Back to Journals » International Journal of General Medicine » Volume 14
An Autophagy-Related Long Non-Coding RNA Prognostic Signature for Patients with Lung Squamous Carcinoma Based on Bioinformatics Analysis
Received 30 July 2021
Accepted for publication 13 September 2021
Published 12 October 2021 Volume 2021:14 Pages 6621—6637
DOI https://doi.org/10.2147/IJGM.S331327
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Boxuan Liu,1 Yun Zhao,2 Shuanying Yang1
1Department of Critical Care and Respiratory Medicine, The Second Affiliated Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, 710004, People’s Republic of China; 2Department of Cardiology, The First Affiliated Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, 710004, People’s Republic of China
Correspondence: Shuanying Yang
Department of Critical Care and Respiratory Medicine, The Second Affiliated Hospital, Xi’an Jiaotong University, 157 Xiwu Road, Xi’an, Shaanxi, 710004, People’s Republic of China
Tel +8613991392919
Email [email protected]
Purpose: Lung cancer is the most common and deadly cancer type affecting humans. Although huge progress has been made on early diagnosis and precision treatment, the overall 5 year survival rate remains low. In this study, we constructed an autophagy-related long non-coding RNA (lncRNA) prognostic signature for guiding clinical practice.
Methods: From The Cancer Genome Atlas, we retrieved mRNA and lncRNA expression matrices of patients with lung squamous carcinoma. We then established a prognostic risk model using Lasso regression and multivariate Cox regression. The model generated a risk score to differentiate high- and low-risk groups. An ROC curve and nomogram were used to visualize the predictive ability of the current signatures. Finally, we used Gene Set Enrichment Analysis to determine gene ontology and pathway enrichment.
Results: After screening 1248 autophagy-related lncRNAs, we selected seven lncRNAs (LUCAT1, AC022150.2, AL035425.3, AC138976.2, AC106786.1, GPRC5D-AS1 and AP006545.2) for our signature. Univariate (hazard ratio [HR] = 2.147, 95% confidence interval [CI]: 1.681– 2.743, P < 0.001) and multivariate (HR = 2.096, 95% CI: 1.652– 2.658, P < 0.001) Cox regression analyses revealed that the risk score is an independent predictive factor for LUSC patients. Further, areas under the receiver operating characteristic curve were 0.622, 0.699, and 0.721, respectively, for the 1 year, 3 year, and 5 year risk scores—indicating a reliable model. Selected lncRNAs were primarily enriched in autophagy, metabolism, MAPK pathway, and JAK/STAT pathway. Further drug sensitivity analysis revealed that low-risk patients were more sensitive to Cisplatin, Docetaxel, Vinblastine, and Vinorelbine. Finally, a multi-omics analysis found that lncRNA-linked proteins IKBKB and SQSTM1 were expressed at low levels and significantly correlated in tumor samples, compared with normal tissue.
Conclusion: Our prognostic model successfully predicted patient prognosis in lung cancer.
Keywords: non-small cell lung cancer, cancer therapy, non-coding RNA, cancer prognosis, The Cancer Genome Atlas
Introduction
Lung cancer is the leading cause of cancer-related deaths worldwide. In the United States alone, newly diagnosed patients exceeded 2 million a year. Non-small cell lung cancer (NSCLC), including lung adenocarcinoma and lung squamous carcinoma (LUSC), accounts for nearly 80% of all new cases.1 Novel treatment options such as targeted therapy and immune therapy have dramatically improved prognosis of lung adenocarcinoma. However, little progress has been made with respect to LUSC treatment, with most patients still subjected to traditional platinum-based chemotherapy (eg, gemcitabine and pemetrexed).2 One way to address this issue is the development of a new prognostic model that can help physicians formulate patient-specific treatment.
Autophagy is a protein degradation pathway that is vital to maintaining whole-body homeostasis in eukaryotes. This pathway plays a critical part in various disorders, including heart disease and neurological malfunction.3,4 Recent studies indicate the involvement of autophagy in tumor occurrence, maintenance, and progression; indeed, autophagy is a tumor suppressor mechanism that enhances the chemotherapy response.5,6 Several cancer-related pathways, including PI3K/AKT/mTOR, p53/DRAM, JAK-STAT, and AMPK/CaMKK exhibit a crosstalk with autophagy. Thus, several researchers have suggested that key molecules in autophagy may serve as therapeutic targets or biomarkers for lung cancer.7 For example, Xie et al found that the increased frequency of three single nucleotide polymorphisms in the autophagy gene ATG10 was significantly correlated with overall survival, while the elevated expression of ATG10 enhanced tumor proliferation and migration.8
In addition to autophagy-related molecules or pathways, long non-coding RNAs (lncRNAs) represent another promising biomarker candidate for cancer. Most of the human genome is transcribed into ncRNA, with lncRNAs (>200 nucleotide-long) being the most extensively studied due to their involvement in tumorigenesis, cell cycle, apoptosis, and chemo-resistance.9–12 The expression of certain lncRNAs has been linked to cancers. Moreover, a five-lncRNA signature was demonstrated to act as an independent survival predictor in NSCLC. Promising findings are available for LUSC as well, with lncRNAs SFTA1P and LINC00519 being associated with the occurrence of LUSC, suggesting that they have the potential to be new diagnostic/prognostic biomarkers.13 Nevertheless, none of the studies have investigated whether autophagy-related lncRNAs (ARlncRNAs) can be used to establish a risk model that predicts LUSC prognosis. Therefore, in this study, we employed mRNA and lncRNA expression profiles derived from The Cancer Genome Atlas (TCGA) data to generate a prognostic model for LUSC patients.
Materials and Methods
Raw Data Acquisition and Initial Analysis
Clinical data of patients with LUSC, along with their mRNA and lncRNA expression profiles, were downloaded from TCGA (https://portal.gdc.cancer.gov/). A Perl script was then used to extract survival time, age, sex, tumor stage, and TNM stage; these data were merged into a single file. After initial screening, patients with a short follow-up time (<30 days) were excluded, yielding a final patient count of 502.
Screening ARlncRNAs
Autophagy-related genes (File S1) were obtained from the Human Autophagy Database (HADb, http://autophagy.lu/clustering/index.html), filtered using expression matrices, and log2-transformed. Pearson’s correlations were run to remove lncRNAs with correlation coefficients |R2| > 0.3 and P < 0.001.
Identification of ARlncRNA Signature for Lung Adenocarcinoma
After initial screening, Cox regression was used to determine if the remaining ARlncRNAs were significantly correlated with prognosis. A prognostic risk score was then constructed using a least absolute shrinkage and selection operator (Lasso) regression, as follows:
with coef(lncRNAn) being lncRNA coefficients and expr(lncRNAn) being lncRNA expression.
All included samples were then allocated into a high‐risk group and a low‐risk group based on the median risk score.
Independent Survival Analysis of the Signature for LUSC Patients
To test the prognostic model, the relationship between patient survival with clinicopathological factors and risk score was assessed using univariate and multivariate Cox regression analyses. Receiver operating characteristic (ROC) curves were generated using the “survivalROC” package in R. A nomogram was generated to evaluate the efficacy of our prognostic signature to predict survival. The index of concordance (C-index) was also calculated to determine the accuracy with which our signature could predict the survival.
Drug Sensitivity Analysis
The gene expression profile of included patients and drug sensitivity data from various cancer cell lines were incorporated in a statistical model generated under “pRRophetic” package from R software.43
Functional Analysis
Gene Set Enrichment Analysis (GSEA, http://software.broadinstitute.org/gsea/index.jsp) calculates whether a set of genes differs significantly between two groups. The analysis was conducted with risk score as the phenotype to identify differentially expressed genes, along with their enriched Gene Ontology (GO) terms and KEGG pathways.
Multi-Omics Analysis of Associated Genes
Gene Expression Profiling Interactive Analysis (GEPIA, http://gepia.cancer-pku.cn/) was used to investigate the differences in mRNA expression between cancer tissues and normal tissues.14 Additionally, differential protein expression between normal tissue and tumors was investigated using the Human Protein Atlas (HPA), an open source database of immunohistochemistry results.
Statistical Analysis
All analyses were performed in R Studio (version 1.1.453). The “limma” package in R was used to differentiate survival-related lncRNAs. The lncRNA‐mRNA co-expression network was established and visualized using Cytoscape and the Sankey diagram, respectively. The “survival” package in R was used to perform the Kaplan‒Meier analysis; P-values were derived using Log rank tests. Univariate and multivariate Cox regression analyses were employed to assess the relationship between risk score and clinicopathological parameters. The “rms” package in R was used to generate the nomograms. Model fit was determined using area under the ROC curve (AUC); AUC > 0.60 was deemed a good fit (reliable predictive ability). Significance was set at P < 0.05 (two-tailed).
Results
Establishment of a Co-Expression Network
We obtained 14,142 lncRNAs from patient expression profiles. The HADb yielded 223 autophagy-related genes (File S1), and our TCGA-LUSC expression matrix extracted 210 genes. Using a cut-off of |R2| > 0.3 and P < 0.001, we constructed a co-expression network between autophagy-related genes and correlated lncRNAs. We then filtered out 1248 ARlncRNAs for further analysis.
Development of a Prognostic Risk Model Using ARlncRNAs
Using univariate Cox regression analyses, we identified 27 ARlncRNAs that were significantly correlated with patient survival in LUSC (Supplementary Table 1). Of these, 21 were favorable factors (hazard ratio, HR < 1) and 6 were harmful. Lasso regression analysis identified 18 lncRNAs that were the most highly correlated with patient survival in LUSC (Figures 1A and B).
 |
Figure 1 Selection of lncRNA using Lasso regression. (A) Lasso coefficient of the five included lncRNAs. (B) Profiles of Lasso coefficients. (C) Univariate analysis of included lncRNAs from the samples. Abbreviations: *P<0.05; **P<0.01. |
Multivariate Cox regression analysis further revealed that seven lncRNAs, ie, LUCAT1, AC022150.2, AL035425.3, AC138976.2, AC106786.1, GPRC5D-AS1, and AP006545.2, can constitute a prognostic signature for patient survival in LUSC (Figure 1C, Table 1). Kaplan‒Meier analyses revealed that LUCAT1 and AC022150.2 were negatively correlated with the survival of LUSC patients while the remainder were positively correlated (Figure 2). We then established a co-expression network between these ARlncRNAs and autophagy-related genes (Figures 3A and B, Table 2). Multivariate Cox regression analyses allowed us to generate risk scores and allocate patients into high- and low-risk groups based on the following formula: 
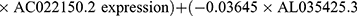
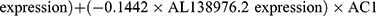

 .
.
 |
Table 1 Survival Analysis of the Seven lncRNAs Obtained Using TCGA-LUSC Data |
 |
Table 2 Correlation Coefficients of Genes Associated with the Seven lncRNAs Obtained Using TCGA-LUSC Data |
 |
Figure 2 The Kaplan-Meier survival analysis of seven included lncRNAs. LUCAT1, AC022150.2, AL035425.3, AL138976.2, AC106786.1, GPRC5D-AS1 and AP006545.2 were independent protective factors for lung squamous carcinoma. (A) Survival analysis of AC022150.2 in LUSC patients; (B) survival analysis of AC106786.1 in LUSC patients; (C) survival analysis of AL035425.3 in LUSC patients; (D) survival analysis of AL138976.2 in LUSC patients; (E) survival analysis of AP006545.2 in LUSC patients; (F) survival analysis of GPRC5D-AS1 in LUSC patients; (G) survival analysis of LUCAT1 in LUSC patients. |
 |
Figure 3 The co-expression network of autophagy-related lncRNA-mRNA and Sankey diagram. (A) mRNA–autophagy-related lncRNAs–risk type relationship showed in Sankey diagram. (B) The co-expression network visualized using Cytoscape 3.7.2 software. |
Prognostic Value of the Established Signature
The risk score cut-off was 1.138; this resulted in 234 low-risk patients and 233 high-risk patients. The high-risk group had poorer survival outcomes than the low-risk group (P < 0.001, Kaplan-Meier; Figure 4E). For the time-dependent ROC curve, the 1-year, 3-year, 5-year, and 7-year AUC were 0.622, 0.699, 0.721, and 0.733, respectively. These values indicate that this signature reliably predicted the patient survival (Figures 4A–D). Next, the risk score and survival distribution between high- and low-risk groups indicated that the former had a poorer probability of survival (Figures 5A and C). A heatmap illustrating lncRNA expression patterns in high- and low-risk patients also indicated that LUCAT1 and AC022150.2 were risk factors (Figure 5B).
 |
Figure 4 The prognostic indicators of the five autophagy-related lncRNAs signature. (A) 1-year survival ROC curve for LUSC patients. (B) 3-year survival ROC curve for LUSC patients. (C) 5-year survival ROC curve for LUSC patients. (D) 7-year survival ROC curve for LUSC patients. (E) Kaplan-Meier survival curve of the high-risk and low-risk groups for LUSC patients. |
 |
Figure 5 The analysis of the risk score from the generated risk model. (A) Expression profiles of lncRNAs in high and low risk groups; (B) the risk curve of each sample in high and low risk group; (C) the survival plot of each sample based on the risk score. |
Clinical Value of ARlncRNA Signature
Univariate Cox regression analyses revealed that age (HR = 1.020, 95% CI: 1.000–1.041, P < 0.05), tumor stage (HR = 1.311, 95% CI: 1.084–1.586, P = 0.005), tumor size (HR = 1.312, 95% CI: 1.065–1.615, P = 0.011), and the risk score (HR = 2.147, 95% CI: 1.681–2.743, P < 0.001) were significantly correlated with patient survival (Table 3, Figure 6A). Multivariate Cox regression analyses—with clinical factors (age, sex, tumor size and stage) as covariates of the risk score—identified only age (HR = 1.027, 95% CI: 1.005–1.050, P = 0.014) and risk score (HR = 2.096, 95% CI: 1.652–2.658, P < 0.001) as being significantly correlated with the overall survival (Table 4, Figure 6B). An ROC curve also indicated that risk score had the highest AUC value (0.720) among all clinicopathological parameters (Figure 6C). A detailed stratification of the clinicopathological characteristics did not reveal any significant correlations with the risk score (Table 5).
 |
Table 3 Univariate Cox Regression Analysis to Identify a Correlation Between the Clinicopathological Characteristics and Risk Scores |
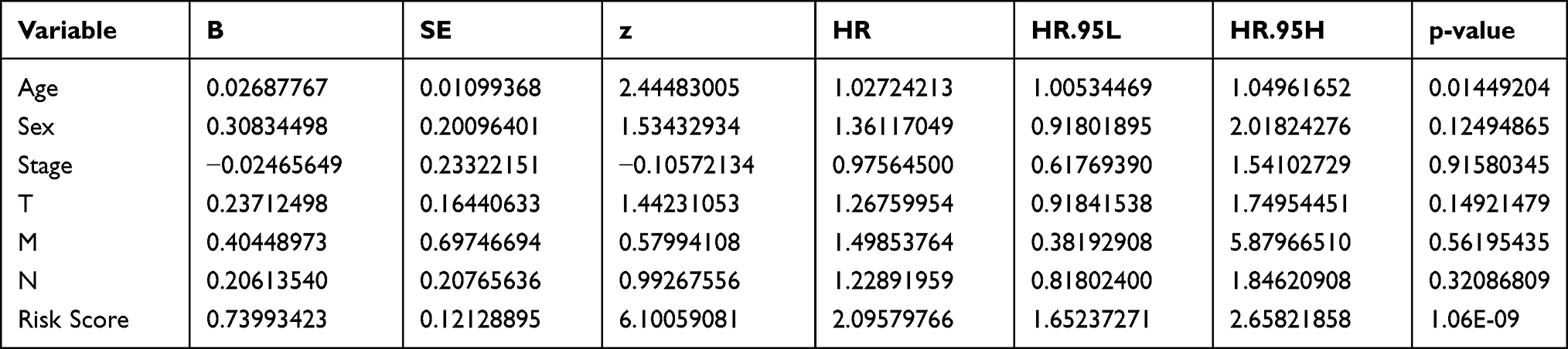 |
Table 4 Multivariate Cox Regression Analysis to Identify a Correlation Between Clinicopathological Characteristics and Risk Scores |
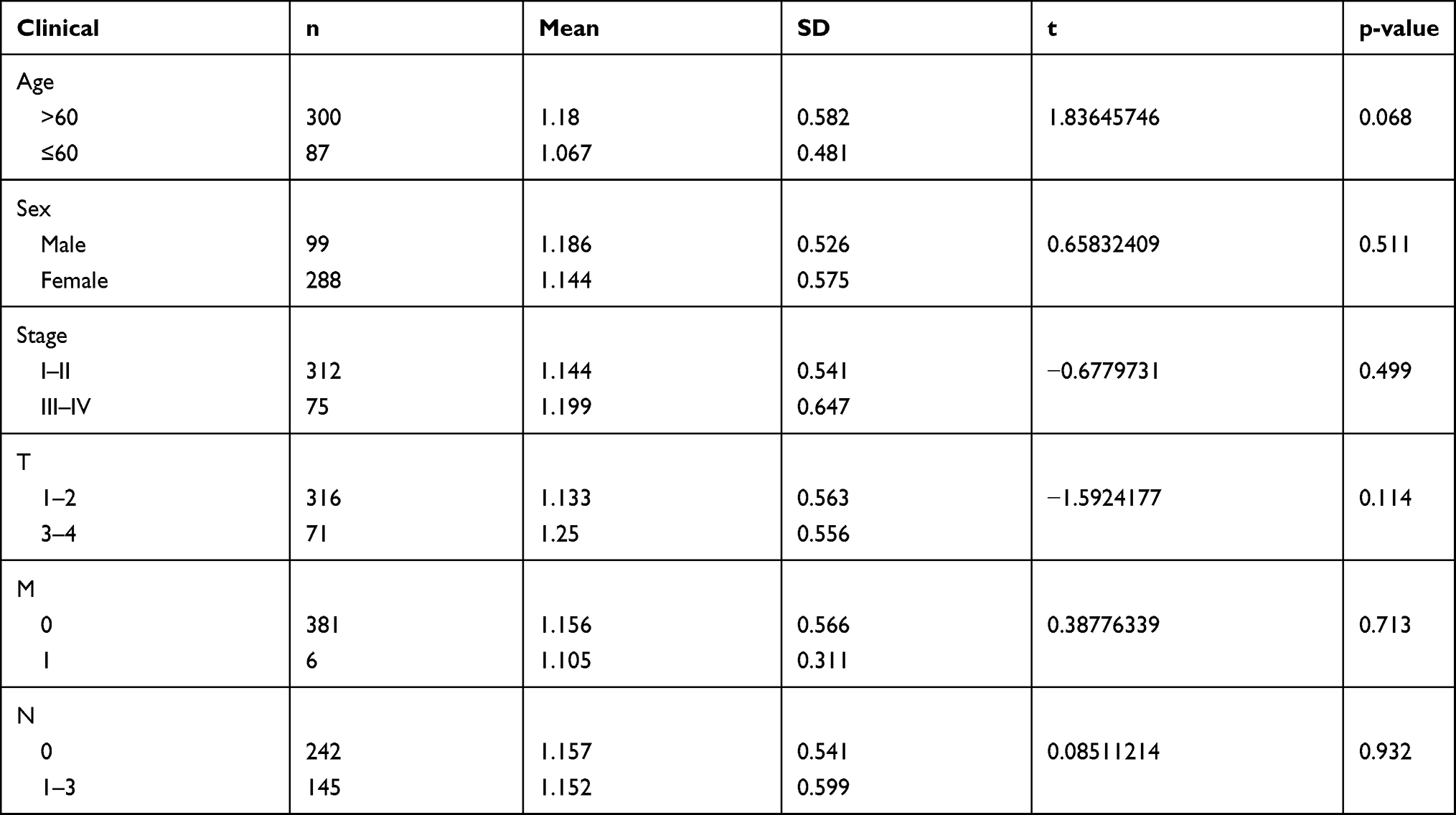 |
Table 5 Relationship Between Clinicopathological Characteristics and Risk Score |
 |
Figure 6 The evaluation of the constructed signature’s prognostic credibility in LUSC patients. (A and B) The univariate and multivariate Cox regression analysis of risk score and clinicopathological characteristics. (C) The integrated ROC curves of risk score and clinical features. |
Establishment of a Nomogram Prediction Model
A nomogram incorporates multiple clinicopathological characteristics to quantify an individual’s prognosis. Here, we employed a nomogram as a rating system to predict the 1-year, 3-year, and 5-year survival probability (Figure 7A). Calibration curves of these three durations matched well, with a C-index of 0.657 (95% CI: 0.633–0.681), indicating that the predictions of the nomogram are reliable (Figures 7B–D).
 |
Figure 7 Nomogram prediction model in LUSC patients. (A) The nomogram of 1-year, 3-year or 5-year survival predictability based on risk score, age and TNM stage. (B–D) The calibration plots for predicting 1-year, 3-year or 5-year survival. |
Drug Sensitivity Analysis of ARlncRNAs
To further explore the usage of our signature in clinical setting, we applied durg sensitivity analysis using “pRRophetic” package in software R to predict the IC50 of the conventional chemotherapy agent between high-risk and low-risk group. We found that low-risk group patients are more sensitive in treating with Cisplatin, Docetaxel, Vinblastine, and Vinorelbine (Figure 8A, B, E and F), indicating a better drug response in low-risk group. As for Gemcitabine and Paclitaxel, there were no significant difference been found (Figures 8C and D).
 |
Figure 8 Drug sensitivity analysis between high-risk and low-risk group. (A) The IC50 of Cisplatin between the two groups; (B) The IC50 of Docetaxel between the two groups; (C) The IC50 of Gemcitabine between the two groups; (D) The IC50 of Paclitaxel between the two groups; (E) The IC50 of Vinblastine between the two groups; (F) The IC50 of Vinorelbine between the two groups. **P<0.01; ***P<0.001. Abbreviation: ns, not significant. |
Functional Analysis
The KEGG pathway analysis identified 51 gene sets in the high-risk group that were significantly associated (P < 0.01) with cancer-related pathways, ie, JAK/STAT signaling, MAPK signaling, cytokine receptor, chemokine signaling, focal adhesion, and cell adhesion (Figures 9A and B, Supplementary Table 2). In the low-risk group, enriched genes were mainly associated with RNA synthesis, RNA degradation, and neurodegenerative disease (eg, Parkinson’s, Huntington’s, and Alzheimer’s) pathways. Gene Ontology analysis showed that the high-risk group (Figure 9C) was significantly associated with immune response (NES = 2.08, P < 0.0001), epithelial cell proliferation (NES = 2.21, P < 0.0001), and endothelial cell proliferation (NES = 2.69, P < 0.0001). Enriched GO terms in the low-risk group (Figure 9D) were RNA methylation (NES = −2.23, P < 0.0001) and protein mannosylation (NES = −1.97, P < 0.0001) pathways. Most of the enriched GO terms and KEGG pathways were closely related to the occurrence and development of lung adenocarcinoma, indicating that the five lncRNAs are involved in functions related to lung cancer.
 |
Figure 9 The functional enrichment analysis based on autophagy-related lncRNAs. (A and B) Upregulated KEGG pathway in high risk group and low risk group; (C and D) Upregulated Gene Ontology terms in high risk group and low risk group. |
Multi-Omics Validation of the lncRNA Signature and Correlated Genes
Correlation analysis (Supplementary Table 3) revealed that the genes EIF4EBP1 (AP006545.2), ATG4A (AL035425.3), CAPN10 (AC022150.2), IKBKB (AC106786.1), ATG2B (AL138976.2), GAPDH (GPRC5D-AS1), and SQSTM1 (LUCAT1) were most highly correlated with selected lncRNAs (in parentheses) (Table 2). Because only AC106786.1 and LUCAT1 were available in GEPIA, we explored the mRNA and protein expression of these two pairs (IKBKB-AC106786.1 and SQSTM1-LUCAT1), along with their clinical significance.
GEPIA showed that the transcript levels of IKBKB were significantly lower in LUSC patients, whereas the transcript levels of AC106786.1 were slightly higher (Figures 10A and B). Both SQSTM1 and LUCAT1 were expressed at low levels in tumor tissues (Figures 10C and D). Survival analysis revealed that low LUCAT1 expression was associated with favorable patient outcomes (Figures 10E–H). Spearman correlations confirmed that IKBKB-AC106786.1 and SQSTM1-LUCAT1 exhibited a significant association only in tumor samples (and not in normal tissues) (Figures 11A–D).
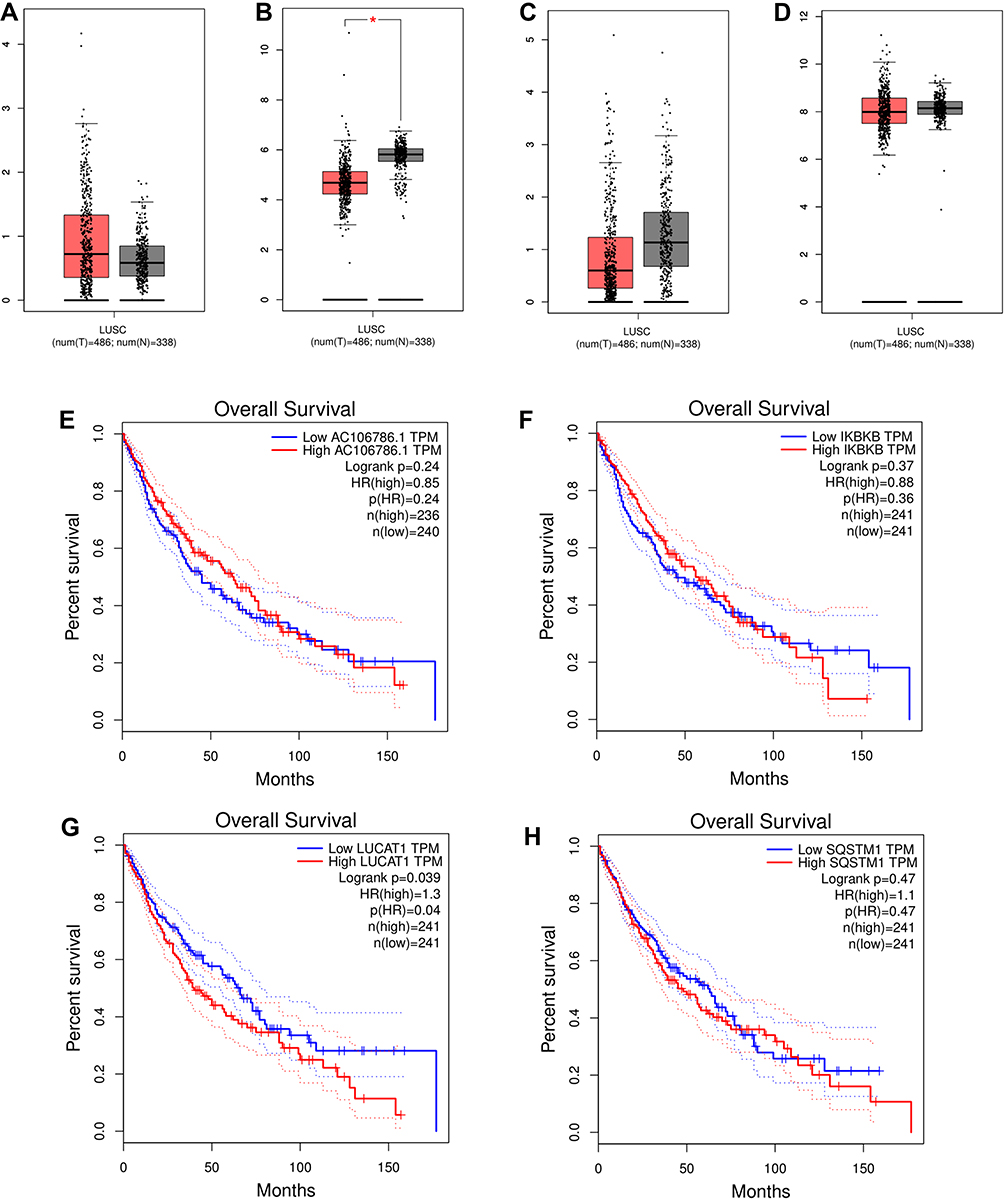 |
Figure 10 Validation of IKBKB-AC106786.1 and SQSTM1-LUCAT1 in GEPIA database. (A) The mRNA expression levels of AC106786.1; (B) The mRNA expression levels of IKBKB; (C) The mRNA expression levels of LUCAT1; (D) The mRNA expression levels of SQSTM1; (E) Kaplan-Meier survival analysis of AC106786.1; (F) Kaplan-Meier survival analysis of IKBKB; (G) Kaplan-Meier survival analysis of LUCAT1; (H) Kaplan-Meier survival analysis of SQSTM1. |
 |
Figure 11 Correlation test in GEPIA and protein expression in HPA. (A) Spearman correlation tests for IKBKB-AC106786.1 in tumor sample; (B) Spearman correlation tests for IKBKB-AC106786.1 in normal lung sample; (C) Spearman correlation tests for SQSTM1-LUCAT1 in tumor sample; (D) Spearman correlation tests for SQSTM1-LUCAT1 in normal lung sample; (E) the expression level of IKBKB in normal lung sample; (F) the expression level of IKBKB in tumor sample; (G) the expression level of SQSTM1 in normal lung sample; (H) the expression level of SQSTM1 in tumor sample. |
We then analyzed the protein expression of these molecules using the HPA database. IKBKB protein was expressed at low levels in tumor samples, consistent with the mRNA expression data. However, in tumor samples the expression of SQSTM1 at the protein level was higher than that at the mRNA level (Figures 11E–H).
Discussion
The treatment of patients with LUSC is still limited to traditional chemotherapy despite recent progress in treatments for various lung cancers. Cancer research has focused on autophagy as it can enhance the efficacy of the existing treatment and even reverse drug resistance, owing to its role in stress response and cell death.15,16 Certain autophagy proteins are significantly correlated with the survival of cancer patients; for instance, high ATG10 and LC3A expression is linked to poor survival outcomes.8,17 Because of these associations, researchers have employed autophagy genes to construct prognostic signatures that can predict the overall survival with some reliability.18,19 Similarly, lncRNAs have received attention for their involvement in tumorigenesis, suggesting a role as biomarkers in cancer diagnosis and prognosis.20 For example, a lncRNA-based predictive model was recently developed for lung adenocarcinoma with potential for future clinical use. These advances led us to investigate whether specific ARlncRNA signatures could be used to predict the prognosis of patients with LUSC.
We initially filtered out 1248 lncRNAs that significantly correlated with 210 autophagy-related genes from an LUSC cohort in TCGA. Using Lasso and multivariate Cox regressions, we established an independent prognostic model of seven ARlncRNAs to predict patient survival and calculated a risk score for each patient. Of the lncRNAs included in our model, LUCAT1 is the most extensively studied, with numerous studies demonstrating its vital role in various cancers, such as gastric cancer,21 esophageal cancer,22 liver cancer,23 and lung cancer.24 Consistent with our findings, LUCAT1 is known to be expressed at significantly higher levels in lung tumors than in the paracancerous tissue and is a negative indicator of prognosis.24 Overexpression of LUCAT1 may inhibit apoptosis and enhance cisplatin resistance development via IGF-2.25 This mechanism likely explains why LUSC patients with LUCAT1 overexpression have a poorer prognosis, given that the conventional treatment for this cancer is platinum-based. Although no cancer-related studies have been conducted on AC106786.1, it was correlated with the autophagy gene IKBKB, which plays a negative role in inhibiting cisplatin-induced apoptosis in lung adenocarcinoma.26 Likewise, EIF4EBP1—the gene correlated with AP006545.2, an under-researched lncRNA—plays a role in several cancers via the mTOR signaling pathway.27 EIF4EBP1 is also a key gene for predicting the prognosis of patients with lung cancer.19
Autophagy-related (ATG) proteins play a dual role in cancer, ie, they can inhibit as well as accelerate cancer progression. Even though little is known about the lncRNAs AC022150.2, AL035425.3, AC138976.2, and GPRC5D-AS1, they were all associated with ATGs in our study. Of the 16 known mammalian ATGs, ATG2, ATG4, ATG8, and ATG16 are most correlated with cancer in our established signature. ATG2 is an essential part of the ATG9/ATG12-WIPI complex that facilitates ATG9 recruitment to expand autophagosomes.4 ATG2B is associated with oncogenesis, and a frameshift mutation affecting this protein is implicated in the development of gastric and colorectal cancer.28 ATG4 activates LC3 by cleaving LC3-I to produce LC3-II.29 Enhanced ATG4 expression is positively correlated with the occurrence and progression of colorectal cancer,30 as well as with increased susceptibility to ovarian and lung cancer.31,32 As ATG4 can function as an alternative to anti-lysosomal treatment, there is widespread interest in developing drugs targeting ATG4B, ie, generation of several ATG4 inhibitors that have a potential clinical value.30,33,34 Finally, the ATG8 family member, GABARAPL1 is involved in cellular trafficking and cancer, in addition to autophagy. High GABARAPL1 expression is correlated with longer survival time in renal, prostate, breast, and liver cancers.35–37 In terms of mechanism, GABARAPL1 upregulation appears to induce cell arrest via the P53 pathway, thereby decreasing cell viability, proliferation, and migration.38
With respect to clinical significance, the high AUC values of our seven ARlncRNAs across 1, 3, and 5 years suggests that the model reliably predicted patient survival. Subsequent multivariate Cox analyses confirmed that the risk score generated using our model can be used as an independent prognostic indicator. Overall, our results suggest that the lncRNA signature has the potential to predict patient survival.
Functional enrichment analysis showed that the most significantly enriched KEGG pathways were MAPK and JAK/STAT signaling pathways, both known to be implicated in various cancers. In lung cancer, the MAPK axis crosstalks with some lncRNAs to promote cancer cell invasion and drug resistance.39,40 Additionally, STAT3 overexpression is observed in nearly 55% of the patients with lung cancer and most lung cancer cell lines.41 Furthermore, JAK1/STAT1 activation is positively correlated with cancer stage, especially with nodal metastasis.42 Beyond this, few studies have investigated the relationships between lncRNAs and various signaling pathways, a topic that we intend to pursue in future work.
Our study has several limitations. First, the small number of patients included means that our results may not be extrapolatable to a larger population. Second, the established prognostic model has not been tested, and a prospective study is needed to verify its predictive efficacy. Third, we did not detect lncRNA expression in tumor samples or identify potential molecular mechanisms, highlighting the need for additional basic biological experiments.
Conclusions
In conclusion, our study provided a comprehensive analysis of ARlncRNAs in LUSC. We constructed a co-expression network that points to a potential mechanism underlying the effects of the seven ARlncRNAs. The lncRNA-based prognostic signature was significantly correlated with patient survival, and the derived risk score was an independent predictive factor for LUSC progression. Moreover, genes associated with the seven ARlncRNAs were potential therapeutic targets in LUSC. Overall, we believe that the identified lncRNAs have clinical value as diagnostic or prognostic tools, and as candidates for drug development.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61(2):69–90. doi:10.3322/caac.20107
2. Rizvi NA, Hellmann MD, Brahmer JR, et al. Nivolumab in combination with platinum-based doublet chemotherapy for first-line treatment of advanced non-small-cell lung cancer. J Clin Oncol. 2016;34(25):2969–2979. doi:10.1200/JCO.2016.66.9861
3. Hsu HC, Li SJ, Chen CY, Chen MF. Eicosapentaenoic acid protects cardiomyoblasts from lipotoxicity in an autophagy-dependent manner. Cell Biol Toxicol. 2018;34(3):177–189. doi:10.1007/s10565-017-9406-9
4. Levine B, Kroemer G. Biological functions of autophagy genes: a disease perspective. Cell. 2019;176(1–2):11–42. doi:10.1016/j.cell.2018.09.048
5. Tsai WT, Lo YC, Wu MS, et al. Mycotoxin patulin suppresses innate immune responses by mitochondrial dysfunction and p62/sequestosome-1-dependent mitophagy. J Biol Chem. 2016;291(37):19299–19311. doi:10.1074/jbc.M115.686683
6. Mah LY, Ryan KM. Autophagy and cancer. Cold Spring Harb Perspect Biol. 2012;4(1):a008821. doi:10.1101/cshperspect.a008821
7. Levy JMM, Towers CG, Thorburn A. Targeting autophagy in cancer. Nat Rev Cancer. 2017;17(9):528–542. doi:10.1038/nrc.2017.53
8. Xie K, Liang C, Li Q, et al. Role of ATG10 expression quantitative trait loci in non-small cell lung cancer survival. Int J Cancer. 2016;139(7):1564–1573. doi:10.1002/ijc.30205
9. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
10. Liu Y, Sharma S, Watabe K. Roles of lncRNA in breast cancer. Front Biosci. 2015;7:94–108.
11. Thin KZ, Liu X, Feng X, Raveendran S, Tu JC. LncRNA-DANCR: a valuable cancer related long non-coding RNA for human cancers. Pathol Res Pract. 2018;214(6):801–805. doi:10.1016/j.prp.2018.04.003
12. Gibb EA, Brown CJ, Lam WL. The functional role of long non-coding RNA in human carcinomas. Mol Cancer. 2011;10(1):38. doi:10.1186/1476-4598-10-38
13. Kumar P, Khadirnaikar S, Shukla SK. A novel LncRNA-based prognostic score reveals TP53-dependent subtype of lung adenocarcinoma with poor survival. J Cell Physiol. 2019;234(9):16021–16031. doi:10.1002/jcp.28260
14. Yin YZ, Yao SH, Li CG, et al. Systematic analysis using a bioinformatics strategy identifies SFTA1P and LINC00519 as potential prognostic biomarkers for lung squamous cell carcinoma. Am J Transl Res. 2021;13(1):168–182.
15. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
16. Shintani T, Klionsky DJ. Autophagy in health and disease: a double-edged sword. Science. 2004;306(5698):990–995. doi:10.1126/science.1099993
17. White E, DiPaola RS. The double-edged sword of autophagy modulation in cancer. Clin Cancer Res. 2009;15(17):5308–5316. doi:10.1158/1078-0432.CCR-07-5023
18. Uberall I, Gachechiladze M, Joerger M, et al. Tumor autophagy is associated with survival outcomes in patients with resected non-small cell lung cancer. Lung Cancer. 2019;129:85–91. doi:10.1016/j.lungcan.2019.01.001
19. Liu Y, Wu L, Ao H, et al. Prognostic implications of autophagy-associated gene signatures in non-small cell lung cancer. Aging. 2019;11(23):11440–11462. doi:10.18632/aging.102544
20. Zhu J, Wang M, Hu D. Development of an autophagy-related gene prognostic signature in lung adenocarcinoma and lung squamous cell carcinoma. PeerJ. 2020;8:e8288. doi:10.7717/peerj.8288
21. Jin KT, Lu ZB, Lv JQ, Zhang JG. The role of long non-coding RNAs in mediating chemoresistance by modulating autophagy in cancer. RNA Biol. 2020;17(12):1727–1740. doi:10.1080/15476286.2020.1737787
22. Chi J, Liu T, Shi C, et al. Long non-coding RNA LUCAT1 promotes proliferation and invasion in gastric cancer by regulating miR-134-5p/YWHAZ axis. Biomed Pharmacother. 2019;118:109201. doi:10.1016/j.biopha.2019.109201
23. Rashid F, Shah A, Shan G. Long non-coding RNAs in the cytoplasm. Genomics Proteomics Bioinformatics. 2016;14(2):73–80. doi:10.1016/j.gpb.2016.03.005
24. Jiao Y, Li Y, Ji B, Cai H, Liu Y. Clinical value of lncRNA LUCAT1 expression in liver cancer and its potential pathways. J Gastrointestin Liver Dis. 2019;28(4):439–447. doi:10.15403/jgld-356
25. Sun Y, Jin SD, Zhu Q, et al. Long non-coding RNA LUCAT1 is associated with poor prognosis in human non-small lung cancer and regulates cell proliferation via epigenetically repressing p21 and p57 expression. Oncotarget. 2017;8(17):28297–28311. doi:10.18632/oncotarget.16044
26. Wang W, Dong ML, Zhang W, Liu T. Long noncoding LUCAT1 promotes cisplatin resistance of non-small cell lung cancer by promoting IGF-2. Eur Rev Med Pharmacol Sci. 2021;25(2):567. doi:10.26355/eurrev_202101_24593
27. Qi K, Li Y, Li X, Zhang F, Shao Y, Zhou Q. [Expression and significance of IKBKB in pulmonary adenocarcinoma A549 cells and its cisplatin-resistant variant A549/DDP]. Zhongguo Fei Ai Za Zhi. 2014;17(5):363–368. Chinese. doi:10.3779/j.issn.1009-3419.2014.05.01
28. Karlsson E, Perez-Tenorio G, Amin R, et al. The mTOR effectors 4EBP1 and S6K2 are frequently coexpressed, and associated with a poor prognosis and endocrine resistance in breast cancer: a retrospective study including patients from the randomised Stockholm tamoxifen trials. Breast Cancer Res. 2013;15(5):R96. doi:10.1186/bcr3557
29. Kang MR, Kim MS, Oh JE, et al. Frameshift mutations of autophagy-related genes ATG2B, ATG5, ATG9B and ATG12 in gastric and colorectal cancers with microsatellite instability. J Pathol. 2009;217(5):702–706. doi:10.1002/path.2509
30. Li M, Hou Y, Wang J, Chen X, Shao ZM, Yin XM. Kinetics comparisons of mammalian Atg4 homologues indicate selective preferences toward diverse Atg8 substrates. J Biol Chem. 2011;286(9):7327–7338. doi:10.1074/jbc.M110.199059
31. Liu PF, Leung CM, Chang YH, et al. ATG4B promotes colorectal cancer growth independent of autophagic flux. Autophagy. 2014;10(8):1454–1465. doi:10.4161/auto.29556
32. Liao YP, Chen LY, Huang RL, et al. Hypomethylation signature of tumor-initiating cells predicts poor prognosis of ovarian cancer patients. Hum Mol Genet. 2014;23(7):1894–1906. doi:10.1093/hmg/ddt583
33. He Q, Lu Y, Hu S, et al. An intron SNP rs807185 in ATG4A decreases the risk of lung cancer in a southwest Chinese population. Eur J Cancer Prev. 2016;25(4):255–258. doi:10.1097/CEJ.0000000000000174
34. Shu CW, Madiraju C, Zhai D, et al. High-throughput fluorescence assay for small-molecule inhibitors of autophagins/Atg4. J Biomol Screen. 2011;16(2):174–182. doi:10.1177/1087057110392996
35. Xu D, Xu Z, Han L, et al. Identification of new ATG4B inhibitors based on a novel high-throughput screening platform. SLAS Discov. 2017;22(4):338–347. doi:10.1177/1087057116639202
36. Su W, Li S, Chen X, et al. GABARAPL1 suppresses metastasis by counteracting PI3K/Akt pathway in prostate cancer. Oncotarget. 2017;8(3):4449–4459. doi:10.18632/oncotarget.13879
37. Uhlen M, Zhang C, Lee S, et al. A pathology atlas of the human cancer transcriptome. Science. 2017;357(6352):eaan2507. doi:10.1126/science.aan2507
38. Berthier A, Seguin S, Sasco AJ, et al. High expression of gabarapl1 is associated with a better outcome for patients with lymph node-positive breast cancer. Br J Cancer. 2010;102(6):1024–1031. doi:10.1038/sj.bjc.6605568
39. Rho SB, Byun HJ, Kim BR, et al. GABAA receptor-binding protein promotes sensitivity to apoptosis induced by chemotherapeutic agents. Int J Oncol. 2013;42(5):1807–1814. doi:10.3892/ijo.2013.1866
40. Zhang YX, Yuan J, Gao ZM, Zhang ZG. LncRNA TUC338 promotes invasion of lung cancer by activating MAPK pathway. Eur Rev Med Pharmacol Sci. 2018;22(2):443–449. doi:10.26355/eurrev_201801_14193
41. Yu T, Bai W, Su Y, Wang Y, Wang M, Ling C. Enhanced expression of lncRNA ZXF1 promotes cisplatin resistance in lung cancer cell via MAPK axis. Exp Mol Pathol. 2020;116:104484. doi:10.1016/j.yexmp.2020.104484
42. Yu H, Jove R. The STATs of cancer–new molecular targets come of age. Nat Rev Cancer. 2004;4(2):97–105. doi:10.1038/nrc1275.
43. Geeleher P, Cox N, Huang RS, Barbour JD. pRRophetic: an R package for prediction of clinical chemotherapeutic response from tumor gene expression levels. PLoS One. 2014;9(9):e107468. doi:10.1371/journal.pone.0107468
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.