Back to Journals » Infection and Drug Resistance » Volume 12
Antimicrobial Resistance And Molecular Characteristics Among Neisseria gonorrhoeae Clinical Isolates In A Chinese Tertiary Hospital
Authors Zheng Z ![]() , Liu L, Shen X, Yu J, Chen L, Zhan L, Chen H, Lin C, Jiang Y, Xia H, Wang L, Yu F
, Liu L, Shen X, Yu J, Chen L, Zhan L, Chen H, Lin C, Jiang Y, Xia H, Wang L, Yu F
Received 27 June 2019
Accepted for publication 25 September 2019
Published 23 October 2019 Volume 2019:12 Pages 3301—3309
DOI https://doi.org/10.2147/IDR.S221109
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Zhou Zheng,1 Li Liu,1 Xiaofei Shen,1 Jingyi Yu,1 Lijiang Chen,1 Lingling Zhan,1 Han Chen,1 Chunchan Lin,1 Ye Jiang,1 Hong Xia,1 Liangxing Wang,2 Fangyou Yu1,3
1Department of Clinical Laboratory, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang, People’s Republic of China; 2Department of Respiratory Medicine, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou 325000, People’s Republic of China; 3Department of Laboratory Medicine, Shanghai Pulmonary Hospital, Tongji University School of Medicine, Shanghai 200443, People’s Republic of China
Correspondence: Fangyou Yu
Department of Clinical Laboratory, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang, People’s Republic of China
Email [email protected]
Liangxing Wang
Department of Respiratory Medicine, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou 325000, People’s Republic of China
Email [email protected]
Purpose: The resistance of N. gonorrhoeae to antimicrobial agents has been increasing year by year due to the overuse of antibiotics. The primary aims of the present study were to investigate the molecular characteristics of the clinical isolates of Neisseria gonorrhoeae and the resistance to azithromycin in a Chinese tertiary hospital.
Methods: From January 2014 to May 2017, a total of 55 clinical isolates of N. gonorrhoeae were collected. Genes associated with azithromycin resistance (AZM-R), including mutations in 23S rRNA alleles, the mtrR promoter and coding regions, and rplD and rplV were evaluated by PCR and DNA sequencing. All clinical isolates were subjected to N. gonorrhoeae multiantigen sequence typing (NG-MAST), while the AZM-R isolates were further characterized by multilocus sequence typing (MLST).
Results: The AZM-R rate in this study was 23.64% (13/55), and a single (A)-nucleotide deletion mutation in the mtrR promoter region, a G45D mutation in the mtrR coding region, a point mutation in rplD, and an A2047G mutation in 23S rRNA alleles were detected in 13, 4, 3 and 4 isolates, respectively; no mutations were found in rplV. There was no significant difference in the mtrR coding region mutation rate between the azithromycin-sensitive and AZM-R groups (P > 0.05); however, there was a significant difference in the mutation rate of the mtrR promoter region (P < 0.05). Among the 55 isolates studied, 43 distinct NG-MAST were determined, while the AZM-R isolates were allocated into 10 distinct MLST/NG-MAST combinations. All three isolates with high-level AZM-R belonged to the sequence types (STs) NG-MAST ST1866 and MLST ST10899.
Conclusion: N. gonorrhoeae clinical isolates from Wenzhou, eastern China, showed considerable genetic diversity. Measures should be implemented to monitor the spread of the NG-MAST ST1866 and MLST ST10899 N. gonorrhoeae clones, which exhibit high-level AZM-R in eastern China.
Keywords: Neisseria gonorrhoeae, azithromycin, antimicrobial resistance, NG-MAST, MLST
Introduction
Gonorrhea is the second most common sexually transmitted bacterial infection in the world. According to a recent estimation of WHO, the number of gonococcal infections occurred worldwide reached 78.3 million cases in the year 2012.1 Antimicrobial resistance (AMR) of N. gonorrhoeae has become a major public health problem worldwide. In recent years, azithromycin has been increasingly used for the treatment of gonorrhea, and the resistance rate of N. gonorrhoeae to azithromycin has risen alongside this elevated use.2,3 In many countries, the sporadic development of high-level azithromycin resistance (AZM-R) and numerous clinical failures have led to great concern and threaten the long-term efficacy of the currently recommended approach for treatment of gonorrhea.4–6
Multidrug resistance of N. gonorrhoeae is related to the overexpression of efflux pumps. The most important efflux mechanism is the MtrC-MtrD-MtrE system, encoded by the mtr operon, in which mtrR is a regulatory gene and mtrCDE are the structural genes.7 Another efflux pump, encoded by the mef gene, was originally identified in some of the gram-positive organisms and has since also been discovered in clinical strains of N. gonorrhoeae.8
The ribosomal proteins, L4 (encoded by rplD) and L22 (encoded by rplV), bind to domain I of 23S rRNA and act as channels, through which macrolide antibiotics enter the ribosome. Point mutations in rplD and rplV, conferring resistance to macrolides, have been described in Escherichia coli,9 and in rplD for Streptococcus pneumoniae;10 however, such mutations are rarely detected in N. gonorrhoeae.11 In contrast, 23S rRNA point mutations have been described, including C2611T (numbering refers to the E. coli genome), conferring low-to-moderate levels of AZM-R (minimum inhibitory concentrations (MICs) = 2 to 32 μg/mL) or A2059G (E. coli numbering), conferring high-level AZM-R (MICs ≥ 256 μg/mL).12
As a commonly used molecular typing method, N. gonorrhoeae multiantigen sequence typing (NG-MAST) detects nucleotide sequence variations in both porB and tbpB and determines the sequence types (STs) of isolates.13 A single N. gonorrhoeae can rapidly diversify into multiple NG-MAST STs by the introduction of mutations or re-combinational replacement events.14 Thus, the evaluation of NG-MAST STs is useful for short-term epidemiology. Over longer periods of time, rapid diversification can conceal the fact that several NG-MAST STs may be minor variants of the same N. gonorrhoeae strain.15
Multilocus sequence typing (MLST) is used to characterize isolates by amplifying sequences from seven housekeeping loci and is commonly used to track the spread of N. gonorrhoeae strains, where indexed genetic variation accumulates over longer periods of time.13
Little is known for types of AZM-R gonococci currently circulating in China until 2013; hence, levels of AZM-R and the molecular characteristics of AZM-R N. gonorrhoeae remain unclear.6,11,16,17 In the absence of new antimicrobial agents for the treatment of gonorrhea, the classification and evaluation of the dynamics of AZM-R N. gonorrhoeae strains on a regional and national basis are important for successful review of treatment recommendations. The resistance levels of N. gonorrhoeae to azithromycin in Wenzhou, eastern China, are unclear. The purpose of this study was to investigate the prevalence and genetic characteristics of N. gonorrhoeae from a teaching hospital in Wenzhou.
Materials And Methods
Bacterial Strains
From January 2014 to May 2017, a total of 55 nonrepetitive N. gonorrhoeae isolates were selected from 189 strains by simple random sampling in the First Affiliated Hospital of Wenzhou Medical University, of which 11 were from 2014, 10 from 2015, 17 from 2016, and 17 from 2017, where more than one sample was obtained from the same patient and only the first isolate was retained. All strains were identified by Gram-staining, oxidase reaction, and sugar fermentation tests and further confirmed as N. gonorrhoeae using a fully automatic rapid microbial mass spectrometry detection system (VITEK MS from bioMérieux, SA, France).
Antimicrobial Susceptibility Testing
Susceptibility of N. gonorrhoeae to antimicrobials, including penicillin, tetracycline, ciprofloxacin, spectinomycin, ceftriaxone, and azithromycin, was determined using the E-test method (Wenzhou Kangtai Biological Technology Co., Ltd.). E-test results were interpreted based on the Clinical and Laboratory Standards Institute (CLSI 2019) guidelines criteria for all antibiotics.18 A single N. gonorrhoeae isolate was streaked onto Thayer-Martin (T-M) selective medium for separation and purification. Inoculated plates were incubated at 36°C in 5% CO2 for 24–48 hrs. Then, bacteria at 0.5 M turbidity were smeared on six 9-cm diameter GC plates three times uniformly and densely and incubated for 24 to 48 hrs at 35°C in a 5% CO2 atmosphere with six E-test strips to determine MIC values. All strains were preserved in glycerin broth and stored at −80°C until used. The quality control strain of the antibiotic susceptibility test is N. gonorrhoeae ATCC 49226, which is provided by the Clinical Laboratory of the Ministry of Health of the People’s Republic of China and kept by the laboratory. The strain is stored in the glycerol broth liquid medium.
Extraction Of Bacterial Genomic DNA
Genomic DNA was extracted by boiling. Briefly, 3–5 N. gonorrhoeae colonies were suspended in 200 μL of sterile distilled water and boiled for 10 mins at 100°C, then centrifuged at 13,800 g for 15 mins at 4°C. Finally, supernatants were removed and stored at −20°C for further use.
PCR And DNA Sequencing Of Genes Associated With Azithromycin Resistance
In an attempt to identify point mutations, sequences of genes related to AZM-R were determined, including 23S rRNA alleles, rplD and rplV (encoding ribosomal proteins L4 and L22, respectively),19 and the mtrR promoter and coding regions. Primers (listed in Table 1) for mtrR were designed using primer premier 5.0, while those for rplD and rplV have been published previously.20
 |
Table 1 The Resistance Gene Primer Sequences And Target Fragment Sizes |
PCR conditions for the mtrR promoter region were 10 mins at 94°C for predenaturation, followed by 30 cycles of 1 min at 94°C for denaturation, 1 min at 50°C for annealing, and extension at 72°C for 1 min. PCR conditions for the mtrR coding region were 10 mins at 94°C, then 30 cycles of 1 min at 94°C, 45 s at 48°C, and 1 min at 72°C. PCR conditions for rplD were 4 mins at 94°C, then 30 cycles of 30 s at 94°C, 30 s at 60°C, and 45 s at 72°C. Conditions for amplification of rplV were 4 mins at 94°C, then 30 cycles of 30 s at 94°C, 30 s at 55°C, and 1 min at 72°C.
A two-step PCR method was developed to identify mutations within domain V of each of the four copies of the 23S rRNA gene. The first step used the Upstream-F primer, paired with a primer specific for each of the 23S rRNA alleles, as previously described (Table 1).21 PCR conditions were 30 cycles of 1 min at 94°C, 1.5 mins at 68°C (for alleles 1 and 4) or 66°C (for alleles 2 and 3), and 2.5 mins at 72°C. The products obtained were used as a template in a second PCR using the Upstream-F and Downstream-R2 primers to amplify the four copies of the 23S rRNA gene for sequence determination. For the second PCR step, the conditions were 35 cycles of: 1 min at 94°C, 1 min at 59°C, and 1 min at 72°C.21
DNA samples amplified by PCR were subjected to electrophoresis at 110 V for 30 mins. Then, all amplified products were photographed using a gel imager and confirmed to be single bands. Next, PCR products were purified using magnetic beads (by Beijing Qingke New Industry Biotechnology Co., Ltd. Hangzhou Branch) and directly sequenced using first-generation sequencing technology. DNA sequences were aligned using BLAST and GenBank programs (http://www.ncbi.nlm.nih.gov/blast/) to identify gene mutations.
N. gonorrhoeae Multiantigen Sequence Typing (NG-MAST) And Multi-Locus Sequence Typing (MLST)
All N. gonorrhoeae isolates were subjected to molecular epidemiologic analysis using NG-MAST, which assigns sequence types (STs) based on a combination of two highly variable alleles (porB and tbpB). Alignments of the porB and tbpB nucleotide sequences were modified using EditSeq software (LaserGene software version 7.1; DNAStar Corp) before submission to the NG-MAST website (www.ng-mast.net) for NG-MAST STs. MEGA 7.0 software, with the maximum-likelihood method, was used to create N. gonorrhoeae phylogenetic trees by concatenating porB and tbpB alleles.11 AZM-R were further typed using MLST, for which seven housekeeping genes (abcZ, adk, aroE, fumC, gdh, pdh, and pgm) were analyzed and the sequencing results submitted to the database (http://pubmlst.org/neisseria/) to obtain allele numbers. Strain sequences were then obtained based on allele numbers (STs).
Results
Patient Data
Among 55 patients with gonorrhea, there were 52 males and 3 females whose age distribution range was 16–78 years (mean ± SD, 32.0 ± 11.2 years); 65.5% (36/55) were married and the main diagnosis is urethritis.
Antimicrobial Susceptibility
Among the 55 N. gonorrhoeae strains tested, 52 (94.5%), 19 (34.5%), 49 (90.1%), 0 (0.0%), 0 (0.0%), and 13 (23.6%) were resistant to penicillin, tetracycline, ciprofloxacin, spectinomycin, ceftriaxone, and azithromycin, respectively. MIC values for azithromycin ranged from 0.064 to >256 μg/mL, with an MIC50 of 0.5 μg/mL and an MIC90 of 1.0 μg/mL. The rates of resistance to azithromycin for the years from 2014 to 2017 were 18.2% (2/11), 10.0% (1/10), 29.4% (5/17), and 29.4% (5/17), respectively. There were 21 multidrug resistant (MDR) strains (strains with resistance to three or more types of antimicrobial agent) among the 55 strains of N. gonorrhoeae, and all 13 AZM-R strains were multidrug resistant. Of the 13 AZM-R strains, all were resistant to penicillin and ciprofloxacin, five were resistant to tetracycline, and the remaining eight exhibited intermediate resistance to tetracycline. Among the 13 AZM-R isolates, eight isolates with MIC values of 1 μg/mL were classified as exhibiting low-level resistance, two with MIC values of 4 μg/mL showed moderate-level resistance, and three with MIC values of>256 μg/mL exhibited high-level resistance.
Detection Of Mutations In Genes Associated With Azithromycin Resistance
The following mutations were detected in 13, 4, and 4 of the 13 AZM-R isolates, respectively: a single-nucleotide (A) deletion in the mtrR promoter region, G45D in the mtrR coding region, and A2047G (N. gonorrhoeae numbering, GenBank accession number: X67293.1) in 23S rRNA alleles (Table 2). Four allelic mutations in 23S rRNA were detected in two of three (66.7%) N. gonorrhoeae isolates exhibiting high-level AZM-R. Point mutations in rplD were identified in three isolates (G68D, G70D, and a combination of T69I and G70S). No mutations were detected in rplV.
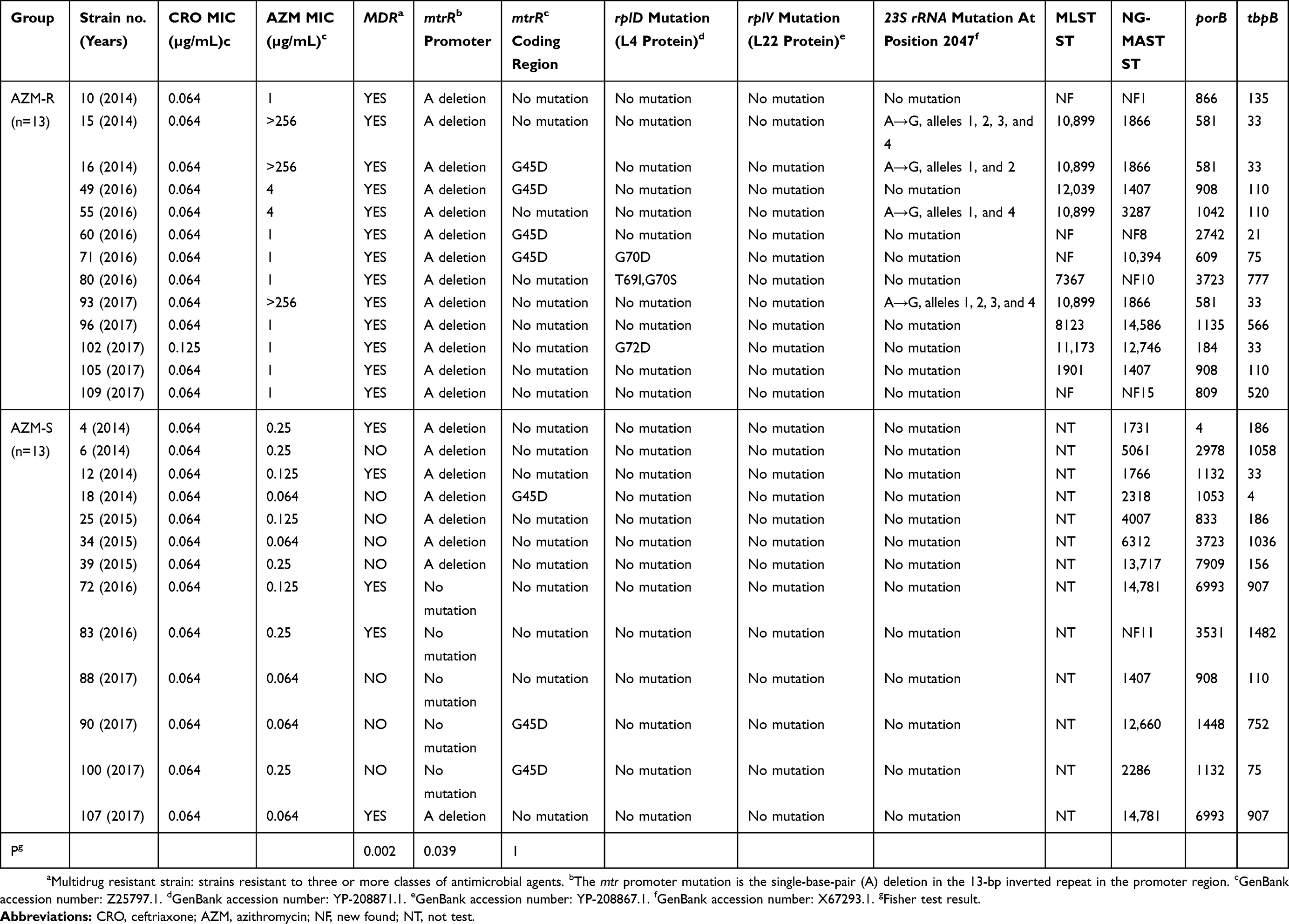 |
Table 2 Characterization And Molecular Typing Results Of Clinical Isolates Of Neisseria gonorrhoeae |
Thirteen strains were randomly selected from the 42 azithromycin-sensitive (AZM-S) strains as a control group. Mutations detected in these controls included: a single-nucleotide (A) deletion in the mtrR promoter region and G45D in the mtrR coding region in eight and three isolates, respectively (Table 2). No mutations were detected in the rplD and rplV genes or the 23S rRNA alleles in the AZM-S group.
Comparison between the AZM-R and AZM-S groups indicated that the mutation rate in the mtrR coding region was not significantly different between them (P > 0.05), whereas there was a significant difference in the rate of mutation of the mtrR promoter region between the two groups (P < 0.05).
Molecular Epidemiologic Typing
The 55 N. gonorrhoeae clinical isolates were typed by NG-MAST analysis and 43 different sequence types (STs) were identified, with 21 (48.8%, 21/43) represented by only a single isolate. ST14781 was the most prevalent ST, accounting for four isolates, followed by ST1766 (three isolates), ST1866 (three isolates), ST5061 (three isolates), ST1407 (two isolates), ST4007 (two isolates), and ST4846 (two isolates). Fifteen of the STs (34.9%,15/43) identified in the present study have not been reported previously in the NG-MAST database. Among the 13 AZM-R isolates, ST1866 was the most prevalent (23.1%, 3/13), followed by ST1407 (15.4%, 2/13). All three NG-MAST ST1866 isolates (which were not epidemiologically linked to one another) exhibited high-level AZM-R.
A total of 10 different STs, including four novel STs determined by MLST, were identified among the 13 AZM-R isolates (Table 2), of which ST10,899 accounted for four isolates (30.8%, 4/13). Three NG-MAST ST1866 isolates with high-level AZM-R belonged to MLST ST10899, indicating that they originated from the same clone.
Based on phylogenetic analysis, four large clusters (including ≥6 isolates) were identified, which excluded 15 new-found types (Figure 1). Cluster A isolates included six different STs, half of them with tbpB 75 (ST2286, ST10394, and STNF7 isolates). Cluster B isolates included five different STs with tbpB 186 (ST2422) or tbpB 33 (ST1766, ST1866, ST12746, and STNF3). Cluster C isolates were represented by nine different STs (ST1407 and ST3287 isolates with tbpB 110 and ST1731, ST4007, ST4056, and STNF2 isolates with tbpB 186). Finally, AZM-S cluster D contained five different STs, including ST505 and STNF12, both with porB 90; ST1660, ST4846, and STNF13 with porB 1448.
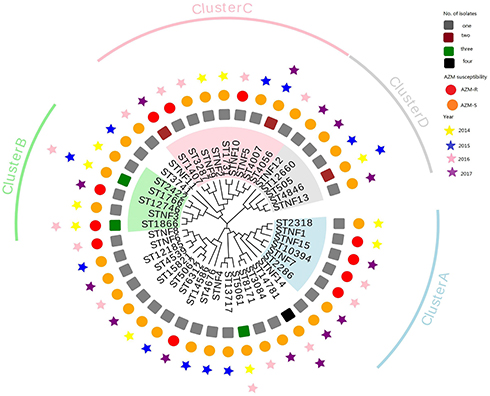 |
Figure 1 Phylogenetic tree constructed using MEGA7.0 for NG-MAST STs of 55 N. gonorrhoeae isolates from Wenzhou, China, 2014 to 2017. |
Discussion
Gonorrhea is the second most common sexually transmitted infection (STI) in the world and remains a major public health problem in China. From 2015 to 2016, the number of gonorrhea cases reported in China increased by 14.7% (100,245 to 115,024).38 In recent years, due to the overuse of antimicrobial agents (in China, it is common for doctors in small clinics to use antibiotics at will) and the evolution of bacteria, the resistance of N. gonorrhoeae to antibiotics has increased annually,21 bringing enormous challenges for the clinical treatment of N. gonorrhoeae infections. The resistance rates of N. gonorrhoeae to penicillin and ciprofloxacin in the present study (>90%) were similar to those reported previously,11 indicating that these two antibacterial drugs should not be used for the treatment of gonorrhea. Resistance to tetracycline and azithromycin was 34.5% and 23.6% in our study; therefore, their use is not at all advisable (based on the WHO recommendations: “in general; an antibiotic should not be used when more than 5% of strains are resistant to it”; Tapsal 2001; WHO/CDS/CSR/DRS/2001.3). All strains were sensitive to spectinomycin and ceftriaxone in our study, which are widely used in China for the treatment of gonorrhea; hence these can still be used as first-line drugs to treat gonorrhea (in China, the extended-spectrum cephalosporin ceftriaxone has been recommended as a monotherapy for gonorrhea since 200739). However, ceftriaxone-resistant N. gonorrhoeae has been detected in several studies;22–24 therefore, it is necessary to strengthen surveillance for ceftriaxone resistance.
In this study, the rate of AZM-R was 29.41% in both 2016 and 2017, similar to other recent reports from China. For example, analysis of 126 isolates from Hefei from 2014 to 2015 showed that 29% were AZM-R, including 10% exhibiting high-level
AZM-R.17 Furthermore, a similar incidence was reported among 384 isolates from Nanjing during 2013–2014, with 32% of isolates displaying AZM-R and 10% showing high-level resistance.11 Notably, the levels of resistance to azithromycin observed in earlier studies from other regions in China were lower.25,26 As two neighboring countries in Asia, the resistance rates of N. gonorrhoeae isolated from China and Japan to azithromycin are much higher than those from Europe and America.2,27–29 Antimicrobial resistant N. gonorrhoeae was first reported in East Asia, subsequently appearing in other parts of the world.30
Mutations in the promoter or coding sequence of the mtrR gene in macrolide-resistant N. gonorrhoeae strains can result in decreased expression of the MtrR repressor, thereby upregulating the MtrCDE efflux pump.31 In gonococcal strains with a single-base-pair (A) deletion in the 13-bp inverted repeat sequence of the mtrR promoter region, mtrR expression is abrogated, while levels of mtrCDE are elevated, most likely because of an increased binding affinity of RNA polymerase for mtrCDE.32 Missense mutations in the mtrR gene, for example, a G45D mutation in the helix-turn-helix motif in the MtrR repressor, can reduce binding of the repressor to the mtrCDE promoter.7 In the present study, the mutation rates of the mtrR promoter and coding regions in AZM-R isolates were 100.0% and 30.8%, respectively. Compared with the AZM-S group, the mutation rate of the mtrR promoter region in the AZM-R group was significantly higher, while there was no significant difference in the mutation rate of the mtrR coding region. This is in accordance with previously reported findings indicating that mutations in the mtrR promoter region play a more important role in the resistance of N. gonorrhoeae to azithromycin than alterations in the mtrR coding region.12 The mutations A40D, A39T, H105Y, and mtr120 reported by other investigators11,12,17,33 were not found in our study.
Previous studies reported that one or more of the four alleles of the rrl gene within domain V of 23S rRNA are associated with AZM-R, including the mutations, A2143G and C2599T (corresponding to A2059G and C2611T in E. coli, respectively).34,35 The mutation A2047G (N. gonorrhoeae numbering) in domain V of detected in the present study in four AZM-R N. gonorrhoeae, but was not found in the AZM-S group. The numbers of each 23S rRNA allele in which the A2047G mutation occurred were 4, 4, 2, and 2, respectively, and the corresponding MIC values were 256, 256, 256, and 4 μg/mL, respectively. It is worth noting that the A2047G mutation was detected in all strains with an MIC value of >256 μg/mL; therefore, we believe that having multiple alleles of A2047G is a major determinant of high-level azithromycin resistance.
Three N. gonorrhoeae isolates with MIC values for azithromycin of 1.0 μg/mL were found to harbor mutations in rplD (one isolate with G68D, one with G70D, and another with both T69I and G70S mutations). To our best knowledge, this is the first report of the mutations, G70D, T69I, and G70S, in rplD, while the G68D mutation has been described previously by Jacobsson.33 Therefore, mutations at positions 68–70 of the N. gonorrhoeae rplD gene appeared to be associated with low-to mid-level AZM resistance. The strains with rplD gene mutations belonged to clusters A and C, with NG-MAST classifications of ST10394, ST12746, and NF, indicating that N. gonorrhoeae strains with this mutation are genetically diverse.
N. gonorrhoeae typing methods, such as NG-MAST and MLST, assist in understanding of the spread of gonorrhea. In the present study, 34.9% (15/43) of STs determined by NG-MAST were newly recognized, and 48.8% (21/43) of these STs were represented by only a single isolate, indicating that these N. gonorrhoeae clinical isolates exhibit considerable genetic diversity. According to the phylogenetic tree, AZM-R strains are scattered throughout each year. Furthermore, phylogenetic analysis demonstrated that AZM-R isolates were widely divergent and did not belong to any specific clusters. This may be related to the large numbers of temporary residents in Wenzhou (according to the main data bulletin of Wenzhou City’s sixth national census in 2010, the city’s permanent population was 9,122,100, of which the number of inflows from outside the city was 2,842,200, accounting for 31.16% of the city’s permanent population), which may provide a greater opportunity for the emergence of the N. gonorrhoeae isolates with new and multiple NG-MAST STs; however, all three isolates with high-level AZM-R belonged to NG-MAST ST1866 and MLST ST10899, unlike those reported from other countries, such as Sweden (NG-MAST ST285, ST332, and ST8727);5 Wales, England, and the United States (NG-MAST ST649);36,37 and Argentina (NG-MAST ST696).4 It is worth mentioning that the high level of AZM-R N. gonorrhoeae found in this study has the same NA-MAST and MLST type as the high-level AZM-R N. gonorrhoeae found in the Liu YH report,40 which indicates that the spread of NG-MAST ST1866, MLST ST10899 N. gonorrhoeae has crossed the Taiwan Strait. Furthermore, the NG-MAST ST1866 clone has previously been described in Nanjing, Hangzhou, and Hefei,6,11,17 indicating that this clone with high-level AZM-R has spread in eastern China, which is a cause for concern. In addition, we also noted that the extensive drug-resistant (XDR) strain mentioned in the Jennison AV41 report is highly resistant to azithromycin while resistant to ceftriaxone. The XDR is the NG-MAST 16848 and MLST 12039, different from the high level of AZM-R N. gonorrhoeae found in our study. However, further analysis revealed that MLST 12039 and MLST 10899 have six identical housekeeping genes, which differ only in the pdhc allele. This may mean that high-level AZM-R N. gonorrhoeae is only one step away from the XDR strain. This must be a cause for concern.
Conclusion
Although the sample size of this study is limited, it is of great significance to study the resistance of N. gonorrhoeae to azithromycin in Wenzhou, eastern China. Measures should be implemented to monitor the spread of the NG-MAST ST1866 and MLST ST10899 N. gonorrhoeae clones, which exhibit high-level AZM-R in eastern China.
Ethics Approval
Because the N. gonorrhoeae were part of the routine hospital laboratory procedure, ethics approval was not required.
Acknowledgments
The authors specially thank professor Yu Fangyou for his guidance and support. This work was supported by Wenzhou Municipal Science and Technology Bureau, China (No. Y20170808).
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Newman L, Rowley J, Vander Hoorn S, et al. Global estimates of the prevalence and incidence of four curable sexually transmitted infections in 2012 based on systematic review and global reporting. PLoS One. 2015;10(12):e0143304. doi:10.1371/journal.pone.0143304
2. Kirkcaldy RD, Harvey A, Papp JR, et al. Neisseria gonorrhoeae antimicrobial susceptibility surveillance - the gonococcal isolate surveillance project, 27 sites, United States, 2014. MMWR Surveill Summ. 2016;65(7):1–19. doi:10.15585/mmwr.ss6507a1
3. Cole MJ, Spiteri G, Jacobsson S, et al. Overall low extended-spectrum cephalosporin resistance but high azithromycin resistance in neisseria gonorrhoeae in 24 European countries, 2015. BMC Infect Dis. 2017;17(1):617. doi:10.1186/s12879-017-2757-2
4. Galarza PG, Alcala B, Salcedo C, et al. Emergence of high level azithromycin-resistant Neisseria gonorrhoeae strain isolated in Argentina. Sex Transm Dis. 2009;36(12):787–788. doi:10.1097/OLQ.0b013e3181b61bb1
5. Unemo M, Golparian D, Hellmark B. First three Neisseria gonorrhoeae isolates with high-level resistance to azithromycin in Sweden: a threat to currently available dual-antimicrobial regimens for treatment of gonorrhea? Antimicrob Agents Chemother. 2014;58(1):624–625. doi:10.1128/AAC.02093-13
6. Ni C, Xue J, Zhang C, Zhou H, van der Veen S. High prevalence of Neisseria gonorrhoeae with high-level resistance to azithromycin in Hangzhou, China. J Antimicrob Chemother. 2016;71(8):2355–2357. doi:10.1093/jac/dkw131
7. Lucas CE, Balthazar JT, Hagman KE, Shafer WM. The MtrR repressor binds the DNA sequence between the mtrR and mtrC genes of Neisseria gonorrhoeae. J Bacteriol. 1997;179(13):4123–4128. doi:10.1128/jb.179.13.4123-4128.1997
8. Luna VA, Cousin S
9. Gregory ST, Dahlberg AE. Erythromycin resistance mutations in ribosomal proteins L22 and L4 perturb the higher order structure of 23 S ribosomal RNA. J Mol Biol. 1999;289(4):827–834. doi:10.1006/jmbi.1999.2839
10. Tait-Kamradt A, Davies T, Cronan M, et al. Mutations in 23S rRNA and ribosomal protein L4 account for resistance in pneumococcal strains selected in vitro by macrolide passage. Antimicrob Agents Chemother. 2000;44(8):2118–2125. doi:10.1128/aac.44.8.2118-2125.2000
11. Wan C, Li Y, Le WJ, et al. Increasing resistance to azithromycin in neisseria gonorrhoeae in Eastern Chinese cities: resistance mechanisms and genetic diversity among isolates from Nanjing. Antimicrob Agents Chemother. 2018;62(5). doi:10.1128/AAC.02499-17
12. Demczuk W, Martin I, Peterson S, et al. Genomic epidemiology and molecular resistance mechanisms of azithromycin-resistant neisseria gonorrhoeae in Canada from 1997 to 2014. J Clin Microbiol. 2016;54(5):1304–1313. doi:10.1128/JCM.03195-15
13. Mavroidi A, Tzelepi E, Siatravani E, et al. Analysis of emergence of quinolone-resistant gonococci in Greece by combined use of Neisseria gonorrhoeae multiantigen sequence typing and multilocus sequence typing. J Clin Microbiol. 2011;49(4):1196–1201. doi:10.1128/JCM.02233-10
14. Martin IM, Ison CA, Aanensen DM, Fenton KA, Spratt BG. Rapid sequence-based identification of gonococcal transmission clusters in a large metropolitan area. J Infect Dis. 2004;189(8):1497–1505. doi:10.1086/383047
15. Choudhury B, Risley CL, Ghani AC, et al. Identification of individuals with gonorrhoea within sexual networks: a population-based study. Lancet. 2006;368(9530):139–146. doi:10.1016/S0140-6736(06)69003-X
16. Liang JY, Cao WL, Li XD, et al. Azithromycin-resistant Neisseria gonorrhoeae isolates in Guangzhou, China (2009-2013): coevolution with decreased susceptibilities to ceftriaxone and genetic characteristics. BMC Infect Dis. 2016;16:152. doi:10.1186/s12879-016-1987-z
17. Jiang FX, Lan Q, Le WJ, Su XH. Antimicrobial susceptibility of Neisseria gonorrhoeae isolates from Hefei (2014-2015): genetic characteristics of antimicrobial resistance. BMC Infect Dis. 2017;17(1):366. doi:10.1186/s12879-017-2757-2
18. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. 29th ed.CLSI Supplement M100. Wayne (PA): CLSI; 2019.
19. Depardieu F, Courvalin P. Mutation in 23S rRNA responsible for resistance to 16-membered macrolides and streptogramins in Streptococcus pneumoniae. Antimicrob Agents Chemother. 2001;45(1):319–323. doi:10.1128/AAC.45.1.319-323.2001
20. Allen VG, Farrell DJ, Rebbapragada A, et al. Molecular analysis of antimicrobial resistance mechanisms in Neisseria gonorrhoeae isolates from Ontario, Canada. Journal. 2011;55(Issue):703–712.
21. Ng LK, Martin I, Liu G, Bryden L. Mutation in 23S rRNA associated with macrolide resistance in Neisseria gonorrhoeae. Antimicrob Agents Chemother. 2002;46(9):3020–3025. doi:10.1128/aac.46.9.3020-3025.2002
22. Shimuta K, Watanabe Y, Nakayama S, et al. Emergence and evolution of internationally disseminated cephalosporin-resistant Neisseria gonorrhoeae clones from 1995 to 2005 in Japan. BMC Infect Dis. 2015;15:378. doi:10.1186/s12879-015-1110-x
23. Yin YP, Han Y, Dai XQ, et al. Susceptibility of Neisseria gonorrhoeae to azithromycin and ceftriaxone in China: a retrospective study of national surveillance data from 2013 to 2016. PLoS Med. 2018;15(2):e1002499. doi:10.1371/journal.pmed.1002593
24. Yan J, Xue J, Chen Y, et al. Increasing prevalence of Neisseria gonorrhoeae with decreased susceptibility to ceftriaxone and resistance to azithromycin in Hangzhou, China (2015-17). J Antimicrob Chemother. 2019;74(1):29–37. doi:10.1093/jac/dky412
25. Li W, Zhu BY, Qin SQ, et al. Surveillance of antibiotic susceptibility patterns of Neisseria gonorrhoeae from 2013 to 2015 in Guangxi Province, China. Jpn J Infect Dis. 2018;71(2):148–151. doi:10.7883/yoken.JJID.2017.169
26. Zhao L, Liu A, Li R, Zhao S. Trends in antimicrobial resistance in Neisseria gonorrhoeae and molecular characteristics of N. gonorrhoeae with decreased susceptibility to ceftriaxone in Shandong, China, 2007 to 2014. Int J Antimicrob Agents. 2018;51(1):52–56. doi:10.1016/j.ijantimicag.2017.06.004
27. Lahra MM, Enriquez RP. Australian Gonococcal Surveillance Programme, 1 July to 30 September 2015. Commun Dis Intell Q Rep. 2016;40(1):E179–81.
28. Yasuda M, Hatazaki K, Ito S, et al. Antimicrobial Susceptibility of Neisseria gonorrhoeae in Japan from 2000 to 2015. Sex Transm Dis. 2017;44(3):149–153. doi:10.1097/OLQ.0000000000000556
29. Gonococcal Antimicrobial Susceptibility Surveillance in Europe-Results summary 2017. European Centre for Disease Prevention and Control(ECDC).2019.doi:10.2900/415493.
30. Centers for Disease Control and Prevention (CDC). Cephalosporin susceptibility among Neisseria gonorrhoeae isolates–united States, 2000-2010. MMWR Morb Mortal Wkly Rep. 2011;60(26):873–877.
31. Lahra MM. Annual report of the Australian gonococcal surveillance programme, 2011. Commun Dis Intell Q Rep. 2012;36(2):E166–E173.
32. Hagman KE, Shafer WM. Transcriptional control of the mtr efflux system of Neisseria gonorrhoeae. J Bacteriol. 1995;177(14):4162–4165. doi:10.1128/jb.177.14.4162-4165.1995
33. Jacobsson S, Golparian D, Cole M, et al. WGS analysis and molecular resistance mechanisms of azithromycin-resistant (MIC >2 mg/L) Neisseria gonorrhoeae isolates in Europe from 2009 to 2014. J Antimicrob Chemother. 2016;71(11):3109–3116. doi:10.1093/jac/dkw279
34. Allen VG, Seah C, Martin I, Melano RG. Azithromycin resistance is coevolving with reduced susceptibility to cephalosporins in Neisseria gonorrhoeae in Ontario, Canada. Antimicrob Agents Chemother. 2014;58(5):2528–2534. doi:10.1128/AAC.02608-13
35. Belkacem A, Jacquier H, Goubard A, et al. Molecular epidemiology and mechanisms of resistance of azithromycin-resistant Neisseria gonorrhoeae isolated in France during 2013-14. J Antimicrob Chemother. 2016;71(9):2471–2478. doi:10.1093/jac/dkw182
36. Katz AR, Komeya AY, Soge OO, et al. Neisseria gonorrhoeae with high-level resistance to azithromycin: case report of the first isolate identified in the United States. Clin Infect Dis. 2012;54(6):841–843. doi:10.1093/cid/cir929
37. Chisholm SA, Neal TJ, Alawattegama AB, et al. Emergence of high-level azithromycin resistance in Neisseria gonorrhoeae in England and Wales. J Antimicrob Chemother. 2009;64(2):353–358. doi:10.1093/jac/dkp188
38. Epidemic of infectious diseases in China, 2016. Chinese Center for Disease Control and Prevention. 2018. http://www.nhfpc.gov.cn/jkj/s3578/201702/38ca5990f8a54ddf9ca6308fec406157.shtml
39. Wang QQ, Zhang GC. Guidelines for Diagnosis and Treatment of Sexually Transmitted Diseases [In Chinese]. Shanghai: Shanghai Science and Technology Press; 2007.
40. Liu YH, Wang YH, Liao CH, Hsueh PR. Emergence and Spread of Neisseria gonorrhoeae Strains with High-level Resistance to Azithromycin in Taiwan from 2001 to 2018. Antimicrob Agents Chemother. 2019. doi:10.1128/AAC.00773-19
41. Jennison AV, Whiley D, Lahra MM, et al. Genetic relatedness of ceftriaxone-resistant and high-level azithromycin resistant Neisseria gonorrhoeae cases, United Kingdom and Australia, February to April 2018. Euro Surveill. 2019;24(8). doi:10.2807/1560-7917.ES.2019.24.8.1900118
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.