Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 16
Metabolomics Reveals Molecular Signatures for Psoriasis Biomarkers and Drug Targets Discovery
Authors Song Q, Chen Y, Ma J, Zhou W, Song J, Wu C, Liu J ![]()
Received 28 August 2023
Accepted for publication 19 October 2023
Published 4 November 2023 Volume 2023:16 Pages 3181—3191
DOI https://doi.org/10.2147/CCID.S433280
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Jeffrey Weinberg
Qian Song,1,* Ying Chen,2,3,* JianQing Ma,1 Wei Zhou,4 JunYan Song,1 ChunFu Wu,5 Jie Liu2
1Department of Medical Laboratory, North China Medical & Health Group Xingtai General Hospital, Orthopedic Hospital of Xingtai, Xingtai, People’s Republic of China; 2BGI Genomics, BGI-Shenzhen, Shenzhen, People’s Republic of China; 3College of Life Sciences, University of Chinese Academy of Sciences, Beijing, People’s Republic of China; 4China National Genebank, BGI-Shenzhen, Shenzhen, People’s Republic of China; 5Yantai Harbor Hospital, Yantai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jie Liu, BGI Genomics, BGI-Shenzhen, Shenzhen, 518083, People’s Republic of China, Email [email protected] ChunFu Wu, Yantai Harbor Hospital, Yantai, 264000, People’s Republic of China, Email [email protected]
Purpose: Psoriasis is a chronic, multi-system skin disease that can be influenced by immunological, environmental, and genetic factors. Plasma metabolomic analysis can provide a great deal of information on potential diagnostic biomarkers, pathogenesis and personalized treatment. However, the role of metabolites in psoriasis is unknown.
Patients and Methods: We performed an untargeted metabolomic analysis of plasma based on high-resolution liquid chromatography mass spectrometry from 10 plaque psoriasis patients and 10 healthy controls.
Results: A total of 301 differential metabolites were detected, of which 10 metabolites were possible potential biomarkers, including vitamins, amino acids, and lipids. At the same time, KEGG pathway enrichment analysis was performed for all detected differential metabolites, and it was found that protein digestion and absorption, amino acid metabolism and lipid metabolism may be jointly involved in regulating the pathogenesis of psoriasis. In addition, the proteins ESR1, OPRM1 and HSD11B1 were identified as possible potential topical therapeutic targets for psoriasis through analysis of the metabolite-protein interaction network.
Conclusion: In this study, we identified 10 differential metabolites as possible potential combinatorial biomarkers for the diagnosis of psoriasis. 12 metabolic pathways were significantly enriched that may be closely related to the occurrence and development of psoriasis. Three proteins, ESR1, OPRM1, and HSD11B1, were identified as possible potential therapeutic targets for psoriasis.
Keywords: psoriasis, metabolomics, biomarker, KEGG pathway, therapeutic target
Introduction
Psoriasis is a non-infectious, chronic, systemic inflammatory skin disease that can manifest at any age and significantly impact the quality of life (QoL) of patients.1,2 According to the World Health Organization (WHO), at least 100 million people worldwide suffered from psoriasis in 2016.3 According to the clinical manifestations of psoriasis, five types of psoriasis are distinguished, namely plaque psoriasis (also known as psoriasis vulgaris, which accounts for about 90% of all cases), erythrodermic psoriasis, pustular psoriasis or generalized pustular psoriasis, guttate or eruptive psoriasis and inverse psoriasis.4 Psoriasis significantly affects the appearance of a patient’s skin and results in a risk for psychiatric comorbidity and suicide.5
Currently, the clinical diagnosis of psoriasis vulgaris mainly includes clinical symptoms, localization and histopathologic changes, and lesion characteristics.6 However, a reliable diagnosis requires a comprehensive evaluation to avoid misdiagnosis and unnecessary treatment. The diagnosis of psoriasis remains a huge challenge due to the non-specific symptoms and lack of diagnostic biomarkers.7 Early studies found that patients with psoriasis often have metabolic abnormalities or disorders, including fatty acids, vitamins, carbohydrates, lipids, and so on.8,9 Therefore, metabolites can be used as biomarkers for the diagnosis of psoriasis and provide a direction for further research into the pathogenesis of psoriasis and safe and effective individualized treatments.
Undoubtedly, the understanding of the immunologic, genetic, and molecular aspects of psoriasis has contributed significantly to the identification of novel drugs, new targets, and holistic approaches in recent decades, leading to the development of increasingly personalized therapeutic targets such as cytokines (tumor necrosis factor [TNF], interleukins [IL] 12/23, 17, and 23) and personalized treatment drugs such as biologics, JAK /TYK inhibitors, and phosphodiesterase-4 (PDE-4) inhibitors.10–12 However, their clinical manifestations still need to be defined and evaluated, so there is still a long way to go before achieving the dream goal of personalized treatment for this disease.11,12 Therefore, it is also imperative to continue to search for new targets that target the characteristics of various psoriasis subtypes.
Metabolomics is a new “omics” discipline that has evolved in recent years into a comprehensive and systematic study of metabolic changes in organisms.13–15 The discovery and identification of small molecule metabolites or alterations in metabolic pathways are also beneficial for understanding the pathophysiological processes of diseases and aiding in the identification of therapeutic targets.15 In recent years, metabolomics research has shown that amino acid metabolism, lipid metabolism, carnitine metabolism, and glycolysis play important roles in psoriasis,16–20 providing valuable perspectives for early diagnosis, disease progression monitoring and targeted therapy of diseases. Although some progress has been made in understanding the complex pathogenesis and in discovering biomarkers for psoriasis. However, despite this progress, further research is needed in some areas, such as diagnostic biomarkers, pathogenesis and targeted therapies.
In this research, we used untargeted metabolomics to detect changes in plasma metabolites in psoriasis based on ultra-high performance liquid chromatography-quadrupole time-of-flight mass spectrometry (UPLC-Q-TOF/MS), to discover the potential biomarkers for distinguishing psoriasis patients from healthy individuals, and to perform the related Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis and metabolite-protein interaction network analysis. This will be helpful for the early diagnosis of psoriasis and provide some reference values for understanding the pathogenesis of the disease and formulating an individualized therapeutic target plan.
Materials and Methods
Materials and methods are detailed in Supplementary Appendix S1.
Briefly, first, the study was approved by the Medical Ethics Committee of the North China Medical and Health Group Xingtai General Hospital (No. ZCKT-2021-0011) and conducted in accordance with the guidelines of the Declaration of Helsinki, and plasma samples were collected from 20 participants after informed consent. Second, an untargeted metabolome based on UPLC-Q-TOF/MS was performed to investigate the changes of metabolites and metabolic pathways in plasma samples of psoriasis. Finally, biomarker analysis and interaction analysis were performed for biomarkers and drug targets. (Table 1)
 |
Table 1 Clinical Characteristics of Participants |
Results
Metabolomic Profile of Plasma Samples from Psoriasis and Healthy Groups
A total of 7597 and 6042 features were detected in positive and negative modes, respectively. As can be seen from the negative and positive TIC overlay charts (Figure 1A and B) of all QC samples, these QC samples completely overlapped without significant retention time and intensity drift, indicating good signal consistency throughout the data acquisition process. In addition, the principal component analysis (PCA) results (Figure 1C and D) showed that the pooled QC samples were well clustered together in both negative and positive ion modes, indicating that the LC-MS data acquisition process was qualified. However, the psoriasis and healthy control groups cannot be distinguished in PCA. Moreover, the two groups are significantly different in the PLS-DA analysis (Figure 2A and B). The models performance was evaluated by monitoring its goodness of fit (R2) and predictive ability (Q2) values. To ensure the reliability of the model, 200 permutation tests were conducted on R2 and Q2, as showed in Figure 2C (positive) and 2D (negative). These results provide a robust assessment of the model’s accuracy and predictive power. The VIP value of a metabolite is the importance of a metabolite to the PLS-DA model, and represents the overall contribution of each metabolite to the model.
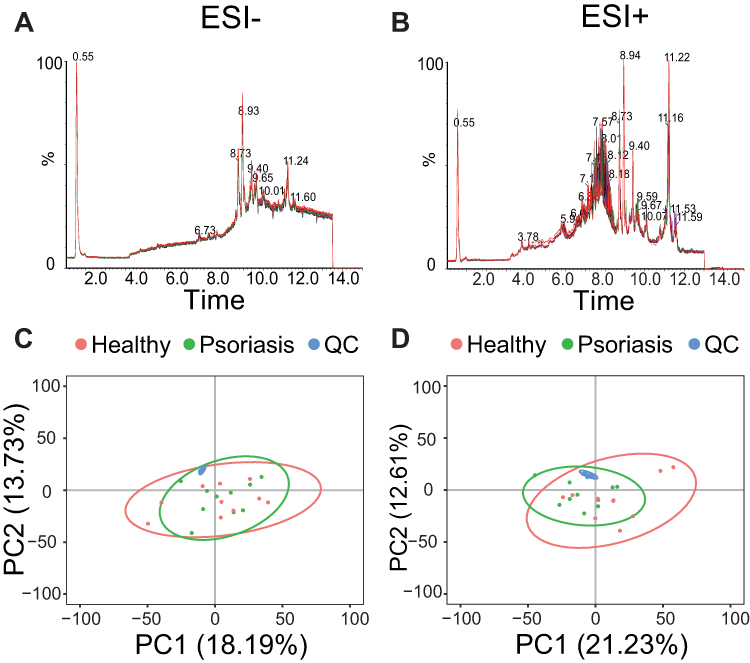 |
Figure 1 Quality control. Overlayed TICs of QC samples in negative mode (A) and positive mode (B). PCA score plots of psoriasis (green), healthy (red), and QC (blue) samples in negative mode (C) and positive mode (D). |
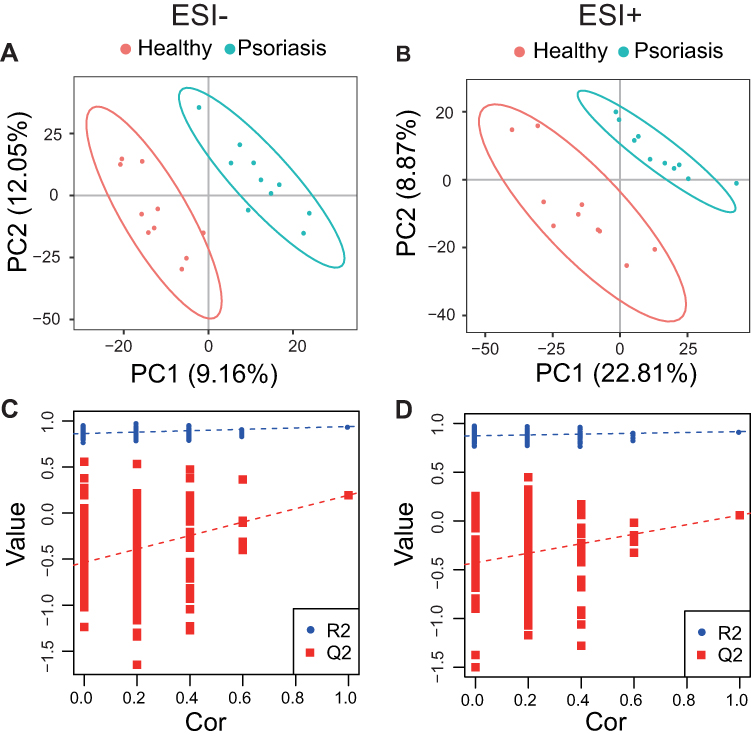 |
Figure 2 PLS-DA analysis. PLS-DA score plots from the healthy and psoriasis groups in (A) negative mode (R2 = 0.916, Q2 = 0.059) and (B) positive mode (R2 = 0.940, Q2 = 0.194). Validation plots were obtained from 200 permutation tests in (C) negative mode and (D) positive mode. |
A total of 3404 metabolites were identified and quantified after data pre-treatment. They included both endogenous and exogenous (eg, food and drugs) metabolites. A total of 301 differentially expressed metabolites were selected by statistical analysis between psoriasis and healthy control groups. These differentially expressed metabolites include lipids (such as sphingolipids, glycerolipids, glycerophospholipids, lysophospholipids, steroids, fatty acyls, prenol lipids), amino acids, carnitines, cholines, phenols and others, as shown in Table S1. Some of these differential metabolites have also been described as biomarkers or hub metabolites.18,20,21
Potential Biomarker Screening and Identification
All differential metabolites were used for univariate ROC analysis, and metabolites with an AUC ≥ 0.8 were selected as potential biomarker candidates. Then, metabolites of endogenous origin were selected from these potential candidate biomarkers, and multivariate ROC analysis was performed using the support vector machine (SVM) model. Finally, 10 differential metabolites with an optimal AUC combination were selected as potential biomarkers, as shown in Table 2. In the multivariate ROC analysis of the SVM model, the AUC of 10 metabolites in the training data set reached 0.989, and the 95% CI was 0.803–1 (Figure 3A). PCA analysis and hierarchical cluster analysis of 10 potential biomarkers can clearly distinguish the two groups of samples (Figures 3B and C). The results of univariate ROC analysis for 10 potential biomarkers are shown in Figure S1.
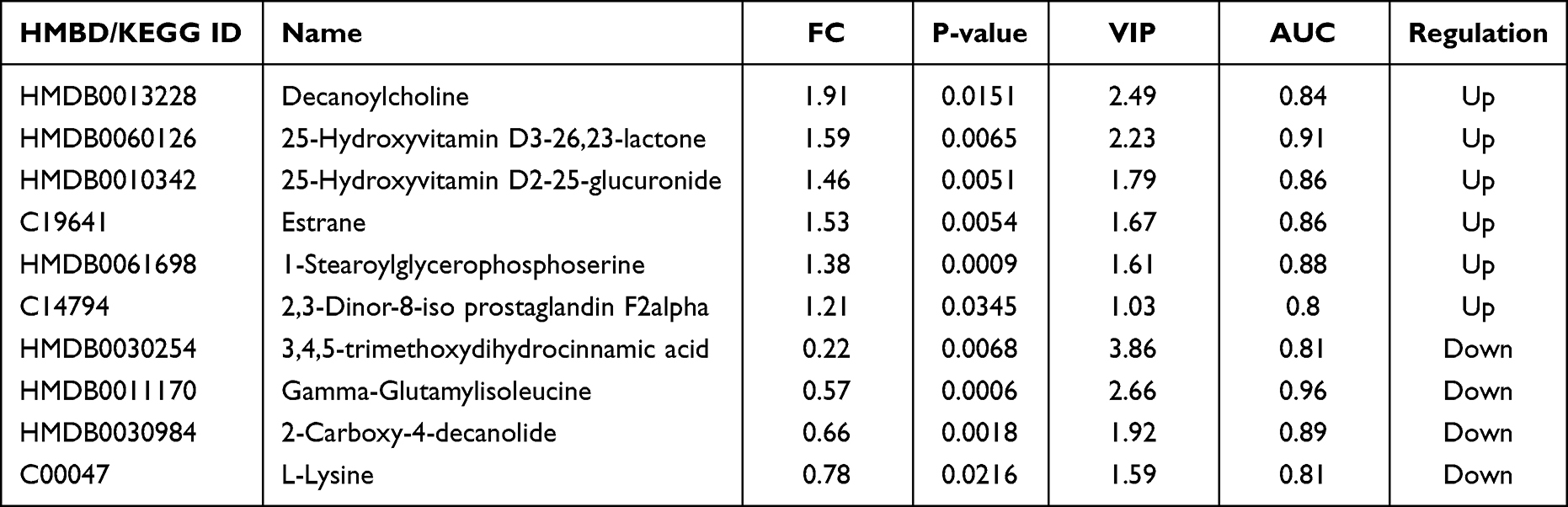 |
Table 2 Potential Biomarkers |
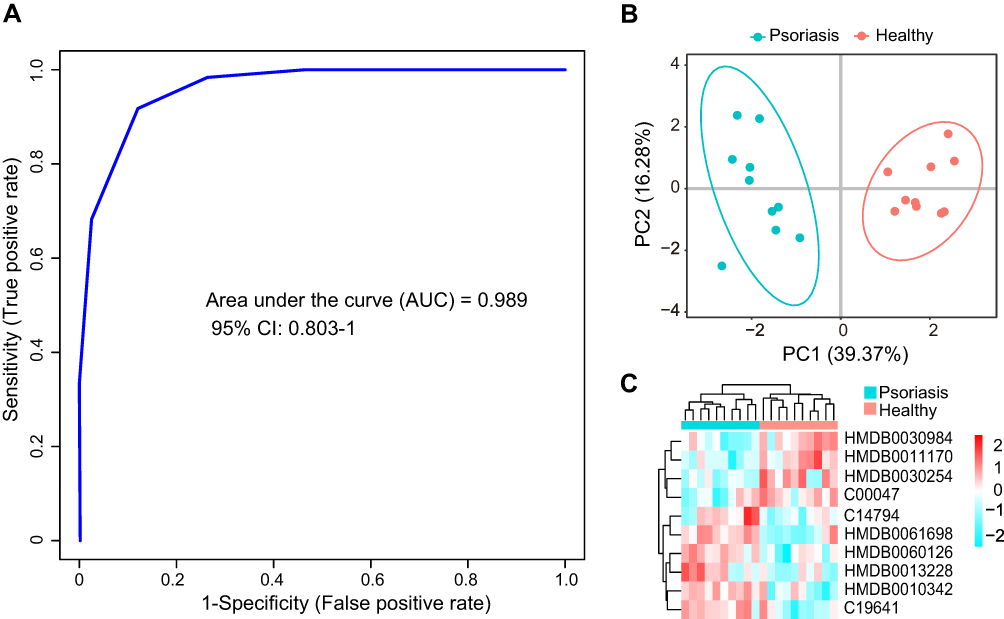 |
Figure 3 Biomarker analysis. (A), multivariate ROC curve analysis based on SVM model to distinguish the predictive power of the psoriasis group from the healthy group. (B), principal component analysis of the psoriasis group and the healthy group based on 10 selected biomarkers. (C), hierarchical cluster analysis of 10 selected biomarkers. |
Metabolic Pathway Enrichment Analysis
Pathway enrichment analysis was performed using information from the KEGG database. The KEGG pathways with p-value less than 0.05 were considered as significantly enriched pathways. The results showed that protein digestion and absorption, biosynthesis of amino acids, valine, leucine and isoleucine degradation, valine, leucine and isoleucine biosynthesis, aminoacyl-tRNA biosynthesis, sphingolipid metabolism, neuroactive ligand-receptor interaction, alpha-linolenic acid metabolism, ABC transporters, biotin metabolism, mineral absorption and mTOR signaling pathways were significantly enriched in patients (Figure 4A). These results suggest that these metabolic pathways may be involved in the regulation of psoriasis. The differential metabolites in these pathways are closely related to psoriasis, especially amino acids and lipids, which have been repeatedly reported to be involved in the process of psoriasis. A network map of enriched metabolic pathways and differential metabolites was generated using Cytoscape software (Figure 4B). Among them, C00123 (L-leucine), C00183 (L-valine), C00047 (L-lysine), C01120 (sphingosine 1-phosphate) and C06124 (sphingosine1-phosphate) connect multiple metabolic pathways, suggesting that these metabolites may be important regulatory molecules of psoriasis.
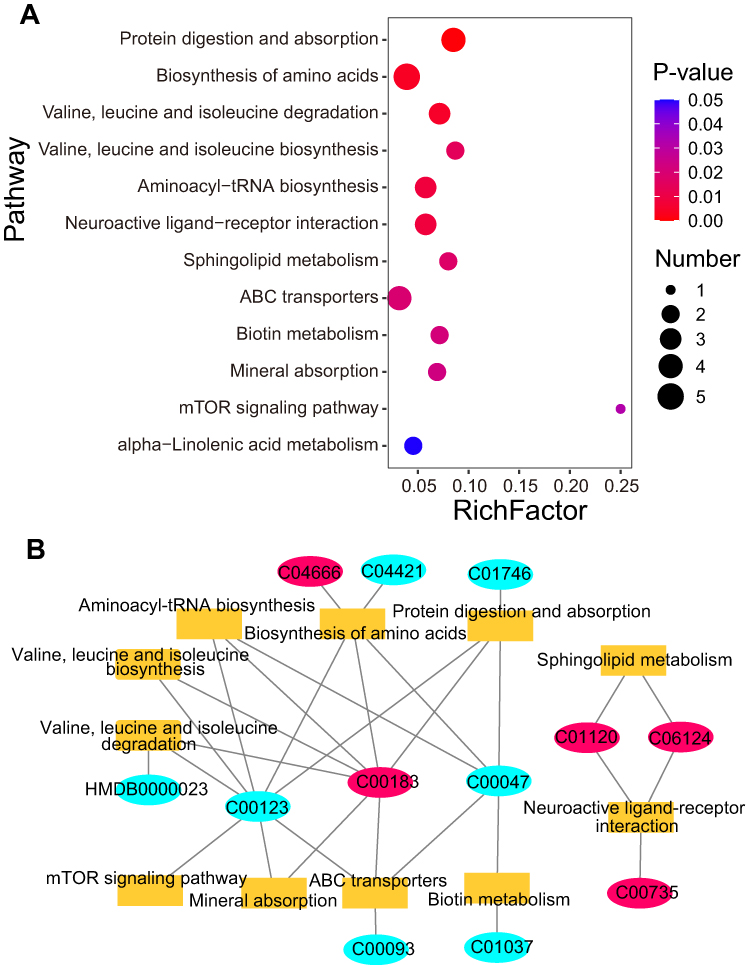 |
Figure 4 KEGG pathway enrichment analysis of differentially expressed metabolites. (A) is significantly enriched KEGG pathway. The X-axis represents the enrichment factor (RichFactor), which represents the number of differential metabolites in each pathway divided by the number of all metabolites in that pathway. The Y-axis represents the names of pathways that are significantly enriched. (B) is the network diagram of enrichment pathways and corresponding metabolites. The rectangle represents pathway, and the oval represents the metabolites. Red represents the up-regulated metabolites in psoriasis, and blue represents the down-regulated metabolites in psoriasis. |
Potential Topical Therapeutic Drug Targets Discovery
Currently, the main drug targets are proteins, such as receptor proteins, enzymes, and ion channels. In search of potential topical therapeutic drug targets for psoriasis, we performed an interaction network analysis using differential metabolites with proteins specifically expressed in the skin and involved in immune response, inflammatory response, or cellular proliferation (considered as psoriasis-related proteins). Network analysis revealed that 14 proteins interacted directly with differential metabolites (Figure 5) and could be potential targets for topical drugs. These proteins included receptors (NR3C1, ESR1, OPRM1, CCR5, HTR1A, FASLG), enzymes (ERBB2, DPP4, HSD11B1, CASP8), and regulators (CFLAR, FADD, KRT18, POMC). In addition, there were another 10 proteins that interacted with the above 14 proteins, which could also serve as indirect drug targets. The total 24 interacting proteins are listed in Table S2. In contrast, there are 11 different metabolites that interact directly with proteins. Among these metabolites, tamoxifen simultaneously interacts with 11 proteins and is a drug for the treatment of hormone receptor-positive breast cancer. In 1996, Ferrari et al22 reported a case of psoriasis and advanced breast cancer in a woman who was treated with tamoxifen for breast cancer and also effectively eliminated the psoriasis. Cortisol interacts with five proteins and is a glucocorticoid secreted and released by the adrenal glands. Cortisol plays a number of important roles in the body, including suppressing inflammation, regulating the stress response, metabolism, blood pressure and blood sugar.
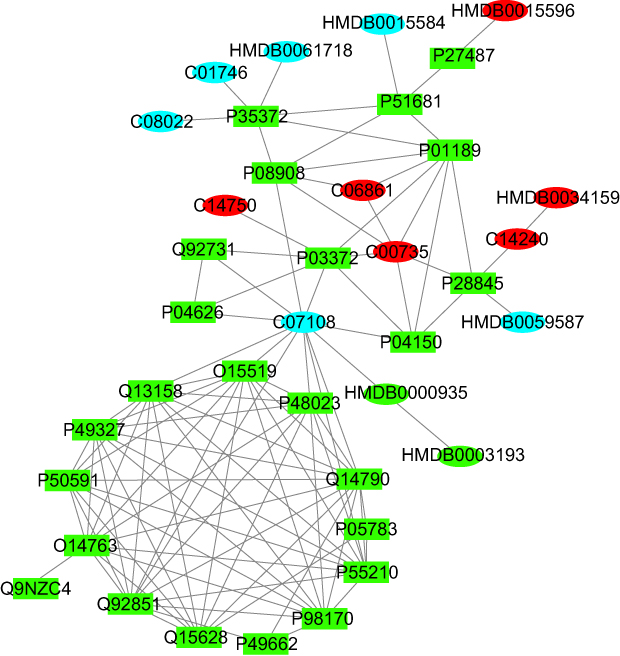 |
Figure 5 Metabolite-protein interaction network analysis. Rectangles represent proteins and ellipses represent metabolites, of which red represents the metabolites that are up-regulated in psoriasis, blue represents the metabolites that are down-regulated in psoriasis, and green represents other substances that interact with the database. |
Discussion
In this study, metabolomics based on UPLC-Q-TOF/MS technique was used to analyze the change of plasma metabolites in patients with psoriasis and healthy control group. The results showed that there were significant differences in metabolites between the two groups. Through univariate statistics and multivariate statistical analysis, 301 differentially expressed metabolites were selected and 10 potential biomarkers were identified by ROC analysis, including vitamins, amino acids, cholines, lipids, and so on, Pathway enrichment analysis of these differential metabolites was performed to find the pathway significantly related to psoriasis. Some of these biomarkers are consistent with previous reports, such as vitamins, amino acids and lipids.18,23–25
Vitamin D is synthesized in the epidermis of the skin and can also be absorbed through the diet. Evidence suggests that vitamin D is involved in the regulation of the immune system and inflammatory processes, is closely related to keratinocyte apoptosis, and is involved in skin barrier integrity.23,26 Because of this ability, vitamin D plays an important role in psoriasis. Previous studies have shown that patients with psoriasis have low vitamin D levels and treatment with vitamin D derivatives can relieve their symptoms. Our result showed that 25-hydroxyvitamin D3-26,23-lactone and 25-hydroxyvitamin D2-25-glucuronide, the vitamin D derivatives, were significantly higher in plasma of psoriasis patients than in healthy controls. We hypothesized that vitamin D is rapidly metabolized in psoriasis due to its involvement in immune and inflammatory responses, resulting in elevation of these two vitamin D derivatives. Therefore, its post-metabolic products 25-hydroxyvitamin D3-26,23-lactone and 25-hydroxyvitamin D2-25-glucuronide can also be used as potential biomarkers for the diagnosis of psoriasis. Excess 25-hydroxyvitamin D3-26,23-lactone and 25-hydroxyvitamin D2-25-glucuronide indicates vitamin D deficiency, which may exacerbate the symptoms of psoriasis. Homeostasis is achieved by the exchange of essential amino acids with non-essential amino acids and the transfer of amino groups from oxidized amino acids to amino acid biosynthesis.27 Therefore, it is not surprising that we have found abnormalities in plasma amino acid levels in psoriasis patients, especially lysine. The essential amino acid lysine, which contributes to the stability of the extracellular matrix as an exogenous inhibitor of plasmin-induced proteolysis,28 has not yet been identified as a potential biomarker for psoriasis. However, it has been widely used as a major component of dietary supplements in the treatment of psoriasis.28,29 Disruption of amino acid metabolism may promote the development of psoriasis by regulating the immune response via nitrogen levels in the body.30 Gamma-glutamyl dipeptide is a bioactive peptide involved in inflammation, oxidative stress, and glucose regulation. Gamma-glutamyl leucine has been widely reported to be associated with the risk of metabolic diseases such as obesity, metabolic syndrome, and type 2 diabetes.31 Additionally, gamma-glutamyl peptides have been identified as potential biomarkers for various diseases, including cancer, diabetes, and liver diseases.32 In our results, we found that gamma-glutamylisoleucine, was significantly reduced in psoriasis and may also be a potential biomarker for psoriasis. Lipids are the major components of the cell membrane and are also involved in the regulation of cell proliferation, apoptosis, immunity, angiogenesis, inflammation, and other biological processes.18,33,34 Lipid disorders are a pathological feature of many diseases, including psoriasis, diabetes, hypertension, cardiovascular disease, and other autoimmune diseases.18,35,36 These lipids may further exacerbate psoriasis by affecting lipid metabolism or cell composition Therefore, some lipids may serve as important biomarkers. In this study, lipids showed significant differences between psoriasis patients and healthy groups, including 1-stearoylglycerophosphoserine, estrane and 2.3-dinor-8-iso prostaglandin F2alpha. Among them, 1-stearoylglycerophosphoserine was reported by Ribeiro et al37 to be associated with sickle cell anaemia and is a putative biomarker for it. Estran is a steroid hydrocarbon that is the basis of several female sex hormones, which is used as biomarkers in hormone-dependent diseases such as breast/ovarian cancer, Turner’s syndrome, precocious puberty and so on.38 2.3-dinor −8-iso prostaglandin F2 alpha is a metabolite of prostaglandin F2 isomerase, which is a useful biomarker for human oxidative status, and could potentially be used as a measurement of oxidative stress.39 In addition, this study found that decanoylcholine, 2-carboxy-4-decanolide and 3,4,5-trimethoxydihydrocinnamic acid, which were first reported as potential specific biomarkers for psoriasis, were also significantly different. The relationship between them and the occurrence and development of psoriasis remains unclear.
Metabolites are substrates and products in the metabolic process and are closely associated with specific phenotypes. Not only can they be produced directly by the host organism or microorganisms, but they can also be ingested from exogenous sources such as food. They are the basic functional substances that drive cells, such as energy production and storage, signal transduction and apoptosis.40 Thus, the metabolome can be used not only to screen of biomarkers but also the related metabolic pathways to improve our understanding of pathophysiology and disease mechanisms.40,41 In our study, metabolic pathway enrichment analysis of differential metabolites was performed. The results showed that some KEGG metabolic pathways were significantly enriched (P<0.05), including amino acid metabolism (including biosynthesis of amino acids, valine, leucine and isoleucine degradation or biosynthesis, aminoacyl-tRNA biosynthesis), protein digestion and absorption, lipid metabolism (including alpha-linolenic acid metabolism and sphingolipid metabolism), neuroactive ligand-receptor interaction, ABC transporters, biotin metabolism, mineral absorption, and mTOR signaling pathway. Differential changes in amino acids indicate disruption of amino acid metabolic pathways, including biosynthesis of amino acids and valine, leucine and isoleucine degradation or biosynthesis. Previous studies have shown that amino acid metabolism is essential for maintaining physiological concentrations of amino acids and nitrogen balance in the body and has evolved to control immune responses, thus being involved in immune-related diseases such as psoriasis, type 1 diabetes, multiple sclerosis, rheumatoid arthritis, and inflammatory bowel disease.30 In addition, our results showed that the protein digestion and absorption pathway was significantly enriched, speculating that it regulates the amino acid metabolism pathway by adjusting amino acid levels in patients with psoriasis. There is evidence that sphingolipid metabolism and inflammatory signaling pathways regulate each other.42 The pathogenesis of psoriasis is also closely related to the inflammatory pathway.43 This suggests that sphingolipid metabolism is also involved in the occurrence of psoriasis. Our results support the above point. In patients with psoriasis, dietary supplementation with α-linolenic acid reduces activation of the T-cell signaling pathway, thereby significantly reducing proliferation and abnormal differentiation of psoriatic keratinocytes.44 When Chen et al45 examined the miRNA-146a target gene in psoriasis, the neuroactive ligand-receptor interaction pathway was found to be significantly enriched, and it was speculated that this pathway is involved in the regulation of psoriasis progression. In this study, the pathway of neuroactive ligand-receptor interaction was also significantly enriched, further suggesting that this pathway may be involved in the regulation of psoriasis progression.
Psoriasis, as an immune-mediated inflammatory skin disease, primarily involves the processes of inflammation, immune response, and cell proliferation. These processes are mediated by the interaction of numerous components, including genes, proteins, and metabolites, which form a complex network.46 Network pharmacology-based drug-target interaction (DTI) has become a paradigm of drug discovery. Not only does it have the advantage of being inexpensive, taking little time, and not relying on 3D structures of targets, but it can also be used in a variety of areas, such as drug target prediction, new drug discovery, and elucidation of potential molecular mechanisms for therapeutic efficacy or drug safety.47 Current topical drugs for the treatment of psoriasis primarily include corticosteroids, vitamin D derivatives, glucocorticosteroids and calcineurin inhibitors.4 Analysis of metabolite-protein interaction network revealed that 14 proteins, including receptors, enzymes, and regulators, interact directly with metabolites commonly used as drug targets. ESR1, also known as estrogen receptor, is a nuclear hormone receptor involved in the regulation of cell proliferation, differentiation, and eukaryotic gene expression. It is commonly used as a therapeutic target for breast cancer.48 Zeng et al49 found that ESR1 is the hub gene in psoriasis as revealed by microarray gene expression profiles and is involved in regulating apoptosis of psoriatic cells. OPRM1, known as the mu-type opioid receptor, is the main target of clinical opioid analgesics.48 Kupczyk et al50 showed that imbalanced expression of OPRM1 in psoriasis may lead to disturbances in neuro epidermal homeostasis, which could exacerbate itch. HSD11B1, or 11-beta-hydroxysteroid dehydrogenase 1, is involved in corticosteroid receptor-mediated anti-inflammatory responses as well as homeostasis and metabolic processes. Research by Sarkar et al51 has shown that HSD11B1 plays a critical role in the regulation of cortisol synthesis and promotes the conversion of cortisone to cortisol, whereas cortisol, a glucocorticoid hormone, inhibits the inflammatory response of psoriatic keratinocytes. Currently, HSD11B1 is being used as a drug target in 12 clinical trials, 4 discontinued and preclinical drugs for the treatment of lupus, influenza virus infection, type 2 diabetes, metabolic disorders and so on.48 Interestingly, the expression of ESR1, OPRM1 and HSD11B1 proteins was down-regulated in patients with psoriasis,49–51 suggesting that activation or overexpression of these proteins may alleviate the symptoms of psoriasis. Our network interaction analysis revealed that these three proteins interact with most different metabolites, so we speculated that these three proteins are the most likely drug targets for the treatment of psoriasis. Activators of these three proteins could be beneficial for the treatment of psoriasis.
With advances in understanding the pathophysiology of psoriasis, the discovery of new key molecules, and the development of novel drugs and targets, treatment options for this disease have expanded significantly.12 However, personalized therapy for psoriasis still faces challenges due to the unique disease characteristics of each individual patient. Even in the same patient, psoriasis characteristics can vary widely.10 Therefore, it may be difficult to meet the needs of patients with a single drug for the treatment of psoriasis and its complications. In this study, we selected psoriasis patients with the same characteristics while searching for biomarkers and therapeutic targets by monitoring their metabolite changes (which can provide real-time feedback on changes in the patient’s phenotype). This could open a new potential avenue for early diagnosis and personalized, comprehensive treatment of specific diseases.
However, due to financial reasons, the sample size is not large, the representativeness of all results may have certain defects. If funding for follow-on applications is sufficient, we will conduct further investigation and verification of potential biomarkers and drug targets for screening, such as targeted quantitative verification with a large sample and validation of animal models of disease. At the same time, the pathogenesis of psoriasis has not been explored extensively enough. Follow-up should also incorporate various “omics” data, including genomics, transcriptomics, proteomics and epigenomics, to explore the regulatory mechanism of psoriasis and provide a basis for personalized diagnosis and treatment.
Conclusion
In summary, we identified 10 possible potential combinatorial biomarkers for psoriasis using metabolomics that can be used for auxiliary diagnosis and clinical application. At the same time, 12 metabolic pathways were found to be significantly involved in psoriasis regulation occurrence and development, deepening the understanding of disease pathogenesis. Finally, Network pharmacology integrates network analysis identified three optimal possible potential topical therapeutic targets. Therefore, it can provide more information on biochemical changes and metabolic alterations for the study of psoriasis, which is of great importance for the comprehensive prevention and treatment of the disease.
Data Sharing Statement
The raw data of mass spectrometry are deposited in the MetaboLights database52 and can be accessed at www.ebi.ac.uk/metabolights/MTBLS5185.
Funding
This study received financial support from the Xingtai Science and Technology Plan in 2017 (Project number 2017ZC051).
Disclosure
The authors report no conflict of interest.
References
1. Prignano F, D’Erme AM, Bonciolini V, Lotti T. Mucosal psoriasis: a new insight toward a systemic inflammatory disease. Int J Dermatol. 2011;50(12):1579–1581. doi:10.1111/j.1365-4632.2010.04864.x
2. Michalek IM, Loring B, John SM. A systematic review of worldwide epidemiology of psoriasis. J Eur Acade Dermatol Venereol. 2017;31(2):205–212. doi:10.1111/jdv.13854
3. Organization WH. Global Report on Psoriasis. World Health Organization; 2016.
4. Boehncke WH, Schön MP. Psoriasis. Lancet. 2015;386(9997):983–994. doi:10.1016/S0140-6736(14)61909-7
5. Pompili M, Innamorati M, Trovarelli S, et al. Suicide risk and psychiatric comorbidity in patients with psoriasis. J Int Med Res. 2016;44(IS):61–66. doi:10.1177/0300060515593253
6. McHugh NJ, Balachrishnan C, Jones SM. Progression of peripheral joint disease in psoriatic arthritis: a 5-yr prospective study. Rheumatology. 2003;42(6):778–783. doi:10.1093/rheumatology/keg217
7. Rida MA, Chandran V. Challenges in the clinical diagnosis of psoriatic arthritis. Clin immunol. 2020;214:108390. doi:10.1016/j.clim.2020.108390
8. Mallbris L, Granath F, Hamsten A, Ståhle M. Psoriasis is associated with lipid abnormalities at the onset of skin disease. J Am Acad Dermatol. 2006;54(4):614–621. doi:10.1016/j.jaad.2005.11.1079
9. Murzaku EC, Bronsnick T, Rao BK. Diet in dermatology: part II. Melanoma, chronic urticaria, and psoriasis. J Am Acad Dermatol. 2014;71(6):1053.e1–1053.e16. doi:10.1016/j.jaad.2014.06.016
10. Claudia C-D, María-Elena V-H, Josué V-E, María-Carmen B-C, Alain-Raimundo R-O, Martha-Estrella G-P. Small molecules under development for psoriasis: on the road to the individualized therapies. Arch Dermatol Res. 2020;312(9):611–627. doi:10.1007/s00403-020-02056-3
11. Camela E, Potestio L, Fabbrocini G, Ruggiero A, Megna M. New frontiers in personalized medicine in psoriasis. Expert Opin Biol Ther. 2022;22(12):1431–1433. doi:10.1080/14712598.2022.2113872
12. Camela E, Potestio L, Fabbrocini G, Pallotta S, Megna M. The holistic approach to psoriasis patients with comorbidities: the role of investigational drugs. Expert Opin Investig Drugs. 2023;32(6):537–552. doi:10.1080/13543784.2023.2219387
13. Rinschen MM, Ivanisevic J, Giera M, Siuzdak G. Identification of bioactive metabolites using activity metabolomics. Nat Rev Mol Cell Biol. 2019;20(6):353–367. doi:10.1038/s41580-019-0108-4
14. Wishart DS. Metabolomics for Investigating Physiological and Pathophysiological Processes. Physiol Rev. 2019;99(4):1819–1875. doi:10.1152/physrev.00035.2018
15. Qiu S, Cai Y, Yao H, et al. Small molecule metabolites: discovery of biomarkers and therapeutic targets. Signal Transduct Target Ther. 2023;8(1):132. doi:10.1038/s41392-023-01399-3
16. Kamleh MA, Snowden SG, Grapov D, et al. LC-MS metabolomics of psoriasis patients reveals disease severity-dependent increases in circulating amino acids that are ameliorated by anti-TNFα treatment. J Proteome Res. 2015;14(1):557–566. doi:10.1021/pr500782g
17. Kang H, Li X, Zhou Q, et al. Exploration of candidate biomarkers for human psoriasis based on gas chromatography-mass spectrometry serum metabolomics. Br J Dermatol. 2017;176(3):713–722. doi:10.1111/bjd.15008
18. Zeng CW, Wen B, Hou GX, et al. Lipidomics profiling reveals the role of glycerophospholipid metabolism in psoriasis. Gigascience. 2017;6(10):1–11. doi:10.1093/gigascience/gix087
19. Li SS, Liu Y, Li H, et al. Identification of psoriasis vulgaris biomarkers in human plasma by non-targeted metabolomics based on UPLC-Q-TOF/MS. Eur Rev Med Pharmacol Sci. 2019;23(9):3940–3950. doi:10.26355/eurrev_201905_17823
20. Chen C, Hou GX, Zeng CW, Ren Y, Chen X, Peng C. Metabolomic profiling reveals amino acid and carnitine alterations as metabolic signatures in psoriasis. Theranostics. 2021;11(2):754–767. doi:10.7150/thno.51154
21. Li L, Lu CJ, Han L, et al. Untargeted serum metabonomics study of psoriasis vulgaris based on ultra-performance liquid chromatography coupled to mass spectrometry. Oncotarget. 2017;8.
22. Ferrari VD, Jirillo A. Psoriasis and tamoxifen therapy: a case report. Tumori. 1996;82(3):262–263.
23. Barrea L, Savanelli MC, Di Somma C, et al. Vitamin D and its role in psoriasis: an overview of the dermatologist and nutritionist. Rev Endocr Metab Disord. 2017;18(2):195–205. doi:10.1007/s11154-017-9411-6
24. Hambly R, Kirby B. The relevance of serum vitamin D in psoriasis: a review. Arch Dermatol Res. 2017;309(7):499–517. doi:10.1007/s00403-017-1751-2
25. Souto-Carneiro M, Tóth L, Behnisch R, et al. Differences in the serum metabolome and lipidome identify potential biomarkers for seronegative rheumatoid arthritis versus psoriatic arthritis. Ann Rheum Dis. 2020;79(4):499–506. doi:10.1136/annrheumdis-2019-216374
26. Mattozzi C, Paolino G, Richetta AG, Calvieri S. Psoriasis, vitamin D and the importance of the cutaneous barrier’s integrity: an update. J Dermatol. 2016;43(5):507–514. doi:10.1111/1346-8138.13305
27. Bröer S, Bröer A. Amino acid homeostasis and signalling in mammalian cells and organisms. Biochem J. 2017;474(12):1935–1963. doi:10.1042/BCJ20160822
28. Rath M, Pauling L. Plasmin-induced proteolysis and the role of apoprotein (a), lysine and synthetic analogs. J Orthomolecular Med. 1992;7:17–23.
29. Wong AP, Kalinovsky T, Niedzwiecki A, Rath M. Efficacy of nutritional treatment in patients with psoriasis: a case report. Exp Ther Med. 2015;10(3):1071–1073. doi:10.3892/etm.2015.2631
30. Mondanelli G, Iacono A, Carvalho A, et al. Amino acid metabolism as drug target in autoimmune diseases. Autoimmun Rev. 2019;18(4):334–348. doi:10.1016/j.autrev.2019.02.004
31. Wu Q, Li J, Zhu J, et al. Gamma-glutamyl-leucine levels are causally associated with elevated cardio-metabolic risks. Front Nutr. 2022;9:936220. doi:10.3389/fnut.2022.936220
32. Thacker JB, He C, Pennathur S. Quantitative analysis of γ-glutamylisoleucine, γ-glutamylthreonine, and γ-glutamylvaline in HeLa cells using UHPLC-MS/MS. J Sep Sci. 2021;44(15):2898–2907. doi:10.1002/jssc.202001266
33. Huse M, Le Floc’h A, Liu X. From lipid second messengers to molecular motors: microtubule-organizing center reorientation in T cells. Immunol Rev. 2013;256(1):1. doi:10.1111/imr.12116
34. Huse M. Lipid-based patterning of the immunological synapse. Biochem Soc Trans. 2014;42(6):1506–1511. doi:10.1042/BST20140191
35. Yan D, Afifi L, Jeon C, et al. The metabolomics of psoriatic disease. Psoriasis. 2017;7.
36. Angelakopoulou A, Shah T, Sofat R, et al. Comparative analysis of genome-wide association studies signals for lipids, diabetes, and coronary heart disease: cardiovascular Biomarker Genetics Collaboration. Eur Heart J. 2012;33(3):393–407. doi:10.1093/eurheartj/ehr225
37. Ribeiro PR, Teixeira RDS, Souza AR, et al. Blood plasma metabolomics of children and adolescents with sickle cell anaemia treated with hydroxycarbamide: a new tool for uncovering biochemical alterations. Br J Haematol. 2021;192(5):922–931. doi:10.1111/bjh.17315
38. Gea M, Toso A, Schilirò T. Estrogenic activity of biological samples as a biomarker. Sci Total Environ. 2020;740:140050. doi:10.1016/j.scitotenv.2020.140050
39. Zhang H. Quantification of the oxidative damage biomarker 2,3-dinor-8-isoprostaglandin-F(2alpha) in human urine using liquid chromatography-tandem mass spectrometry. Anal Biochem. 2010;399(2):302–304. doi:10.1016/j.ab.2009.12.024
40. Johnson CH, Ivanisevic J, Siuzdak G. Metabolomics: beyond biomarkers and towards mechanisms. Nat Rev Mol Cell Biol. 2016;17(7):451–459. doi:10.1038/nrm.2016.25
41. Laíns I, Gantner M, Murinello S, et al. Metabolomics in the study of retinal health and disease. Prog Retin Eye Res. 2019;69:57–79. doi:10.1016/j.preteyeres.2018.11.002
42. Norris GH, Blesso CN. Dietary and Endogenous Sphingolipid Metabolism in Chronic Inflammation. Nutrients. 2017;9(11):11. doi:10.3390/nu9111180
43. Gisondi P, Bellinato F, Girolomoni G, Albanesi C. Pathogenesis of Chronic Plaque Psoriasis and Its Intersection With Cardio-Metabolic Comorbidities. Front Pharmacol. 2020;11:117. doi:10.3389/fphar.2020.00117
44. Morin S, Simard M, Rioux G, Julien P, Pouliot R. Alpha-Linolenic Acid Modulates T Cell Incorporation in a 3D Tissue-Engineered Psoriatic Skin Model. Cells. 2022;11(9):9. doi:10.3390/cells11091513
45. Chen HX, Liu YS, Zhang XJ. TargetScore used to reveal potential targets of miRNA203 and miRNA-146a in psoriasis by integrating microRNA overexpression and microarray data. Medicine. 2018;97(41):. doi:10.1097/MD.0000000000012671
46. Gu J, Li L, Wang D, et al. Deciphering metabonomics biomarkers-targets interactions for psoriasis vulgaris by network pharmacology. Ann Med. 2018;50(4):323–332. doi:10.1080/07853890.2018.1453169
47. Mashaki Ceyhan E, Gürsöz WH, Alkan A. The Turkish Medicines and Medical Devices Agency: comparison of Its Registration Process with Australia, Canada, Saudi Arabia, and Singapore. Front Pharmacol. 2018;9:9. doi:10.3389/fphar.2018.00009
48. Zhou Y, Zhang YT, Lian XC, et al. Therapeutic target database update 2022: facilitating drug discovery with enriched comparative data of targeted agents. Nucleic Acids Res. 2022;50(D1):D1398–D1407. doi:10.1093/nar/gkab953
49. Zeng X, Zhao JJ, Wu XH, et al. PageRank analysis reveals topologically expressed genes correspond to psoriasis and their functions are associated with apoptosis resistance. Mol Med Rep. 2016;13(5):3969–3976. doi:10.3892/mmr.2016.4999
50. Kupczyk P, Reich A, Hołysz M, et al. Opioid Receptors in Psoriatic Skin: relationship with Itch. Acta Derm Venereol. 2017;97(5):564–570. doi:10.2340/00015555-2595
51. Sarkar MK, Kaplan N, Tsoi LC, et al. Endogenous Glucocorticoid Deficiency in Psoriasis Promotes Inflammation and Abnormal Differentiation. J Invest Dermatol. 2017;137(7):1474–1483. doi:10.1016/j.jid.2017.02.972
52. Haug K, Cochrane K, Nainala VC, et al. MetaboLights: a resource evolving in response to the needs of its scientific community. Nucleic Acids Res. 2019;48(D1):D440–D444.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Fibrinogen-Like Protein 1 as a Novel Biomarker of Psoriasis Severity
Sun X, Liu L, Chen S, Wang J, Cai X, Song J, Zhou M, Guo D, Kuai L, Ding X, Li B, Li X
Journal of Inflammation Research 2022, 15:4637-4647
Published Date: 15 August 2022
The SELP, CD93, IL2RG, and VAV1 Genes Associated with Atherosclerosis May Be Potential Diagnostic Biomarkers for Psoriasis
Liu S, Liu F, Zhang Z, Zhuang Z, Yuan X, Chen Y
Journal of Inflammation Research 2023, 16:827-843
Published Date: 27 February 2023
A Nomogram for Early Diagnosis of Community-Acquired Pneumonia Based on Bronchoalveolar Lavage Fluid Metabolomics
Chen S, Su M, Lei W, Wu Z, Wu S, Liu J, Huang X, Chen G, Zhang Q, Zhong H, Rong F, Li X, Xiao Q
Infection and Drug Resistance 2023, 16:1237-1248
Published Date: 1 March 2023
Association of Complete Blood Cell Count-Derived Inflammatory Biomarkers with Psoriasis and Mortality
Zhao Y, Yang XT, Bai YP, Li LF
Clinical, Cosmetic and Investigational Dermatology 2023, 16:3267-3278
Published Date: 13 November 2023
Distinct Airway Microbiome and Metabolite Profiles in Eosinophilic and Neutrophilic Asthma
Liu S, Lin Z, Zhou J, Yang X, You L, Yang Q, Li T, Hu Z, Zhan X, Jiang Y, Sun B
Journal of Asthma and Allergy 2025, 18:1003-1022
Published Date: 13 June 2025