Back to Journals » Neuropsychiatric Disease and Treatment » Volume 11
An association study between DLGAP1 rs11081062 and EFNA5 rs26728 polymorphisms with obsessive–compulsive disorder in a Chinese Han population
Authors Li J, Cui J, Wang X, Ma J, Niu H, Ma X, Zhang X ![]() , Liu S
, Liu S
Received 26 September 2014
Accepted for publication 12 February 2015
Published 1 April 2015 Volume 2015:11 Pages 897—905
DOI https://doi.org/10.2147/NDT.S75009
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Wai Kwong Tang
Jiang Li,1 Jiajia Cui,2 Xiuhai Wang,3 Jianhua Ma,4 Haitao Niu,4 Xu Ma,5–7 Xinhua Zhang,2,4 Shiguo Liu8
1Physiatry Department, The Affiliated Hospital of Qingdao University, 2Department of Psychiatry, 3Department of Genetics, Medical College, Qingdao University, 4The Affiliated Hospital of Qingdao University, Qingdao, People’s Republic of China; 5Graduate School, Peking Union Medical College, 6National Research Institute for Family Planning, 7World Health Organization Collaborating Centre for Research in Human Reproduction, Beijing, People’s Republic of China; 8Prenatal Diagnosis Center, The Affiliated Hospital of Qingdao University, Qingdao, People’s Republic of China
Background: A recent genome-wide association study indicated that the single nucleotide polymorphisms (SNPs) rs11081062 in DLGAP1 and rs26728 in EFNA5 were associated with obsessive–compulsive disorder (OCD) in Caucasians. The present case–control association study assessed the global relevance of these two SNPs with respect to OCD subtypes in a Chinese Han population.
Methods: We recruited 320 OCD patients and 431 age- and sex-matched controls from a Chinese Han population. rs11081062 and rs26728 SNPs were genotyped by real-time TaqMan polymerase chain reaction, and the chi-squared test was used to compare allele and genotype frequencies of variants between the two groups.
Results: No significant differences were found in allele or genotype frequencies of DLGAP1 rs11081062 and EFNA5 rs26728 between the OCD and control groups. Moreover, consistently negative results were observed when classifying by sex, onset age, and comorbidity. However, on analyzing OCD subphenotypes, significant associations were observed between rs11081062 and the presence of contamination obsessions and cleaning compulsions (χ2=7.724, P=0.021 by genotype; χ2=3.745, P=0.053 by allele; and χ2=0.821, P=0.365 by genotype, χ2=27.809, P=0.000 by allele, respectively), and rs26728 with the presence of repeating compulsions (χ2=8.285, P=0.004 by genotype; χ2=7.512, P=0.006 by allele).
Conclusion: Although we found no association between DLGAP1 rs11081062 and EFNA5 rs26728 SNPs with OCD in a Chinese Han population, obvious associations were observed with OCD subphenotypes. Therefore, it appears to be useful to divide OCD into more homogeneous subphenotypes to help understand the complex genetic basis of this disorder. Further investigations are needed to replicate these findings using larger sample sizes, different populations, and other polymorphisms.
Keywords: DLGAP1, EFNA5, obsessive-compulsive disorder, polymorphism
Introduction
Obsessive–compulsive disorder (OCD) is a neuropsychiatric disease characterized by obsessions (intrusive unwanted thoughts and/or images) and compulsions (repetitive behavior). Its lifetime prevalence ranges from 1% to 3%,1,2 making it one of the most common mental disorders. OCD has a great influence on the daily life and social capacity of patients. Its symptoms vary but are always accompanied by other psychiatric diseases such as Tourette syndrome3 or anxiety,4 which leads to difficulties in selecting the most appropriate therapy. Early-onset (EO) OCD affects 20% of all OCD patients, according to the US National Comorbidity Survey Replication.5
Twin and family studies have revealed the involvement of genetic factors in the etiology of OCD. Pauls reported that the OCD concordance rate of monozygotic twins was 47%–50%, compared with that of dizygotic twins at 80%–87%.6 Moreover, the pathogenesis of OCD demonstrated family convergence, with relatives of affected patients being more prone to developing OCD than those of controls.7,8 van Grootheest et al concluded that obsessive–compulsive symptoms are heritable in both children and adults, reporting genetic influences of 45%–65% and 27%–47%, respectively.9 On this basis, screening of candidate OCD susceptibility genes has been attempted,10–12 but the results are inconsistent and controversial. Indeed, none of the candidate genes were found to be involved in the direct pathogenesis of OCD. Recently, Stewart et al conducted an OCD genome-wide association study (GWAS) involving trios and unselected controls. Although no single nucleotide polymorphisms (SNPs) were shown to be associated with OCD at the genome-wide significant level, two SNPs (rs11081062 in DLGAP1 and rs26728 in EFNA5) were suggested to play a role in the etiology of OCD in Caucasians.13
DLGAP1, located on chromosome 18p11.31, encodes a postsynaptic scaffold protein with five conserved 14 amino acid repeats in its N-terminal,14 which interact with the guanylate kinase-like domain of postsynaptic density protein 95 (PSD-95). This interaction was linked with the synaptic plasticities in OCD.15 EFNA5 encodes the brain protein ephrin-A5, which plays an important role in cell morphology and behavior through Eph-A/B receptor signaling,16 particularly in the nervous system. EFNA5 has also previously been associated with susceptibility to Alzheimer’s disease17 and Parkinson’s disease.18
Recent models indicated that the main difficulty in identifying susceptibility genes in OCD derives from its symptom heterogeneity. Therefore, to resolve the complex genetic basis of the disease, dissection of OCD into different subphenotypes could be an effective method. Thus, the present case–control study genotyped the DLGAP1 rs11081062 and EFNA5 rs26728 polymorphisms using TaqMan real-time polymerase chain reaction (PCR) to analyze the possible relationship between these variants and OCD subphenotypes, including EO OCD and OCD with comorbid tic disorders, in a Chinese Han population.
Materials and methods
Patient population
The proband sample consisted of 320 individuals (178 males, 142 females) recruited from the Affiliated Hospital of Qingdao University (Qingdao, People’s Republic of China) with an average age of 29.270±13.961 years. Inclusion criteria were 1) OCD diagnosed according to the Diagnostic and Statistical Manual of Mental Disorders, 4th edition (DSM-IV) criteria19 by means of the Structured Clinical Interview for DSM-IV; 2) Chinese Han individual; and 3) first diagnosed or treatment-free or medication-free for at least 6 weeks. Exclusion criteria were 1) presence of any serious psychiatric diseases other than OCD, or related family history; 2) history of alcohol and/or drug abuse/dependence; 3) presence of any serious concomitant general medical condition or neurological disease, with the exception of Tourette syndrome; 4) history of medical disorders that may have a causal relationship with OCD; and 5) pregnancy or lactation. The symptoms and severity of OCD were evaluated using the Yale–Brown Obsessive Compulsive Scale symptom checklist20 including seven obsessive items (aggressive, contamination, sexual, symmetry, exactness, miscellaneous, and somatic) and seven compulsive items (cleaning, washing, checking, repeating, ordering, arranging, and miscellaneous) by an experienced physician.
In parallel, a control group of 431 disease-free individuals were selected from the Physical Examination Center of the Affiliated Hospital of Qingdao University. These were matched with the patients for age (mean age, 28.773±9.254 years), sex (males 239, females 192), geographic region, and ethnic origin, and had a negative family history of related disease. All subjects provided written informed consent for participation in the study.
Genotyping
Venous blood samples from patients and controls were collected, and DNA was extracted by standard procedures. TaqMan genotyping was performed for SNPs rs11081062 and rs26728 using the following reactions: 5 U/L super mix, 0.9 ng forward primer, 0.9 ng downstream primer, 0.5 ng FAM (6-carboxyfluorescein), 0.5 ng VIC (4,7,2′-trichloro-7′-phenyl-6-carboxyfluorescein), and 1 U/L genomic DNA. PCR was conducted in an Applied Biosystems® 7500 real-time PCR system with the following conditions: 95°C for 3 minutes, followed by 40 cycles of 95°C for 15 seconds, and 60°C for 1 minute. Positive and negative controls were included with all reactions. Genotype data were interpreted using SDS software (Thermo Fisher Scientific, Waltham, MA, USA).
Statistical analysis
Data analysis was carried out using SPSS software (version 17.0 for Windows; SPSS, Inc., Chicago, IL, USA). Age comparisons between OCD and control groups were made using the t-test, while the chi-squared test compared allelic and genotypic frequencies and sex ratios between patients and controls, estimated the Hardy–Weinberg equilibrium, and verified differences in genotypic frequencies between OCD symptom-present and symptom-absent as well.
Results
Demographic data
No significant difference was detected between the age or sex of the patients and controls (total t=0.587, P=0.558; male t=0.401, P=0.689; female t=0.870, P=0.385; χ2=0.02, P=0.962). Genotypic frequencies for rs11081062 and rs26728 in the control group followed the Hardy–Weinberg equilibrium distribution (rs11081062, χ2=1.807, P=0.179; rs26728, χ2=2.349, P=0.125).
Case–control study of DLGAP1 rs11081062
The T allele of DLGAP1 rs11081062 had a frequency of 0.363 in the OCD group and 0.393 in the control group, but was not significantly associated with OCD according to the allelic χ2 association test (χ2=1.476, P=0.224). Additionally, no significant differences in genotype distribution were found between the OCD group and controls as a whole (χ2=3.094, P=0.213) (Table 1). To eliminate the bias exerted by sex, we classified the OCD subjects into males and females and then compared them with their control counterparts. No significant differences in genotypic or allelic frequency distributions of rs11081062 were detected in the two sexes between patients and controls. We also divided the OCD group into EO OCD and late-onset (LO) OCD according to the age of onset of obsessive and/or compulsive behavior, and further subdivided these groups according to sex into female EO, female LO, male EO, and male LO to explore the role of age in the etiology of OCD. The complete list of allele frequencies and genotype rates are shown in Table 1. Regardless of sex, the onset age of OCD was found to be an irrelevant factor in the progress of OCD: genotypic and allelic variants were not significantly different between EO and OCD, LO and OCD, male EO and male effects, male LO and male OCD, female EO and female effects, or female LO and female OCD.
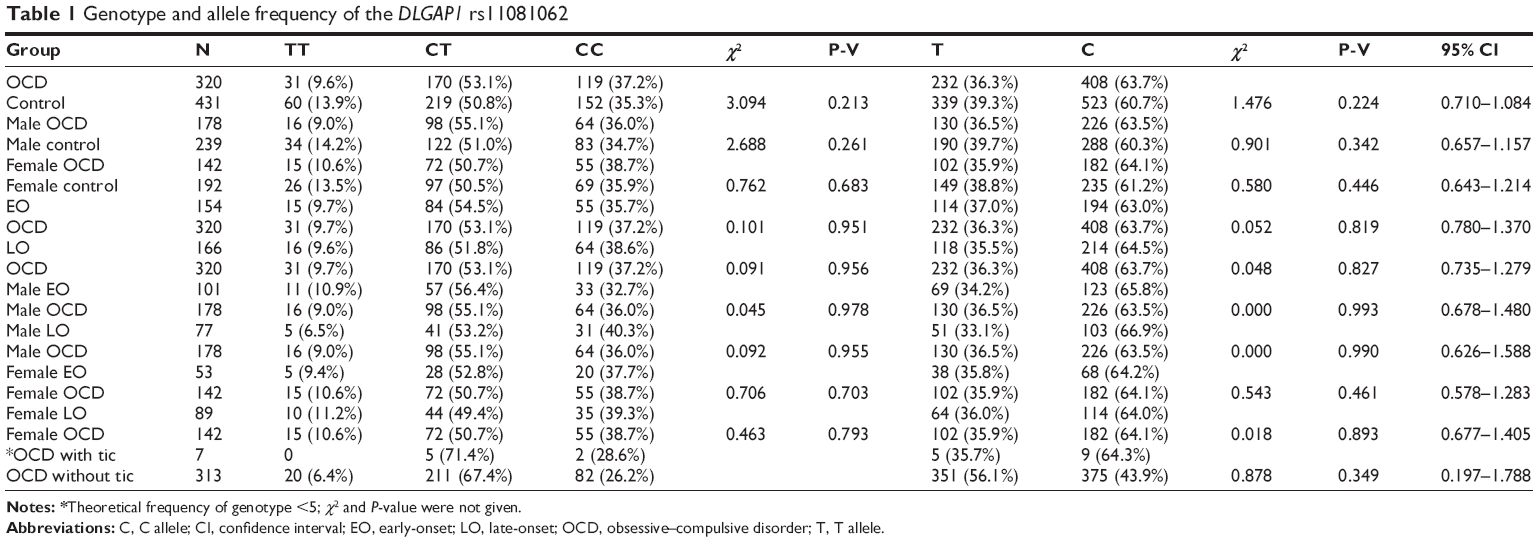 | Table 1 Genotype and allele frequency of the DLGAP1 rs11081062 |
We performed an additional investigation to explore the relationship between rs11081062 and different OCD symptoms. Interestingly, patients with contamination obsessions or cleaning compulsions were found to have significant differences in the ratio of the rs11081062 T allele versus the C allele compared with those OCD patients lacking these symptoms (contamination obsessive genotype, χ2=7.724, P=0.021, allele, χ2=3.745 P=0.053; cleaning compulsive genotype, χ2=0.821, P=0.365, allele, χ2=27.809, P=0.000) (Table 2).
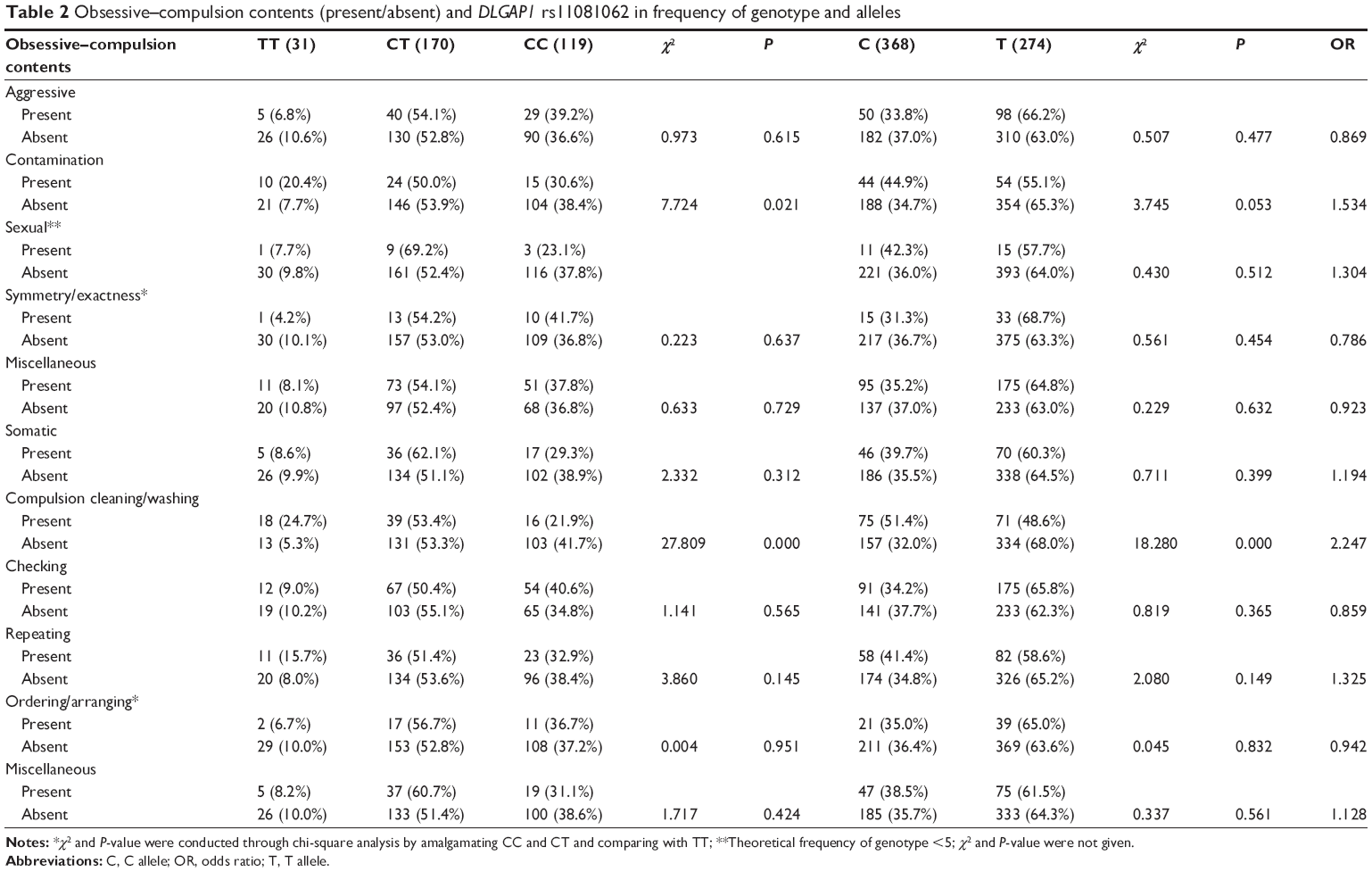 | Table 2 Obsessive–compulsion contents (present/absent) and DLGAP1 rs11081062 in frequency of genotype and alleles |
Case–control study of EFNA5 rs26728
The distribution of genotypic and allelic frequencies showed no significant differences between OCD and control groups as a whole (genotype, χ2=1.314, P=0.252; allele, χ2=1.421, P=0.233) (Table 3). Similarly, no significant differences were observed after analyzing the data according to sex or age of onset. However, patients with a repeating compulsion were positively associated with SNP rs26728 in terms of genotypic frequencies and ratio of the T allele compared with OCD patients lacking this symptom (genotype, χ2=8.285, P=0.004; allele, χ2=7.512, P=0.006). No other associations with rs26728 genotype or allele frequencies were identified between OCD patients with and without particular symptoms (Table 4).
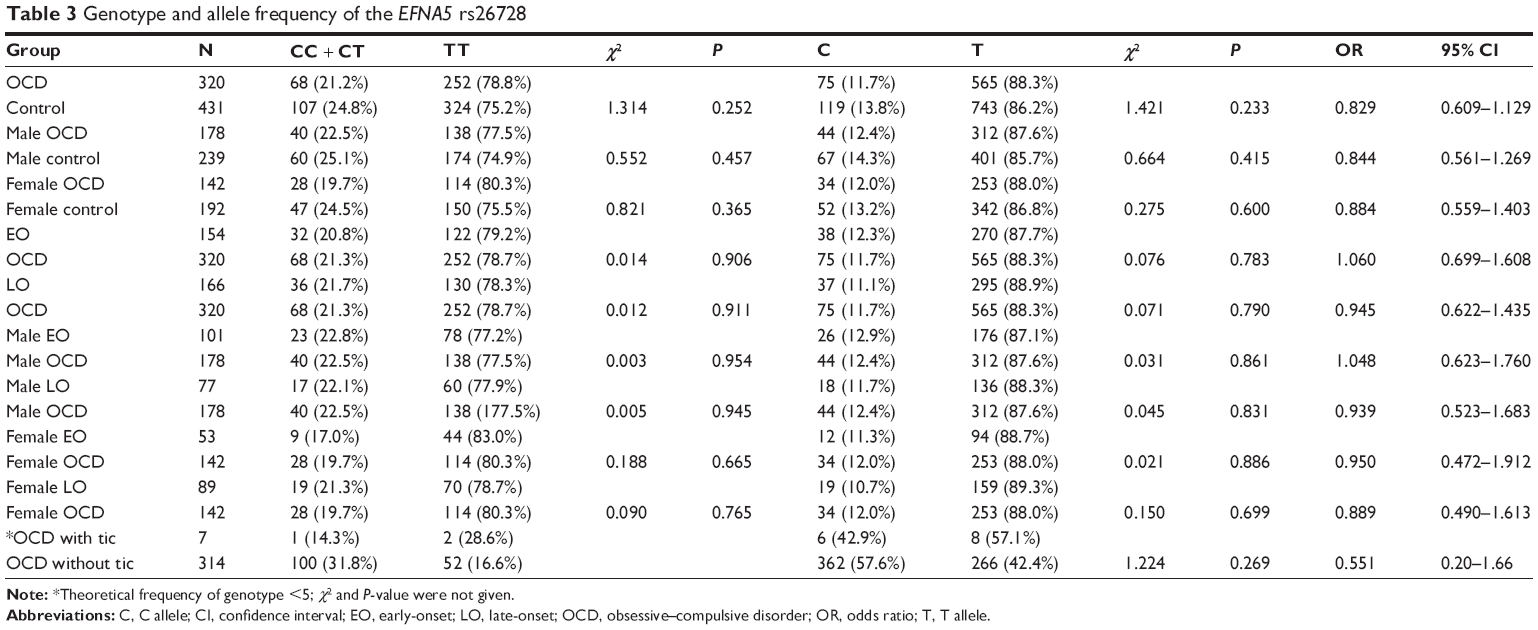 | Table 3 Genotype and allele frequency of the EFNA5 rs26728 |
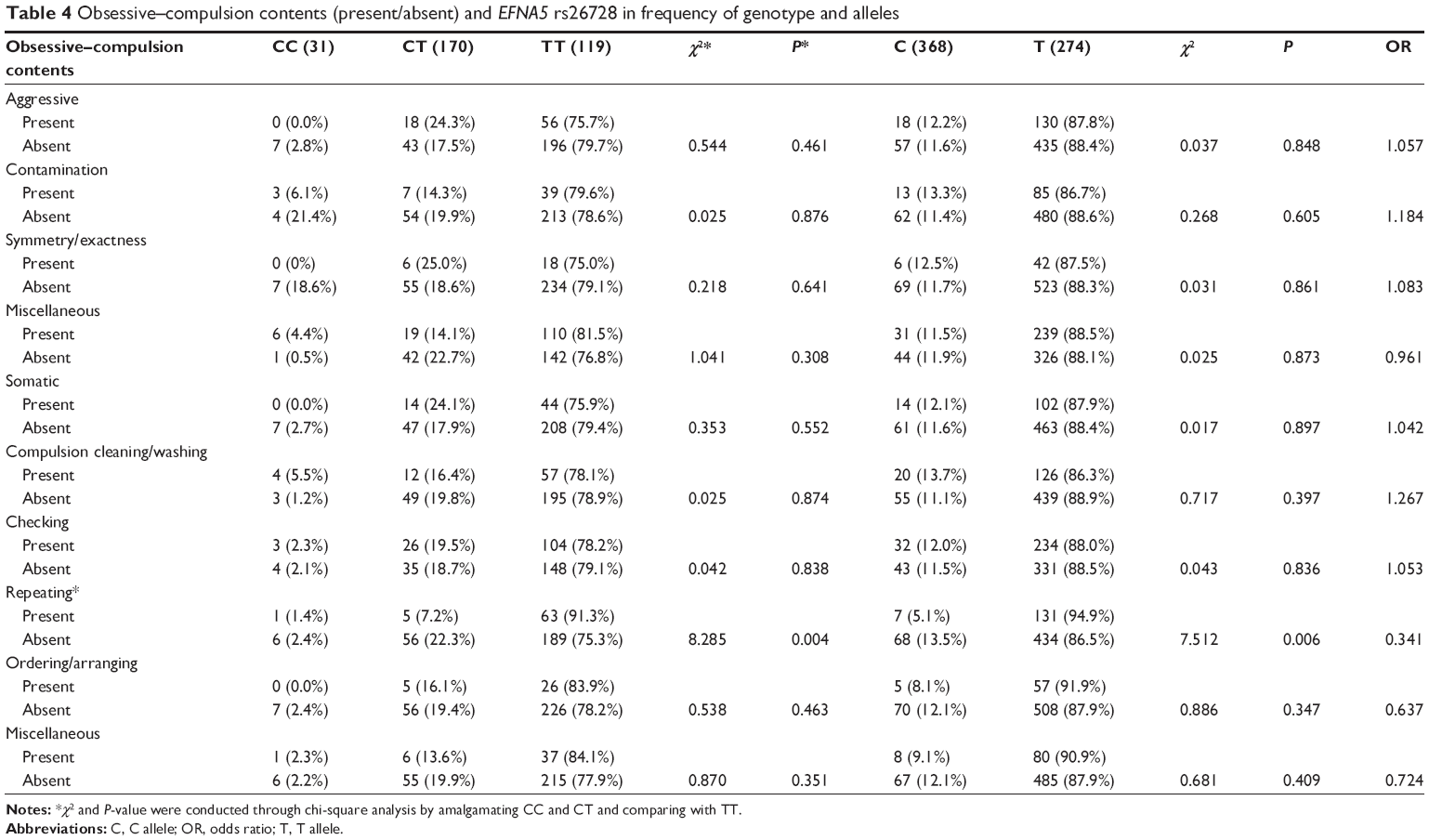 | Table 4 Obsessive–compulsion contents (present/absent) and EFNA5 rs26728 in frequency of genotype and alleles |
Discussion
The aim of our study was to verify the functional variants of DLGAP rs11081062 and EFNA5 rs26728 in the etiology of OCD, which was initially suggested in a GWAS.13 The role of genetic factors in OCD was purported to be based on psychological evidence, and many different system-related candidate genes have been investigated for their involvement in disease, including those from the dopamine system,21 glutamine system,22 and serotonin.11,12 Such studies have produced inconsistent and controversial findings, which could reflect differences in ethnicities or sex, genotype inaccuracies, or small sample sizes. The GWAS by Stewart et al investigated 1,465 OCD cases, 5,557 ancestry-matched controls, and 400 complete trios from a Caucasian population.13 This study identified SNPs in DLGAP1 and EFNA5 as being important in the development of OCD.
DLGAP1 encodes DLGAP, which is also known as SAP90/PSD-95-associated protein (SAPAP), and guanylate kinase-associated protein, both of which interact with the guanylate kinase domain of PSD-95. PSD-95 is a member of the PSD-95 synaptic ion channel clustering protein family,23,24 which is crucial in the structural and functional organization of postsynaptic neurotransmitter receptor proteins and in the adhesion of the postsynapse to presynaptic terminals.25 SAPAP and guanylate kinase binding plays an important role in the anchoring of channel/PSD-95 clusters to the postsynaptic density. By combining with other proteins, guanylate kinase-associated protein may function as a molecular scaffold that links ion channel/PSD-95 clusters to the subsynaptic cytoskeleton or to downstream signaling molecules.26 Recent research has shown that increased expression of SAPAP occurs in the nucleus accumbens of patients with schizophrenia and in phencyclidine-treated rats, suggesting that abnormalities in intracellular signal transduction involving interactions between PSD-95 and N-methyl-d-aspartate receptors might be closely linked with schizophrenia.27
EFNA5, a member of the ephrin gene family, encodes EFNA5, a tyrosine kinase receptor, which stimulated axon fasciculation and prevented its bundling in cocultures of cortical neurons with astrocytes.28 The EPH and EPH-related receptors function as receptor tyrosine kinases and have been implicated in the mediation of developmental events, particularly in the nervous system.28,29 Ephrin-A5 has two isoforms, the canonical full-length (ephrinA5L) and a shorter variant (ephrinA5S). Both isoforms inhibited neurite outgrowth of dorsal root ganglia in an earlier study.30
In our study of 320 OCD patients and 431 healthy controls in a Chinese Han population, no significant variants were found in allele or genotype frequencies of rs11081062 and rs26728 SNPs between the two groups. Hall et al previously reported that the brain-derived neurotrophic factor (BDNF) V66M polymorphism might be significantly associated with EO OCD rather than adult-onset OCD,31 while Enoch et al32 and Lochner et al33 showed that the HTR2A variant played a critical role in the etiology of female OCD. Based on these sex and age of disease onset preferences, we carried out an additional classification of the two groups into males and females and EO and LO, but still found no significant differences between groups with respect to the two SNPs. Although these results indicated that the etiology of OCD might not be associated with DLGAP1 and EFNA5 in a Chinese Han population, ethnic differences should be taken into consideration as an important factor exerting a potential false negative effect given the small sample size. Moreover, other potential OCD-related genes linked to DLGAP1 or EFNA5 might also influence the results.
To resolve the complex genetic basis of OCD, division of the disorder into different subphenotypes has been proposed as an effective technique to produce more etiologically homogeneous groups closely linked to the action of genes. A case–control study indicated that glutamatergic variants might contribute to the genetic vulnerability of certain OCD manifestations,22 while the frequency of the BDNF Val allele was shown to be significantly higher among OCD patients with hoarding compulsive behavior than those without.34 We divided our OCD patients into positive symptom and negative symptom groups to analyze the relationship between rs11081062 and rs26728 and different OCD subtypes. Individuals with rs11081062 T alleles were found to be at a significantly higher risk of being classified as cleaning compulsive and contamination obsessive than those carrying C alleles. Allele-wise comparisons also revealed a significant difference in the frequency of rs26728 T alleles and genotype between the repeating group and the nonrepeating group. Taken together, these results indicate that SNPs rs11081062 and rs26728 might be involved in the etiopathology of OCD subphenotypes. Moreover, the present study also supports the division of OCD into more homogeneous subphenotypes to better understand the complex genetic basis of this disorder.
Acknowledgments
We thank all probands for their participation in this study. This work was supported by the National Natural Science Foundation of China (81371499 and 30971586).
Disclosure
The authors report no conflicts of interest in this work.
References
Fontenelle LF, Mendlowicz MV, Versiani M. The descriptive epidemiology of obsessive-compulsive disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2006;30(3):327–337. | ||
Ruscio AM, Stein DJ, Chiu WT, Kessler RC. The epidemiology of obsessive-compulsive disorder in the National Comorbidity Survey Replication. Mol Psychiatry. 2010;15(1):53–63. | ||
Leckman JF, Goodman WK, Anderson GM, et al. Cerebrospinal fluid biogenic amines in obsessive compulsive disorder, Tourette’s syndrome, and healthy controls. Neuropsychopharmacology. 1995;12(1):73–86. | ||
Nutt D, Malizia A. Anxiety and OCD – the chicken or the egg? J Psychopharmacol. 2006;20(6):729–731. | ||
Kessler RC, Berglund P, Demler O, Jin R, Merikangas KR, Walters EE. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Arch Gen Psychiatry. 2005;62(6):593–602. | ||
Pauls DL. The genetics of obsessive-compulsive disorder: a review. Dialogues Clin Neurosci. 2010;12(2):149–163. | ||
Pauls DL, Alsobrook JP, Goodman W, Rasmussen S, Leckman JF. A family study of obsessive-compulsive disorder. Am J Psychiatry. 1995;152(1):76–84. | ||
Nestadt G, Samuels J, Riddle M, et al. A family study of obsessive-compulsive disorder. Arch Gen Psychiatry. 2000;57(4):358–363. | ||
van Grootheest DS, Cath DC, Beekman AT, Boomsma DI. Twin studies on obsessive-compulsive disorder: a review. Twin Res Hum Genet. 2005;8(5):450–458. | ||
Stewart SE, Fagerness JA, Platko J, et al. Association of the SLC1A1 glutamate transporter gene and obsessive-compulsive disorder. Am J Med Genet B Neuropsychiatr Genet. 2007;144B(8):1027–1033. | ||
Wu H, Wang X, Xiao Z, et al. Association between SLC1A1 gene and early-onset OCD in the Han Chinese population: a case-control study. J Mol Neurosci. 2013;50(2):353–359. | ||
Tibrewal P, Kumar HB, Shubha GN, et al. Association of serotonin transporter gene polymorphisms with obsessive-compulsive disorder (OCD) in a south Indian population. Indian J Med Res. 2010;132:690–695. | ||
Stewart SE, Yu D, Scharf JM, et al; North American Brain Expression Consortium; UK Brain Expression Database. Genome-wide association study of obsessive-compulsive disorder. Mol Psychiatry. 2013;18(7):788–798. | ||
Zhu J, Shang Y, Xia C, Wang W, Wen W, Zhang M. Guanylate kinase domains of the MAGUK family scaffold proteins as specific phospho-protein-binding modules. EMBO J. 2011;30(24):4986–4997. | ||
Welch JM, Lu J, Rodriguiz RM, et al. Cortico-striatal synaptic defects and OCD-like behaviours in Sapap3-mutant mice. Nature. 2007;448(7156):894–900. | ||
Himanen JP, Chumley MJ, Lackmann M, et al. Repelling class discrimination: ephrin-A5 binds to and activates EphB2 receptor signaling. Nat Neurosci. 2004;7(5):501–509. | ||
Potkin SG, Guffanti G, Lakatos A, et al; Alzheimer’s Disease Neuroimaging Initiative. Hippocampal atrophy as a quantitative trait in a genome-wide association study identifying novel susceptibility genes for Alzheimer’s disease. PLoS One. 2009;4(8):e6501. | ||
Lesnick TG, Papapetropoulos S, Mash DC, et al. A genomic pathway approach to a complex disease: axon guidance and Parkinson disease. PLoS Genet. 2007;3(6):e98. | ||
First MB, Spitzer RL, Gibbon M, Williams JBW. (1994). Structured Clinical Interview for DSM-IV Axis I Disorders, Patient Edition (SCID-P), Version 2. New York, NY: University of NY. | ||
Goodman WK, Pice LH, Rasmussen SA, Mazure C, Fleischmann RL, Hill CL, et al. The Yale-Brown Obsessive Compulsive Scale. I. Development, use, and reliability. Arch Gen Psychiatry. 1989;46(11):1006–1011. | ||
Schneier FR, Martinez D, Abi-Dargham A, et al. Striatal dopamine D(2) receptor availability in OCD with and without comorbid social anxiety disorder: preliminary findings. Depress Anxiety. 2008;25(1):1–7. | ||
Arnold PD, Rosenberg DR, Mundo E, Tharmalingam S, Kennedy JL, Richter MA. Association of a glutamate (NMDA) subunit receptor gene (GRIN2B) with obsessive-compulsive disorder: a preliminary study. Psychopharmacology (Berl). 2004;174(4):530–538. | ||
Froehner SC. Regulation of ion channel distribution at synapses. Annu Rev Neurosci. 1993;16:347–368. | ||
Hall ZW, Sanes JR. Synaptic structure and development: the neuromuscular junction. Cell. 1993;72 Suppl:99–121. | ||
Ziff EB. Enlightening the postsynaptic density. Neuron. 1997;19(6):1163–1174. | ||
Kim E, Naisbitt S, Hsueh YP, et al. GKAP, a novel synaptic protein that interacts with the guanylate kinase-like domain of the PSD-95/SAP90 family of channel clustering molecules. J Cell Biol. 1997;136(3):669–678. | ||
Kajimoto Y, Shirakawa O, Lin XH, et al. Synapse-associated protein 90/postsynaptic density-95-associated protein (SAPAP) is expressed differentially in phencyclidine-treated rats and is increased in the nucleus accumbens of patients with schizophrenia. Neuropsychopharmacology. 2003;28(10):1831–1839. | ||
Winslow JW, Moran P, Valverde J, et al. Cloning of AL-1, a ligand for an Eph-related tyrosine kinase receptor involved in axon bundle formation. Neuron. 1995;14(5):973–981. | ||
Martínez A, Soriano E. Functions of ephrin/Eph interactions in the development of the nervous system: emphasis on the hippocampal system. Brain Res Brain Res Rev. 2005;49(2):211–226. | ||
Lai KO, Ip FC, Ip NY. Identification and characterization of splice variants of ephrin-A3 and ephrin-A5. FEBS Lett. 1999;458(2):265–269. | ||
Hall D, Dhilla A, Charalambous A, Gogos JA, Karayiorgou M. Sequence variants of the brain-derived neurotrophic factor (BDNF) gene are strongly associated with obsessive-compulsive disorder. Am J Hum Genet. 2003;73(2):370–376. | ||
Enoch MA, Greenberg BD, Murphy DL, Goldman D. Sexually dimorphic relationship of a 5-HT2A promoter polymorphism with obsessive-compulsive disorder. Biol Psychiatry. 2001;49(4):385–388. | ||
Lochner C, Hemmings SM, Kinnear CJ, et al. Corrigendum to “gender in obsessive-compulsive disorder: clinical and genetic findings” [Eur Neuropsychopharmacol. 2004;14:105–113]. Eur Neuropsychopharmacol. 2004;14(5):437–445. | ||
Timpano KR, Schmidt NB, Wheaton MG, Wendland JR, Murphy DL. Consideration of the BDNF gene in relation to two phenotypes: hoarding and obesity. J Abnorm Psychol. 2011;120(3):700–707. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.