")
Back to Journals » Drug Design, Development and Therapy » Volume 17
Antitumor Activity of Berberine by Activating Autophagy and Apoptosis in CAL-62 and BHT-101 Anaplastic Thyroid Carcinoma Cell Lines
Authors Shi XZ, Zhao S, Wang Y , Wang MY , Su SW, Wu YZ, Xiong C
Received 30 January 2023
Accepted for publication 6 June 2023
Published 26 June 2023 Volume 2023:17 Pages 1889—1906
DOI https://doi.org/10.2147/DDDT.S406354
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Jianbo Sun
Xiang-Zhe Shi,1,2 Sheng Zhao,3 Yan Wang,1,2 Meng-Yao Wang,1,2 Su-Wen Su,1,2 Yan-Zhao Wu,3 Chen Xiong1,2
1The Key Laboratory of Neural and Vascular Biology, Ministry of Education, Hebei Medical University, Shijiazhuang, 050017, People’s Republic of China; 2The Key Laboratory of Pharmacology and Toxicology for New Drugs, Department of Pharmacology, Hebei Medical University, Shijiazhung, 050017, People’s Republic of China; 3Department of Otorhinolaryngology-Head and Neck Surgery, 4th Hospital of Hebei Medical University, Shijiazhuang, 050011, People’s Republic of China
Correspondence: Chen Xiong; Yan-Zhao Wu, Email [email protected]; [email protected]
Introduction: Anaplastic thyroid carcinoma (ATC) is the most lethal thyroid carcinoma. Doxorubicin (DOX) is the only drug approved for anaplastic thyroid cancer treatment, but its clinical use is restricted due to irreversible tissue toxicity. Berberine (BER), an isoquinoline alkaloid extracted from Coptidis Rhizoma, has been proposed to have antitumor activity in many cancers. However, the underlying mechanisms by which BER regulates apoptosis and autophagy in ATC remain unclear. Thus, the present study aimed to assess the therapeutic effect of BER in human ATC cell lines CAL-62 and BHT-101 as well as the underlying mechanisms. In addition, we assessed the antitumor effects of a combination of BER and DOX in ATC cells.
Methods: The cell viability of CAL-62 and BTH-101 with treatment of BER for different hours was measured by CCK-8 assay, and cell apoptosis was assessed by clone formation assay and flow cytometric analysis. The protein levels of apoptosis protein, autophagy-related proteins and PI3K/AKT/mTORpathway were determined Using Western blot. Autophagy in cells was observed with GFP-LC3 plasmid using confocal fluorescent microscopy. Flow cytometry was used to detect intracellular ROS.
Results: The present results showed that BER significantly inhibited cell growth and induced apoptosis in ATC cells. BER treatment also significantly upregulated the expression of LC3B-II and increased the number of GFP-LC3 puncta in ATC cells. Inhibition of autophagy by 3-methyladenine (3-MA) suppressed BER-induced autophagic cell death. Moreover, BER induced the generation of reactive oxygen species (ROS). Mechanistically, we demonstrated that BER regulated the autophagy and apoptosis of human ATC cells through the PI3K/AKT/mTOR pathways. Furthermore, BER and DOX cooperated to promote apoptosis and autophagy in ATC cells.
Conclusion: Taken together, the present findings indicated that BER induces apoptosis and autophagic cell death by activating ROS and regulating the PI3K/AKT/mTOR signaling pathway.
Keywords: anaplastic thyroid carcinoma, berberine, autophagy, apoptosis, PI3K/AKT/mTOR, reactive oxygen species
Introduction
Anaplastic thyroid carcinoma (ATC) is one of the most lethal aggressive malignancies and is characterized by high levels of rapid proliferation, extrathyroidal invasion, and distant metastasis. Unfortunately, unlike differentiated thyroid carcinoma, most ATC patients are resistant to conventional treatment, including surgery, radiotherapy, chemotherapy (multimodal therapy), and immunotherapy.1,2 Due to a lack of effective treatment measures, ATC is the major cause of all thyroid carcinoma-related deaths with a median survival time of 3 to 9 months, and the 1-year overall survival rate of ATC patients after diagnosis is only 20%.3,4
In recent years, natural compounds from traditional Chinese medicinal plants have received increasing attention due to their potential antitumor activities and few side effects.5,6 Berberine (BER) is a type of isoquinoline alkaloid. As the main component of Coptis chinensis, BER has been widely used in the clinical treatment of gastroenteritis and other intestinal infectious diseases with almost no toxic side effects. Modern studies have also found that BER has many drug properties, such as antitumor and cardioprotective effects.7–11 Although studies have shown that BER inhibits a variety of tumor cells, only a few studies included thyroid cancer. Through a literature search, we found that BER inhibits the proliferation of thyroid cancer cells in a dose–time-dependent manner, while the cytotoxicity of normal thyroid cells to BER is relatively low.12,13
Apoptosis is a process of programmed cell death. Unlike apoptosis, autophagy is normally a cell-survival process that maintains cellular homeostasis by removing misfolded proteins and damaged organelles, thereby providing additional nutrients and energy to the cell.14 In cancer therapy, autophagy behaves as a double-edged sword because it has a cytoprotective effect in some situations but is an alternative cell death pathway (namely, type II PCD) in other situations.15 Thus, autophagy-dependent cell death provides molecular mechanisms and implications for cancer therapy. Growing evidence has demonstrated that BER, as a potent natural autophagic modulator, induces or inhibits autophagy in a variety of cancer cells.16–18 Thus, further research is needed to determine whether BER regulates the growth of ATC cells by modulating autophagy.
Many studies have shown that the phosphatidylinositide-3 kinase (PI3K)/protein kinase B (AKT) signaling pathway activates mammalian rapamycin target protein (mTOR), thus regulating autophagy.19,20 More importantly, the PI3K/AKT/mTOR signaling pathway plays critical roles in the development and progression of various human cancers, including ATC.21,22 However, it is unclear whether PI3K/AKT/mTOR signaling mediates BER-mediated autophagy in ATCs.
DOX is the only drug approved by the Food and Drug Administration (FDA) for ATC treatment based on the guidelines.23 However, its clinical usage and efficacy are hindered due to the cumulative and irreversible cardiotoxicity. Previous studies have shown that BER combined with DOX does not interfere with the antitumor effect of DOX but significantly promotes the sensitivity and antitumor effect of DOX on a variety of human cancer cells.24,25 Moreover, our recent studies have reported that BER alleviates DOX-induced cardiotoxicity in vitro and in vivo.26,27 However, it has not been reported whether BER enhances the anti-ATC effects of DOX, and the underlying molecular mechanisms of BER have not yet been elucidated.
The aims of the present study were to investigate the anticancer effects of BER on ATC cancer cell lines in vitro. We also analyzed the level of autophagy and apoptosis after BER treatment and elucidated the role of the PI3K/Akt/mTORsignaling pathway in the regulation of cell death. Additionally, we aimed to further investigate the effects of BER on the sensitivity of ATC cells to DOX.
Materials and Methods
Cell Culture and Chemicals
The CAL-62 and BHT-101 human ATC cell lines were obtained from the China Center for Type Culture Collection (CCTCC, Wuhan, China) and cultured in Roswell Park Memorial Institute (RPMI) 1640 medium and DMEM (Gibco, USA), respectively, with 10% fetal bovine serum (ExCell Bio, China) and 1% penicillin/streptomycin (BI, Israel). All cells were incubated at 37 °C with 5% CO2. Berberine chloride was purchased from Solarbio (China) and dissolved in water and then RPMI-1640 or DMEM culture medium was added to the desired concentration, waiting to be used. Bafilomycin A1 (BafA1), rapamycin (RA), SC79, wortmannin (WOR), MK-2206,3-methyladenine (3-MA) and N-acetylcysteine (NAC) were purchased from MCE (NJ, USA).
Cell Survival Assay
A CCK-8 kit (Roche Diagnosis, Indianapolis, IN, USA) was used to measure cell viability. ATC cells were plated in 96-well plates at a density of 4×103 cells per well 24 h before treatment. ATC cells were then treated with various concentrations of BER for 24, 48, and 72 h. At each time point, 10 µL of CCK-8 reagent was added into each well for an additional 2 h at 37 °C, and a microplate reader (NanoDrop, USA) was utilized to detect the absorbance at 450 nm. The cell survival rate is presented as the absorbance relative to that of controls.
Flow Cytometry Assay
The flow cytometric apoptosis assays were performed according to the manufacturer's instructions. In brief, the Annexin V-FITC Apoptosis Detection kit (BD Pharmingen, USA) was used to detect cell apoptosis. Cells were treated with various concentrations of BER for 72 h. Both attached and floating cells were harvested, washed twice with ice-cold PBS, and suspended in 300 µL of binding buffer. Cells were stained with 5 µL of Annexin V-FITC and incubated at room temperature for 15 min in the dark, and they were then stained with 5 µL of propidium iodide (PI) and incubated at room temperature for 5 min in the dark. Finally, 200 µL 1× binding buffer was added to each sample, and apoptosis was analyzed by flow cytometry (Agilent Novocyte, USA).
Colony Formation Assay
A colony-forming assay was performed to determine clonogenic cell death. Monolayer culture was performed to measure colony formation. ATC cells (700/well) were seeded into 6-well plates and treated with various doses of BER, and the medium was refreshed every 72 h. After 15 days of culture for colony formation, cells were fixed with 4% paraformaldehyde for 10 min and then washed with PBS, and the colonies were stained with a 1% crystal violet solution for 15 min. Each assay was performed in triplicate, and the surviving fraction was determined.
Western Blot Analysis
After treatment with BER for 72 h, cells were harvested and lysed with RIPA buffer (Solarbio, China) containing PMSF (Solarbio, China), and the protein concentration was quantified using BCA assays (Lianke Biotech). The proteins were electrophoresed using 8–12% SDS‒PAGE gels and then transferred to PVDF membranes (Millipore). The membranes were then blocked in 5% nonfat milk for 1.5 h and incubated at 4 °C overnight with the following primary antibodies: P62 (MBL, 1:1000), LC3B (GeneTex, 1:1000), cleaved caspase 3 (Abcam, 1:500), cleaved PARP1 (Abcam, 1:1000), PI3K (Abcam, 1:1000), p-PI3K (Abcam, 1:1000), AKT (Abcam, 1:10000), p-AKT (Abcam, 1:1000), mTOR (Abcam, 1:1000), p-mTOR (Abcam, 1:1000) and GAPDH (ABclonal, 1:6000). The membranes were further incubated with the corresponding secondary antibodies (Rockland, USA) for 2 h at RT. The protein bands were detected using the Odyssey system.
Quantification of GFP-LC3 Puncta
ATC cells were transfected with a pCDNA2 GFP-LC3 plasmid using Lipofectamine 3000 (Invitrogen, USA) in serum- and antibiotic-free medium for 12 h followed by treatment with BER for 12 h. The accumulation of GFP-LC3 was examined by fluorescence microscopy (Olympus, Japan).
Measurement of Cytosolic ROS
An ROS Assay Kit (Beyotime, Shanghai, China) was used to determine ROS levels using 2′,7′-dichlorofluorescein diacetate (DCFH-DA) by flow cytometry analysis. ATC cells were treated with BER and the ROS inhibitor, NAC, for 24 h at 37 °C, and they were then incubated in RPMI-1640 medium containing 5 µmol/mL DCHF-DA for 30 min at room temperature. The fluorescence intensity was determined using a flow cytometer (Agilent Novocyte, USA), and the data were analyzed using FlowJo software.
Statistical Analysis
The data analysis was performed at least three times using GraphPad Prism 8.0, and the results are expressed as the mean ± SD. Student’s t-tests were performed to evaluate the significant differences between the control and treated groups, and P values <0.05 were considered statistically significant.
Results
BER Inhibits the Viability and Proliferation of ATC Cells
To investigate the effects of BER on the proliferation of ATC cells, the viability of two anaplastic thyroid carcinoma cell lines (CAL-62 and BHT-101) was evaluated. We first evaluated the cytotoxicity of BER at different concentrations (10, 20, 40, 80, 160 and 200 μM) after 24, 48 and 72 h of treatment by the CCK-8 assay. The results showed that BER significantly inhibited the viability of both ATC cell lines in a time- and dose-dependent manner (Figure 1A). The half-maximal inhibitory concentration (IC50) of BER in CAL-62 cells was 124.60, 40.18 and 30.39 µM at 24, 48 and 72 h, respectively. Similarly, the IC50 values of BER in BHT-101 cells treated for 24, 48 and 72 h were 70.29, 38.44 and 22.92 µM, respectively. The strongest effect of BER on the inhibition of cell growth was observed 72 h after treatment. According to the CCK-8 assay results of the examined cell lines, BHT-101 was the most sensitive cell line to BER. Hence, we subsequently used 80 and 40 µΜ BER for 72 h to treat CAL-62 and BHT-101 cells. Furthermore, the number of colonies in the BER-treated cells was significantly decreased with increasing BER concentrations (Figure 1B). Taken together, these data suggested that BER inhibits cell growth and proliferation in CAL-62 and BHT-101 cells.
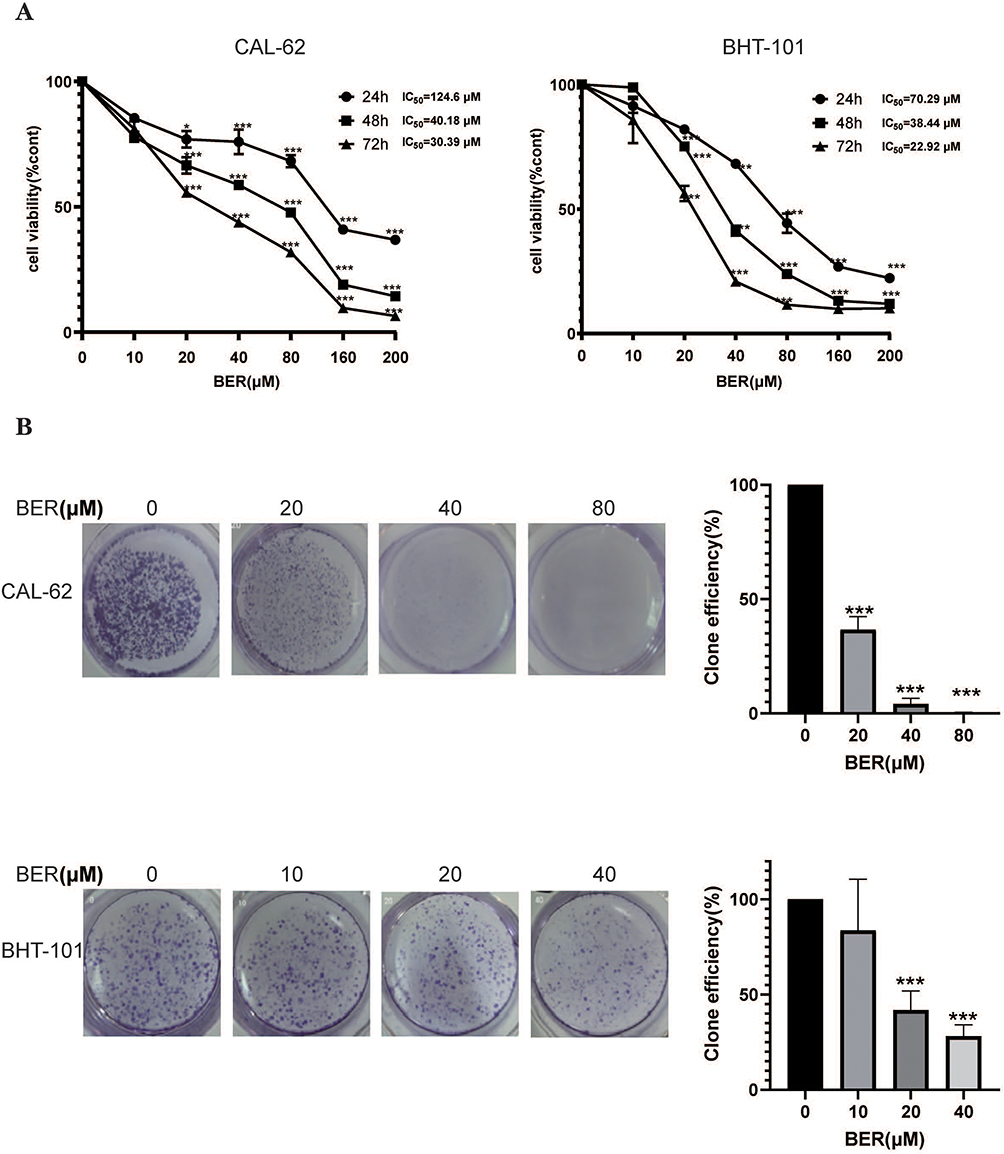 |
Figure 1 BER inhibits the viability and proliferation of ATC cells. (A) The CAL-62 and BHT-101 ATC cell lines were incubated with increasing doses of BER (0–200 µM) for 24, 48, and 72 h to analyze cell proliferation using the CCK-8 assay. (B) BER inhibited colony formation of ATC cell lines. Representative images of CAL-62 and BHT-101 cell contact-dependent colony formation. Each experiment is representative of three independent experiments. The data are presented as the mean ± SD of three separate experiments; *P <0.05, **P < 0.01, and ***P < 0.001 vs 0 µM BER, berberine; IC50, half-maximal inhibitory concentration. |
BER Induces Apoptosis in ATC Cells
To determine whether the inhibitory proliferative effect of BER in CAL-62 and BHT-101 cells was associated with apoptosis, the expression levels of cleaved caspase 3 and cleavedPARP1, two apoptosis protein biomarkers,28 in BER-treated ATC cells were examined. Figure 2A shows that BER significantly increased the protein expression of cleaved caspase 3 and cleavedPARP1 in both ATC cell lines in a dose-dependent manner. Furthermore, CAL-62 and BHT-101 cells were exposed to various concentrations of BER for 72 h and analyzed by flow cytometry (Figure 2B). Flow cytometry analysis indicated that the apoptosis rates induced by BER treatment at 20, 40 and 80 µM were 46.32±1.96%, 56.17±2.57% and 66.27±1.09%, respectively, in CAL-62 cells. In addition, the apoptosis rates induced by BER treatment at 10, 20 and 40 µM in BHT-101 cells were 4.02±0.41%, 8.90±0.32% and 7.55±0.91%, respectively (Figure 2C).
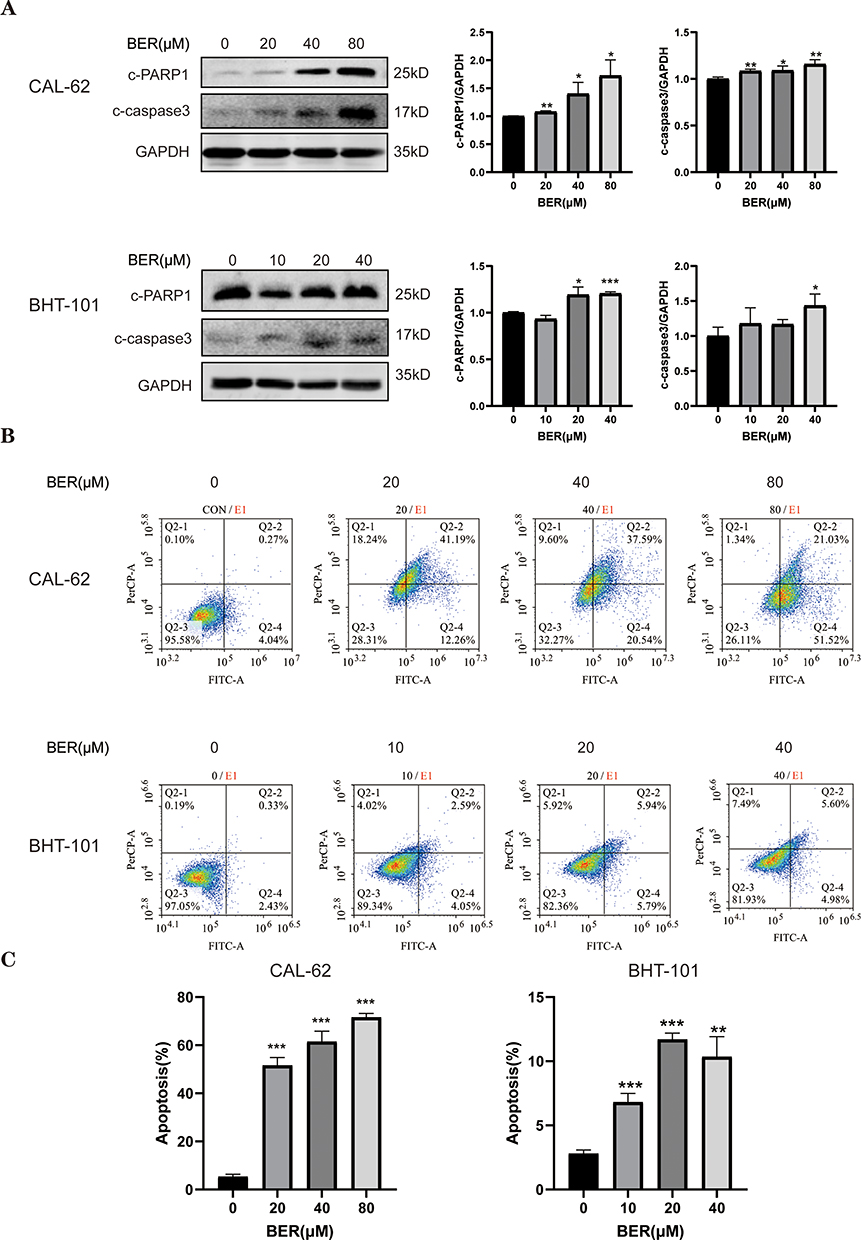 |
Figure 2 BER induces apoptosis in ATC cells. (A) The expression of apoptosis-related proteins was evaluated by Western blot analysis. CAL-62 and BTH-101 cells were treated with a series of concentrations of BER for 72 h, and Western blot analysis was used to evaluate the levels of apoptosis pathway-related proteins using anti-PARP1 and anti-caspase-3 antibodies. (B) AV/PI flow cytometric analysis was used to detect the effect of BER on the apoptosis of ATC cells. To determine the percentage of early and late apoptotic populations, the cells were dual stained with dUTP-FITC and PI and detected by flow cytometric analysis.(C) The cell apoptosis rate was calculated from the flow cytometry results. Data are presented as the mean ± SD. All data are representative of three independent experiments. *P< 0.05, **P< 0.01, and ***P < 0.001 vs 0 µMBER. |
BER Induces Autophagy in ATC Cells
BER exerts anticancer activity in various cancer types by inducing autophagy.29 To explore whether BER triggers autophagy in ATC cells, we first detected the protein expression of autophagy-related factors, including LC3 and p62, in CAL-62 and BHT-101 cells. Because the conversion of LC3-I to LC3-II is commonly used to evaluate autophagy,30 we evaluated the conversion of LC3-I to LC3-II. After BER treatment for 72 h, the LC3‑II/LC3‑I ratio (LC3B) was significantly increased, and the levels of p62, a substrate of autophagosome degradation,31 were also increased (Figure 3A). To further confirm that BER causes autophagy in ATC cells, an GFP-LC3 puncta autophagy assay was used to monitor autophagy through direct fluorescence microscopy. In CAL-62 and BHT101 cells treated with 80 and 40 µM BER, respectively, GFP-LC3 puncta were significantly increased compared to untreated cells (Figure 3B), which demonstrated that BER induced autophagy in ATC cells.
 |
Figure 3 BER-induced autophagy contributes to cell death in ATC cells. (A) The expression of autophagy-related proteins was evaluated in CAL-62 and BHT-101 cells after treatment with the indicated concentrations of BER for 72 h by Western blot analysis. GAPDH was used as a control. Representative column diagrams show the results of relative protein expression. (B) CAL-62 and BHT-101 cells were transiently transfected with GFP-LC3 expression plasmids for 12 h followed by treatment with 80 or 40 μM BER for 72 h. Scale bar, 100 μm. (C) ATC cells were pretreated with or without 2 μM BafA1for 2 h followed by treatment with or without BER for 72 h. Cells were then harvested for Western blot analysis to examine the LC3‑II/LC3‑I ratio (LC3B) and p62 levels. GAPDH was used as a control. Representative column diagrams showing the results of relative protein expression. (D) The cells were cultured with BER for 72 h in the absence or presence of 2 mM 3-MA, and Western blot analysis of the LC3‑II/LC3‑I ratio (LC3B) and c-PARP1 was performed. (E) Cells were treated as in D, and apoptosis was detected by flow cytometry. (F) The cell apoptosis rate was calculated from the flow cytometry results. (G) Cells were treated as in D, and the inhibitory rate was measured by CCK-8 assays. Data are expressed as the mean ± SD from three independent experiments. *P < 0.05, **P < 0.01, and ***P < 0.001 vs 0 µM BER or the control group; #P < 0.05, ##P < 0.01, and ###P < 0.001 vs the BER group. |
Because the accumulation of LC3-II may be attributed to an increase in autophagosome formation or a decrease in lysosomal fusion and degradation, cells were incubated with BafA1, an inhibitor of the autophagosome, to block autophagic flux. The results showed that BafA1 (2μM) treatment elicited a significant increase in the LC3‑II/LC3‑I ratio (LC3B) and p62 expression in ATC cells treated with BER (Figure 3C), which excluded the possibility of lysosomal dysfunction causing LC3-II accumulation. These data revealed that BER treatment induces autophagy in ATC cells.
Because the above data indicated that BER promotes both apoptosis and autophagy in ATC cells, we further investigated the role of BER-induced autophagy in cell death by pretreating cells the 3-MA (2 mM), an autophagy inhibitor for 2 h followed by BER treatment for 72 h. We then calculated the cell viability of BER-treated ATC cells with or without pretreatment with 3-MA. When ATC cells were cotreated with BER and 3-MA, the BER‐induced accumulation of LC3B was significantly blocked, and the expression of c-PARP1 was significantly decreased (Figure 3D). Flow cytometry analysis was utilized to further explore the combination of BER and 3-MA. Compared to the BER group, cotreatment with BER and 3-MA efficiently reduced cell apoptosis (Figure 3E and F). In addition, ATC cells pretreated with 3-MA showed significantly enhanced viability compared to BER-treated ATC cells (Figure 3G). These results indicated that the antiproliferative and proapoptotic effects of BER on ATC cells were decreased under cotreatment with BER and 3-MA. Thus, these findings suggested that autophagy may play a partial but significant role in BER-induced cell growth inhibition.
BER Induces Cytotoxicity via the PI3K/AKT/mTOR Signaling Pathway
As the PI3K/AKT/mTOR pathway has been demonstrated to be related to cancer progression and cell autophagy,19,20 we investigated whether the PI3K/AKT/mTOR signaling pathway is involved in the effects of BER in ATC. Compared to control cells, Western blot analysis demonstrated that the ratios of p-PI3K/PI3K and p-mTOR/mTOR were significantly lower in both types of cancer cells treated with BER, while the ratios of p-AKT/AKT were increased in CAL-62 and BHT-101 cells treated with BER (Figure 4A). Interestingly, although BER blocked mTOR activation, it simultaneously activated AKT. These results suggested that the BER-induced mTOR inhibition was not dependent on its effect on AKT but that BER-induced AKT activation may be due to mTOR inhibition. Thus, the AKT/mTOR pathway, as well as its feedback loop, are more likely involved in these processes.32 These results indicated that the mechanism by which BER inhibits the proliferation of ATC cells may involve modulation of the PI3K/AKT/mTOR signaling pathway.
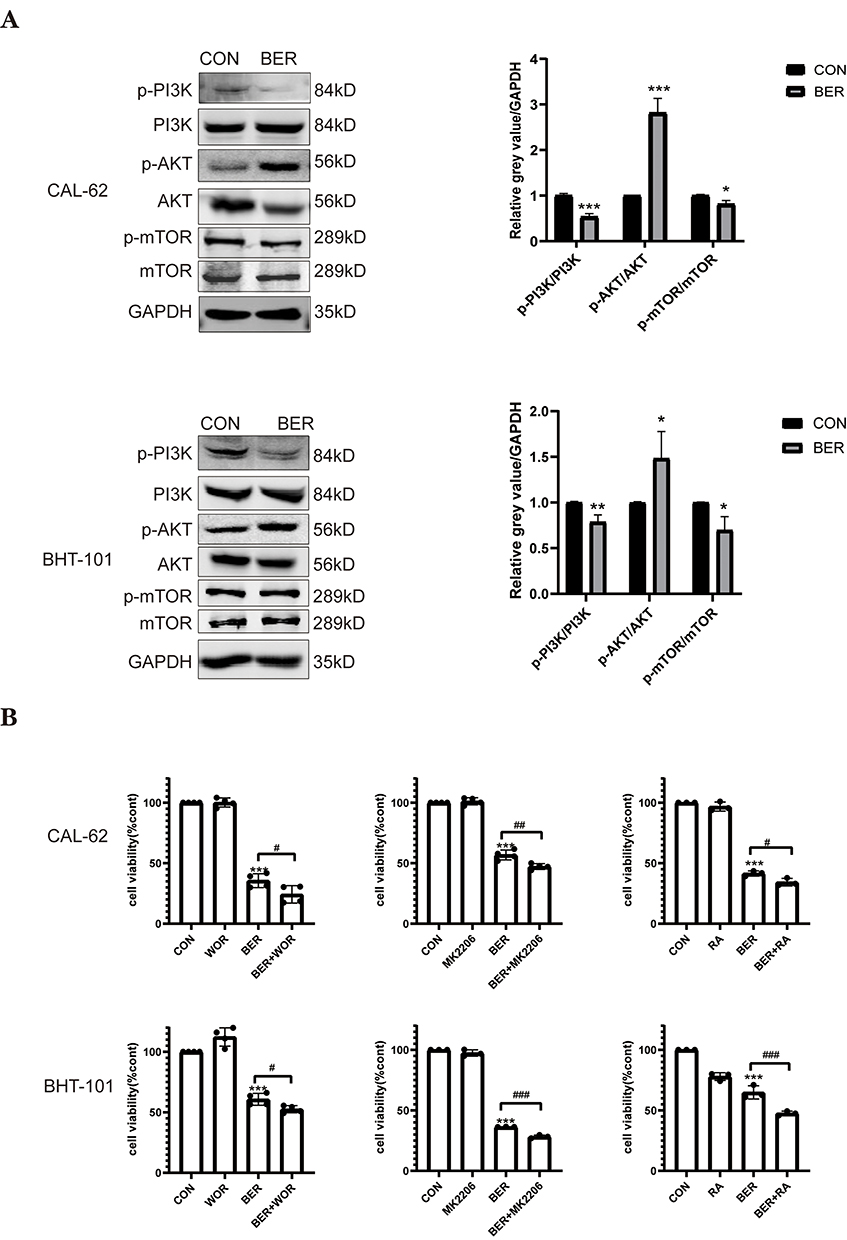 |
Figure 4 BER induce cytotoxicity via the PI3K/AKT/ mTOR signaling pathway. (A) The phosphorylation levels of PI3K, AKT, and mTOR in ATC cells treated with or without BER for 72 h were detected by Western blot analysis. Quantification of the relative gray value of bands compared to GAPDH. (B) Effects of the PI3K/AKT/mTOR signaling pathways on BER-induced inhibition of ATC cell viability. CAL-62 and BHT-101 cells were pretreated with 1 μMWOR (a PI3K inhibitor), 0.5 μMMK2206 (an AKT inhibitor), or 0.2 µMRA (an mTOR inhibitor) for 2h prior to 80 or 40 μM BBR treatment for 72 h, and cell viability was detected using CCK-8 assays. Data are expressed as the mean ± SD from three independent experiments. *P<0.05, **P<0.01, and ***P<0.001 vs. the control group; #P < 0.05, ##P < 0.01, and ###P < 0.001 vs the BER group. |
To further confirm whether the PI3K/AKT/mTOR pathway participates in BER-triggered cytotoxicity in anaplastic thyroid cancer cells, we tested whether cotreatment with WOR (a PI3K inhibitor), MK-2206 (an AKT inhibitor), and RA (an mTOR inhibitor) enhances the BER-induced reduction in cellular growth. The CCK8 assay demonstrated that although BER inhibited the viability of ATC cells, RA, WOR, and MK-2206 pretreatment increased the inhibitory effect of BER on ATC cell viability (Figure 4B). Therefore, these data demonstrated that BER suppresses ATC cell growth by regulating the PI3K/AKT/mTOR signaling pathway.
BER Mediates Autophagy and Apoptosis by Regulating the PI3K/Akt/mTOR Signaling Pathway
We next investigated whether the PI3K/AKT/mTOR pathway participates in BER-induced apoptosis and autophagy in ATC cells. Cells were treated with BER for 72 h with or without pretreatment with the WOR, SC79 (an AKT activator) and RA for 30 min. Western blot analysis showed that the expression of cleaved PARP and the LC3‑II/LC3‑I ratio (LC3B) were increased in cells treated with both WOR and BER compared to BER-treated cells (Figure 5A). Flow cytometry analysis showed that ATC cells treated with BER and WOR had a higher apoptotic rate than those treated with BER alone (Figure 5B). Moreover, the apoptosis and autophagy-related protein levels were similarly increased after BER treatment alone and cotreatment with RA, an inhibitor of mTOR (Figure 5E). Moreover, flow cytometry analysis indicated that the apoptotic rates of cells cotreated with BER and RA were greater than those in cells treated with BER alone (Figure 5F). Additionally, Western blot analysis demonstrated that pretreatment of ATC cells with SC79 significantly reduced the BER-induced the LC3II/I ratio and cleavedPARP1 expression (Figure 5C). Flow cytometry analysis showed that cells treated with BER and SC79 had a lower apoptotic rate than cells treated with BER alone (Figure 5D). Hence, activation of AKT via SC79 treatment may relieve the autophagy and apoptosis induced by BER. Taken together, these findings indicated that the PI3K/AKT/mTOR signaling pathway may have a vital role in the regulation of BER‐induced autophagy and apoptosis.
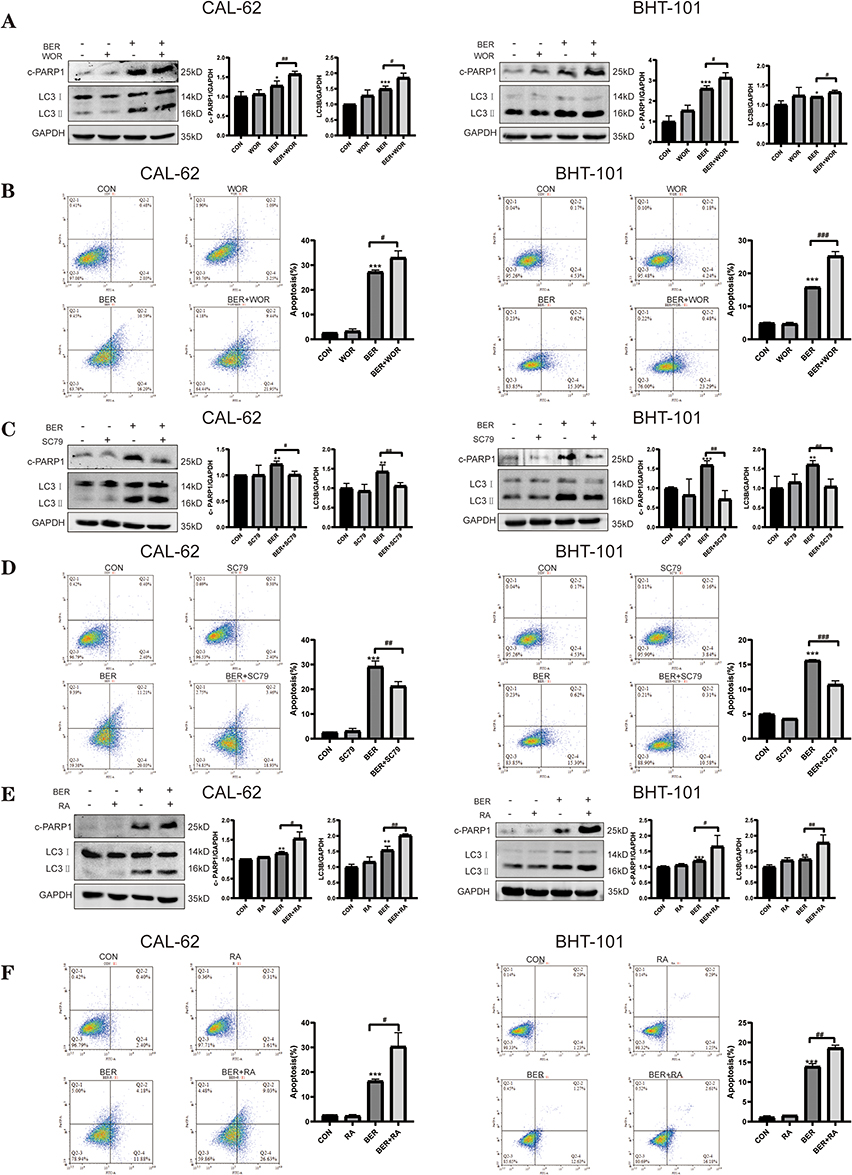 |
Figure 5 BER mediates autophagy and apoptosis by regulating the PI3K/AKT/mTOR signaling pathway. CAL-62 and BHT-101 cells were pretreated with 1 μMWOR (a PI3K inhibitor), 4 μg/mlSC79 (an AKT activator), and 0.2 μM RA (anmTOR inhibitor) for 2 h prior to 80 or 40 μM BBR treatment for 72 h. (A) Cells were treated with or without BER in combination with or without 1 μM WOR, and the levels of the apoptosis-related protein, cleaved PARP1, and the autophagy-related protein, LC3B, were determined by Western blot analysis. (B) Cells were treated as in A, and apoptosis was detected using flow cytometry. (C) Cells were treated with or without BER in combination with or without 4 μg/mL SC79, and the levels of the apoptosis-related protein, cleaved PARP1, and the autophagy-related protein, LC3B, were determined by Western blot analysis. (D) Cells were treated as in C, and apoptosis was detected using flow cytometry. (E) Cells were treated with or without BER in combination with or without 0.2 μM RA, and the levels of the apoptosis-related protein, cleaved PARP1, and the autophagy-related protein, LC3B, were determined by Western blot analysis. (F) Cells were treated as in E, and apoptosis was detected using flow cytometry analysis. Data are expressed as the mean ± SD of triplicate experiments. *P < 0.05, **P < 0.01, and ***P < 0.001 vs the control group; #P < 0.05, ##P < 0.01, and ###P < 0.001 vs the BER group. |
BER Induces ATC Cell Autophagy and Apoptosis via ROS Accumulation
Previous studies have demonstrated that ROS are a major factor in BER-induced apoptosis.33,34 Excessive ROS production induces tumor cell autophagy and apoptosis.35,36 To further investigate the mechanism by which BER promotes autophagy and apoptotic responses in ATC cells, the DCHF-DA oxidation-activated fluorescent dye was used to detect intracellular ROS via flow cytometry analysis. BER treatment significantly increased ROS levels, whereas pretreatment with the NAC, an ROS inhibitor significantly decreased ROS levels, which suggested that BER stimulated ROS generation in ATC cells (Figure 6A). Furthermore, pretreatment with NAC significantly reversed the BER-induced decrease in cell viability in ATC cells (Figure 6B). To further investigate whether BER-induced ROS triggers autophagy and apoptosis, the LC3‑II/LC3‑I ratio (LC3B) and cleavedPARP1 were detected by Western blot analysis, which demonstrated that NAC inhibited BER-induced autophagy and apoptosis (Figure 6C). These results demonstrated that increased ROS production is one of the upstream events contributing to BER-induced activation of autophagy and apoptosis. Furthermore, the effects of NAC and BER on apoptosis were detected by flow cytometry analysis. Figure 6D shows that NAC alone did not affect the apoptosis rate of ATC cells but that NAC reduced the apoptosis rate of BER-treated ATC cells (P < 0.01) or P <0.001). Collectively, these data indicated that BER induces cellular ROS accumulation, thereby promoting apoptosis and autophagic cell death.
 |
Figure 6 BER induces ATC cell autophagy and apoptosisvia ROS accumulation. (A) After being treated with 80 or 40 µM BER and/or 2 mM NAC (an ROS inhibitor) for 72 h, CAL-62 and BHT-101 cells were stained with the DCHF-DA oxidation-activated fluorescent dye and analyzed using flow cytometry to detect intracellular ROS. (B) Cell viability was evaluated by the CCK-8 assay. (C) ATC cells were treated as in A, and Western blot analysis was used to measure the protein levels of cleaved PARP1 (an apoptosis-related protein), and the protein levels of LC3B (an autophagy-related protein). (D) Cells were treated as in A, and apoptosis was detected using flow cytometry. Data are expressed as the mean ± SD of triplicate experiments. **P< 0.01 and ***P< 0.001 vs the control group; #P< 0.05, ##P < 0.01, and ###P < 0.001 vs the BER group. |
BER Promotes the Sensitivity of ATC Cells to DOX
DOX has shown encouraging clinical activity as a chemotherapeutic for ATC.23 Because BER has antitumor activities toward ATC cells, we next tested the effect of a combination of BER and DOX on the proliferation of ATC cells. We first treated CAL-62 cells with increasing concentrations of DOX for different time points (48–72 h), and cell viability was measured and quantified by the CCK-8 assay. As shown in Figure 7A, DOX inhibited cell growth in a dose- and time-dependent manner. The IC50 values of DOX were 0.298 and 0.098 μM at 48 and 72 h in CAL-62 cells, respectively. To investigate the combinatorial anticancer effects of BER and DOX, we treated CAL-62 cells with DOX at concentrations of 0.2, 0.5 and 1 μM combined with 40 μM BER. Compared to 0.5 and 1 μM DOX alone, the combination of DOX and BER did not induce a reduction in cell viability. When the concentration of DOX was reduced to 0.2 μM, the cell viability was significantly reduced by adding BER (Figure 7B), which indicated that BER reduced the dosage of low concentrations of DOX. These findings suggested that BER enhanced the sensitivity of ATC cells to DOX treatment. Furthermore, Chou-Talalay analysis37 was used to calculate the combination index (CI) of BER and DOX for ATC cells. CI values less than one are defined as synergism, while a CI greater than one is defined as antagonism. As shown in Table 1, BER combined with DOX exhibited a synergistic inhibitory effect in CAL-62 cells with CI values of 0.80–0.97 (CI < 1). Thus, we selected BER (40 μM) and DOX (0.2 μM) as the final experimental concentrations for further analysis. The above results revealed that BER may improve the sensitivity of ATC cells to DOX treatment.
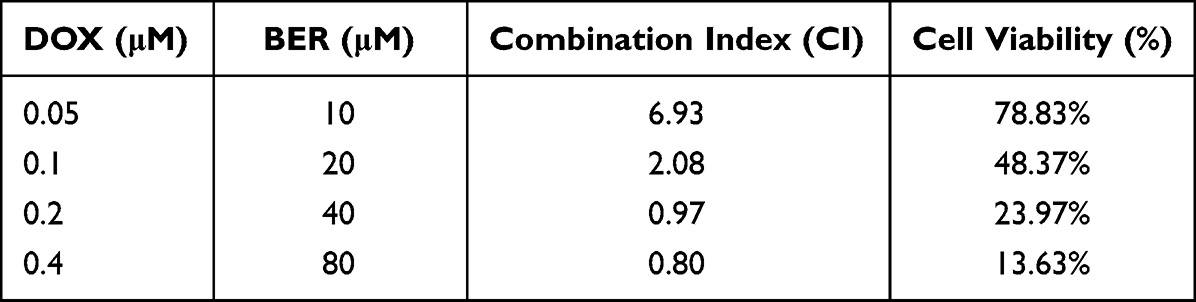 |
Table 1 The Combination Index (CI) of BER and DOX Was Calculated for the CAL-62 Cell |
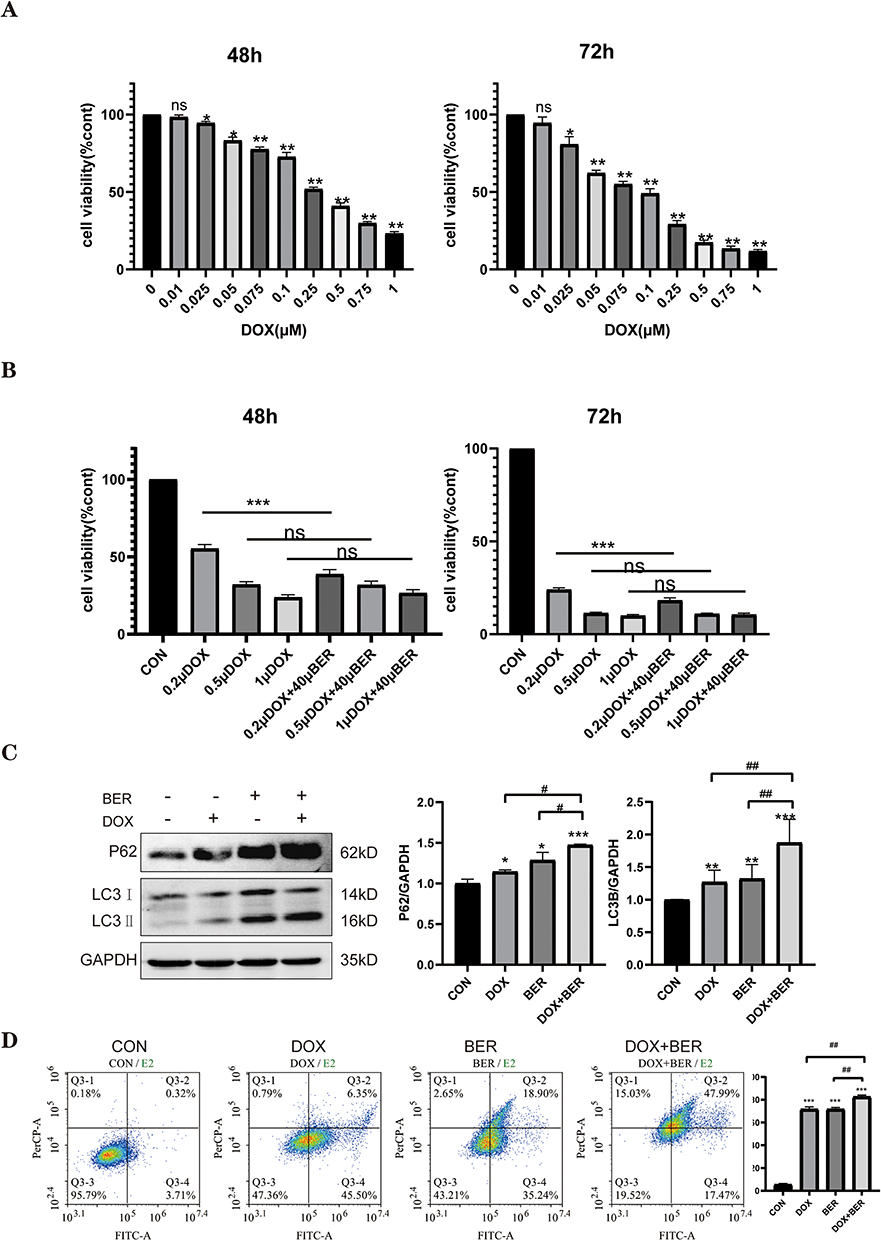 |
Figure 7 Effects of BER, DOX, and their combination on the growth of ATC cell lines. (A) CAL-62 cells were treated with different concentrations of DOX for 48 or 72 h. Cell viability was detected using a CCK-8 assay. ns indicates no significant difference. (B) CAL-62 cells were treated with different concentrations of DOX (0.2, 0.5, and 1 μM) in the presence or absence of BER (40 μM) for 72 h. ns indicates no significant difference. (C) The expression of the autophagy-related proteins, LC3 and p62, was evaluated by Western blot analysis. (D) Flow cytometry was employed to evaluate cell apoptosis. Data are expressed as the mean ± SD of triplicate experiments. *P< 0.05, **P< 0.01 and ***P< 0.001 vs the control group; #P< 0.05 and ##P < 0.01 vs the BER group or DOX group. |
Because BER induces autophagic death in ATC cells, we investigated whether cotreatment with DOX and BER enhances autophagy induction. As shown in Figure 7C, both BER and DOX alone increased the relative expression of the LC3B and p62 autophagy‐promoting proteins in CAL-62 cells compared to the control (P< 0.05 or P< 0.01). Interestingly, BER and DOX combination therapy had higher expression levels of LC3B and p62 compared to either treatment alone (P < 0.05 or P< 0.01). Furthermore, the apoptotic effect of the combination of BER and DOX was further explored by flow cytometry analysis. The results showed that a combination of BER and DOX enhanced ATC cell apoptosis compared to DOX monotherapy (Figure 7D). Collectively, these results provided additional evidence that the combination of BER with DOX promotes apoptosis and autophagy activation in human ATC cells.
Discussion
BER, the main medicinal component of Coptis chinensis, is a natural plant product with many applications. BER is accepted as a promising anticancer agent in different types of tumors through multiple mechanisms.38 Recently, Gao et al reported that BER enhances the sensitivity of breast cancer cells to different conventional chemotherapeutic drugs.39 Pretreatment with BER is effective in promoting the antitumor effects of 5-fluorouracil and cisplatin in laryngeal cancer cells.40 Importantly, BER has attracted much attention because it has low toxicity in relatively high doses and exhibits significant therapeutic activities without major adverse effects.41 However, the efficacy of BER against ATC has rarely been reported, and the potential molecular mechanism remains unknown. The present study showed that BER induced a decrease in cell viability, apoptosis, and autophagy in CAL-62 and BHT-101 cells. Furthermore, combining DOX with BER generated a synergistic anticancer effect in ATC cells.
Autophagy is commonly considered to play a dual role in tumor suppression and cancer promotion, which may provide novel opportunities for anticancer drug development.15 BER, as a potential autophagy modulator, may trigger or inhibit autophagy and exert its anticancer effects.29 For instance, BER induces autophagic cell death in breast cancer cells while inhibiting autophagy in lung cancer cells, suggesting the cytoprotective role of autophagy.42,43 Further research is consequently needed to determine whether BER exhibits anticancer effects through modulation of autophagy in ATC cells. As previously reported, the presence of LC3-II has been considered an indicator of autophagosome formation.29 In the present study, we found that BER triggered autophagy and promoted the conversion of LC3-I to LC3-II. Interestingly, the level of p62 also increased, which is related to the degradation of autophagy. As a stress protein, p62 is also affected by oxidative stress and toxic substances. When these conditions act on cells, p62 also increases, which may be accompanied by an increase in autophagy.44 BER also promoted an increase in the LC3-II green fluorescent puncta number in the cytoplasm. In the presented study, we used BafA1 to understand the role of BER-induced autophagy. When cells were cotreated with BER and BafA1, the BER-induced LC3-II and p62 accumulation was greater than that of cells treated with BER alone. Thus, these findings indicated that autophagy occurred in BER-treated ATC cancer cells. To further investigate the role of autophagy in BER-induced apoptosis of ATC cells, we used 3-MA, a classical inhibitor of autophagy. The combination of BER with 3-MA significantly attenuated the production of LC3-II and cleavedPARP1compared to treatment with BER alone, which partly indicated that the BER-regulated autophagy may be a form of cell death and not a protective mechanism. These results suggested that BER represses human ATC cell growth by inducing cytostatic autophagy.
Autophagy and apoptosis are two distinct processes that maintain homeostasis, both of which are associated with the death of tumor cells. The present results indicated that the protein levels of cleaved caspase 3 and cleavedPARP1 were significantly upregulated by BER in a concentration-dependent manner. In addition, the inhibition of ATC cell proliferation was decreased after combined treatment with BER and 3‐MA compared to BER treatment alone. RA, an autophagy inducer that targets mTOR,45 significantly promoted the BER-reduced cell viability. Moreover, flow cytometry analysis suggested that RA pretreatment enhanced BER-induced apoptosis, while 3-MA reduced BER-induced cell death in ATC cells. These findings indicated that BER‐induced apoptosis is reduced or increased after the inhibition or promotion of autophagy by pharmaceutical tools of autophagy. The present findings also revealed that BER induces apoptosis and autophagy in ATC cells. Therefore, the relationship between apoptosis and autophagy may be mutually regulated with treatment to achieve anticancer effects. Additionally, both autophagy and apoptosis may be triggered by common upstream signals, resulting in the activation of combined autophagy and apoptosis.
Because the PI3K/AKT signaling pathway plays a fundamental role in thyroid carcinogenesis and progression, it is a potential therapeutic target in ATC treatment.21,46 Previous studies have demonstrated that inactivation of the PI3K/AKT/mTOR signaling pathway is involved in autophagy induction.19,20 Thus, the PI3K/AKT/mTOR pathway may be associated with both apoptosis and autophagy, and it may play a crucial role in BER-induced apoptosis and autophagy in ATC cells. Here, Western blot analysis indicated that BER significantly blocked the activation of the PI3K and mTOR proteins as evidenced by the decreased phosphorylation of these proteins but increased phosphorylation of AKT. Interestingly, our results were consistent with previous studies demonstrating that BER treatment activates the AKT signaling pathways in chondrosarcoma and HepG2 cells, resulting in apoptosis.47,48 Furthermore, blocking the PI3K/AKT/mTOR pathway by WOR, MK2206, and RA enhanced the BER-induced cell growth inhibition. Therefore, we further confirmed that BER represses ATC cell growth through the PI3K/AKT/mTOR pathway. Importantly, in the present study, we simultaneously investigated the roles of PI3K/AKT/mTOR signaling in BER-induced autophagy and apoptosis, and we found that inhibition of PI3K and mTOR significantly increased BER-induced cytostatic autophagy and apoptosis, leading to significant inhibition of cell proliferation, while SC79 (an AKT activator) elicited the opposite effect. Therefore, these results indicated that Akt activation played a negative role on BER-induced cytotoxicity under this situation. The functional roles of Akt signaling in the modulation of BER-induced cancer cell death have been controversially discussed. For example, BER reduces the viability of gastric cancer cell through inhibiting the activity of Akt,16 whereas BER induces apoptosis in human hepatoma cells by activating Akt.47 This may be because the effects were dependent on cell types and ligands. Taken together, the results suggested that PI3K/AKT/mTOR signaling is involved in BER-induced autophagy and apoptosis.
Because the ability of BER to produce free radicals has been reported in various cancer cell lines,33,34 we investigated the ability of BER to alter intracellular redox potential in ATC cells. In the present study, BER induced a significant increase in ROS generation, which was clearly inhibited by an ROS scavenger (NAC). In addition, a decreased level of ROS through pretreatment with NAC reversed the BER-induced cell viability decrease, apoptosis, and autophagy. These findings demonstrated that BER induces the generation of ROS, which contributes to cell death via apoptosis and/or autophagy.
DOX has been approved by the American Thyroid Association (ATA) guidelines as a conventional single agent in ATC treatment.49 However, the antitumor activities of combining DOX with BER in ATC are unknown. Thus, we tested the synergistic anticancer effects of DOX and BER in combination on ATC cancer cell lines. The combination therapy of DOX and BER significantly inhibited ATC cell proliferation and induced cell apoptosis compared to DOX or BER monotherapy. In addition, BER combined with DOX exhibited a synergistic inhibitory effect in ATC cells with CI values less than one, indicating that these two compounds in combination markedly exert synergistic growth inhibitory effects on ATC cells. Furthermore, concomitant treatment with DOX and BER enhanced cell autophagy by enhancing the expression of p62 and LC3-II. These data demonstrated that cotreatment with DOX and BER markedly exerts a synergistic growth inhibitory effect on ATC cells by triggering apoptosis and autophagic cell death. Therefore, cotreatment with DOX and BER significantly inhibits cell growth and may be a promising strategy for ATC chemoprevention. The fate of tumor cells is not determined by only one pathway and BER exerted its anti-cancer role through modulating different cell signaling pathways.50 In gastric cancer cells, BER may also cause autophagic cell death through modulation of the AKT signaling pathways.51 Another study based on the BER-induced apoptosis of melanoma A375 cells is associated with the AMPK activation.52 In addition, BER suppressed human keratinocytes cells’ growth through the JAK-STAT3 signaling pathway.53 Therefore, it is the direction of our future research to explore and investigate whether these signaling pathways is involved in the mechanism of anti-cancer by BBR treatment. Owing to time constraints, this study is only a preliminary in vitro study. We will continue to study in more cell lines and in depth in future research.
Conclusions
In conclusion, the present results demonstrated that BER exerts antitumor effects in ATC by inhibiting proliferation, promoting apoptosis, and inducing autophagy through the PI3K/AKT/mTOR signaling pathway. In this process, ROS act as a potential target of BER-mediated induction of autophagy and apoptosis. Furthermore, we demonstrated that cotreatment with DOX and BER exhibits synergistic chemopreventive effects by inducing apoptosis and autophagic cell death. Taken together, these results suggested that cotreatment with DOX and BER may be a potential therapeutic approach for ATC. Our findings may shed light on the underlying mechanisms of the anti-ATC effects of BER (Figure 8), which may serve as a promising drug for treating ATC. However, further studies exploring BER combined with DOX as a chemotherapeutic strategy for clinical translation in future.
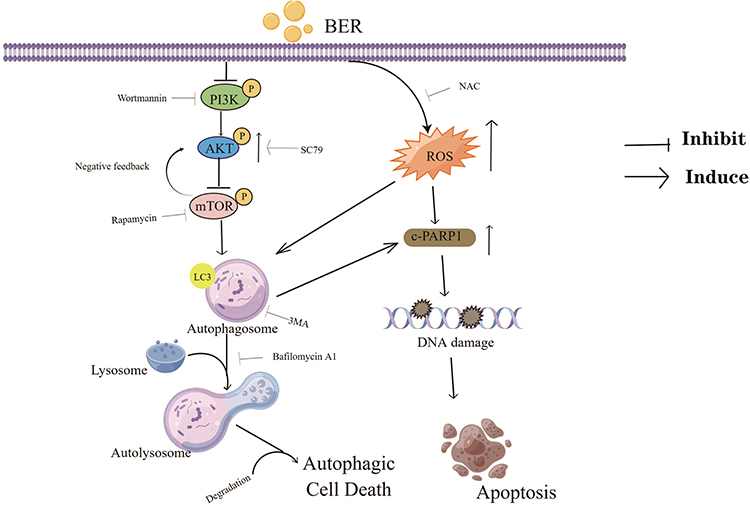 |
Figure 8 Schematic diagram illustrating the potential pathway associated with BER-induced apoptosis and autophagy. BER promotes the downregulation of PI3K/AKT/mTOR signaling and the upregulation of ROS-mediated cell dysfunction, ultimately resulting in autophagic cell death. |
Abbreviations
ATC, anaplastic thyroid carcinoma; DOX, doxorubicin; BER, berberine; PI3K, phosphatidy linositide-3 kinase; AKT, protein kinase B; mTOR, mammalian rapamycin target protein; ROS, reactive oxygen species; 3-MA, 3-methyladenine; RA, rapamycin; Wor, wortmannin; BafA1, bafilomycin A1; NAC, N-acetylcysteine.
Data Sharing Statement
This article had included all data.
Author Contributions
Chen Xiong and Yan‐Zhao Wu designed and conceptualized this article. Xiang-Zhe Shi, Sheng Zhao, Yan Wang and Meng-Yao Wang carry out experiments and analyzed data. Xiang-Zhe Shi and Chen Xiong wrote the manuscript and drew all the figures. Su‐Wen Su and Yan‐Zhao Wu helped to revise the manuscript. All authors read and approved the final manuscript.
All authors made substantial contributions to conception and design, Chen Xiong and Yan‐Zhao Wu acquisited of data, analysis and interpretation of data; Xiang-Zhe Shi, Sheng Zhao, Yan Wang and Meng-Yao Wang took part in drafting the article and revising it critically for important intellectual content; All authors agreed to submit to the current journal; All authors gave final approval of the version to be published and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Natural Science Foundation of Hebei Province (H2020206273 and H2021206012).
Disclosure
There is no conflict of interest in this article.
References
1. Chen W, Zheng R, Zhang S, et al. Cancer incidence and mortality in China in 2013: an analysis based on urbanization level. Chin J Cancer Res. 2017;29(1):1–10.
2. Deeken-Draisey A, Yang GY, Gao J, Alexiev BA. Anaplastic thyroid carcinoma: an epidemiologic, histologic, immunohistochemical, and molecular single-institution study. Hum Pathol. 2018;82:140–148.
3. Kim JH, Leeper RD. Treatment of anaplastic giant and spindle cell carcinoma of the thyroid gland with combination Adriamycin and radiation therapy. A new approach. Cancer. 1983;52(6):954–957.
4. Ranganath R, Shah MA, Shah AR. Anaplastic thyroid cancer, Current opinion in endocrinology. Diabetes Obes. 2015;22(5):387–391.
5. Liu C, Yang S, Wang K, et al. Alkaloids from traditional Chinese medicine against hepatocellular carcinoma. Biomed Pharmacother. 2019;120:109543.
6. Qi F, Zhao L, Zhou A, et al. The advantages of using traditional Chinese medicine as an adjunctive therapy in the whole course of cancer treatment instead of only terminal stage of cancer. Biosci Trends. 2015;9:16–34.
7. Tan HL, Chan KG, Pusparajah P, et al. Rhizoma coptidis: a potential cardiovascular protective agent. FronPharmacol. 2016;7:362.
8. Tabeshpour J, Imenshahidi M, Hosseinzadeh H. A review of the effects of Berberis vulgaris and its major component, berberine, in metabolic syndrome. Iran J Basic Med Sci. 2017;20(5):557–568.
9. Zou K, Li Z, Zhang Y, et al. Advances in the study of berberine and its derivatives: a focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol Sin. 2017;38(2):157–167.
10. Gong C, Hu X, Xu Y, et al. Berberine inhibits proliferation and migration of colorectal cancer cells by downregulation of GRP78. Anticancer Drugs. 2020;31:141–149.
11. Li D, Zhang Y, Liu K, et al. Berberine inhibits colitis-associated tumorigenesis via suppressing inflammatory responses and the consequent EGFR signaling-involved tumor cell growth. Lab Invest. 2017;97(1):1343–1353.
12. Li L, Wang X, Sharvan R, Gao J, Qu S. Berberine could inhibit thyroid carcinoma cells by inducing mitochondrial apoptosis, G0/G1 cell cycle arrest and suppressing migration via PI3K-AKT and MAPK signaling pathways. Biomed Pharmacother. 2017;95:1225–1231.
13. Park KS, Kim JB, Bae J, et al. Berberine inhibited the growth of thyroid cancer cell lines 8505C and TPC1. Yonsei Med J. 2012;53(2):346–351.
14. Levy JMM, Towers CG, Thorburn A. Targeting autophagy in cancer. Nat Rev Cancer. 2017;17(9):528–542.
15. Singh SS, Vats S, Chia AY, et al. Dual role of autophagy in hallmarks of cancer. Oncogene. 2018;37(9):1142–1158.
16. Zhang Q, Wang X, Cao S, et al. Berberine represses human gastric cancer cell growth in vitro and in vivo by inducing cytostatic autophagy via inhibition of MAPK/mTOR/p70S6K and Akt signaling pathways. Biomed Pharmacother. 2020;128:110245.
17. Liu J, Liu P, Xu T, et al. Berberine induces autophagic cell death in acute lymphoblastic leukemia by inactivating AKT/mTORC1 signaling. Drug Des Devel Ther. 2020;14:1813–1823.
18. Wang Y, Liu Y, Du X, Ma H, Yao J. Berberine reverses doxorubicin resistance by inhibiting autophagy through the PTEN/Akt/mTOR signaling pathway in breast cancer. Onco Targets Ther. 2020;13:1909–1919.
19. Wang H, Dong R, Fan WW, Zheng XC, Li AM, Wang WD. Timosaponin A-III induces autophagy of T-cell acute lymphoblastic leukemia Jurkat cells via inhibition of the PI3K/Akt/mTOR pathway. Oncol Rep. 2019;41(5):2937–2944.
20. Fan XJ, Wang Y, Wang L, Zhu M. Salidroside induces apoptosis and autophagy in human colorectal cancer cells through inhibition of PI3K/Akt/mTOR pathway. Oncol Rep. 2016;36(6):3559–3567.
21. Saji M, Ringel MD. The PI3K-Akt-mTOR pathway in initiation and progression of thyroid tumors. Mol Cell Endocrinol. 2010;321(1):20–28.
22. Petrulea MS, Plantinga TS, Smit JW, Georgescu CE, Netea-Maier RT. PI3K/Akt/mTOR: a promising therapeutic target for non-medullary thyroid carcinoma. Cancer Treat Rev. 2015;41(8):707–713.
23. Haddad RI, Lydiatt WM, Ball DW, et al. Anaplastic thyroid carcinoma, version 2.2015. J Natl Compr Can Netw. 2015;13(9):1140–1150.
24. Mittal A, Tabasum S, Singh RP. Berberine in combination with doxorubicin suppresses growth of murine melanoma B16F10 cells in culture and xenograft. Phytomedicine. 2014;21(3):340–347.
25. Tong N, Zhang J, Chen Y, et al. Berberine sensitizes multiple human cancer cells to the anticancer effects of doxorubicin in vitro. Oncol Lett. 2012;3(6):1263–1267.
26. Xiong C, Wu YZ, Zhang Y, et al. Protective effect of berberine on acute cardiomyopathy associated with doxorubicin treatment. Oncol Lett. 2018;15(4):5721–5729.
27. Wu YZ, Zhang L, Wu ZX, Shan TT, Xiong C. Berberine ameliorates doxorubicin-induced cardiotoxicity via a SIRT1/p66Shc-mediated pathway. Oxid Med Cell Longev. 2019;2019:2150394.
28. Cohen GM. Caspases: the executioners of apoptosis. Biochem J. 1997;326(Pt 1):1–16.
29. Mohammadinejad R, Ahmadi Z, Tavakol S, Ashrafizadeh M. Berberine as a potential autophagy modulator. J Cell Physiol. 2019;2019:1.
30. Klionsky DJ, Abdel-Aziz AK, Abdelfatah S, et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition) 1. Autophagy. 2021;17(1):1–382.
31. Pankiv S, Clausen TH, Lamark T, et al. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J Biol Chem. 2007;282(33):24131–24145.
32. Wan X, Harkavy B, Shen N, Grohar P, Helman LJ. Rapamycin induces feedback activation of Akt signaling through an IGF-1R-dependent mechanism. Oncogene. 2007;26:1932–1940.
33. Fang J, Huang X, Yang Y, Wang X, Liang X, Liu J. Berberine-photodynamic induced apoptosis by activating endoplasmic reticulum stress-autophagy pathway involving CHOP in human malignant melanoma cells. Biochem Biophys Res Commun. 2021;552:183–190.
34. Kim SY, Hwangbo H, Kim MY, et al. Coptisine induces autophagic cell death through down-regulation of PI3K/Akt/mTOR signaling pathway and up-regulation of ROS-mediated mitochondrial dysfunction in hepatocellular carcinoma Hep3B cells. Arch Biochem Biophys. 2021;697:108688.
35. Liu JZ, Hu YL, Feng Y, et al. BDH2 triggers ROS-induced cell death and autophagy by promoting Nrf2 ubiquitination in gastric cancer. J Exp Clin Cancer Res. 2020;39(1):123.
36. Wen C, Wang H, Wu X, et al. ROS-mediated inactivation of the PI3K/AKT pathway is involved in the antigastric cancer effects of thioredoxin reductase-1 inhibitor chaetocin. Cell Death Dis. 2019;10(11):809.
37. Chou TC, Talalay P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv Enzyme Regul. 1984;22:27–55.
38. Liu D, Meng X, Wu D, Qiu Z, Luo H. Anaturalisoquinolinealkaloidwith antitumor activity: studies of the biological activities of berberine. FrontPharmacol. 2019;10:9.
39. Gao X, Wang J, Li M, et al. Berberine attenuates XRCC1-mediated base excision repair and sensitizes breast cancer cells to the chemotherapeutic drugs. J Cell Mol Med. 2019;23(10):6797–6804.
40. Palmieri A, Iapichino A, Cura F, et al. Pre-treatment with berberine enhances effect of 5-fluorouracil and cisplatin in HEP2 laryngeal cancer cell line. J Biol Regul Homeost Agents. 2018;32(2 Suppl. 1):167–177.
41. Kumar A, Chopra K, Mukherjee M, Pottabathini R, Dhull DK. Current knowledge and pharmacological profile of berberine: an update. Eur J Pharmacol. 2015;761:288–297.
42. Wang K, Zhang C, Bao J, et al. Synergistic chemopreventive effects of curcumin and berberine on human breast cancer cells through induction of apoptosis and autophagic cell death. Sci Rep. 2016;6:26064.
43. Meng M, Geng S, Du Z, et al. Berberine and cinnamaldehyde together prevent lung carcinogenesis. Oncotarget. 2017;8(44):76385–76397.
44. Moscat J, Karin M, Diaz-Meco MT. p62 in cancer: signaling adaptor beyond autophagy. Cell. 2016;167(3):606–609.
45. Pan T, Rawal P, Wu Y, Xie W, Jankovic J, Le W. Rapamycin protects against rotenone-induced apoptosis through autophagy induction. Neuroscience. 2009;164(2):541–551.
46. Xing M. Genetic alterations in the phosphatidylinositol-3 kinase/Akt pathway in thyroid cancer. Thyroid. 2010;20(7):697–706.
47. Hyun MS, Hur JM, Mun YJ, Kim D, Woo WH. BBR induces apoptosis in HepG2 cell through an Akt-ASK1-ROS-p38MAPKs-linked cascade. J Cell Biochem. 2010;109(2):329–338.
48. Eo SH, Kim JH, Kim SJ. Induction of G(2)/M arrest by berberine via activation of PI3K/Akt and p38 in human chondrosarcoma cell line. Oncol Res. 2014;22(3):147–157.
49. Smallridge RC, Ain KB, Asa SL, et al. American thyroid association guidelines for management of patients with anaplastic thyroid cancer. Thyroid. 2012;22(11):1104–1139.
50. Almatroodi SA, Alsahli MA, Rahmani AH. Berberine: an important emphasis on its anticancer effects through modulation of various cell signaling pathways. Molecules. 2022;27:5889.
51. Yi T, Zhuang L, Song G, Zhang B, Li G, Hu T. Akt signaling is associated with the berberine-induced apoptosis of human gastric cancer cells. Nutr Cancer. 2015;67:523–531.
52. Kim HS, Kim MJ, Kim EJ, Yang Y, Lee MS, Lim JS. Berberine-induced AMPK activation inhibits the metastatic potential of melanoma cells via reduction of ERK activity and COX-2 protein expression. Biochem Pharmacol. 2012;83:385–394.
53. Sun S, Zhang X, Xu M, et al. Berberine downregulates CDC6and inhibits proliferation via targeting JAK-STAT3 signaling in keratinocytes. Cell Death Dis. 2019;10:274.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.