Back to Journals » Journal of Inflammation Research » Volume 14
Anti-Inflammatory Effects of Four Psilocybin-Containing Magic Mushroom Water Extracts in vitro on 15-Lipoxygenase Activity and on Lipopolysaccharide-Induced Cyclooxygenase-2 and Inflammatory Cytokines in Human U937 Macrophage Cells
Authors Nkadimeng SM ![]() , Steinmann CM, Eloff JN
, Steinmann CM, Eloff JN
Received 30 April 2021
Accepted for publication 29 June 2021
Published 5 August 2021 Volume 2021:14 Pages 3729—3738
DOI https://doi.org/10.2147/JIR.S317182
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Ning Quan
Sanah Malomile Nkadimeng,1 Christiaan ML Steinmann,2 Jacobus N Eloff1
1Phytomedicine Programme, Paraclinical Sciences Department, University of Pretoria, Onderstepoort, Pretoria, Gauteng, 0110, South Africa; 2Physiology Department, Sefako Makgatho Health Sciences University, Ga-Rankuwa, Gauteng, 0208, South Africa
Correspondence: Sanah Malomile Nkadimeng
Phytomedicine Programme, Paraclinical Sciences Department, University of Pretoria, P/Bag X04, Onderstepoort, Pretoria, Gauteng, 0110, South Africa
Tel +27 12 529 8187
Email [email protected]
Purpose: During a pathological inflammation, macrophages are activated to produce accumulation of inflammatory mediators such as induced-cyclooxygenase-2 (COX-2), 15-lipoxygenase (15-LOX) and pro-inflammatory cytokines. Pathological inflammation is a significant problem in many chronic diseases. As a result, more research into natural remedies with anti-inflammatory potential is crucial. Since ancient times, psilocybin-containing mushrooms, also known as magic mushrooms, were used for mind healing and also to advance the quality of life. However, not much is known about their anti-inflammatory potential. This study aimed at investigating the anti-inflammatory effects of four psilocybin-containing mushrooms (Panaeolus cyanescens, Psilocybe natalensis, Psilocybe cubensis and Psilocybe cubensis leucistic A+ strain) from genus Panaeolus and Psilocybe for the first time in vitro on 15-LOX activity and also on lipopolysaccharide (LPS)-induced inflammation in human U937 macrophage cells.
Methods: Mushrooms were grown and extracted with boiling hot water. Effects of the four water extracts on 15-LOX activity were determined. Confluent human U937 cells were differentiated with phorbol 12-myristate 13-acetate and treated with the hot-water extracts (25 and 50 μg/mL) 2 hours before being stimulated with 1 μg/mL LPS over 24 hours. Quercetin was used as a positive control. Control cells were differentiated but not LPS-induced nor treated. Tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, and IL-10 concentrations were measured. Levels of COX-2 and mitochondrial activity were also determined.
Results: The four water extracts had poor 15-LOX inhibition activity with IC50 > 250 μg/mL. Extracts were safe at the concentration studied and inhibited the LPS-induced production of pro-inflammatory mediators, TNF-α and IL-1β significantly and lowered IL-6 and COX-2 concentrations in treated human U937 macrophage cells. Water extracts also increased percentage viability of treated cells and levels of anti-inflammatory IL-10 non-significantly.
Conclusion: The study suggested that the hot-water extracts of the four psilocybin-containing magic mushrooms have potential anti-inflammatory effects executed by down-regulating pro-inflammatory mediators.
Keywords: Panaeolus cyanescens, Psilocybe natalensis, Psilocybe cubensis, Psilocybe cubensis A+ strain, pro-inflammatory cytokine, COX-2, 15-LOX
Introduction
Pathological inflammation is central to a number of chronic diseases including cardiovascular, cancer, arthritis, neurodegeneration, chronic depression and aging.1,2 Inflammation is a natural essential response of our innate immune system upon exposure to a foreign agent or injury, with the purpose of preventing damage and to remove the harmful agents.3 Altogether, immunity does not only differentiates between self and not-self but also between dangerous and non-dangerous agents.4 However, when inflammation is not properly regulated and is prolonged, it leads to an unhealthy state as it relates to many chronic diseases. As a result, during pathological inflammation, macrophages found in the body are activated to produce an unhealthy accumulation of inflammatory mediators.5
Cytokines are small proteins released by cells that may either act on the cells that secreted them, on neighboring cells or in some occasions on distant cells.6 There are both pro-inflammatory and anti-inflammatory cytokines and although made by many different cell populations, cytokines are predominantly produced by helper T cells and macrophages.6 In particular, pro-inflammatory cytokines are produced predominantly by activated macrophages and are involved in the up-regulation of inflammatory reactions.6 Imbalance in the creation of inflammatory cytokines such as Interleukin (IL)-6, tumor necrosis factor-α (TNF-α), IL-1 and IL-10 were found to be involved in dysfunctions in immunity and also in mediating inflammation of tissues and organ damage.7 There is also profuse evidence that pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α are involved in the process of pathological pain by directly activating nociceptive sensory neurons.6 Studies have shown that during cell injury, foreign invasion, infections and inflammation, IL-1β cytokine is released predominantly by monocytes and macrophages and also by non-immune cells, such as fibroblasts and endothelial cells.6 Furthermore, IL-1β cytokine was also found to have an increasing effect on the production of substance P and prostaglandin endoperoxide synthase 2 (PGE2) in numerous neuronal and glial cells.6 Inhibition of IL-1β receptors by use of agents such as IL-a which is a specific IL-1 receptor antagonist were found to prevent and weaken cytokine-mediated inflammatory hyperalgesia and nerve-injury induced allodynia.6 The TNF-α is also a major pro-inflammatory cytokine that acts on various signaling pathways by binding on its two cell surface receptors, TNF receptor-1 and TNF receptor-2 to regulate apoptotic pathways, NF-kB activation of inflammation, and activate stress-activated protein kinases.6 Blockage of TNF-α in some studies resulted in a total elimination of hyperalgesia which is normally observed after administration of lipopolysaccharide (LPS).6 Cytokine IL-6 has also been found to have a key role in the neuronal reaction to nerve injury whereby its inhibition in some studies led to reduced regenerative effects.6
Interleukin-10 on the other hand is considered an important anti-inflammatory modulator and was found to have a preventative role in a pathological inflammation-mediated condition.8
One of the well-known metabolic precursors are the arachidonic acids (AA) and they are involved in a number of inflammation pathways.9 Arachidonic acids are 20-carbon unsaturated fatty acids and in a resting position they are generally disseminated in a lipid bilayer membrane.10 However, various studies have shown that many external and internal factors may stimulate phospholipase A2, which upon stimulation cleaves the membrane-bound AA from the phospholipids in order to render it available for three major pathways of inflammation that include cytochrome P-450 monooxygenase, lipoxygenase (LOX) and cyclooxygenase (COX) pathways.11 Cyclooxygenase and LOX are the most studied and crucial enzymes responsible for the progression of inflammation.11
The COX pathway is initiated with the formation of hydroperoxy endoperoxide (PGG2) from AA which will result with oxygen molecules being added to the AA structure.12 Then PGG2 binds to the PGG2 specific position, where it is reduced through lipid peroxidation, and results in the production of prostaglandin H2 (PGH2), a major metabolic substrate for the prostaglandin and thromboxane associated synthases.13,14 The COX enzymes are characterized into two distinctive iso-enzymes, COX-1 which is constitutive in nature and largely expressed in different parts of the body, and COX-2 which can be easily stimulated by different endogenous and exogenous stimulus including pro-inflammatory cytokines IL-1, IL- 6, TNF-α, lipopolysaccharide (LPS) and stress.11 Once induced COX-2 can cause an excessive production of prostaglandin endoperoxide synthase 2 (PGE2) (a major metabolic product of COX-2) from PGH2 via prostaglandin E synthase (PGES). The PGE2 together with other prostaglandins lowers the threshold of pain and stimulates pain, sensitizes nerve endings in both the central and peripheral nervous systems, increases vascular permeability and henceforth creates a route for inflammatory associated disorders or diseases.11,15 As a result, inhibition of COX-2 enzyme will provide a significant improvement in the treatment of pathological inflammation.
Lipoxygenases on the other hand, are non-heme iron-containing dioxygenases that are known to catalyze the stereo-specific peroxidation of polyunsaturated fatty acids such as linoleic acid and AA to the corresponding hydroperoxy derivatives.16 Lipoxygenase enzymes are expressed in epithelial, tumor and immune cells that display a diversity of physiological functions which include skin disorders, tumorigenesis and inflammation.17 The enzymes are widely spread in animals, plant, and fungi and they are also present in the human body where they are involved in the synthesis of prostaglandins and leukotrienes and play an important role in the stimulation of inflammatory reactions.18 The LOX enzymes are associated with disease development and their inhibition is considered to be a crucial step in disease prevention.19 In particular, the role of 15-LOX has been implicated in various inflammation-related diseases.17 The pro-inflammatory role of 15-LOX and its metabolite 15(S)-HETE were demonstrated by various studies including the Namgaladze et al.20 study where LPS-induced expression of pro-inflammatory cytokines such as IL-6 and IL-12 was reduced by 15-LOX inhibition in macrophages.
Reactive oxygen species (ROS) are classified as partially reduced metabolites of oxygen that possess strong oxidizing capabilities and are also key signaling molecules that play an important role in the progression of inflammation.21 ROS are created as by-products of cellular metabolism through the electron transport chain in mitochondria and also via cytochrome P450.21 ROS act both as a signaling molecule and a mediator of inflammation and they are harmful to cells in high levels or if not properly regulated.21 They oxidize protein and lipid cellular constituents and damage the DNA, as a result causing cell injury and death.21 Chronic or prolonged ROS production is central to the progression of inflammatory disease.22 Excessive amounts of ROS are also known to cause inflammation by stimulating the release of cytokines and subsequent activation of COX and LOXs signaling.17
Numerous studies have showed that over time, prolonged inflammation may actually cause damage to the tissues around the affected region.3 At present non-steroidal anti-inflammatory drugs (NSAIDs) such as aspirin, which act by inhibiting COX-2 activation, are available to treat inflammatory diseases.5 However, studies also reported the adverse effects associated with their use which include disturbances in the upper gastrointestinal system and heart burn.5 As a result, more research into remedies with anti-inflammatory effects is crucial. Natural products with medicinal properties have been used to treat all sorts of inflammatory conditions and studies have shown that natural products have advantageous structural diversity over synthetic compounds, which thus make them a potential source of novel compounds with potent anti-inflammatory activity.11
Psilocybin-containing mushrooms, commonly known as magic mushrooms, have been used since ancient times for their mind healing properties and to improve the quality of life. The mushrooms are considered safe with lethal concentrations of 17 kg/70 kg in humans.23 Fatal incidences with the mushrooms are rare and mainly reported to be in combination with other drugs.23 Psilocybin-containing mushrooms are mainly studied with reference to their anti-depressant effects and very little information is known towards their anti-inflammatory potential. We have previously reported in vitro antioxidant and anti-inflammatory effects of Psilocybe natalensis, one of the well-known magic mushrooms that grow in South Africa on the RAW 267.4 macrophage.24 This study aimed at investigating further the anti-inflammatory effects of four psilocybin-containing mushrooms (Panaeolus cyanescens, Psilocybe natalensis, Psilocybe cubensis and Psilocybe cubensis leucistic A+ strain) from genus Panaeolus and Psilocybe for the first time in vitro on 15-lipoxygenase activity, and also on lipopolysaccharide (LPS)-induced inflammation in human U937 macrophage cells. The human U937 macrophage cell line has been widely used and categorized as a model of mammalian cellular response to various inflammatory stimuli.25 We used hot-water extracts of the four mushrooms, which is one of the common methods of consumption used by magic mushroom users.
Materials and Methods
Ethical Clearances
This study’s protocol was submitted and approved by the University of Pretoria Research ethic committee with the protocol number REC045-18. In South Africa psilocybin mushrooms are schedule 7 substances, and approval by the South African Department of Health Medical Control Council (MCC) was applied for, and a permit license POS 223/2019/2020 granted for the project.
Growing the Mushrooms and Making Water Extracts
The spore prints of Panaeolus (Copelandia) cyanescens (Pan cyanescens) commonly known as “Natal Blue Meanie”, Psilocybe natalensis (P. natalensis) commonly known as “Natal super strength”, Psilocybe cubensis (P. cubensis) commonly known as “Golden teacher” and Psilocybe cubensis leucistic A+ strain (P. A+ strain) mushrooms were verified with SKU number TBMN-1, TEA-1, NSS-1 and AAP-1 respectively, together with the growing sterile substrate kit (SSK-2) and were all purchased from Spore Spot Company, Durban, South Africa. On arrival the spores were inoculated, grown and harvested and extracted with boiling hot water according to Nkadimeng et al.’s method.24 The extracts were stored in dark in a fridge until use.
15-Lipoxygenase Activity Measurement
The anti-inflammatory activity of extracts was evaluated in vitro via the soybean 15-lipoxygenase (15-LOX) (Sigma, Aldrich) inhibitory assay according to the method of Ondua et al.26 in the presence of linoleic acid based on the formation of the complex Fe3+/xylenol orange (Sigma-Aldrich) with absorption at 560 nm. Percentage inhibition of 15-lipoxygenase activity was determined and the inhibition concentrations (IC50) were determined using the non-linear regression curve of the 15-LOX percentage inhibition against the logarithm of concentrations tested.
Effects of the Extracts on LPS-Induced U937 Macrophages
Human U937 Macrophage Cell Culture
The human U937 macrophage cell line was obtained from ATCC and the cells were grown in RPMI 1640 (Sigma-Aldrich) medium supplemented with 10% fetal bovine serum and 1% of penicillin (100 units/mL) and streptomycin (100 µg/mL) (Celtic Molecular Diagnostics) at 37 °C under 5% CO2. Cells were harvested once the cell confluency reached approximately 80–90%. For all the experiments, the U937 cells were differentiated to obtain macrophage like phenotype by addition of (200 nM) phorbol 12-myristate 13-acetate (PMA) (Biocom Africa) for 24 h according to Harikrishnan et al.’s method.5 After 24 h the cells were washed with normal medium once and then incubated for 24 h with serum free media for recovery phase.
Treatment with the Extracts
Cells were then treated according to Harikrishnan et al.’s method5 with modifications. Briefly, differentiated cells were seeded in 25 cm2 flasks (Lasec) at a density of 2×106 and then treated with the hot-water extracts (25 and 50 µg/mL) and quercetin (50 µg/mL) (Sigma-Aldrich), a well-known antioxidant and a flavonol found in many fruits and plants, for 2 h before being induced with LPS (1 µg/mL) (Sigma-Aldrich) and incubated at 37°C in a 5% CO2 incubator. LPS cells were differentiated, induced with LPS but not treated. Control cells were differentiated but neither induced with LPS nor treated. The cells were exposed and treated over 24 h. After 24 h medium was removed and stored in −80°C freezer until day of analysis.
COX-2 Activity Measurements
The effects of the extracts on COX-2 levels were determined using the human PTGS2/COX-2 Prostaglandin endoperoxide synthase 2 (PGE2) ELISA kit (Elabscience, Biocom Africa) according to the manufacturer’s manual protocol. The absorbance was directly proportional to the concentrations of PTGS2/COX-2 in the sample medium. Concentrations of human COX-2 in the cell culture media samples were calculated from the standard curve.
Cytokine Activity Measurements
The effects of the extracts on the concentrations of TNF-α, IL1 β, IL6 and IL10 were determined and quantified using the human ELISA kits E-EL-H0109, E-EL-H0149, E-EL-H0102 and E-EL-H0103, (Elabscience, Biocom Africa) respectively following the instruction manual on the cell culture medium. Concentrations of the cytokines were calculated from standard curves.
Cytotoxicity of Extracts on LPS-Induced Human U937 Macrophages
Differentiated human U937 macrophage cells were seeded at a density of 5×105 cells/well into the 24-well plates (NEST, Whitehead Scientific) and incubated for 24 h at 37°C in 5% CO2. The cells were pre-treated with the four hot-water extracts (25 and 50 µg/mL) and positive control, quercetin (50 µg/mL) for 2 h and then induced with (1 µg/mL) LPS over 24 h same as before. After 24-h incubation mitochondrial activity was measured using the Resazurin assay kit AR002 (R & D, Whitehead Scientific) according to the manufacturer’s manual. Viability of cells in percentages was calculated using the formula: % Viability = ((Sample Absorbance/control Absorbance) × 100). The experiments were repeated three times on different occasions.
Statistical Analysis
Results are expressed as mean ± standard deviations and statistically significant values were compared using one-way ANOVA and pairwise multiple comparison procedures using Holm-Sidak method. Normality test was done using Shapiro–Wilk and equal variance test using Brown-Forsythe. The p-value of ≤ 0.050 was considered statistically significant.
Results
Inhibition Effects of the Mushroom Extract On 15-LOX Activity
The four hot-water extracts had very low 15-LOX inhibition activities with IC50> 250 µg/mL in comparison with the positive control, quercetin with IC50 of 12.35 µg/mL, Table 1.
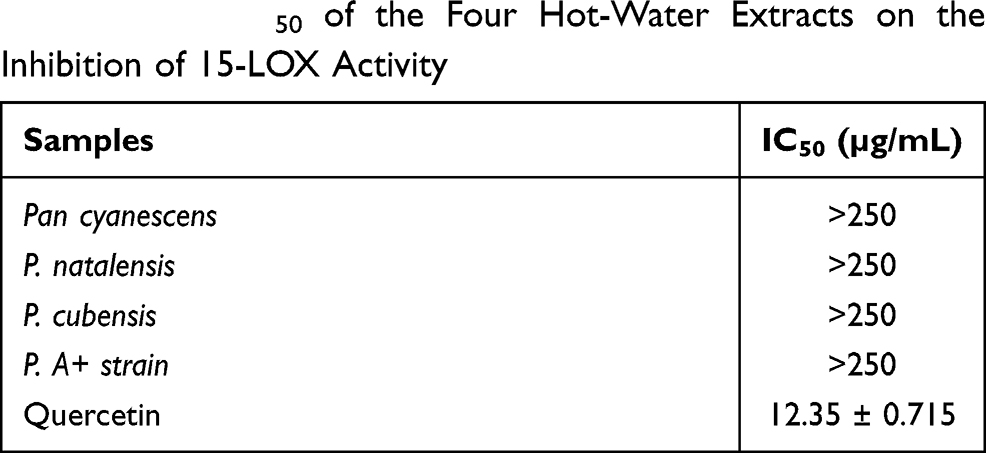 |
Table 1 The IC50 of the Four Hot-Water Extracts on the Inhibition of 15-LOX Activity |
Inhibition Effects of the Mushroom Extracts on LPS-Induced COX-2 Production
Stimulation of the human U937 macrophage cells with LPS increased the concentration of COX-2 significantly (p = 0.009) compared with the control non-stimulated cells, Figure 1. Treatment with the four hot-water extracts reduced the levels of COX-2 and the effect was significant (p = 0.039) with the lowest concentration (25 µg/mL) of the P. A+ strain mushroom extracts, Figure 1.
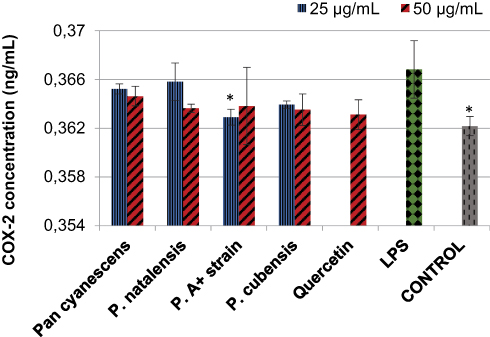 |
Figure 1 Effects of the hot-water extracts (25 and 50 µg/mL) of Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain mushrooms and the positive control; quercetin (50 µg/mL) on LPS-induced COX-2 concentration over 24 h. Control cells were differentiated but neither induced with LPS nor treated. LPS: differentiated and LPS-induced. (*Statistically significant). |
Effects of the Mushroom Extracts on LPS-Induced Pro-Inflammatory TNF-α Cytokine Levels
LPS increased significantly (p = 0.003) the production of TNF-α cytokine levels in stimulated cells compared with the control cells, Figure 2. The positive control quercetin reversed this effect (p< 0.001) significantly. The four mushroom extracts also inhibited TNF-α concentration significantly with both the 25 and 50 µg/mL concentrations, Figure 2.
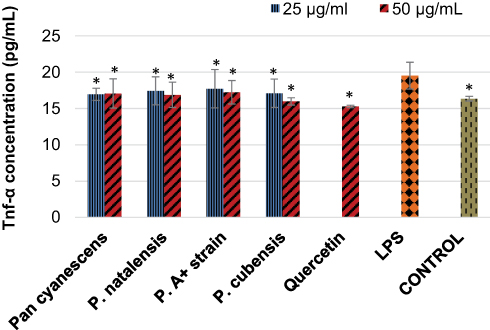 |
Figure 2 Inhibitory effects of the hot-water extract (25 and 50 µg/mL) of Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain mushrooms and the positive control, quercetin (50 µg/mL) on the LPS-induced TNF-α production over 24 h. Control cells were differentiated but neither induced with LPS nor treated. LPS: differentiated and LPS-induced. (*Statistically significant). |
Effects of the Mushroom Extracts on LPS-Induced Pro-Inflammatory IL-1β and IL-6
Stimulation with LPS increased significantly the IL-1β production (p< 0.001) in comparison to the control, Figure 3. The four hot-water extracts inhibited the LPS-induced IL-1β significantly to the levels very close to the positive control, Figure 3. LPS also increased production of IL-6 significantly (p = 0.010) compared with the control and quercetin reversed this effect significantly (p = 0.003), Figure 3. The four hot-water extracts reduced the IL-6 production in treated cells however; the inhibition was only significant with 25 and 50 µg/mL concentrations of P. natalensis (p = 0.025 and p = 0.015 respectively) and the highest 50 µg/mL concentration of P. cubensis (p = 0.040), Figure 3.
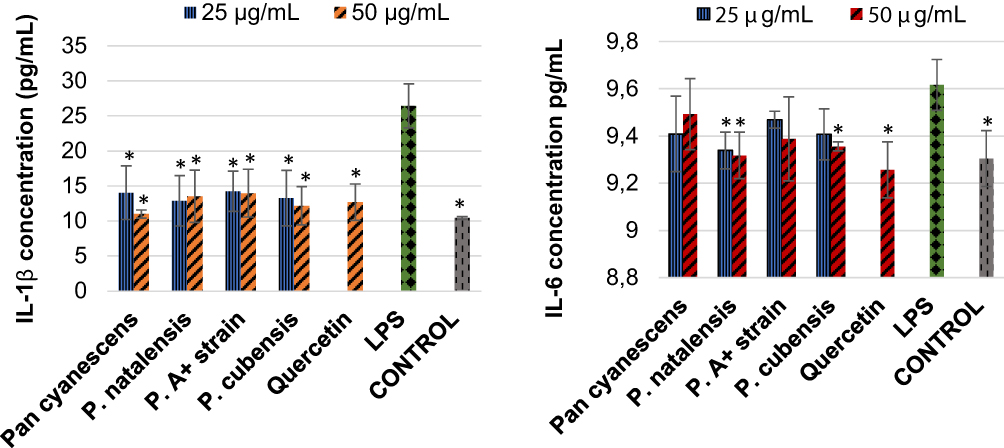 |
Figure 3 Inhibitory effects of the hot-water extracts (25 and 50 µg/mL) of Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain mushrooms and quercetin on the LPS-induced production of pro-inflammatory IL-1β and IL-6 cytokines over a 24-h treatment period. Control cells were differentiated but neither induced with LPS nor treated. LPS: differentiated and LPS-induced. (*Statistically significant). |
Effects of the Mushroom Extracts on the Anti-Inflammatory IL-10 Cytokine
Stimulation with LPS decreased non-significantly the IL-10 concentration of the induced cells in comparison to the control, Figure 4. The four water extracts increased non-significantly the LPS-induced IL-10 production of the treated cells, the same as quercetin. Effects were more pronounced with the P. cubensis mushroom, Figure 4.
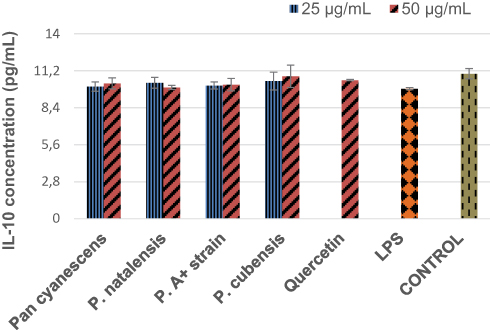 |
Figure 4 The effects of the hot-water extract (25 and 50 µg/mL) of Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain mushrooms and quercetin on the anti-inflammatory IL-10 cytokines levels over a 24-h treatment period. LPS: differentiated and LPS-induced. |
Effects of the Extracts on % Cell Viability
The treatment of the LPS-induced cells with the four extracts increased percentage viability of the cells above 100%, similar to the positive control quercetin, Figure 5. Pan cyanescens, P. cubensis and P. natalensis had the highest percentage viability with the lower concentrations and displayed a deceleration dose-dependent pattern such that percentage cell viability decreased with increase in concentrations while the P. A+ strain increased percentage viability as concentration increased, the same as quercetin, Figure 5.
 |
Figure 5 Effects of Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain mushroom extracts and quercetin on the percentage cell viability of LPS-induced human U937 macrophage cells over a 24-h treatment period. |
Discussion
Although psilocybin-containing magic mushrooms have been used for centuries for their mind healing properties, very little is known about their anti-inflammatory potential. Many studies have shown that pathological inflammation is at the center of a number of chronic diseases including cardiovascular, cancer, arthritis, neurodegeneration, chronic depression and aging.2,27,28 Numerous studies have also showed that over time, prolonged inflammation may cause harm to the tissues around the affected area.3 Furthermore, studies have also reported the adverse effects associated with the uses of the current NSAIDs as treatment for inflammatory diseases which include disturbances in the upper gastrointestinal system, leading to the need for more research into remedies (particularly natural products) with anti-inflammatory properties.5,11 The purpose of this study was to investigate the anti-inflammatory effects of four well-known psilocybin-containing magic mushrooms, Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain, for the first time in vitro on 15-LOX activity, and also on LPS-induced inflammation in human U937 macrophage cells, in order to establish their potential use in inflammatory diseases. We used the hot-water extraction method, which is one of the most common methods of consumption used by the magic mushroom users.
The potential inhibitory ability of the mushroom extracts on the 15-LOX activity, which is one of the vital enzymes in the biosynthesis of leukotrienes that play an important role in the pathophysiology of many inflammatory diseases, was examined in this study. Our study revealed that the hot-water mushroom extracts of all four psilocybin-contain mushrooms had very low inhibition properties as indicated by very high IC50 > 250 μg/mL on the 15-LOX activity.
We further investigated the anti-inflammatory effects of the four hot-water mushroom extracts on LPS-induced human U937 macrophage cells, an extensively used cell line categorized as a model of mammalian cellular response to various inflammatory stimuli.25 Lipopolysaccharides activate macrophages to produce an unhealthy accumulation of inflammatory mediators such as inducible COX-2 and nitric oxide synthase (iNOS), and pro-inflammatory cytokines such as TNF-α, IL-1β and IL-6.5 In our study, LPS stimulation increased concentrations of COX-2, TNF-α, IL-1β and IL-6 significantly in the stimulated cells compared with non-stimulated control cells in agreement with previous studies.5
The LPS-induced COX-2 effects were reversed significantly by quercetin and the 25 μg/mL concentration of P. A+ strain mushroom extracts. Treatment with other mushroom extracts also lowered the LPS-induced COX-2 concentration even though non-significantly. Inducible COX-2 enzymes play a critical role in pathological inflammation where they are responsible for edema, pain and tissue damage associated with the disease process.29 As a result, remedies with the ability to inhibit of COX-2 expression or concentrations have been identified by many studies as valuable tools in the prevention or treatment of pathological inflammation disease.29
In our study, treatment with the four hot-water mushroom extracts reversed the LPS-induced TNF-α and IL-1β and inhibited significantly the concentrations of these two key pro-inflammatory cytokines in a dose-dependent manner and very close to the positive control, quercetin. Studies have shown that TNF-α and IL-1β are among the main pro-inflammatory cytokines secreted in the development of chronic inflammatory disease. As a result, by inhibiting these two key proinflammatory cytokines, the extracts showed potential potent anti-inflammatory properties that could be beneficial in chronic inflammatory disease treatment. One disease highly affected by inflammation in particular is osteoarthritis (OA), which affects about 25% of the global population and is mainly characterized by destruction in the functioning and structure of articular cartilage.30 Studies have shown pro-inflammatory cytokine IL-1B as an important key factor in the development and progression of OA implemented by stimulating both the pro-catabolic and the pro-inflammatory chondrocyte (cartilage cells) response.30 Chondrocytes play a fundamental role in maintaining normal tissue synthesis and also the renewal of old extracellular matrix.30 Consequently IL-1β-induced catabolic response promotes suppression of cartilage-related genes expression and increases expression of matrix-degrading related genes that include collagenases and matrix metalloproteinases.30 Furthermore, IL-1β also stimulates chondrocytes to increase expression of pro-inflammatory related proteins and genes such as iNOS and COX-2 which when induced further decrease the primary components of cartilage extracellular matrix.32 Studies have also shown that inhibition of COX-2 expression have suppressive effects on OA.33 Moreover, in the study of Yang et al.31 where IL-1β was used to induce a degenerated chondrocyte model of hypertrophy, there was an up-regulated increase in the expressions of many proteins and enzymes including COX-2, PGE-2, and collagen X which were reversed by treatment with a potent COX-2 inhibitor used in the study.31 As a result, the ability of the hot-water extracts of the four magic mushrooms to significantly inhibit especially the induced-IL-1β and also lower the induced-COX-2 levels (significantly with P. A+ strain treatment), make them a great agent for use in treatment of inflammatory diseases such as AO.
Furthermore, P. natalensis and P. cubensis mushroom extracts inhibited the LPS-induced IL-6 concentrations significantly while the other mushroom extracts lowered the levels non-significantly in human macrophages. We also observed that there was a reduction in anti-inflammatory cytokine IL-10, an important modulator with a preventative role in pathological inflammation conditions, in the LPS-induced human macrophage in our study which was increased, however non-significantly by the treatment with the four mushroom extracts and quercetin.
All these effects were not due to cytotoxicity as indicated by the increase in percentage cell viability of the treated cells beyond 100% with all the four mushroom extracts.
In summary, the present study demonstrated a significant increase in pro-inflammatory mediators, COX-2, pro-inflammatory cytokines TNF-α, IL-1β and IL-6 which were significantly reversed by quercetin in LPS-stimulated human U937 macrophage cells. The hot-water extracts of P. A+ strain mushroom inhibited significantly the LPS-induced COX-2 while the other extracts lowered the levels non-significantly. This effect is important as remedies with COX-2 inhibition are regarded as valuable tools in the prevention or treatment of pathological inflammatory disease.29 The study also revealed that the four hot-water mushroom extracts of Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain inhibited significantly the two key pro-inflammatory cytokines TNF-α, IL-1β in a dose-dependent manner and very close to quercetin, a well-known potent anti-inflammatory flavanol and antioxidant found in many fruits and plants. In particular, the significant suppressing effect on the induced IL-1β and the lowering of COX-2 observed with the mushroom treatments in the study indicated their potential use especially in inflammation-related diseases such AO. Further investigation on the potential of these magic mushrooms to alleviate OA is highly recommended. Furthermore, P. natalensis and P. cubensis also inhibited significantly the LPS-induced IL-6 in human U937 macrophage cells while the other hot-water extracts lowered the levels, although non-significantly. The extracts also increased non-significantly the concentrations of anti-inflammatory cytokine IL-10 in the treated human macrophage cells. These effects were not due to toxicity as indicated by increased viability of cells above 100% in the treated cells. Previous studies have revealed the presence of mycochemical compounds in psilocybin-containing mushrooms such as alkaloids and saponins which are known as potent antioxidants that neutralize free radicals and flavonoids with antioxidant and anti-inflammatory activities and also tannins with antioxidant properties especially in Pan cyanescens and P. cubensis.32,33 We have also previously reported the presence of well-known anti-inflammatory and antioxidant compounds in P. natalensis mushroom extracts.24 Moreover, our previous study also showed a significant inhibition of ROS displayed by Pan cyanescens and P. cubensis in a pathological hypertrophy condition.34 Excessive amounts of ROS are known to cause inflammation by stimulating the release of cytokines and subsequent activation of COX and LOXs signaling in a pathological inflammation disease.17 The presence of these compounds in the mushroom extracts could have played a role in the inhibition of key pro-inflammatory cytokines and reduction in COX-2 observed in this study. However, the study showed poor inhibition of 15-LOX activity with the four hot-water mushroom extracts suggesting that the extracts may not necessary induce their anti-inflammatory effects via this pathway.
Conclusion
In conclusion, this study demonstrated that the hot-water extracts of Pan cyanescens, P. natalensis, P. cubensis and P. A+ strain magic mushrooms suppressed inflammatory response induced by LPS stimulation in human U937 macrophages. The study also proposed that these effects were executed by mechanisms that involved inhibition of the pro-inflammatory mediators such as COX-2 and pro-inflammatory cytokines. A further study into the underlying mechanisms of action in vivo and in vitro are recommended.
Acknowledgments
We appreciate the support of Mr Llewellyn Morland who assisted with the growing of mushrooms, and Ms Lebogang E Moagi for assistance with statistics.
Funding
The study was funded by the Health and Welfare Sector Education and Training Authority (HWSETA) and MJ Medtech Grants to S.M. Nkadimeng. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Ferrucci L, Fabbri E. Inflammageing: chronic inflammation in ageing, cardiovascular disease, and frailty. Nat Rev Cardiol. 2018;15(9):505–522. doi:10.1038/s41569-018-0064-2
2. Liu Y-Z, Wang Y-X, Jiang C-L. Inflammation: the common pathway of stress-related diseases. Front Hum Neurosci. 2017;11:316. doi:10.3389/fnhum.2017.00316
3. Muniandy K, Gothai S, Badran KMH, Suresh Kumar S, Esa NM, Arulselvan P. Suppression of proinflammatory cytokines and mediators in LPS-Induced RAW 264.7 macrophages by stem extract of alternanthera sessilis via the inhibition of the NF-κB pathway. J Immunol Res. 2018;2018:3430684. doi:10.1155/2018/3430684
4. Matzinger PT. Danger, and the extended family. Annu Rev Immunol. 1994;12(1):991–1045. doi:10.1146/annurev.iy.12.040194.005015
5. Harikrishnan H, Jantan I, Haque MA, Kumolosasi E. Anti-inflammatory effects of Phyllanthus amarus Schum. & Thonn. through inhibition of NF-κB, MAPK, and PI3K-Akt signaling pathways in LPS-induced human macrophages. BMC Complement Altern Med. 2018;18(1):224. doi:10.1186/s12906-018-2289-3
6. Zhang J-M, An J. Cytokine, inflammation and pain. Int Anaesthesiol Clin. 2009;45(2):27–37. doi:10.1097/AIA.0b013e318034194e
7. Lauwerys BR, Houssiau FA. Involvement of cytokines in the pathogenesis of systemic lupus erythematosus. Adv Exp Med Biol. 2003;520:237–251. doi:10.1007/978-1-4615-0171-8_14
8. Porro C, Gianciulli A, Panaro MA. The regulatory role of IL-10 in neurodegenerative diseases. Biomolecules. 2020;10(7):1017. doi:10.3390/biom10071017
9. Zeldin DC. Epoxygenase pathways of arachidonic acid metabolism. J Biol Chem. 2001;276(39):36059–36062. doi:10.1074/jbc.R100030200
10. Sevanian A, Kim E. Phospholiphase A2 dependent release of fatty acids from peroxidized membranes. Free Radic Biol Med. 1985;1(4):263–271. doi:10.1016/0748-5514(85)90130-8
11. Attiq A, Jalil J, Husain K, Ahmad W. Raging the war against inflammation with natural products. Front Pharmacol. 2018;9:976. doi:10.3389/fphar.2018.00976
12. Samuelsson B, Goldyne M, Granström E, et al. Prostaglandins and thromboxanes. Annu Rev Biochem. 1978;47(1):997–1029. doi:10.1146/annurev.bi.47.070178.005025
13. Sugimoto Y, Inazumi T, Tsuchiya S. Roles of prostaglandin receptors in female reproduction. J Biochem. 2015;157(2):73–80. doi:10.1093/jb/mvu081
14. Kawahara K, Hohjoh H, Inazumi T, Tsuchiya S, Sugimoto Y. Prostaglandin E 2-induced inflammation: relevance of prostaglandin E receptors. Biochim Biophys Acta. 2015;1851(4):414–421. doi:10.1016/j.bbalip.2014.07.008
15. Mbonye UR, Yuan C, Harris CE, et al. Two distinct pathways for cyclooxygenase-2 protein degradation. J Biol Chem. 2008;283(13):8611–8623. doi:10.1074/jbc.M710137200
16. Kuhn H, Humeniuk L, Kozlov N, Roigas S, Adel S, Heydeck D. The evolutionary hypothesis of reaction specificity of mammalian ALOX15 orthologs. Prog Lipid Res. 2018;72:55–74. doi:10.1016/j.plipres.2018.09.002
17. Singh NK, Rao GN. Emerging role of 12/15-Lipoxygenase (ALOX15) in human pathologies. Prog Lipid Res. 2019;73:28–45. doi:10.1016/j.plipres.2018.11.001
18. Lončarić M, Strelec I, Moslavac T, Šubarić D, Pavić V, Molnar M. Lipoxygenase inhibition by plant extracts. Biomolecules. 2021;11(2):152. doi:10.3390/biom11020152
19. Srivastava P, Vyas VK, Variya B, et al. Synthesis, anti-inflammatory, analgesic, 5-lipoxygenase (5-LOX) inhibition activities, and molecular docking study of 7-substituted coumarin derivatives. Bioorg Chem. 2016;67:130–138. doi:10.1016/j.bioorg.2016.06.004
20. Namgaladze D, Snodgrass RG, Angioni C, et al. AMP-activated protein kinase suppresses arachidonate 15-lipoxygenase expression in interleukin 4-polarized human macrophages. J Biol Chem. 2015;290(40):24484–24494. doi:10.1074/jbc.M115.678243
21. Mittal M, Siddiqui MR, Tran K, Reddy SP, Malik AB. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal. 2014;20(7):1126–1167. doi:10.1089/ars.2012.5149
22. Griffith B, Pendyala S, Hecker L, et al. NOX enzymes and pulmonary disease. Antioxid Redox Signal. 2009;11(10):2505–2516. doi:10.1089/ars.2009.2599
23. Amsterdam JGC, Opperhuizen A, van den Brink W. Harm potential of magic mushroom: a review. Regul Toxicol Pharmacol. 2011;59(3):423–429. doi:10.1016/j.yrtph.2011.01.006
24. Nkadimeng SM, Nabatanzi A, Steinmann CML, Eloff JN. Phytochemical, cytotoxicity, antioxidant and anti-inflammatory effects of Psilocybe natalensis magic mushroom. Plants. 2020;9(9):127. doi:10.3390/plants9091127
25. Grkovich A, Johnson CA, Buczynski MW, Dennis EA. Lipopolysaccharide-induced cyclooxygenase-2 expression in human U937 macrophages is phosphatidic acid phosphohydrolase-1-dependent. J Biol Chem. 2006;281(44):32978–32987. doi:10.1074/jbc.M605935200
26. Ondua M, Njoya EM, Abdalla MA, McGaw LJ. Anti-inflammatory and antioxidant properties of leaf extracts of eleven South African medicinal plants used traditionally to treat inflammation. J Ethnopharmacol. 2019;234:27–35. doi:10.1016/j.jep.2018.12.030
27. Nabatanzi A, Nkadimeng SM, Lall N, Kabasa JD, McGaw LJ. Antioxidant and anti-inflammatory activities of Kigelia africana (Lam.) Benth. Evid Based Complement Alternat Med. 2020;2020:1–11. doi:10.1155/2020/4352084
28. Needleman P, Manning PT. Interactions between the inducible cyclooxygenase (COX-2) and nitric oxide synthase (iNOS) pathways: implications for therapeutic intervention in osteoarthritis. Osteoarthr Cartil. 1999;7(4):367–370. doi:10.1053/joca.1998.0237
29. Baraf HS. Efficacy of the newest COX-2 selective inhibitors in rheumatic disease. Curr Pharm Des. 2007;13(22):2228–2236. doi:10.2174/138161207781368819
30. Jia Y, He W, Zhang H, et al. Morusin ameliorates IL-1β-induced chondrocyte inflammation and osteoarthritis via NF-κB signal pathway. Drug Des Devel Ther. 2020;14:1227–1240. doi:10.2147/DDDT.S244462
31. Yang YR, Yang XF, Duan HC, Qiao JQ. Cyclooxygenase-2 inhibitor rofecoxib prevents chondrocytes against hypertrophy via Wnt/β-catenin pathway. J Biol Regul Homeost Agents. 2020;34(3):785–794.
32. Bustillos RG, Dulay RMR, Bauto JJ, et al. Mycochemical profile of mycelia and fruiting body of Panaeolus cyanescens and its optimal submerged culture conditions for antioxidant properties. Int J Pure Appl Biosci. 2014;2(6):175–181.
33. Dhanasekaran D, Latha S, Suganya P, et al. Taxanomic identification and bioactive compounds characterization of Psilocybe cubensis DPT1 to probe its antibacterial and mosquito larvicidal competency. Microb Pathog. 2020;143:104138. doi:10.1016/j.micpath.2020.104138
34. Nkadimeng SM, Steinmann CML, Eloff JN. Effects and safety of Psilocybe cubensis and Panaeolus cyanescens magic mushroom extracts on endothelin-1-induced hypertrophy and cell injury in cardiomyocytes. Sci Rep. 2020;10(1):22314. doi:10.1038/s41598-020-79328-5
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.