Back to Journals » ImmunoTargets and Therapy » Volume 15
An Exploratory in vitro Study of Cytotoxicity and Cytokine Profiling of Cytokine-Induced Killer (CIK) Cells Against HCT-15 Colorectal Cancer (CRC) Cells
Authors Chin SP ![]() , Nor Arfuzir NN, Kong VWP, Cheong SK
, Nor Arfuzir NN, Kong VWP, Cheong SK ![]()
Received 12 January 2026
Accepted for publication 14 April 2026
Published 22 May 2026 Volume 2026:15 594441
DOI https://doi.org/10.2147/ITT.S594441
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sarah Wheeler
Sze-Piaw Chin,1,2 Natasha Najwa Nor Arfuzir,1 Vivian Wai-Ping Kong,3 Soon-Keng Cheong1,2
1Department of Research and Development, Cytopeutics Sdn Bhd, Cyberjaya, Selangor, Malaysia; 2M. Kandiah Faculty of Medicine & Health Sciences, University Tunku Abdul Rahman Sungai Long City Campus, Kajang, Selangor, Malaysia; 3Laboratory Department, CryoCord Sdn Bhd, Cyberjaya, Selangor, Malaysia
Correspondence: Sze-Piaw Chin, Department of Research and Development, Cytopeutics Sdn Bhd, Cyber 8, Cyberjaya, Selangor, 63000, Malaysia, Email [email protected]
Background: Cytokine-induced killer (CIK) cells are a promising adoptive immunotherapy due to their non-major histocompatibility complex (MHC)-restricted cytotoxicity. However, their cytokine-mediated response against CRC remains poorly defined.
Methods: In this exploratory study, CIK cells were generated from three healthy donors and one metastatic CRC patient. Phenotypes were assessed by flow cytometry. Cytotoxicity against the HCT-15 CRC cell line was evaluated at multiple effector-to-target (E:T) ratios (1:5 to 40:1). Combination treatment with 5-fluorouracil (5-FU) and cytokine profiling using a 105-plex array were performed using CIK cells from donor 2 and donor 3, respectively.
Results: CIK cells derived from one CRC patient exhibited phenotypic characteristics (CD3⁺CD8⁺ 85.91%, CD3⁺CD56⁺ 27.81%) comparable to the range observed in three healthy donors. Co-culture with CIK cells from donor 1 resulted in reduced HCT-15 viability at higher E:T ratios, with viability decreasing to 8.98 ± 0.61% and 8.08 ± 2.00% at 20:1 and 40:1, respectively. In combination experiments, HCT-15 viability was reduced to 1.02 ± 1.01% at an E:T ratio of 20:1 when co-cultured with CIK from donor 2 and treated with 5-FU. Similar reductions were observed with CIK cells from donor 3 and the CRC patient, reaching 2.60 ± 5.86% and 0.0% at 20:1, and 0.0% at 40:1 ratio for both donors. CIK cells from all donors showed dose-dependent reduction in cell viability against HCT-15 cells. Cytokine profiling of CIK from donor 3 identified changes in immune-related factors, including upregulation of interferon gamma-inducible chemokines (CXCL9 and CXCL10), alongside downregulation of immunosuppressive mediators such as TIM-3 and VEGF.
Conclusion: This exploratory study demonstrates that functional CIK cells can be generated from healthy individuals and CRC patients, with observed cytotoxic activity against HCT-15 cells. The findings suggest potential immunomodulatory effects and support further investigation of CIK cells in combination strategies; however, larger patient cohorts and functional validation studies are required.
Keywords: cytokine-induced killer, CIK, natural killer, NK, natural killer T, NKT, colorectal cancer, CRC, immunotherapy
Introduction
Colorectal cancer (CRC) is one of the leading causes of gastrointestinal (GI) cancer-related mortality worldwide. According to the Global Cancer Observatory (GLOBOCAN) 2022 report by the International Agency for Research on Cancer (IARC), colorectal cancer (CRC) ranks among the top three cancers and is the leading cause of GI cancer mortality. In 2022, there were an estimated 1.9 million new CRC cases and over 900,000 deaths globally.1 Despite advances in chemotherapy, therapeutic resistance and disease progression, which limit treatment options for advanced-stage disease remain major clinical challenges.1,2 In addition, the majority of CRCs particularly microsatellite-stable subtypes, are considered immunologically “cold” tumors, characterized by low tumor-infiltrating lymphocytes and limited responsiveness to immune checkpoint blockade.3,4 These limitations have driven increasing interest in immunotherapeutic strategies, particularly in adoptive cell therapy, as an alternative approach to enhance antitumor immunity in CRC.
Cytokine-induced killer (CIK) cells are a heterogeneous population of immune effector cells characterized by their non-major histocompatibility complex (MHC)-unrestricted cytotoxicity and ability to target a broad range of tumor cells.5,6 They are primarily composed of T cells (CD3⁺CD8+), natural killer T (NKT) cells (CD3⁺CD56⁺), and a subset of natural killer (NK) cells (CD3−CD56⁺).5 CIK cells can be expanded in vitro from peripheral blood mononuclear cells (PBMCs) through priming with interferon-γ (IFN-γ) and anti-CD3 antibodies, followed by stimulation with interleukin-2 (IL-2).5 Under optimized culture conditions, CIK cells can undergo substantial expansion while retaining potent cytotoxic activity.7,8 Their MHC-unrestricted killing mechanism makes them particularly attractive for targeting tumors with low immunogenicity, such as CRC.7
In addition to cytokine stimulation, the microenvironment plays an important role in regulating immune cell behavior during ex vivo expansion. Extracellular matrix (ECM) components such as laminin support cell adhesion and influence cell activation, proliferation and survival through integrin-mediated signaling pathways.9,10 These ECM-immune cell interactions have been shown to modulate immune cell function and responsiveness.11 Incorporation of ECM-mimicking substrates during culture may therefore provide a supportive platform to enhance CIK cell expansion and functionality. CIK cells exert antitumor effects through multiple mechanisms, including NKG2D-mediated cytotoxicity, leukocyte function-associated antigen-1 (LFA-1)–dependent adhesion to tumor cells, and Fas ligand (FasL)–induced apoptosis.5
Beyond direct cytotoxicity, CIK cells actively modulate the immune microenvironment through the secretion of cytokines and chemokines. Upon tumor stimulation, pro-inflammatory cytokines such as tumor necrosis factor (TNF)-α, IFN-γ, and IL-2 are upregulated, contributing to immune activation and recruitment of effector cells.8 Cytotoxic ligands such as FasL and TNF-related apoptosis-inducing ligand (TRAIL), which induce tumor cell death, are also increased. Emerging evidence suggests that the cytokine milieu plays a critical role in shaping tumor-immune interactions by regulating immune cell recruitment, activation, and immunosuppressive mechanisms within the tumor microenvironment.12 However, despite increasing interest in CIK cell therapy, the comprehensive profiling of cytokine responses in the context of CRC, particularly in relation to tumor-immune interactions, remains insufficiently characterized.
Previous preclinical and clinical studies have demonstrated the therapeutic potential of CIK cells in gastrointestinal malignancies, including CRC.13–15 In particular, combination strategies integrating CIK cells with chemotherapy have shown promising results in enhancing antitumor efficacy and potentially overcoming drug resistance.14,16 However, the underlying immunomodulatory mechanisms, especially those related to cytokine networks within the tumor microenvironment, are not fully understood. A more detailed characterization of these cytokine-mediated interactions may provide important insights into optimizing CIK-based therapies and identifying potential biomarkers of response.
Therefore, this exploratory study aimed to (1) generate Cytopeutics® CIK cells from metastatic CRC patients and healthy donors, (2) compare their cytotoxicity against the HCT-15 cell line, (3) assess the enhanced cytotoxic effects of Cytopeutics® CIK cells in combination with fluorouracil (5-FU) which is a widely used chemotherapeutic agent and backbone of standard treatment regimens for colorectal cancer, and (4) identify the growth factors and cytokines associated with the antitumor activity. These findings provide preliminary insights into the feasibility and mechanistic potential of CIK cell-based immunotherapy for CRC.
Materials and Methods
Cell Lines
The human colorectal cancer (CRC) cell line HCT-15 was obtained from the American Type Culture Collection (ATCC; USA). HCT-15 has been reported to harbor KRAS mutation, and is widely used as an in vitro CRC model for investigating tumor biology and responses to therapeutic agents.17,18 Cells were cultured in RPMI-1640 medium (Gibco, USA) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic-antimycotic solution (all from Gibco, USA), and incubated at 37°C in a humidified 5% CO2 incubator (Memmert GmbH, Germany).
Study Group
Peripheral blood mononuclear cell (PBMC) samples were collected from three healthy donors and one Stage IV CRC patient. Written informed consent was obtained from all participants. The study protocol was approved by the Medical Research and Ethics Committee (MREC), Ministry of Health Malaysia (MOH) (Approval No. 24–03956-VSD) and conducted according to the Declaration of Helsinki (1975, revised 2013). Donor identities were anonymized (Table 1).
 |
Table 1 Characteristics of Healthy Donors and CRC Patient |
Generation of CIK Cells
CIK cells were generated as described previously.19 PBMCs were isolated using Lymphoprep™ density gradient medium (STEMCELL Technologies, Canada) and cultured in a laminin-coated 175 cm2 flask containing LymphoOne T-cell expansion medium (Takara Bio, Japan), supplemented with heat-inactivated autologous plasma. Laminin-coated culture flasks were used to provide ECM-mimicking substrate that facilitates immune cell adhesion and supports cell expansion under ex vivo conditions through integrin-mediated signaling.10 Cultures were primed with recombinant human IFN-γ (PeproTech, USA) at a total dose of 1000 IU per culture for 24 hours, followed by stimulation with 400 ng/mL anti-CD3 monoclonal antibody (T&L Biological Technology Co., Ltd, China) and 1000 U/mL recombinant human IL-2 (Four Rings Bio-Pharmaceutical Co., Ltd, China). Cells were incubated at 37°C in a 5% CO2 incubator for 21 days, with LymphoOne medium and IL-2 replenished every 2–3 days.
Phenotypic Characterization of CIK Cells
After 21 days of culture, CIK cells were harvested and analyzed by flow cytometry to determine the proportions of CD3+CD8+ T cells, CD3+CD56+ NKT cells, and CD3−CD56+ NK cells. A total of 5×105 cells were stained with anti-human antibodies: CD3-APC, CD8-FITC, and CD56-PE for 15 minutes in the dark at room temperature, washed twice, and stained with 7-aminoactinomycin D (7-AAD) for viability assessment. Data from 100,000 events per sample were acquired using a FACSLyrics™ flow cytometer, and analyzed with FACSuite™ Clinical Software (all from BD Biosciences, USA).
In vitro Cytotoxicity Assay of CIK Cells from Healthy Donor 1 Against HCT-15 Cells
The cytotoxicity of CIK cells against HCT-15 cells was evaluated using MTT assay across effector to target (E:T) ratios ranging from 40:1, 20:1, 10:1, 5:1, 1:1, 1:2, and 1:5. HCT-15 cells were seeded overnight in a 96-well plate at a density of 5000 cells/well. CIK cells derived from healthy donor 1 were harvested and resuspended in LymphoOne T-cell expansion medium supplemented with heat-inactivated autologous plasma and 1000 U/mL recombinant human IL-2, and added to the wells at the designated E:T ratios in an equal volume of the target cell medium. After 72 hours of co-culture at 37°C in a 5% CO2 incubator, CIK cells were carefully removed by gentle washing with PBS to eliminate non-adherent effector cells. Subsequently, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was added into each well to reach a final concentration of 0.5 mg/mL, and the plate was incubated for 4 hours. Formazan crystals were solubilized with 150 µL of dimethyl sulfoxide (DMSO), and absorbance was measured at 570 nm (Infinite® M200 PRO; Tecan, Switzerland). Cell viability was normalized to untreated HCT-15 control.
The MTT assay was used to assess cell viability; however, it primarily reflects metabolic activity rather than direct cell death. Therefore, results should be interpreted as an indirect measure of cytotoxicity in co-culture systems.
In vitro Cytotoxicity Assay of CIK Cells from Healthy Donor 2 in Combination with 5-FU Against HCT-15 Cells
To assess the combinatorial effect of 5-fluorouracil (5-FU) with CIK cells, HCT-15 cells (10,000 cells/well) were seeded overnight in a 96-well plate and pre-treated with 5-FU (0.050, 0.075, or 0.100 mg/mL) for 24 hours. CIK cells derived from healthy donor 2 were harvested, resuspended in LymphoOne medium supplemented with heat-inactivated autologous plasma and 1000 U/mL IL-2, and co-cultured with HCT-15 cells at E:T ratios of 1:5, 1:1, 10:1, and 20:1 for 72 hours at 37°C in a 5% CO2 incubator. MTT assay was performed as described above. Cell viability was expressed relative to untreated HCT-15 control.
Comparison of Cytotoxicity of CIK Cells from Healthy Donor 3 versus Cancer Patient Against the HCT-15 Cells
CIK cells derived from healthy donor 3 and a metastatic CRC patient were co-cultured with HCT-15 cells (5000 cells/well) at E:T ratios of 40:1, 20:1, 10:1, 5:1, 1:1, 1:2 and 1:5 for 72 hours at 37°C in a 5% CO2 incubator. Following incubation, MTT assay was conducted as described above to determine relative cytotoxicity between donor and patient-derived CIK cells.
Proteome Profiler Assay
The Proteome Profiler Human XL Cytokine Array Kit (R&D System, USA) was used to identify the relative levels of 105 different human cytokines and chemokines in the conditioned media from CIK:HCT-15 co-culture, following the manufacturer’s protocol. Briefly, HCT-15 cells (150,000 cells/well) were seeded overnight in a 6-well plate. CIK cells from healthy donor 3 were added at E:T ratios of 1:1 and 10:1 and cultured for 72 hours. Conditioned media (supernatant) from CIK-only, HCT-15-only, and the co-culture of CIK:HCT-15 were collected and centrifuged at 5000 g for 10 minutes. Assays were performed according to the manufacturer’s instructions. Chemiluminescent signals were visualized using iBright™ FL1500 Imaging System (Thermo Fisher Scientific, USA), and the pixel density was quantified using iBright™ Analysis Software with background intensity subtracted.
Protein Functional Annotation and Enrichment Analysis
Functional annotation and enrichment analysis of significantly regulated proteins were performed using Database for Annotation, Visualization and Integrated Discovery version 6.7 (DAVID v6.7; USA) for gene ontology (GO) classification.20,21 Corresponding gene lists were mapped to the Homo sapiens reference database to identify enriched biological processes, molecular functions, and cellular components. Subsequently, the interrelationships among significantly regulated proteins identified in the cytokine array were analyzed using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING; version 12.0; https://string-db.org/) database.22 Protein-protein interaction (PPI) networks were constructed with a maximum false discovery rate (FDR) of 0.05, and a minimum strength threshold of 0.01.
Statistical Analysis
Data were analyzed using the GraphPad Prism (version 9.0; GraphPad Software, Inc., USA) and presented as mean ± standard deviation (SD) from technical triplicates of each donor. Statistical analyses were performed as appropriate for each experiment. Cytotoxicity assays involving single-variable comparisons were analyzed using one-way ANOVA, while combination treatment assays (CIK cells + 5-FU) were analyzed using two-way ANOVA, followed by Tukey’s post-hoc test. Comparisons of cytotoxic activity between CIK cells derived from CRC patient and healthy donors were performed using an independent t-test. Immunophenotyping of CIK cells and cytokine array data were analyzed using two-way ANOVA, followed by Tukey’s post-hoc test. Statistical significance was set at p < 0.05.
Results
Generation of CIK Cells from Healthy Donors and a CRC Patient
PBMCs isolated from three healthy donors and one CRC patient were successfully expanded into CIK cells, and their phenotype was analyzed by flow cytometry after 21 days of culture. Flow cytometry (Figure 1 and Supplementary Figure 1) showed an increase in CD3+CD8+ T cells and CD3+CD56+ NKT cells after 21 days, while CD3−CD56+ NK cells decreased. The observed reduction in CD3−CD56+ NK cells may reflect preferential expansion of CD3+CD8+ T and CD3+CD56+ NKT subsets under IL-2-driven culture conditions, as well as the influence of cytokine stimulation and activation-induced changes during in vitro expansion. CIK derived from CRC patient showed comparable subset proportions (CD3+CD8+ 85.91%; CD3+CD56+ 27.81%) to those observed in healthy donors (mean CD3+CD8+ 85.10 ± 2.55%; CD3+CD56+ 31.54 ± 13.15%) (Table 2), suggesting that metastatic CRC patients may be capable of generating similar percentages of CIK cell populations as healthy donors, although this observation is limited by small sample size.
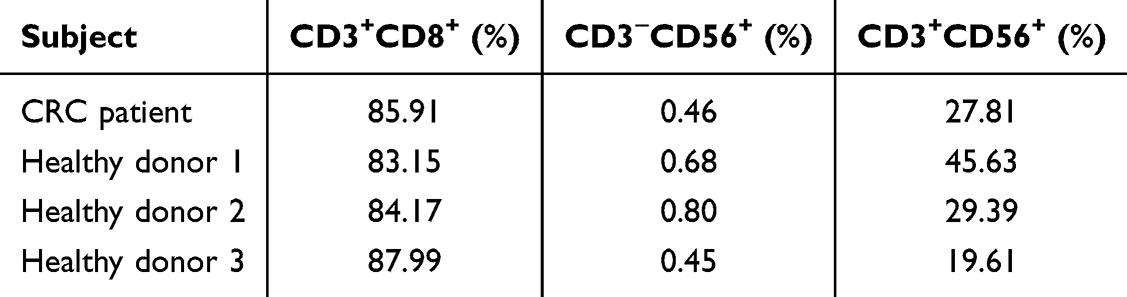 |
Table 2 Phenotypic Characteristics of CIK Cells from Patient with CRC and Healthy Donors After 21 Days of Culture |
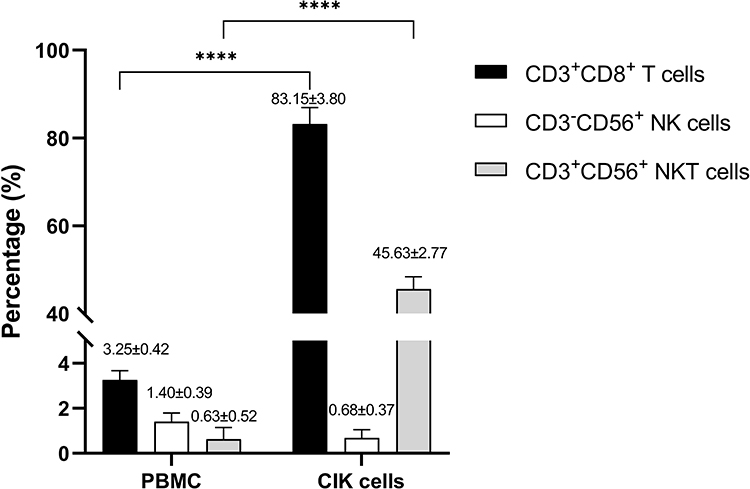 |
Figure 1 Immunophenotypic analysis of CIK cells derived from healthy donor 1 PBMCs. The percentages of T (CD3+CD8+), NK (CD3−CD56+) and NKT (CD3+CD56+) cells were determined by flow cytometry, with results expressed as mean ± SD from three healthy donors and one CRC patient (N = 4). Statistical analysis was assessed using two-way ANOVA. ****p < 0.0001. |
The Viability of HCT-15 Cells Co-Cultured with CIK Cells from Donor 1
To assess cytotoxic potential, CIK cells derived from healthy donor 1 were co-cultured with HCT-15 cells at various E:T ratios, and tumor cell viability was assessed using the MTT assay (Figure 2). At the lowest E:T ratio of 1:5, 47.73 ± 8.38% of HCT-15 cells remained viable. As the E:T ratios increased to 1:2, 1:1, 5:1, 10:1, 20:1, and 40:1, cell viability decreased in a dose-dependent manner. At the highest E:T ratios of 20:1 and 40:1, the viability of HCT-15 cells was 8.98 ± 0.61% and 8.08 ± 2.00% respectively. These results indicate a dose-dependent reduction in tumor cell viability in the presence of CIK cells. As the MTT assay reflects metabolic activity rather than direct cell death, these findings should be interpreted as an indirect measure of cytotoxicity.
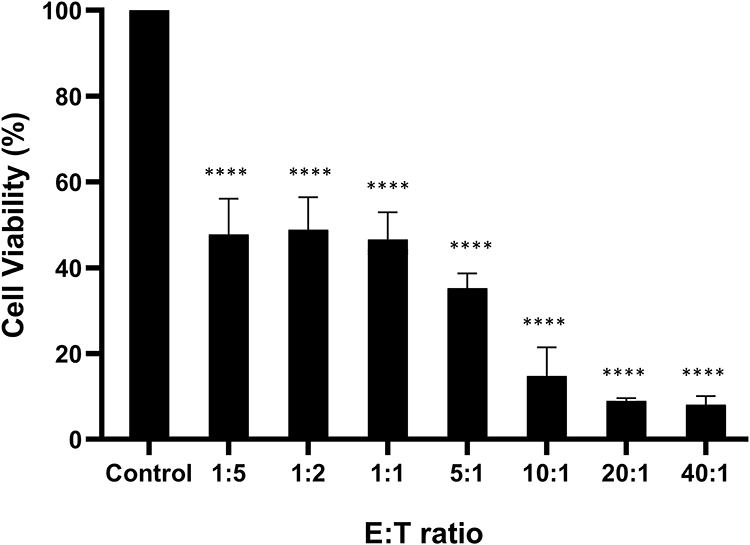 |
Figure 2 In vitro cytotoxicity of CIK cells against HCT-15 CRC cells at various E:T ratios. The percentage of viable tumor cells was normalized to their respective control. Data are presented as mean ± SD from technical triplicates. Statistical analysis was assessed using one-way ANOVA. ****p < 0.0001. |
The Viability of HCT-15 Cells Co-Cultured with CIK Cells from Donor 2 with or Without 5-FU Co-Treatment
To evaluate the combined effect of chemotherapy, HCT-15 cells were pre-treated with the antineoplastic agent, 5-FU, commonly used in the treatment of CRC patients, for 24 hours, followed by co-incubation with CIK cells from healthy donor 2 at various E:T ratios. To determine the optimal dose of 5-FU for combination with CIK cells, HCT-15 cells were treated with varying concentrations of 5-FU (ranging from 0 to 0.150 mg/mL) for 24 hours. Based on these preliminary results (results not shown here), three concentrations, 0.050, 0.075, and 0.100 mg/mL, were selected for co-treatment with CIK cells. The MTT assay revealed that 5-FU alone reduced cell viability at all three concentrations after 72 hours compared to control (Figure 3A). Co-culture with CIK cells also reduced cell viability in a dose-dependent manner (Figure 3B). At the lowest E:T ratio of 1:5, HCT-15 cell viability was 60.05 ± 3.57%, which decreased to 38.46 ± 9.24% at 20:1. The combination of 5-FU and CIK cells resulted in a further reduction in cell viability compared to either treatment alone (Figure 3C). For example, pre-treatment with 0.050 mg/mL 5-FU followed by CIK cells co-culture reduced viability to 22.63 ± 3.06%, 11.47 ± 6.54%, 9.30 ± 4.76%, and 1.02 ± 1.01% at E:T ratios of 1:5, 1:1, 10:1, and 20:1, respectively. Similar trends were observed with other 5-FU dosages co-cultured in CIK cells. These findings suggest an enhanced cytotoxic effect of the combination treatment compared to either treatment alone.
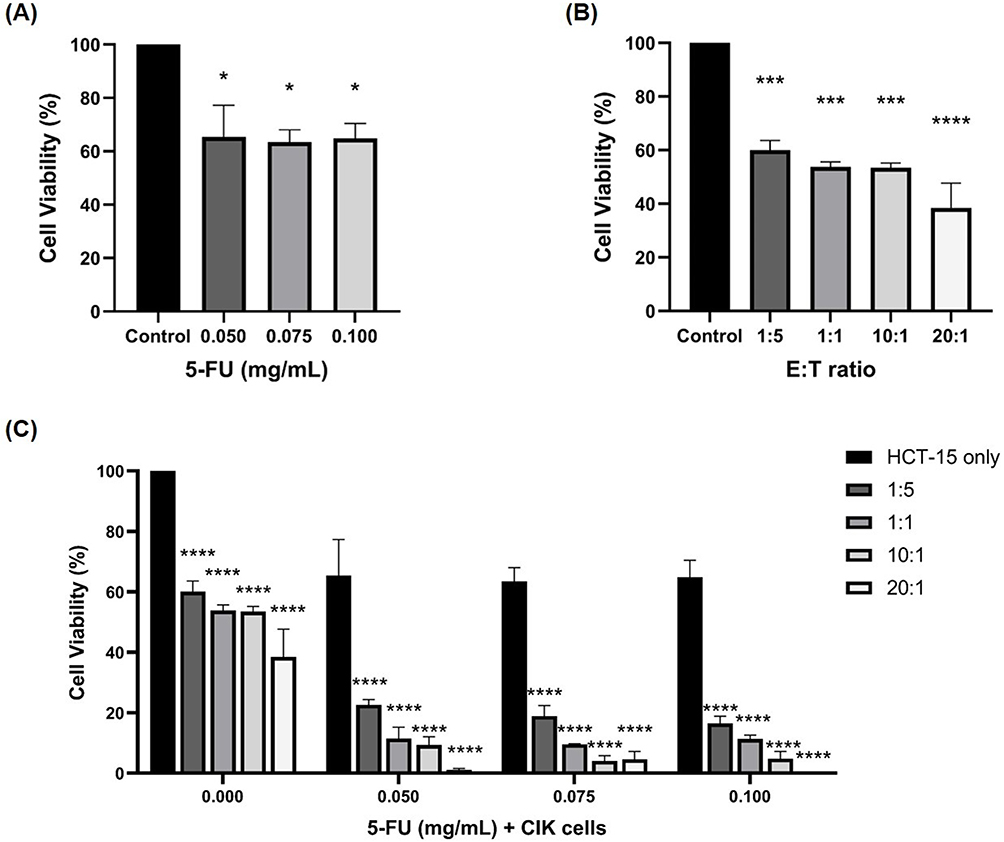 |
Figure 3 Cytotoxic effects of 5-FU and CIK cells on HCT-15 cells. HCT-15 were pre-treated with 5-FU (0.050, 0.075, and 0.100 mg/mL) for 24 hours, followed by co-culture with CIK cells for 72 hours at E:T ratios of 1:5, 1:1, 10:1, and 20:1. (A) Cytotoxic effect of 5-FU on HCT-15 cells. (B) Cytotoxic effect of CIK cells at various E:T ratios on HCT-15 cells. (C) Combined effects of 5-FU and CIK cells on HCT-15 cells. Statistical analyses for (A and B) were performed using one-way ANOVA, while (C) was analyzed using two-way ANOVA. Data are presented as mean ± SD from technical triplicates. *p < 0.05, ***p < 0.001, ****p < 0.0001, compared with the control group. |
The Viability of HCT-15 Cells Co-Cultured with CIK Cells from Donor 3 and CRC Patient
To compare the cytotoxic activities, CIK cells derived from CRC patient and healthy donor 3 were co-cultured with HCT-15 cells at various E:T ratios (1:5, 1:2, 1:1, 5:1, 10:1, 20:1, and 40:1). As shown in Figure 4, CIK cells derived from healthy donor showed higher cytotoxicity at an E:T ratio of 5:1, corresponding to approximately 70% estimated cell killing, compared to approximately 25% estimated cell killing by CRC patient-derived CIK cells. As the E:T ratio increased, the cytotoxic activity of both cell populations improved, with near-complete elimination observed at 20:1 and 40:1. The HCT-15 cell viability at an E:T ratio of 20:1 when co-cultured with CIK from donor 3 and CRC patient was 2.60 ± 5.86% and 0.0% respectively. At an E:T ratio of 40:1, HCT-15 viability when co-cultured with CIK from donor 3 and CRC patient was 0.0%. These findings suggest that CIK cells derived from metastatic CRC patient also displayed cytotoxic functionality at high E:T ratios comparable to CIK from donor 3, although cytotoxic effects may be lower at low E:T ratios.
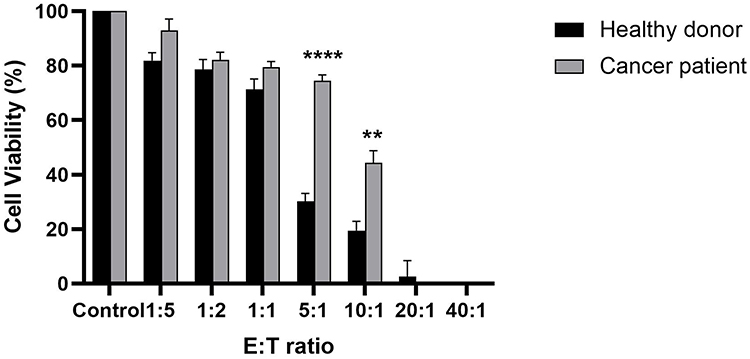 |
Figure 4 In vitro cytotoxic activity of CIK cells against HCT-15 cells. The cytotoxic effect of CIK cells expanded from healthy donor 3 versus CRC patient was assessed at E:T ratios of 1:5, 1:2, 1:1, 5:1, 10:1, 20:1 and 40:1. Cancer cells without CIK cells served as normalizing controls for both healthy donor and cancer patient. Statistical analysis was assessed using independent t-test. Data are presented as mean ± SD from technical triplicates. **p < 0.01, ****p < 0.0001, compared to the respective control. |
Cytokines and Growth Factors Profiles of CIK Cells from Donor 3, HCT-15 Cells, and Co-Culture Conditions
As shown in Figure 2, CIK cells induced effective cytotoxicity at E:T ratios of 1:1 and 10:1, corresponding to estimated killing of > 50% and > 80% of HCT-15 cells, respectively, after 72 hours of incubation. Based on these observations, cytokine profiling was performed to further explore the potential mechanisms underlying CIK-mediated effects in the co-culture system. Cytokines released from both CIK cells derived from healthy donor 3 and HCT-15 cells were therefore compared at E:T ratios of 1:1 and 10:1.
To characterize the secreted factors in the conditioned media of CIK cells in response to HCT-15 cells, the cytokines, chemokines, and growth factors were profiled (Figure 5 and Supplementary Figure 2). The expression levels of 105 cytokines were visualized in a heat map (Figure 6A), and the corresponding pixel density values are provided in Supplementary Table 1. Cytokines with pixel density greater than 300,000 were considered highly expressed. In this study, 15 cytokines related to inflammation, immune cell regulation, proliferation, and chemotaxis were highly expressed in the CIK cells. In contrast, HCT-15 cells exhibited 16 elevated cytokines associated with cellular, inflammatory, and immune responses, and signal transduction and signaling pathways (Figure 6B and Supplementary Table 2). Among these 105 cytokines, nine markers were commonly expressed in both CIK cells and HCT-15 cells, primarily associated with immune regulation, inflammatory responses, and cellular responses (Figure 6B).
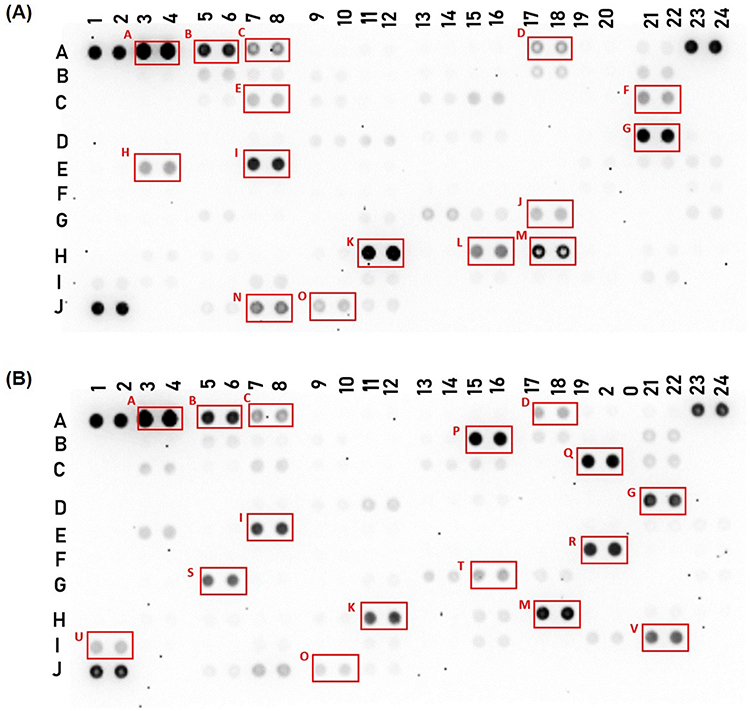 |
Figure 5 The relative levels of cytokines in the conditioned media of (A) CIK cells from donor 3 and (B) HCT-15 cells were visualized using a human cytokine antibody array (Proteome Profiler Human XL Cytokine Array Kit, R&D Systems). The following highly expressed markers in CIK cells and HCT-15 cells culture are indicated by the boxes on the membranes: A: Adiponectin; B: Apolipoprotein A-I; C: Angiogenin; D: Complement component C5/C5a; E: Fas ligand (FasL); F: Granulocyte macrophage colony-stimulating factor (GM-CSF); G: Interleukin-2 (IL-2); H: Interleukin-5 (IL-5); I: Interleukin-8 (IL-8); J: Macrophage inflammatory protein-1 alpha/Macrophage inflammatory protein-1 beta (MIP-1α/MIP-1β); K: Platelet factor 4 (PF4); L: Regulated upon Activation, Normal T Cell Expressed and Presumably Secreted (RANTES); M: Retinol binding protein 4 (RBP-4); N: Cluster of differentiation 31 (CD31); O: T-cell immunoglobulin and mucin domain 3 (TIM-3); P: Dickkopf-related protein 1 (DKK-1); Q: Growth differentiation factor 15 (GDF-15); R: Human interferon-inducible protein 10 (IP-10); S: Lipocalin-2; T: Monokine induced by gamma interferon (MIG); U: Serpin E1; V: Vascular endothelial growth factor (VEGF). Differences in spot intensity reflect relative cytokine expression levels, where darker spots indicate higher expression and lighter spots indicate lower expression. Background signal was subtracted during image analysis. |
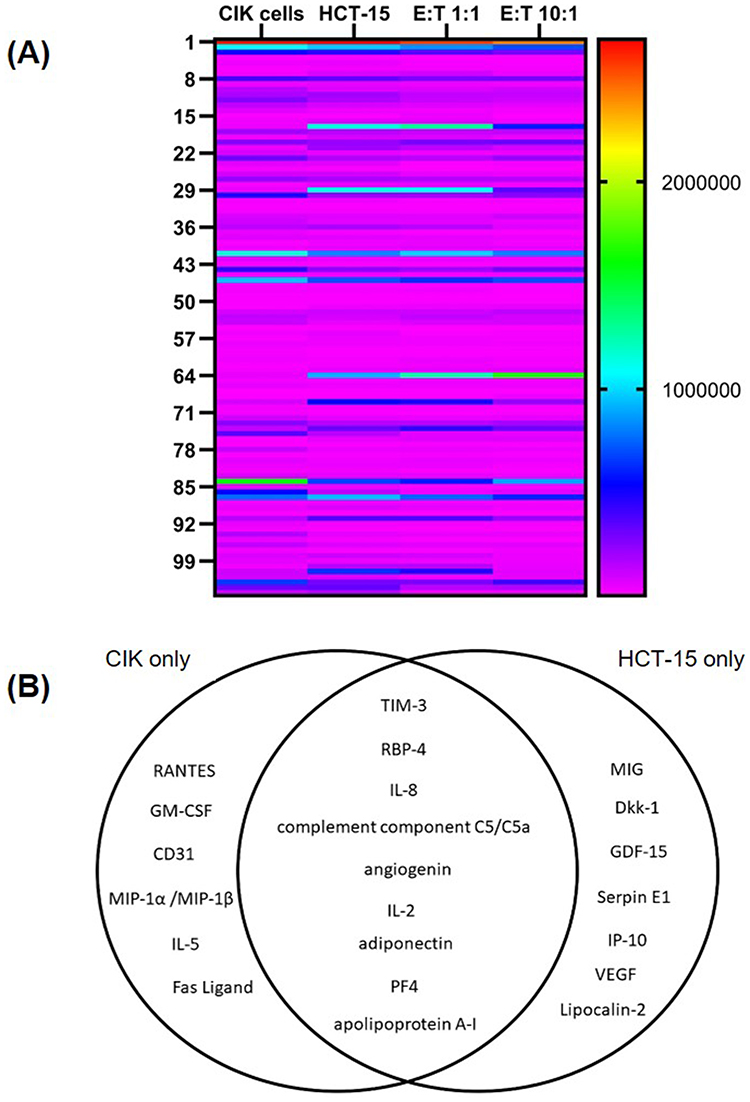 |
Figure 6 Differentially expressed cytokines and growth factors in the conditioned medium of CIK cells from donor 3 alone, HCT-15 cells, and co-culture of CIK cells and HCT-15 cells at E:T ratios of 1:1 and 10:1. (A) Heatmap of 105 cytokines and chemokines. (B) Venn diagram depicting markers that were highly expressed (pixel density >300,000) in CIK cells and HCT-15 cells. |
Cellular Response Associated with Changes in Cytokines and Growth Factors Exhibited by CIK Cells from Donor 3 and HCT-15 Cells
To characterize the secreted factors in the conditioned media, samples from CIK cells derived from healthy donor 3, HCT-15 cells and their co-culture were analyzed. A total of 36 cytokines and growth factors were observed to be differentially regulated in co-culture conditions of CIK cells and HCT-15 cells (Supplementary Table 1). GO enrichment analysis (DAVID v6.7) identified that inflammatory response, immune response, chemotaxis, and cell-cell signaling were the most enriched GO terms associated with the factors secreted by CIK cells in response to HCT-15 cells (Table 3). This suggests that the co-culture with HCT-15 cells may influence the cytokine secretion profile of CIK cells, particularly in terms of immunomodulation.
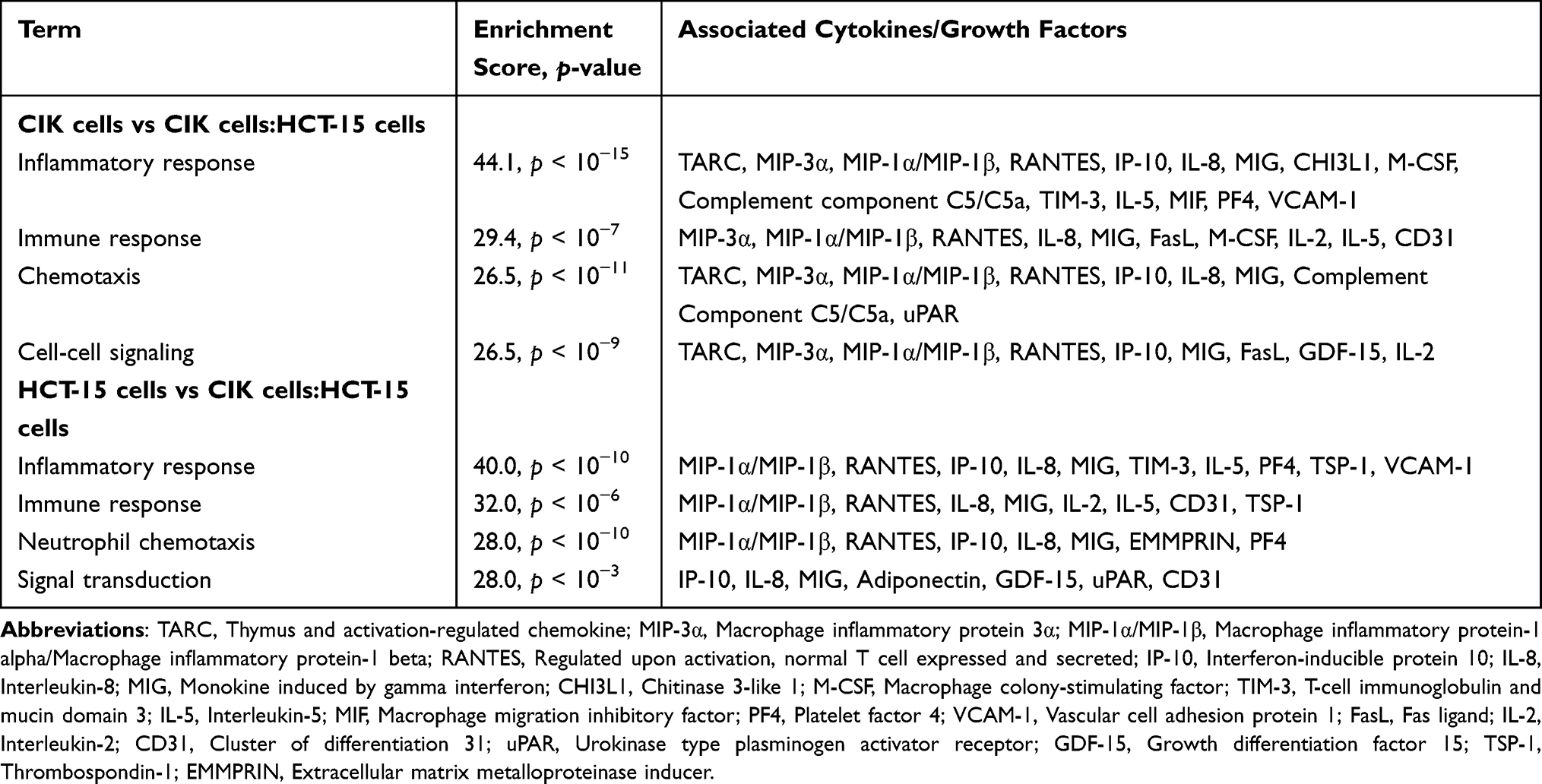 |
Table 3 Functional Annotation of the Significantly Regulated Cytokines and Growth Factors in CIK Cells or HCT-15 Cells Following Co-Culture |
Specifically, thymus and activation-regulated chemokine (TARC/CCL17), macrophage inflammatory protein (MIP)-1α/MIP-1β (CCL3/CCL4), regulated upon activation, normal T cell expressed and secreted (RANTES/CCL5), IL-2, IL-5, IL-8 (CXCL8), chitinase-3-like protein 1 (CHI3L1), macrophage colony-stimulating factor (M-CSF), complement component C5/C5a, T-cell immunoglobulin and mucin domain 3 (TIM-3), macrophage migration inhibitory factor (MIF), platelet factor 4 (PF4/CXCL4), vascular cell adhesion molecule 1 (VCAM-1), FasL, and CD31 were reduced, while MIP-3α (CCL20), monokine induced by IFN-γ (MIG/CXCL9), IFN-γ-induced protein 10 kDa (IP-10/CXCL10), urokinase plasminogen activator surface receptor (uPAR), and growth differentiation factor 15 (GDF-15) were increased.
Conversely, when HCT-15 were exposed to CIK cells, IP-10 and extracellular matrix metalloproteinase inducer (EMMPRIN/CD147) were increased, while MIP-1α/MIP-1β, RANTES, TIM-3, thrombospondin-1 (TSP-1), VCAM-1, and adiponectin were decreased.
Compared with HCT-15 cells alone, at an E:T ratio of 1:1, MIG and IL-2 levels increased, while IL-8 levels declined. At an E:T ratio of 10:1, IL-5 and CD31 levels increased, whereas GDF-15 and uPAR levels decreased (Supplementary Table 1). These factors were associated with processes such as neutrophil chemotaxis and signal transduction, which are involved in inflammatory and immune responses (Table 3).
The relationships between the identified factors were assessed using STRING and a PPI network was constructed (Figure 7). Upon stimulation by HCT-15 cells, CIK cells released factors associated with positive regulation of mononuclear cell proliferation, immune processes, leukocyte migration, and negative and positive regulation of cellular process. In contrast, co-culturing with CIK cells led HCT-15 cells to be associated with positive regulation of the immune system, mitogen-activated protein kinase (MAPK) cascade, cell proliferation, and cell communication. These findings align with the DAVID analysis and suggest a role for these regulated cytokines and growth factors in modulating cell-to-cell interactions, inflammation, and immune responses during the interaction between CIK cells and HCT-15 cells.
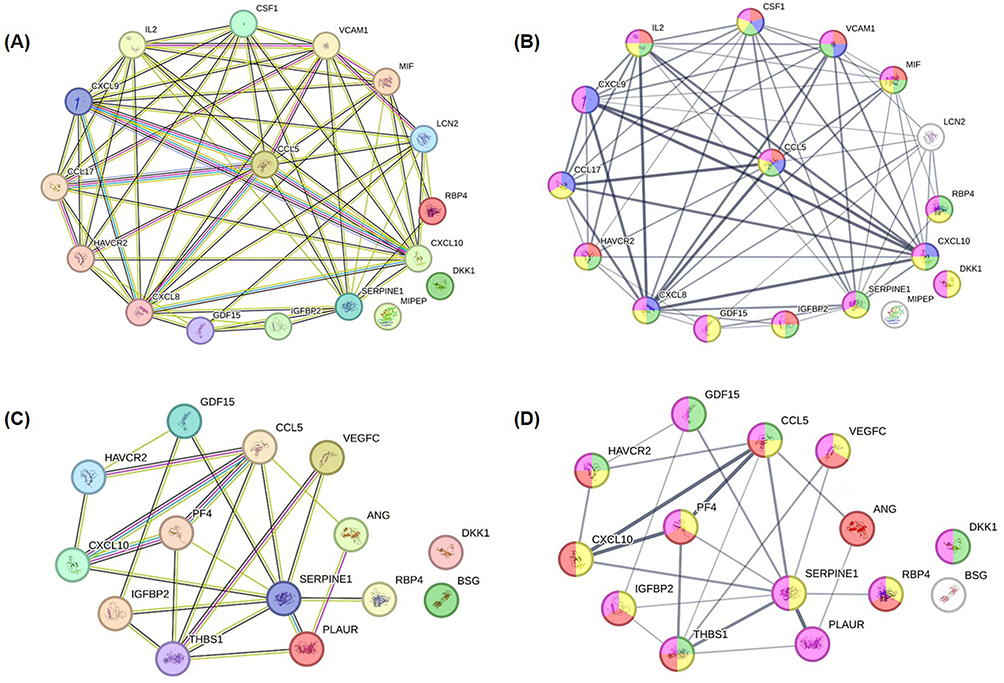 |
Figure 7 Distinct protein-protein interaction among the significantly regulated cytokines and growth factors within (A and B) CIK cells from donor 3 or (C and D) HCT-15 cells during co-culture, as elucidated by STRING. (A) Functional and physical associations of cytokines significantly regulated within CIK cells. (B) Association strengths of these regulated factors in different biological processes. Red: positive regulation of mononuclear cell proliferation; Green: positive regulation of immune system process; Blue: leukocyte migration, Yellow: negative regulation of cellular process; Pink: positive regulation of cellular process. (C) Functional and physical associations of cytokines significantly regulated within HCT-15 cells. (D) Association strengths of these regulated factors in different biological processes. Yellow: positive regulation of immune system process; Green: positive regulation of the MAPK cascade; Red: regulation of cell population proliferation; Pink: regulation of cell communication. The networks were generated using the STRING database. Nodes represent proteins, and edges indicate predicted or known associations. These interactions reflect database-derived relationships and suggest potential functional connectivity but do not imply direct causal interactions. |
Regulation of Tumor Markers in CIK Cells from Donor 3 and HCT-15 Cells
The biological roles of the regulated factors in HCT-15 cells were further investigated by comparing HCT-15 cells cultured alone with HCT-15 cells co-cultured with CIK cells derived from healthy donor 3 at E:T ratios of 1:1 and 10:1. Among the regulated cytokines and growth factors (Supplementary Table 1), 15 were identified as playing key roles in modulating tumor growth or suppression. These factors were categorized into three groups based on their functions: antitumor effects, immunosuppression, and cell proliferation and metastasis (Figure 8). Compared to HCT-15 cells alone, antitumor-related markers such as IP-10, MIG, and EMMPRIN showed increased expression in HCT-15 cells following co-culture with CIK cells, whereas markers associated with immunosuppression, cell proliferation, and metastasis were reduced. Notably, as the number of CIK cells increased, there was a further reduction in the levels of TSP-1, Dickkopf-related protein 1 (DKK-1), retinol binding protein 4 (RBP-4), uPAR, vascular endothelial growth factor (VEGF), insulin-like growth factor binding protein-2 (IGFBP-2), Serpin E1, and GDF-15 (Figure 8). These observations suggest that CIK cells may influence tumor-associated signaling pathways.
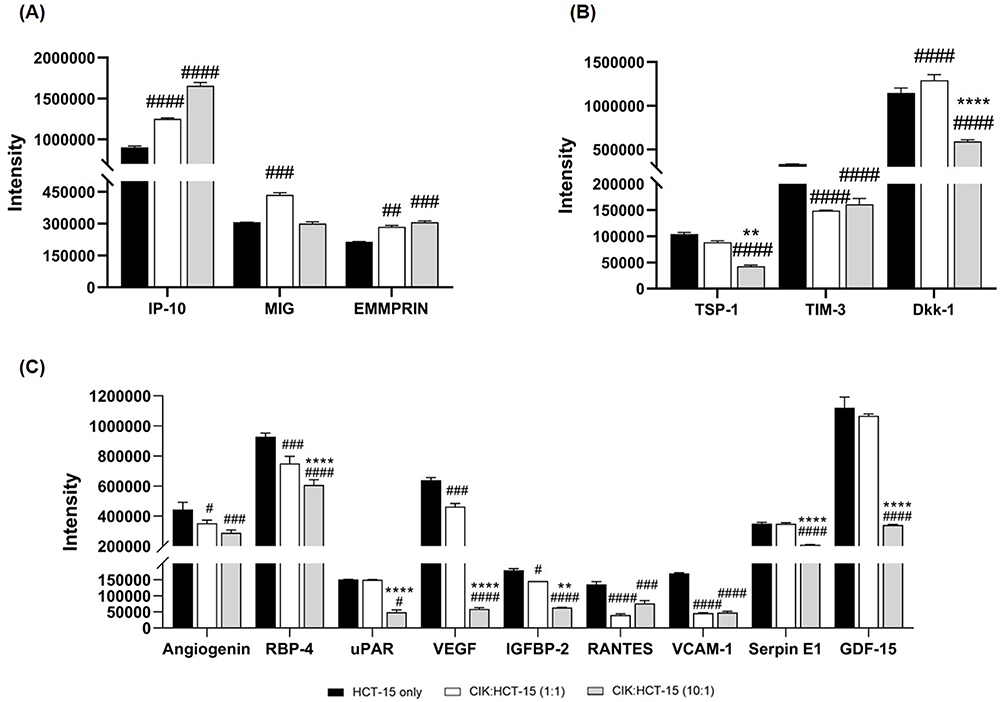 |
Figure 8 Levels of cytokines and growth factors in HCT-15 cells alone and co-cultured with CIK cells from donor 3 at E:T ratios of 1:1 and 10:1. The factors are grouped based on their functions into (A) antitumor, (B) immunosuppression, and (C) cell proliferation and metastasis. The pixel intensity is shown as mean ± SD from technical triplicates. Two-way ANOVA followed by post-hoc Tukey analysis was performed. #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 compared to HCT-15 cells; **p < 0.01, ****p < 0.0001 compared to CIK:HCT-15 at an E:T ratio of 1:1. |
Discussion
This exploratory study demonstrated that CIK cells can be successfully generated from both healthy donors and a metastatic CRC patient, exhibiting cytotoxic activity against HCT-15 cells and modulating tumor-associated cytokine profiles.
CIK cells generated in this study exhibited established phenotypic characteristics reported in previous studies, including enrichment of CD3⁺CD8⁺ T cells and CD3⁺CD56⁺ NKT cells.5,6 The reduction in CD3−CD56⁺ NK cells observed during CIK expansion is consistent with previous findings and may reflect selective proliferation of CD3⁺CD8⁺ T and CD3⁺CD56⁺ NKT subsets under cytokine-driven culture conditions. The generation of CIK cells from several CRC patients with liver metastases also showed increased yields of CD3⁺CD8⁺ T cells and CD3⁺CD56⁺ NKT cells, while the CD3−CD56⁺ NK cell subset was reduced.7 Our results showed that despite donor variability, CIK cells derived from a metastatic CRC patient exhibited comparable expansion and cytotoxic activity to those from healthy donors, supporting their potential applicability in autologous settings. Although reduced cytotoxicity was observed at lower effector-to-target (E:T) ratios, increased effector cell density enhanced tumor cell killing, which aligns with previous findings showing that cytotoxic T cell density influences tumor elimination efficiency.23 Additionally, higher NKT cell levels have been associated with improved antitumor immunity and clinical outcomes, including a subset known as invariant NKT (iNKT) cells, which have been linked to improved survival and tumor control in head and neck squamous cell carcinoma, as well as longer disease-free survival in colorectal cancer.24,25
Clinical studies have demonstrated that CIK cell-based immunotherapy is associated with antitumor activity across multiple malignancies, including gastrointestinal cancers.14,26,27 In metastatic CRC, the combination of CIK cells with chemotherapy has been associated with improved overall and progression-free survival compared to chemotherapy alone.26 Similar observations have been reported in gastric cancer, with potential survival benefit supporting the feasibility of integrating CIK cells with conventional treatment approaches.27 In this study, pre-treatment with 5-FU was associated with enhanced reduction in HCT-15 cell viability when combined with CIK cells. Similar findings have been reported where CIK cells enhance the effects of chemotherapeutic agents, potentially through cytokine-mediated immune activation or increased tumor susceptibility.28,29
Cytokine profiling in this study revealed dynamic modulation of factors involved in immune regulation, chemotaxis, and inflammation following co-culture with HCT-15 cells. Several cytokines associated with immune cell recruitment and activation, including MIP-1β, GM-CSF, and IL-2, were modulated under co-culture conditions. The observed changes in these cytokines may reflect an immunologically responsive environment induced by CIK-tumor interaction, consistent with proposed mechanisms of CIK-mediated cytotoxicity and immune activation (Supplementary Figure 3). Previous studies have shown that MIP-1β chemoattracts CD8⁺ T cells, CD4⁺ T cells, and NK cells and promotes antitumor immune responses, while GM-CSF regulates recruitment and activation of immune cells such as dendritic cells and lymphocytes, and IL-2 plays a central role in T cell proliferation and activation in cancer immunotherapy.30–32 In addition, interferon-inducible chemokines such as MIG (CXCL9) and IP-10 (CXCL10) were increased following co-culture in this study. The upregulation of these chemokines may be associated with enhanced immune-related activity, although functional validation was not performed.33,34
Notably, EMMPRIN (CD147) expression was increased following co-culture. This finding suggests a potential involvement of CD147 in tumor-immune interactions within the experimental system. CD147 is a multifunctional glycoprotein involved in T cell activation and immune regulation.35 However, it has also been pathologically associated with tumor-promoting functions in other contexts, including enhancement of angiogenesis through VEGF regulation, promotion of tumor invasion and metastasis, and suppression of CD8⁺ T cell–mediated antitumor activity.36–38 Given these context-dependent roles, the increase in CD147 observed in this study should be interpreted cautiously, as its functional implication in CRC cannot be determined from expression data alone.
In addition, several factors associated with tumor progression, immune evasion, and metastasis, including TSP-1, DKK-1, TIM-3, uPAR, IGFBP-2, and GDF-15, were reduced under co-culture conditions in this study. The downregulation of these molecules may suggest a shift toward a less tumor-promoting profile within the experimental system, as these factors are implicated in tumor progression, angiogenesis, and immune evasion pathways (Supplementary Figure 4). In relation to our findings, the observed decrease in TSP-1 and DKK-1 may be associated with a less immunosuppressive environment, as these factors have been reported to regulate immune responses and impair CD8⁺ cell-mediated antitumor activity.39,40 Similarly, the reduction in TIM-3 expression may reflect a decrease in signals linked to tumor progression and metastatic potential in CRC.41 The decreased levels of uPAR and GDF-15 observed in this study may also be associated with reduced tumor cell migration, invasion, and epithelial-mesenchymal transition.42,43 In addition, the reduction in IGFBP-2 may suggest a potential decrease in immunosuppressive signaling, as IGFBP-2 has been shown to promote regulatory T cell differentiation.44
Collectively, these findings suggest that CIK cells may influence multiple components of the tumor-associated cytokine network observed in this in vitro system. A schematic summary of CIK-mediated antitumor activity and tumor-associated mechanisms is provided in Supplementary Figures 3 and 4. However, given the exploratory design, limited sample size, and absence of functional validation, these observations should be interpreted as preliminary. In addition, the cytotoxicity assessment in this study was based on the MTT assay, which reflects metabolic activity rather than direct tumor cell death, and may be influenced by effector cell metabolism in co-culture systems. Future studies incorporating larger patient cohorts, more physiologically relevant models, and direct cytotoxicity assays such as flow cytometry-based killing assays (eg, Annexin V/PI or CFSE-based approaches) are required to further elucidate the mechanisms underlying CIK-tumor interactions.
Conclusion
In conclusion, this exploratory study demonstrates that functional CIK cells can be generated from both healthy donors and a metastatic CRC patient, with comparable phenotypic characteristics and cytotoxic activity against HCT-15 cells. The combination of CIK cells with 5-FU was associated with enhanced reduction in tumor cell viability compared to either treatment alone. Cytokine profiling suggests that CIK cells may modulate tumor-associated immune responses through changes in pro-inflammatory and immunoregulatory factors. Notably, factors associated with immune activation were increased, whereas several tumor-associated cytokines were reduced, suggesting that CIK cells may influence the tumor-associated immune environment. These findings provide preliminary insight into cytokine-mediated interactions between CIK cells and HCT-15 cells.
Data Sharing Statement
The data analyzed to support the findings of this study are available from the corresponding author upon request.
Ethics Statement
This study was conducted in accordance with the ethical standards of the Declaration of Helsinki and relevant national guidelines. Ethical approval was obtained from the Medical Research and Ethics Committee (MREC), Ministry of Health Malaysia (MOH), under approval number: 24-03956-VSD. All participants were fully informed about the nature and purpose of the study, and written informed consent was obtained from each participant included in the study.
Acknowledgments
The authors would like to express their sincere gratitude to the Postgraduate Laboratory, M Kandiah Faculty of Medicine and Health Sciences, University Tunku Abdul Rahman, for providing access to essential laboratory instruments. Additionally, we acknowledge the R&D Laboratory, Cryocord Sdn Bhd, Selangor, Malaysia for providing research conduct support. We extend our thanks to the staff at CryoCord Sdn Bhd, Selangor, Malaysia for culturing CIK cells under good manufacturing practice (GMP) conditions. We are deeply grateful to Ms. Tai Lihui and Dr. Looi Chin King for their invaluable contributions in conducting the experiments and drafting the manuscript, respectively. Lastly, we extend our heartfelt appreciation to all the patients who participated in this study.
Author Contributions
All authors made a significant contribution to the work reported, including in the conception, study design, execution, acquisition of data, and analysis and interpretation; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was sponsored and provided by Cytopeutics Unconditional Research Grant POD0008/PreCP/R-1.
Disclosure
Sze-Piaw Chin advises Cytopeutics Sdn Bhd on regulatory, clinical, and research activities. He also reports that he sits on the medical and research advisory board of Cytopeutics which owns the patent (registered in Malaysia) for the method of culture of cytokine-induced killer cells. However the entire research has been carried out independently. Soon-Keng Cheong sits on Cytopeutics Sdn Bhd medical advisory board. Natasha Najwa Nor Arfuzir and Vivian Wai-Ping Kong have no conflicts of interest to declare in this work.
References
1. Favoriti P, Carbone G, Greco M, Pirozzi F, Pirozzi REM, Corcione F. Worldwide burden of colorectal cancer: a review. Updates Surg. 2016;68(1):7–16. doi:10.1007/s13304-016-0359-y
2. Blondy S, David V, Verdier M, Mathonnet M, Perraud A, Christou N. 5-Fluorouracil resistance mechanisms in colorectal cancer: from classical pathways to promising processes. Cancer Sci. 2020;111(9):3142–3154. doi:10.1111/cas.14532
3. Guven DC, Kavgaci G, Erul E, et al. The efficacy of immune checkpoint inhibitors in microsatellite stable colorectal cancer: a systematic review. Oncologist. 2024;29(5):580–600.
4. Ros J, Balconi F, Baraibar I, et al. Advances in immune checkpoint inhibitor combination strategies for microsatellite stable colorectal cancer. Oncology. 2023;13:1–12. doi:10.3389/fonc.2023.1112276
5. Guo Y, Han W. Cytokine-induced killer (CIK) cells: from basic research to clinical translation. Chin J Cancer. 2015;34(3):1–9. doi:10.1186/s40880-015-0002-1
6. Wang Y, Bo J, Dai H, et al. CIK cells from recurrent or refractory AML patients can be efficiently expanded in vitro and used for reduction of leukemic blasts in vivo. Exp Hematol. 2013;41(3):241–252.e3. doi:10.1016/j.exphem.2012.10.014
7. Li CMY, Tomita Y, Dhakal B, et al. Generation and assessment of cytokine-induced killer cells for the treatment of colorectal cancer liver metastases. Cancer Immunol Immunother. 2024;73(1):1–16. doi:10.1007/s00262-023-03591-4
8. Yeh CL, Suk MW, Hui KM. Comparative gene expression profiling of cytokine-induced killer cells in response to acute myloid leukemic and acute lymphoblastic leukemic stimulators using oligonucleotide arrays. Exp Hematol. 2005;33(6):671–681. doi:10.1016/j.exphem.2005.03.005
9. Tsuruta D, Kobayashi H, Imanishi H, Sugawara K, Jones JCR. Laminin-332-Integrin Interaction: a Target For Cancer Therapy? Curr Med Chem. 2010;15(20):1968–1975. doi:doi:10.2174/092986708785132834
10. Yazlovitskaya EM, Viquez OM, Tu T, et al. The laminin binding α3 and α6 integrins cooperate to promote epithelial cell adhesion and growth. Matrix Biol. 2020;77:101–116. doi:10.1016/j.matbio.2018.08.010
11. Simon T, Bromberg JS. Regulation of the Immune System by Laminins. Trends Immunol. 2018;38(11):858–871. doi:10.1016/j.it.2017.06.002
12. Abdul-Rahman T, Ghosh S, Badar SM, et al. The paradoxical role of cytokines and chemokines at the tumor microenvironment: a comprehensive review. Eur J Med Res. 2024;29(1):1–19. doi:10.1186/s40001-024-01711-z
13. Man C, Li Y, Li R, et al. Clinical application of cytokine-induced killer (CIK) cell therapy in colorectal cancer: current strategies and future challenges. Cancer Treat Rev. 2024;122:102665. doi:10.1016/j.ctrv.2023.102665
14. Li CMY, Tomita Y, Dhakal B, et al. Use of cytokine-induced killer cell therapy in patients with colorectal cancer: a systematic review and meta-analysis. J Immunother Cancer. 2023;11(4):1–17. doi:10.1136/jitc-2023-006764
15. Fayyaz F, Yazdanpanah N, Rezaei N. Cytokine - induced killer cells mediated pathways in the treatment of colorectal cancer. Cell Commun Signal. 2022;1–16. doi:10.1186/s12964-022-00836-0
16. Lu D, Li T, Yang Z, Zhao X, Su Y, Nian L. DC-CIK combined with chemotherapy on the efficacy, immune function, and life quality in colorectal cancer patients after radical resection. Am J Transl Res. 2023;15(4):2793–2801.
17. Long S, Wang J, Weng F, et al. ECM1 regulates the resistance of colorectal cancer to 5-FU treatment by modulating apoptotic cell death and transition induction. Pharmacology. 2022;13:1–15. doi:10.3389/fphar.2022.1005915
18. Ahmed D, Eide PW, Eilertsen IA, et al. Epigenetic and genetic features of 24 colon cancer cell lines. Oncogenesis. 2013;2(29). doi:10.1038/oncsis.2013.35
19. Chin S, Kong W, Lian Q, Then K, Cheong S. A safe and simple approach to obtaining sufficient cytokine induced killer (CIK) cells from critically-ill cancer patients without the need for apheresis or peripheral mobilization. Cytotherapy. 2019;21(5):S88. doi:10.1016/j.jcyt.2019.03.516
20. Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57. doi:10.1038/nprot.2008.211
21. Sherman BT, Hao M, Qiu J, et al. DAVID: a web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022;50(W1):W216–W221. doi:10.1093/nar/gkac194
22. Szklarczyk D, Kirsch R, Koutrouli M, et al. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023;51(1 D):D638–D646. doi:10.1093/nar/gkac1000
23. Weigelin B, Den boer AT, Wagena E, et al. Cytotoxic T cells are able to efficiently eliminate cancer cells by additive cytotoxicity. Nat Commun. 2021;12(1):1–12. doi:10.1038/s41467-021-25282-3
24. Schneiders FL, de Bruin RCG, van den Eertwegh AJM, et al. Circulating invariant natural killer T-cell numbers predict outcome in head and neck squamous cell carcinoma: updated analysis with 10-year follow-up. J Clin Oncol. 2012;30(5):567–570. doi:10.1200/JCO.2011.38.8819
25. Al-Mterin MA, Murshed K, Elkord E. PD-1 expression, among other immune checkpoints, on tumor-infiltrating NK and NKT cells is associated with longer disease-free survival in treatment-naïve CRC patients. Cancer Immunol Immunother. 2023;72(6):1933–1939. doi:10.1007/s00262-022-03337-8
26. Pan QZ, Gu JM, Zhao JJ, et al. Retrospective analysis of the efficacy of cytokine-induced killer cell immunotherapy combined with first-line chemotherapy in patients with metastatic colorectal cancer. Clin Transl Immunol. 2020;9(2):1–13. doi:10.1002/cti2.1113
27. Ma X, Peng L, Wang J, et al. Autologous CIK cells combined with chemotherapy as the first-line treatment for locally advanced or metastatic gastric cancer is safe and feasible. Front Immunol. 2023;14:1–12. doi:10.3389/fimmu.2023.1267369
28. Zhao Q, Zhang H, Li Y, Liu J, Hu X, Fan L. Anti-tumor effects of CIK combined with oxaliplatin in human oxaliplatin-resistant gastric cancer cells in vivo and in vitro. J Exp Clin Cancer Res. 2010;29(1):1–13. doi:10.1186/1756-9966-29-118
29. Yang L, Du C, Wu L, et al. Cytokine-induced killer cells modulates resistance to cisplatin in the A549/DDP cell line. J Cancer. 2017;8(16):3287–3295. doi:10.7150/jca.19426
30. Luo X, Yu Y, Liang A, et al. Intratumoral expression of MIP-1beta induces antitumor responses in a pre-established tumor model through chemoattracting T cells and NK cells. Cell Mol Immunol. 2004;1(3):199–204. doi:10.1182/blood.v104.11.5268.5268
31. Shi Y, Liu CH, Roberts AI, et al. Granulocyte-macrophage colony-stimulating factor (GM-CSF) and T-cell responses: what we do and don’t know. Cell Res. 2006;16(2):126–133. doi:10.1038/sj.cr.7310017
32. Mortara L, Balza E, Bruno A, Poggi A, Orecchia P, Carnemolla B. Anti-cancer therapies employing IL-2 cytokine tumor targeting: contribution of innate, adaptive and immunosuppressive cells in the anti-tumor efficacy. Front Immunol. 2018;9:1–11. doi:10.3389/fimmu.2018.02905
33. Liu T, Long X, Zhang Y, Jin J, Chen L, Liu A. IP-10 enhances the amplification capacity and antitumor activity of CAR-T cells in vitro and could influence positive outcomes in MM patients treated with CAR-T cell therapy. Int Immunopharmacol. 2022;112:109253. doi:10.1016/j.intimp.2022.109253
34. Teruya-feldstein BJ, Jaffe ES, Burd PR, et al. The role of mig, the monokine induced by interferon- g, and IP-10, the interferon- g– inducible protein-10, in tissue necrosis and vascular damage associated with epstein-barr virus-positive lymphoproliferative disease. J Am Soc Hematol. 1997;90(10):4099–4105.
35. Hahn JN, Kaushik DK, Yong VW. The role of EMMPRIN in T cell biology and immunological diseases. J Leukoc Biol. 2015;98(1):33–48. doi:10.1189/jlb.3ru0215-045r
36. Chen Y, Xu J, Wu X, et al. CD147 regulates antitumor CD8+ T-cell responses to facilitate tumor-immune escape. Cell Mol Immunol. 2021;18(8):1995–2009. doi:10.1038/s41423-020-00570-y
37. Chen X, Lin J, Kanekura T, et al. A small interfering CD147-targeting RNA inhibited the proliferation, invasiveness, and metastatic activity of malignant melanoma. Cancer Res. 2006;66(23):11323–11330. doi:10.1158/0008-5472.CAN-06-1536
38. Tang Y, Nakada MT, Rafferty P, et al. Regulation of vascular endothelial growth factor expression by EMMPRIN via the PI3K-Akt signaling pathway. Mol Cancer Res. 2006;4(6):371–377. doi:10.1158/1541-7786.MCR-06-0042
39. Stirling ER, Terabe M, Wilson AS, et al. Targeting the CD47/thrombospondin-1 signaling axis regulates immune cell bioenergetics in the tumor microenvironment to potentiate antitumor immune response. J Immunother Cancer. 2022;10(11). doi:10.1136/jitc-2022-004712
40. Sui Q, Liu D, Jiang W, et al. Dickkopf 1 impairs the tumor response to PD-1 blockade by inactivating CD8+ T cells in deficient mismatch repair colorectal cancer. J Immunother Cancer. 2021;9(3):1–17. doi:10.1136/jitc-2020-001498
41. Yu M, Lu B, Liu Y, Me Y, Wang L, Zhang P. Tim-3 is upregulated in human colorectal carcinoma and associated with tumor progression. Mol Med Rep. 2017;15(2):689–695. doi:10.3892/mmr.2016.6065
42. Ding Y, Niu W, Zheng X, et al. Plasminogen activator, urokinase enhances the migration, invasion, and proliferation of colorectal cancer cells by activating the Src/ERK pathway. J Gastrointest Oncol. 2022;13(6):3100–3111. doi:10.21037/jgo-22-1215
43. Li C, Wang J, Kong J, et al. GDF15 promotes EMT and metastasis in colorectal cancer. Oncotarget. 2016;7(1):860–872. doi:10.18632/ONCOTARGET.6205
44. Sun L, Zhang Y, Yang T, Chen J, Zhang X, Liang X. IGFBP2 drives regulatory T cell differentiation through STAT3/IDO signaling pathway in pancreatic cancer. J Pers Med. 2022;12(12). doi:10.3390/jpm12122005
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Machine Learning-Derived Neddylation Gene Signature for Predicting Prognosis and Immunotherapy Benefits in Colorectal Cancer

Yang G, Xiao J, He H, Wang J, Wang Z, Jian L, Chen Q
ImmunoTargets and Therapy 2025, 14:931-952
Published Date: 25 August 2025