Iams JD, Romero R. Epidemiology and causes of preterm birth. Lancet. 2008;371:75–84. doi:10.1016/S0140-6736(08)60074-4
13. Miller FA, Sacco A, David AL, Boyle AK. interventions for infection and inflammation-induced preterm birth: a preclinical systematic review. Reprod Sci. 2022;30:361–379. doi:10.1007/s43032-022-00934-x
14. Poltorak A, He X, Smirnova I, et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science. 1998;282:2085–2088. doi:10.1126/science.282.5396.2085
15. Sheller-Miller S, Radnaa E, Yoo JK, et al. Exosomal delivery of NF-κB inhibitor delays LPS-induced preterm birth and modulates fetal immune cell profile in mouse models. Sci Adv. 2021;4. doi:10.1126/SCIADV.ABD3865
16. Poehlmann TG, Busch S, Mussil B, et al. The possible role of the Jak/STAT pathway in lymphocytes at the fetomaternal interface. Chem Immunol Allergy. 2005;89:26. doi:10.1159/000087907
17. Qu J, Jiang H, Zhang B, et al. Oxidative stress-mediated abnormal polarization of decidual macrophages promotes the occurrence of atonic postpartum hemorrhage. Redox Biol. 2025:81. 10.1016/j.redox.2025.103530.
18. Al-Roub A, Akhter N, Al-Rashed F, Wilson A, Alzaid F. Al-Mulla F, et al.TNFα induces matrix metalloproteinase-9 expression in monocytic cells through ACSL1/JNK/ERK/NF-kB signaling pathways. Sci Rep. 2023;13:14351. doi:10.1038/s41598-023-41514-6
19. Farias-Jofre M, Romero R, Galaz J, et al. Blockade of IL-6R prevents preterm birth and adverse neonatal outcomes. EBioMedicine. 2023;98:104865. doi:10.1016/j.ebiom.2023.104865
20. Cappelletti M, Doll JR, Stankiewicz TE, et al. Maternal regulation of inflammatory cues is required for induction of preterm birth. JCI Insight. 2020:5. 10.1172/jci.insight.138812.
21. A BSL. B M L A.Endoplasmic reticulum stress regulates inflammation and insulin resistance in skeletal muscle from pregnant women. Mol Cell Endocrinol. 2016;425:11–25. doi:10.1016/j.mce.2016.02.016
22. Hessami K, D’Alberti E, Mascio DD, Berghella V. Universal cervical length screening and risk of spontaneous preterm birth: a systematic review and meta-analysis. Am J Obstetrics Gynecol. 2024;6:101343. doi:10.1016/j.ajogmf.2024.101343
23. Ville Y, Rozenberg P. Predictors of preterm birth. Best Pract Res Clin Obstetrics Gynaecol. 2018;52:23–32. doi:10.1016/j.bpobgyn.2018.05.002
24. Tency I. Inflammatory response in maternal serum during preterm labour. Facts Views Vision ObGyn. 2014;6:19.
25. Baloh RH, Tansey MG, Lampe PA, et al. Artemin, a novel member of the GDNF ligand family, supports peripheral and central neurons and signals through the GFRα3–RET receptor complex. Neuron. 1998. doi:10.1016/S0896-6273(00)80649-2
26. Zhu SP, Li YH, Bennett S, et al. The role of glial cell line-derived neurotrophic factor family member artemin in neurological disorders and cancers. Cell Prolif. 2020;53:e12860. doi:10.1111/cpr.12860
27. Ilieva M, Nielsen J, Korshunova I, et al. Artemin and an artemin-derived peptide, artefin, induce neuronal survival, and differentiation through ret and NCAM. Front Mol Neurosci. 2019. doi:10.3389/fnmol.2019.00047
28. Michael P, Jankowski K, Rau K, et al. Enhanced artemin/GFRα3 levels regulate mechanically insensitive, heat-sensitive C-fiber recruitment after axotomy and regeneration. J Neurosci. 2010;30:16272–16283. doi:10.1523/JNEUROSCI.2195-10.2010
29. Huang HM, Huang CC, Poon LYC, Chang YC. Artemin is upregulated by TrkB agonist and protects the immature retina against hypoxic-ischemic injury by suppressing neuroinflammation and astrogliosis. Front Mol Neurosci. 2021;14:645000. doi:10.3389/FNMOL.2021.645000
30. Zhou Q, Yang YL, Wang JF, et al. Inflammation-induced generation of splenic erythroblast-like Ter-Cells alleviates acute lung injury. Eur Resp J. 2024;64:3. doi:10.1165/rcmb.2024-0407OC
31. Han Y, Liu Q, Hou J, et al. Tumor-induced generation of splenic erythroblast-like ter-cells promotes tumor progression. Cell. 2018;173:634–48.e12. doi:10.1016/j.cell.2018.02.061
32. Elahi S, Rezaeifar M, Osman M, Shahbaz S. Exploring the role of galectin-9 and artemin as biomarkers in long COVID with chronic fatigue syndrome: links to inflammation and cognitive function. Front Immunol. 2024;15:1443363. doi:10.3389/fimmu.2024.1443363
33. Gómez E, Martin D, Carrocera S, et al. Expression and localization of ARTEMIN in the bovine uterus and embryos. Theriogenology. 2017:153–162. 10.1016/j.theriogenology.2016.12.007.
34. Hezam K, Jiang J, Sun F, Zhang X, Zhang J. Artemin promotes oncogenicity, metastasis and drug resistance in cancer cells. Rev Neurosci. 2017;29:93–98. doi:10.1515/revneuro-2017-0029
35. Li J, Klein C, Liang C, et al. Autocrine regulation of early embryonic development by the artemin-GFRA3 (GDNF family receptor-alpha 3) signaling system in mice. FEBS Lett. 2009;583:2479–2485. doi:10.1016/j.febslet.2009.06.050
36. Prairie E, Côté F, Tsakpinoglou M, et al. The determinant role of IL-6 in the establishment of inflammation leading to spontaneous preterm birth. Cytokine Growth Factor Rev. 2021. doi:10.1016/j.cytogfr.2020.12.004
37. Gomez-Lopez N, Stlouis D, Lehr MA, Sanchez-Rodriguez EN, Arenas-Hernandez M. Immune cells in term and preterm labor. Cell Mol Immunol. 2014;11. doi:10.1038/cmi.2014.46
38. Thornton P, Hatcher JP, Robinson I, et al. Artemin-GFRα3 interactions partially contribute to acute inflammatory hypersensitivity. Neuroscience Lett. 2013;545:23–28. doi:10.1016/j.neulet.2013.04.007
39. Chen XG, Xu XC, Chen JH, et al. ADAR1 as a placental innate immune rheostat sustaining the homeostatic balance of intrinsic interferon response at the maternal-fetal interface. Adv Sci. 2025;12:e05491. doi:10.1002/advs.202505491
40. Shang HQ, Wang Y, Mao YY, et al. Expression of artemin and GFRα3 in an animal model of migraine: possible role in the pathogenesis of this disorder. J Headache Pain. 2016;17:81. doi:10.1186/s10194-016-0673-2
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Metabolomics Reveals Molecular Signatures for Psoriasis Biomarkers and Drug Targets Discovery
Song Q, Chen Y, Ma J, Zhou W, Song J, Wu C, Liu J
Clinical, Cosmetic and Investigational Dermatology 2023, 16:3181-3191
Published Date: 4 November 2023
Machine Learning and Experimental Validation Reveal MYH11 as a Novel Prognostic Biomarker and Therapeutic Target in Bladder Cancer
Tan Z, Chen X, Fu S, Huang Y, Li H, Gong C, Lv D, Yang C, Wang J, Ding M, Wang H
Journal of Inflammation Research 2025, 18:8357-8387
Published Date: 25 June 2025