Back to Journals » Cancer Management and Research » Volume 17

ALPP Induces Epithelial-Mesenchymal Transition by Activating the Wnt/β-Catenin Signaling Pathway in Colorectal Cancer Cells
Authors Gao B, Li B ![]() , Hu J, Hu X
, Hu J, Hu X ![]() , Su M
, Su M
Received 9 June 2025
Accepted for publication 23 November 2025
Published 22 December 2025 Volume 2025:17 Pages 3227—3240
DOI https://doi.org/10.2147/CMAR.S545808
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Lu-Zhe Sun
Bo Gao, Baokun Li, Jitao Hu, Xuhua Hu, Mingming Su
Second Department Surgery, The Fourth Hospital of Hebei Medical University, Shijiazhuang, Hebei, 050011, People’s Republic of China
Correspondence: Mingming Su, Second Department Surgery, The Fourth Hospital of Hebei Medical University, 12 Jiankang Road, Changan District, Shijiazhuang, Hebei, 050000, People’s Republic of China, Tel +86-13673112014, Email [email protected]
Purpose: Colorectal cancer (CRC) is a prevalent cancer worldwide, with metastasis significantly contributing to its high mortality and poor prognosis. This study focuses on the impact of Alkaline Phosphatase, Placental (ALPP) on epithelial-mesenchymal transition (EMT) in colorectal cancer cells and its role in the Wnt/β-catenin signaling pathway.
Patients and Methods: Differential ALPP expression was first interrogated in metastatic versus non-metastatic CRC samples from The Cancer Genome Atlas (TCGA-CRC) cohort. Functional validation was subsequently performed in vitro with HT29 and HCT116 cell lines engineered for ALPP overexpression or CRISPR/Cas9-mediated knockdown. Proliferation, migration and invasion were quantified by CCK-8, wound-healing and Transwell assays; EMT and Wnt/β-catenin signaling were assessed by Western blot.
Results: Bioinformatics analysis revealed significantly different ALPP expression between metastatic and non-metastatic patients. In vitro experiments further revealed that ALPP overexpression drives proliferation, invasion, migration and, consequently, metastasis of HT29 and HCT116 colorectal cancer cells, whereas ALPP knockdown abolishes these EMT-dependent effects. Increased ALPP expression resulted in increased levels of N-Cadherin, Vimentin, and Snail proteins, along with a decrease in E-cadherin protein expression, in contrast to findings following ALPP knockdown. Furthermore, ALPP overexpression was also associated with Wnt/β-catenin signaling pathway activation.
Conclusion: ALPP was found to act as an oncogenic factor in colorectal cancer cell lines HT29 and HCT116, stimulating cell proliferation and facilitating EMT. Abnormal activation of the Wnt/β-catenin signaling pathway was also found to be linked to increased ALPP expression.
Keywords: alkaline phosphatase, colorectal cancer cells, Wnt/β-catenin signaling pathway, epithelial-mesenchymal transition, cell invasion
Introduction
Colorectal cancer (CRC) is the second most common cancer in the United States and a leading cause of cancer-related deaths. As of 2023, it is estimated that approximately 150,000 individuals will be diagnosed with CRC, resulting in about 50,000 deaths. Despite the gradual decline in CRC incidence rates in the United States from 2019 to 2023, a trend of increase in CRC prevalence among younger age groups is also observed. CRC is often diagnosed at an advanced stage, contributing to rising global incidence rates.1,2 The increase in CRC cases has been linked to factors such as diet, sex differences, geographical factors, and genetic mutations.3 Early diagnosis and treatment of CRC remain challenging.4 American Society of Clinical Oncology (ASCO) guidelines for treatment of metastatic CRC5 recommend the combination of anticancer medications with chemotherapy. However, these conventional treatments often cause significant side effects, affecting the physical and mental well-being of the patients.6
More than half of colorectal-cancer patients eventually develop metastatic disease (mCRC), for whom the 5-year overall survival remains approximately 13%—a figure that underscores the formidable clinical challenge posed by tumour dissemination7 The epithelial-mesenchymal transition (EMT) in CRC tissues contributes to increased cell invasion and migration. During EMT, CRC cells gradually lose their adhesion ability, leading to reduced cell-cell adhesion and fostering a more invasive phenotype.8 Suppression of EMT cells is therefore critical for preventing and treating metastatic CRC. The Wnt/β-catenin signaling pathway plays a significant role in various intracellular processes such as macrophage polarization exosome formation,9 tumor angiogenesis,10 and ferroptosis.11 Activation of Wnt/β-catenin signaling in CRC leads to upregulation of EMT transcription factors such as Snail and Twist, which repress E-cadherin and induce mesenchymal markers like N-cadherin and vimentin.12 Wang et al conducted an RNA-seq analysis of gallbladder cancer and gallstone disease and found that ALPP is a significantly upregulated carcinogenic risk marker. KEGG enrichment analysis showed that the Wnt/β-catenin signaling pathway was involved. However, to date, no experimental studies have confirmed the existence of a direct regulatory axis between ALPP and Wnt signaling. Therefore, the mechanistic dissection of this hypothesized ALPP-Wnt interaction represents a high-value yet unresolved research niche.13
Alkaline phosphatase (ALP) is a type of enzyme widely present in various tissues of the human body. It encompasses multiple isoenzymes, such as the placental - type (ALPP/PLAP), intestinal- type, and liver/bone/kidney-type. These isoenzymes exhibit certain differences in structure and function, and their expression patterns vary in different tissues and disease states.14 Recent research suggests that ALPP is abnormally expressed in breast cancer,15,16 potentially serving as a biomarker for early diagnosis. ALPPL2 has been identified as a potential adverse prognostic indicator of gastric cancer.17,18 High PLAP expression is associated with advanced pathological tumor stage (P = 0.0086), lymph node metastasis (P = 0.0085), lymphatic vessel (P = 0.0007) and vascular invasion (P = 0.0222) in colorectal cancer.19 Additionally, ALPP is highly expressed in the placental trophoblast layer and germ cells,20 with minimal or negligible expression in other tissues. This highlights the potential of therapies targeting ALPP to be remarkably specific for cancer treatment. Li et al21 utilized ALPP as the sole target in CAR-T cell therapy, demonstrating its efficacy and multifaceted therapeutic benefits in CRC. Therefore, due to its high specificity and restricted expression in healthy tissues, targeting ALPP could mitigate the off - target effects of current therapies that often damage normal cells (eg, chemotherapy - induced gastrointestinal toxicity). However, the effect of ALPP on CRC cells, and the mechanisms involved remain to be investigated. To this end, this study aims to elucidate the specific effects of ALPP on CRC cells, particularly those on the Wnt/β-catenin signaling pathway, and thereby to provide a theoretical basis for ALPP as a targeted therapy for CRC.
Materials and Methods
Cell Culture
Human CRC HT29 and HCT116 cell lines and human colonic epithelial NCM460 cells were procured from the Cell Bank of the Chinese Academy of Sciences, Shanghai, China (All cell lines were authenticated by short tandem repeat (STR) profiling and routinely verified as mycoplasma-negative prior to experimentation). The cells were cultured in high-glucose DMEM (11995, Gibco Life Technologies, China) containing 10% fetal bovine serum (YSN0121, ExCell Bio, China) and 1% penicillin-streptomycin solution (P1400, Beijing Solarbio Science & Technology Co., Ltd, China). Cultures were maintained in a 37°C, 5% CO2 incubator (CI-191C, Jiemei Electronics, China), with media changed every two days. The cells were allowed to grow naturally until reaching exponential phase for subsequent experiments.
Cell Transfection
After HT29 and HCT116 have reached a density of 80% during the exponential phase, the medium was removed, and the cells were rinsed with PBS three times. Then, trypsin (P6730, Beijing Solarbio Science & Technology Co., Ltd, China) was added for digestion for 3 min. Digestion was terminated by adding complete DMEM, and the medium containing cells were centrifuged at 1200 g for 3 min. After that, the cells were resuspended in an appropriate amount of culture medium, and 5×105 cells per well were inoculated into a six-well plate. After incubation overnight, Lipofectamine™ 2000 (11668019, Thermo Fisher Scientific, China) pre-mixed with ALPP interference RNA or ALPP plasmid DNA was added to the cells along with serum-free DMEM high-glucose medium according to the transfection kit instructions. Transfection efficiency was confirmed by qPCR and Western blot 48 h post-transfection. In the validation experiment of ALPP targeting the Wnt/β - catenin pathway, after transfecting with siRNA for 48 hours, we added 0.5 μmol /L Wnt/β - catenin agonist 1 (an activator of the Wnt/β - catenin pathway, HY - 114321, MedChemExpress, China) and continued the cultivation for another 48 hours. After the cultivation was completed, the expression levels of proteins related to the Wnt/β - catenin pathway were detected through a Western blot experiment.
Differentially Expressed Genes in Metastatic and Non-Metastatic CRC Cells
Expression data (mRNA expression count values) for TCGA Colon Cancer (GDC TCGA Colon Cancer) were obtained from the UCSC Xena database (https://xenabrowser.net/datapages) (The data in this study were sourced from public platforms, and the data collection and processing methods complied with the regulations on exemption from ethical review in the “Measures for Ethical Review of Human - related Life Science and Medical Research” in China. Specifically, in accordance with Items (1) and (2) of Article 32 of these measures, the research used legally disclosed and anonymized data without involving personal privacy. Therefore, the research protocol was exempt from ethical review and approval). Differential expression analysis of metastatic and non-metastatic CRC samples was performed using the R package DeSeq.22 Genes with |log2FC| > 1 and FDR < 0.05 were selected. CRC metastasis-related targets were extracted from the GeneCards database (https://www.genecards.org). The intersection between this list and the previously obtained differentially expressed genes was determined. A PPI network diagram was established for the resulting list using the STRING database (https://string-db.org) and visualized using Cytoscape software. Univariate and multivariate COX analyses were performed as well.
Survival Analysis
Survival prognostic analysis of NTF4, LRP2, CRHR1, FABP4, CTXND1, and ALPP-related genes in CRC patients assessed using the TCGA cohort. Patients with CRC were categorized into high- and low-expression groups based on median gene expression levels. KM survival analysis for different patient groups was conducted using the R packages “Survminer” and “survival”23 Box plots were generated using the R package “ggpubr”, and Wilcox tests were employed to compare expression differences between different patient groups.24 Patients were categorized into metastasis and non-metastasis groups as follows: patients with uncertain M and N stages were excluded; patients with M stages other than M0 or N stages other than N0 were categorized as having metastasis; and patients with both M and N stages equal to M0 and N0 were categorized as non-metastatic.
CCK8 Assay
The transfected HT29 and HCT116 cells were retrieved, digested, and counted as described above. Cells were seeded at 3000 cells per well in a 96-well plate, with five replicates per group. A control group devoid of cells was established by adhering to procedures identical to those used in the experimental group. The plate was placed in a cell incubator, and 10 μL of CCK8 solution was introduced to each well at intervals of 24 h, 48 h, and 72 h. After incubation for 3 h, absorbance at 450 nm was measured using a microplate reader (Flash, ReadMax 1200).
Transwell Assay
The transwell chambers were coated with Matrigel matrix (C0371, Beyotime Biotech Inc, China). The Matrigel matrix gel was diluted in serum-free DMEM at a 1:10 ratio. A total of 100 μL of diluted Matrigel matrix gel was introduced to the upper chamber of the Transwell and incubated for 4 h in a cell incubator. After 4 h, the Transwell chambers were retrieved. In the lower chamber of the Transwell, each well was filled with 500 μL of high-glucose DMEM containing 20% FBS. Successfully transfected HT29 and HCT116 cells were counted as described previously. For the invasion assay, the cells were diluted to a concentration of 3×106 cells/mL. For the migration assay, the cells were diluted to a concentration of 1×106 cells/mL. Subsequently, 100 μL of the cell suspension was carefully introduced into the upper chamber containing the gel-coated surface. The Transwell chambers were then placed in a cell incubator and incubated for 12 h. After incubation, the chambers were washed thrice with PBS, and then each well in the lower chamber was filled with 500 μL of methanol solution. After 25 min of incubation, the methanol solution was discarded and the wells were thoroughly rinsed three times with PBS. A 0.5% crystal violet solution was added to each well of the lower chamber for 5 min, and excess crystal violet was washed away with PBS. The cells in the upper part of the chamber were gently wiped with a cotton swab and allowed to air dry. Five random fields were selected for photography under a microscope. The number of cells with invasive capabilities was analyzed using the Image J 1.5.2a software.
qPCR Experiment
Transfected and non-transfected HT29, HCT116, and NCM460 cells were collected using sterile cell scrapers and placed in 1.5 mL centrifuge tubes. One milliliter of TRIzol (15596026, Beijing Solarbio Science & Technology Co., Ltd, China) was added, and the tubes were left at room temperature for 5 min. Subsequently, 0.2 mL of chloroform was introduced into the tubes, which were gently inverted multiple times before being allowed to stand at room temperature for a period 3 min. Following this, the samples underwent centrifugation at 12000 g and 4°C for a duration of 15 min. Subsequently, 500 μL of isopropanol was introduced to the samples and thoroughly mixed. After 10 min of static incubation at room temperature, the samples underwent a subsequent centrifugation at 12000 g and 4°C for a duration of 10 min. The resulting supernatant was meticulously removed and 1 mL of 75% ethanol was added. Following this addition, the samples were subjected to centrifugation at 5000 g and 4°C for 5 min, and the ensuing supernatant was subsequently discarded. Upon unsealing the cap of the centrifuge tube, a brief period of 10 min was allowed for the samples to stabilize at room temperature. Subsequently, a suitable quantity of DEPC-treated water (W293452, Shanghai aladdin Biochemical Technology Co., Ltd, China) was introduced into the samples. The samples were either used immediately or stored in a −80°C freezer. RNA concentration was measured using a UV-Vis spectrophotometer (Q5000), and cDNA concentration was determined using a reverse transcription kit (KR118, Tiangen Biotech Co., Ltd, China). Real-time fluorescence quantitative amplification was performed using a QPCR detection kit (FP313, Tiangen Biotech Co., Ltd, China) on a real-time fluorescence quantitative instrument (Bio-RAD, CFX Connect). The thermal cycling conditions are as follows: thermostatic section 95°C, 15 min, cycling section 95°C, 10s; 60°C, 32s, dissolution section 95°C, 15s; 60°C, 1 min; 95°C, 30s, infinite thermostatic section 60°C ∞. The original data were retained for subsequent analyses. The relative expression level is represented by 2 −ΔΔCt, where ΔCt represents the difference between the Ct value of the target gene and that of the internal reference GAPDH.25 ALPP interference RNA, SiNC 5ʹ-UUCUCCGAACGUGUCACGUTT-3ʹ (sense); 5ʹ-ACGUGACAGGUUCGGAGAATT-3ʹ (antisense); SiALPP-1 5ʹ-GACAUACAAUGUAGACAAACA-3ʹ (sense); 5ʹ-UUUGUCUACAUUGUAUGUCUU-3ʹ (antisense) SiALPP-2 5ʹ-CGACCAUGGUCAUCAUGAAAG-3ʹ (sense); 5ʹ-UUCAUGAUGACCAUGGUCGAU-3ʹ (antisense) SiALPP-3 5ʹ-CGGUCCUCCUAUACGGAAACG-3ʹ (sense); 5ʹ-UUUCCGUAUAGGAGGACCGUG-3ʹ (antisense). Primer sequences for qPCR are provided in Table S1. The pcDNA3.1 (+) plasmid was obtained from addgene Company, and the pcDNA3.1 (+) -ALP primer sequence is as follows: pcDNA3.1(+)-ALPP-F:TACCGAGCTCGGATCCATGCTGGGGCCCTGCATGCTG; pcDNA3.1(+)-ALPP-R:GATATCTGCAGAATTCTCAGGGAGCAGTGGCCGTC.
Western Blot Experiment
After 48 h of transfection, HT29 and HCT116 cells were collected using sterile cell scrapers and placed in 1.5 mL centrifuge tubes. A total of 100 μL of cell lysis buffer (R0010, Beijing Solarbio Science & Technology Co., Ltd, China) containing 1% protease inhibitor (P0100, Beijing Solarbio Science & Technology Co., Ltd, China) and 1% protein phosphatase inhibitor (P1260, Beijing Solarbio Science & Technology Co., Ltd, China) was added, and the tubes were left on ice for 25 min. The tubes were vortexed for 20s every 5 min and then centrifuged at 12,000 g for 30 min at 4°C. The supernatant was collected and stored in a −80°C freezer or used immediately. The protein concentration was determined using a BCA protein quantification kit (PC0020, Beijing Solarbio Science & Technology Co., Ltd, China). Equal amounts of protein were loaded onto gels for electrophoresis. Appropriate amounts of protein lysis buffer and 5× buffer (P1040, Beijing Solarbio Science & Technology Co., Ltd, China) were added to adjust the protein concentration for each group. The mixture was then placed in a 95°C water bath and allowed to stand for 15 min. Afterward, it was cooled to room temperature and stored at −80°C. After loading equal amounts of protein samples onto the gel, gel electrophoresis was initiated with a voltage set at 60 V for 45 min, followed by adjustment of the voltage to 120 V, and a subsequent 60-minute waiting period before electrophoresis was completed. Proteins were transferred onto a PVDF membrane for 1 h at a constant current of 260 mA. The membrane was then blocked with 5% skim milk powder for 2 h and was subjected to an overnight incubation at a temperature of 4°C, with appropriate primary antibodies. The secondary antibody was prepared using 5% skim milk powder and incubated for 1 h. The membrane was washed three times with 1× TBST solution, each lasting 3 min. A chemiluminescence reagent was added and the gel was imaged using a gel imager. The ImageJ software was used for subsequent data processing and analysis. Antibodies used: ALPP (ab133602, Abcam, 1:1000), GAPDH (AF7021, Affinity Biosciences, 1:10,000), N-cadherin (AF5239, Affinity Biosciences, 1:1000), E-cadherin (BF0219, Affinity Biosciences, 1:1000), Vimentin (AF7013, Affinity Biosciences, 1:1000), ALPP (DF9033, Abcam, 1:1000), GSK3β (ab32391, Abcam, 1:1000), p-GSK3β (ab75814, Abcam, 1:1000), Snail (AF6032, Affinity Biosciences, 1:1000), β-catenin (ab32572, Abcam, 1:1000), Goat anti-rabbit (K008, Bioss, 1:5000), Goat anti-mouse (K009, Bioss, 1:5000), APC (ab40778, Abcam, 1:1000).
Statistical Analysis
Data were analyzed using SPSS 22.0 software. Measurement data were expressed as mean ± standard deviation (SD). Unpaired t-test was used to test the expression of genes in the TCGA database for CRC patients with and without metastasis. All other data were analyzed by one-way ANOVA followed by post hoc comparison with Tukey’s honestly significant difference test. P < 0.05 indicates a statistically significant difference.
Results
Identification of Potential Targets Influencing Colorectal Cancer Metastasis Through Bioinformatics Analysis
We first used the UCSC Xena database (https://xenabrowser.net/datapages/) to access the TCGA CRC COAD (GDC TCGA Colon Cancer) dataset, and extracted mRNA expression count values. Then, we performed a differential expression analysis using the DeSeq2R package to identify the differences in gene expression between metastatic and non-metastatic CRC samples. Only genes meeting |log2FC| > 1 and FDR < 0.05 were included, which resulted in 585 differentially-expressed genes (152 up-regulated and 433 down-regulated). A volcano plot (Figure 1A) and a heatmap showing a hierarchical cluster analysis (Figure 1B) were generated to visualize the results.
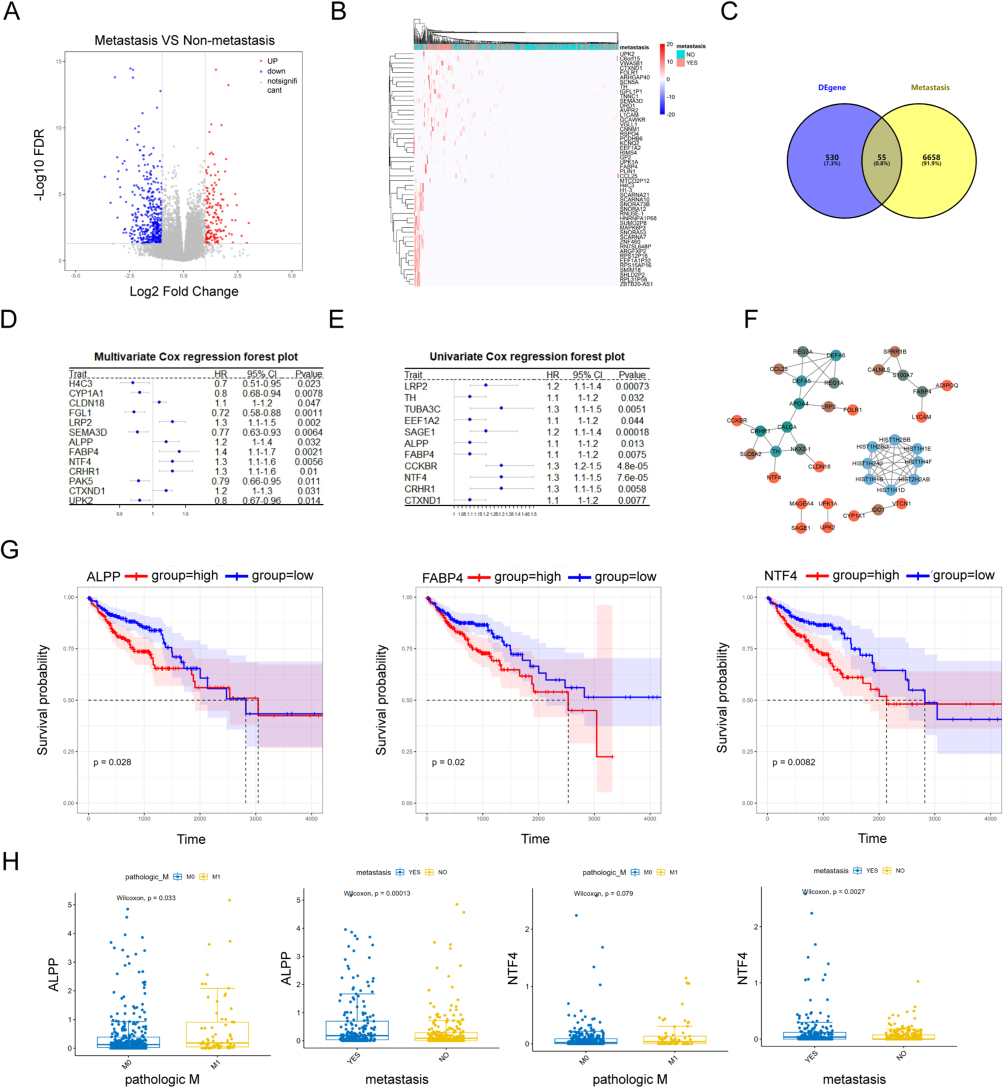 |
Figure 1 Selection of potential therapeutic targets against metastatic CRC based on bioinformatics analysis. (A) Volcano plot (B) clustering heatmap (C) Venn diagram showing intersection of identified genes between GeneCards and UCSC Xena databases (D) forest plot showing intersection genes for single-factor COX regression analysis) (E) forest plot showing intersection genes for multi-factor COX regression analysis) (F) protein-protein interaction (PPI) network, constructed using intersection genes (G) KM survival curves for NTF4, FABP4, and ALPP in TCGA CRC cohort (H) box plots of ALPP and NTF4 expression in TCGA CRC Cohort, stratified by metastasis status and M0/M1 stage. |
Next, we used the GeneCards database (https://www.genecards.org/) to identify a list of 6713 CRC metastasis-related protein-coding genes. A total of 55 genes in this list were found to be present in the previously identified differentially-expressed gene list as well (Figure 1C). A univariate COX analysis on expression levels of these genes revealed 11 statistically significant genes, all of which were identified as risk factors for CRC metastasis (hazard ratio [HR] = 1). A multivariate COX analysis identified 13 significant genes. Among these, six genes consistently appeared to be significant risk factors in both univariate and multivariate COX analyses: NTF4, LRP2, CRHR1, FABP4, CTXND1, and ALPP (Figure 1D and E). Subsequently, a PPI network was constructed using the STRING database (https://string-db.org/), and visualized using the Cytoscape software. The network comprised 37 genes connected by 61 interaction pairs (Figure 1F), yet did not provide any useful insights.
Based on this preliminary analysis, NTF4, LRP2, CRHR1, FABP4, CTXND1, and ALPP were selected as key genes related to metastasis within the TCGA CRC cohort. Samples in the dataset were categorized based on median expression levels of these genes, facilitating the exploration of the relationship between gene expression and prognosis. In the Kaplan - Meier (KM) survival analysis, significant differences were observed in the expression levels of NTF4, FABP4, and ALPP, indicating that high expression levels of these genes were associated with poor prognosis (Figure 1G). These results support the potential of ALPP and other factors as prognostic biomarkers. Detailed survival curves for LRP2, CRHR1, and CTXND1 are provided in Figure S1A. Furthermore, expression levels of ALPP and NTF4 were found to be significantly higher in metastatic samples, compared to levels in non-metastatic CRC samples. In particular, expression of ALPP was more pronounced between M0 and M1 stages, with significantly increased expression in the M1 stage (Figure 1H). A detailed depiction of stage-specific expression differences of FABP4 is provided in Figure S1B. Next, quantitative real-time PCR experiments were conducted to investigate and validate the differences detected in expression of the identified potential metastasis-related target genes in both CRC and intestinal epithelial cells. As expected, the results demonstrated that ALPP mRNA levels showed most pronounced differences between CRC specifically HCT116 and HT29 cells, and the intestinal epithelial cell line NCM460. mRNA levels of LRP2, CRHR1, FABP4, and CTXND1 were also found to be upregulated in CRC cells, corroborating our findings in the previous section (Figure S1C). We also found that the expression level of ALPP protein was significantly higher in HCT116 and HT29 cells compared with that in NCM460 cells (Figure S1D).
ALPP Promotes Proliferation of HCT116 and HT29 Cells
Drawing upon the insights gained from both bioinformatics analysis and qPCR experiments, we next focused on ALPP for subsequent investigations. To investigate the role of ALPP in CRC, we generated ALPP-overexpressing and knock-down cell lines from HCT116 and HT29 cell lines. Western blot and qPCR analyses revealed that 48 h post-transfection of ALPP overexpression plasmids, ALPP mRNA and protein levels both increased (Figure 2A and B). In contrast, transfection with ALPP-interfering RNA (si-ALPP) resulted in decreased mRNA and protein levels of ALPP in both cell lines after 48 h (Figure 2C and D). The most significant reductions were those of Si-ALPP-1 and Si-ALPP-2 in HT29 and HCT116 cells, prompting us to select these two siRNAs for subsequent experiments. Transfection with si-ALPP was found to decrease ALPP protein expression. To validate the effect of ALPP on CRC cell proliferation, CCK8 assay was conducted. These results indicated that ALPP promoted the proliferation of HCT116 and HT29 cells in a time-dependent manner (Figure 2E and F). Taken together, our comprehensive analysis in this section highlighted the pivotal role of ALPP in facilitating cell proliferation within the context of CRC. These effects collectively contribute to enhanced metastatic potential in CRC.
 |
Figure 2 ALPP induces proliferation of HT29 and HCT116 cells. (A) qPCR analysis of ALPP mRNA expression levels after transfection with overexpression plasmids. (B) Western blot experiment to detect protein expression levels of ALPP after overexpression plasmid transfection. (C) QPCR analysis of ALPP mRNA expression levels after transfection with interfering RNA. (D) Western blot experiment to detect protein expression levels of ALPP after interfering RNA transfection. (E) CCK8 assay to assess changes in cell viability after ALPP overexpression. (F) CCK8 assay to assess changes in cell viability after ALPP downregulation. (When compared between the two groups, **P<0.01, ***P<0.001, ****P<0.0001). |
ALPP Induces EMT in HCT116 and HT29 Cells
Having substantiated the ability of ALPP to induce CRC cell proliferation, we next conducted Transwell migration experiments to evaluate changes in invasion and migration capabilities. Our findings revealed a significant reduction in the invasive and migratory abilities of HT29 and HCT116 cells following ALPP knockdown. Conversely, ALPP overexpression clearly increased the invasive and migratory capacities of these cells (Figure 3A and B). We also employed immunofluorescence assays to examine the localization and expression of EMT-associated proteins, namely E-cadherin, N-cadherin, and Vimentin. The results indicated that E-cadherin and N-cadherin were predominantly expressed in the cytoplasm and nucleus, whereas vimentin was predominantly expressed in the cell membrane. Notably, ALPP downregulation led to an increase in E-cadherin expression, along with a reduction in N-cadherin and Vimentin expression. In contrast, ALPP overexpression reduced E-cadherin expression, and increased expression of N-cadherin and Vimentin (Figure 4A and B). The effect of ALPP on EMT-related protein experiment was further investigated by Western blot analysis of E-cadherin, N-cadherin, Snail, and Vimentin. As expected, ALPP overexpression resulted in a decrease in E-cadherin expression, and an increase in the expression levels of N-cadherin, Vimentin, and Snail. Vice versa, ALPP downregulation resulted in an opposite trend in the expression of EMT-related proteins (Figure 5A and B). In summary, these findings demonstrate the regulatory role of ALPP in modulating EMT-related proteins, and thereby in facilitation of EMT in CRC cells.
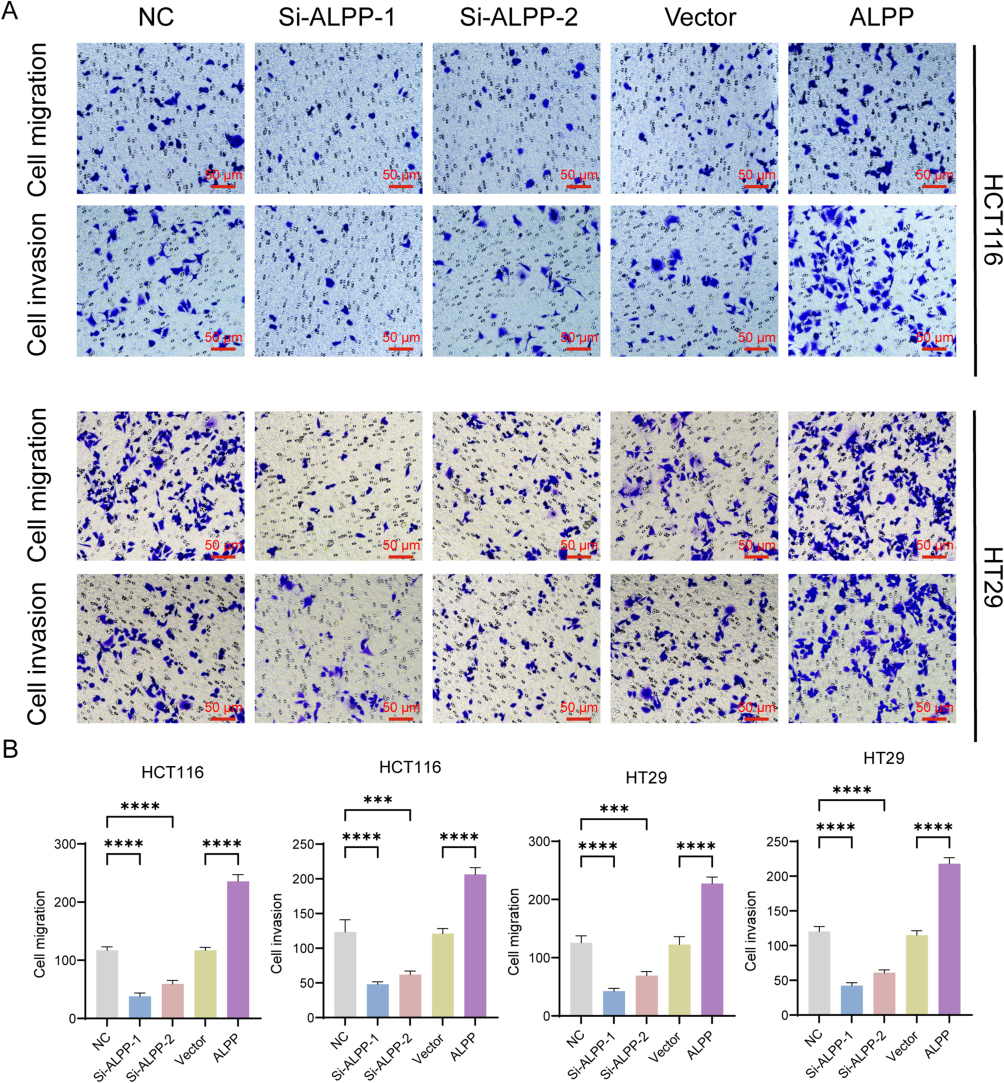 |
Figure 3 ALPP induces invasion and migration in HT29 and HCT116 cells. (A) Transwell assays to assess changes in the invasion and migration capabilities of HT29 and HCT116 CRC cells after transfection with overexpression plasmids or interfering RNA for 48 h (200× magnification, scale bar 50 μm). (B) Bar chart for counting the number of invasive and migratory cells in the Transwell experiment. (When compared between the two groups, ***P<0.001, ****P<0.0001). |
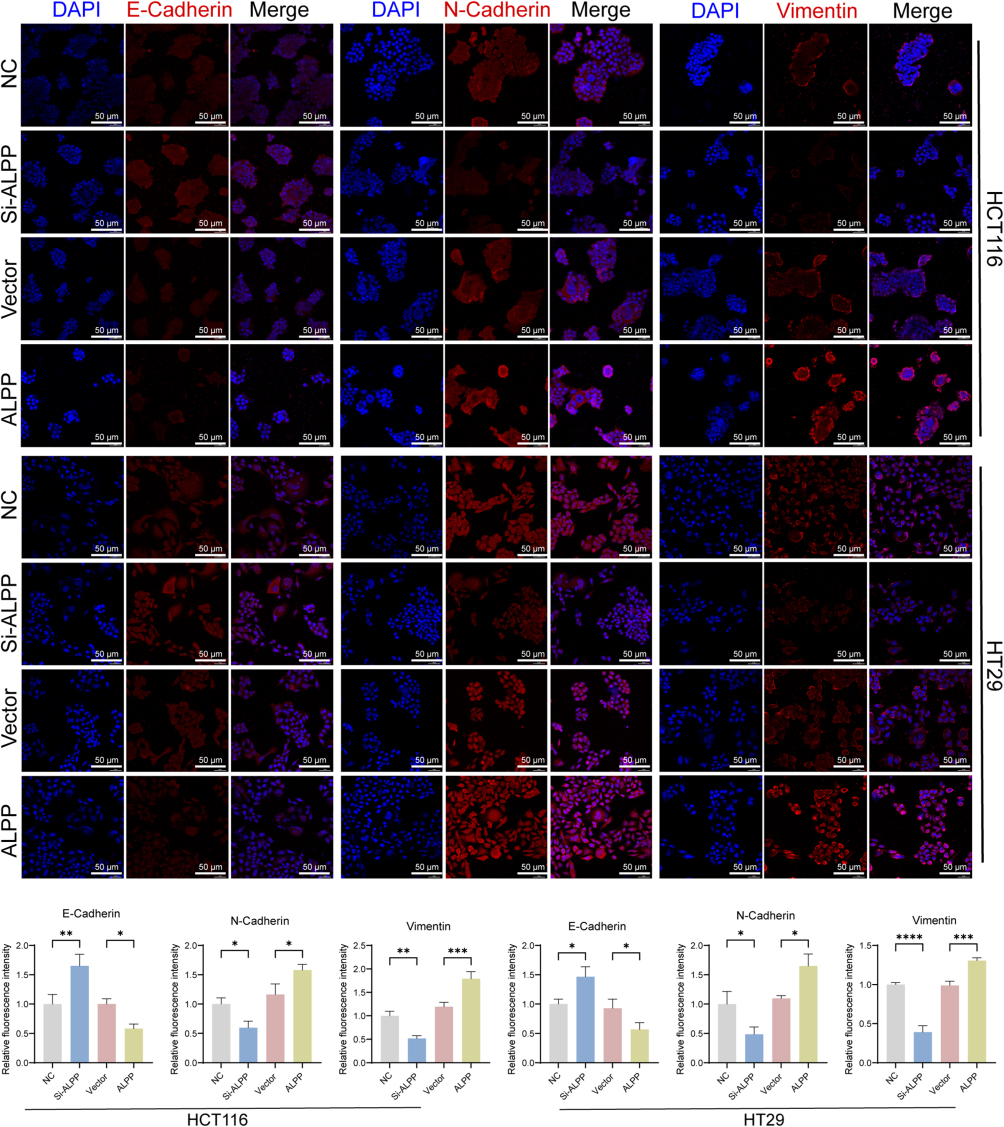 |
Figure 4 The expression and distribution of E - Cadherin, N - Cadherin, and Vimentin in HCT116 and HT29 cells. (A) An immunofluorescence assay was conducted in HT29 and HCT116 CRC cells after transfection with overexpression plasmids or interfering RNA for 48 h to examine the expression, localization, and levels of E-cadherin, N-Cadherin, and Vimentin (200× magnification, scale bar 50 μm). (B) Bar chart for statistical analysis of the average fluorescence intensity of E-cadherin, N-Cadherin, and Vimentin detected by immunofluorescence. (When compared between the two groups, *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). |
 |
Figure 5 ALPP regulates EMT-related protein expression. (A) Western blot assay was performed in HCT116 and HT29 cells to determine the protein expression levels of ALPP, E-cadherin, N-cadherin, Vimentin, and Snail. (B) Bar chart for statistical analysis of the relative expression levels of ALPP, E-cadherin, N-cadherin, Vimentin, and Snail proteins in the Western blot experiment.(When compared between the two groups, *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). |
ALPP-Mediated Activation of the Wnt/β-Catenin Signaling Pathway
While our investigations substantiated the ability of ALPP to induce EMT in CRC cells, the precise signaling pathways involved remain elusive. In order to understand the potential impact of ALPP on signaling cascades, we focused on on the Wnt/β-catenin pathway. Using Western blot analysis, we determined the expression patterns of key Wnt/β-catenin pathway proteins, specifically APC, β-catenin and p-GSK3β. Our findings revealed that ALPP overexpression downregulated APC and upregulated β-catenin and p-GSK3β expression. ALPP downregulation further induced APC activation, which subsequently inhibited β-catenin and p-GSK3β expression (Figure 6A). In summary, ALPP activates the Wnt/β-catenin signaling pathway, reinforcing its role in promoting EMT in CRC cells lines HT29 and HCT116. In HCT116 and HT29 cells, after transfecting with siRNA for 48 hours, 0.5 μmol/L of Wnt/β-catenin agonist 1 (an activator of the Wnt/β-catenin pathway) was added and incubation was continued for another 48 hours. Subsequently, the relative expression levels of APC, GSK3β, p - GSK3β, and β-catenin proteins were detected through Western blot analysis. The results showed that, in HCT116 and HT29 cells, compared with Wnt/β-catenin agonist 1 alone, the addition of Si-ALPP + Wnt/β - catenin agonist 1 could significantly inhibit the ratio of β-catenin and p-GSK3β/GSK3β (Figure 6B).
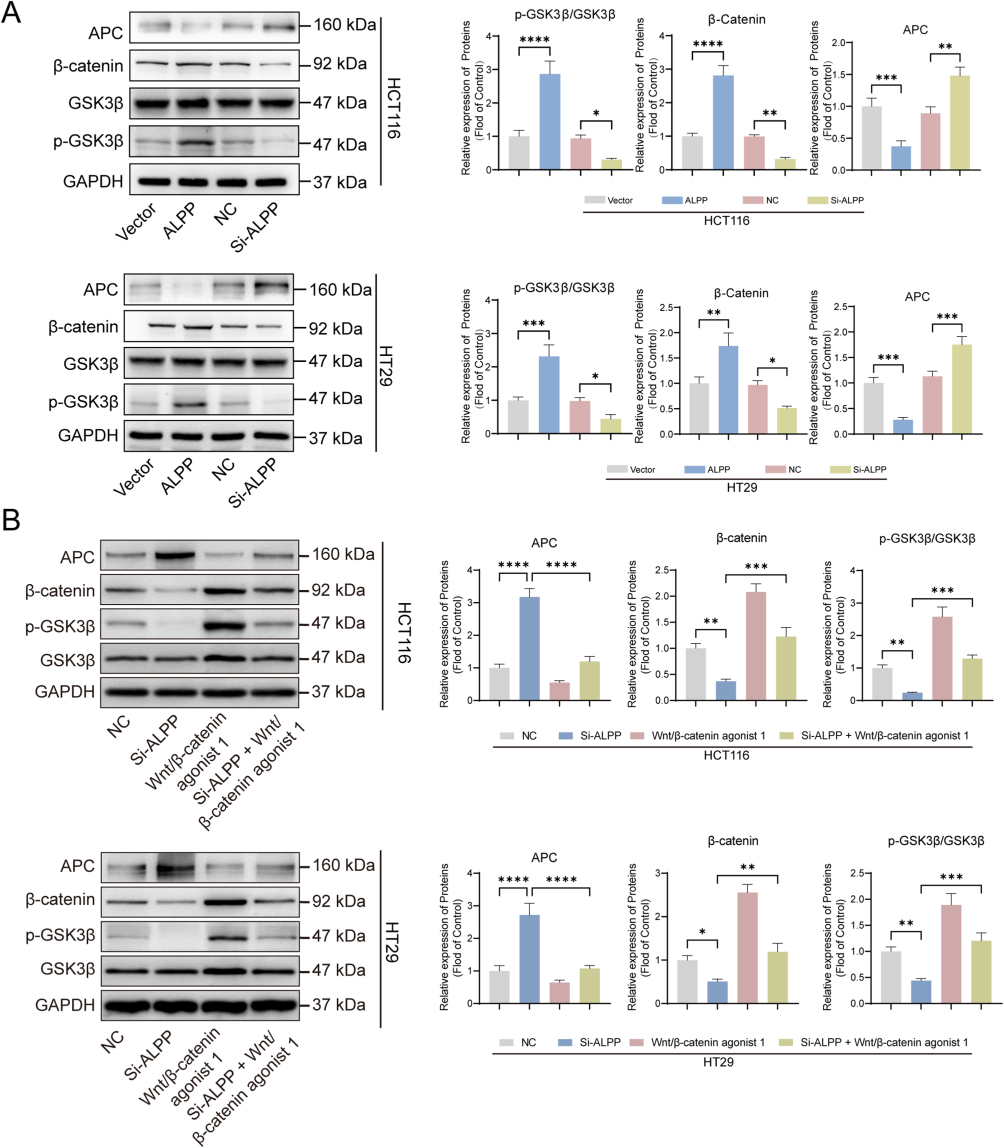 |
Figure 6 ALPP activates Wnt/β-catenin signaling pathway. (A) After transfecting the over-expression and knockdown sequences of ALPP into HCT116 and HT29 cells and incubating for 48 hours, the expression levels of proteins including APC, GSK3β, p-GSK3β, and β - catenin were detected by Western blot. (B) In HCT116 and HT29 cells, after transfecting with siRNA for 48 hours, 0.5 μmol/L of Wnt/β-catenin agonist 1 (an activator of the Wnt/β -catenin pathway) was added and incubation was continued for another 48 hours. Subsequently, the relative expression levels of APC, GSK3β, p-GSK3β, and β-catenin proteins were detected through Western blot analysis.(When compared between the two groups, *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). |
Discussion
Metastasis remains a key factor in high recurrence rates, mortality, and challenging therapeutic outcomes in CRC. While existing treatments effectively eliminate tumor cells, they can also damage adjacent tissues, leading to significant side effects. Thus, there is a need for more effective and targeted therapeutic strategies.26,27 To this end, we investigated CRC metastasis-associated differentially-expressed genes, and identified NTF4, LRP2, CRHR1, FABP4, CTXND1, and ALPP as potential biomarkers for predicting CRC metastasis, with ALPP showing the most pronounced differences. Unlike other potential targets, ALPP is primarily expressed in germ cells and is almost absent in normal cells, highlighting the potentially high efficiency and specificity of therapies targeting ALPP.26 Thus, targeting ALPP against CRC metastasis therapy could minimize harm to normal cells. The main contribution of this study is the pioneering exploration of the effect of ALPP on EMT and the Wnt/β-catenin signaling pathway in CRC cells. To this end, this study provides a theoretical foundation for the development of ALPP-targeting therapeutics.
Our findings revealed that ALPP overexpression promotes proliferation, invasion, and migration of HT29 and HCT116 CRC cells, whereas interfering with ALPP expression suppresses these cellular behaviors. These results clearly demonstrate the role of ALPP as an oncogene in CRC cells, in line with observations in previous related research.22 We also investigated the relationship between ALPP and the EMT. E-cadherin, N-cadherin, Snail, and Vimentin are well-known EMT markers.28 Loss of E-cadherin and aberrant N-cadherin and Vimentin expression, which are prevalent in most tumors, drive cancer cell metastasis.29,30 According to the literature, the transcription factor Snail, acting as a “master switch”, directly inhibits the expression of E-cadherin and activates the transcription of N-cadherin and Vimentin. This leads to the disintegration of cell adhesion junctions and the acquisition of a mesenchymal phenotype. The down-regulation of E-cadherin weakens epithelial integrity, the up-regulation of N-cadherin enhances cell migration, and Vimentin remodels the cytoskeleton. These processes work together to drive cells to detach from the epithelial layer and acquire the ability to invade and metastasize.31 Our findings revealed that increasing ALPP expression was found to lead to decreased E-cadherin expression while elevating N-cadherin and Vimentin levels, ultimately culminating in enhanced invasiveness of HCT116 and HT29 cells. ALPP likely induced EMT in these cells, diminished cell adhesion and increased their invasive potential. It is well known that during the process of EMT, the cells will form a spindle morphology.32 However, we did not observe the appearance of the recognized morphology in the immunofluorescence experiments, and we suspected that it might be related to the characteristics of the cells,33 or it might be possible that the transfection reagents and the treatment of other reagents would affect the morphology changes.
The Wnt/β-Catenin signaling pathway is crucial for human embryonic development. CRC is often triggered by inactivating mutations in the tumor suppressor gene APC, which in turn leads to activation of the Wnt signaling pathway.34,35 Targets that inhibit this pathway have emerged as novel therapeutic strategies against CRC.36 Based on these developments, our investigation of the relationship between the oncogenic ALPP and the Wnt/β-catenin signaling pathway is particularly relevant. As expected, ALPP down-regulation activated the expression of APC proteins, which in turn inhibited the Wnt/β-Catenin signaling pathway. Therefore, we hypothesize that ALPP potentially impacts the CRC cell EMT process via the Wnt/β-catenin signaling pathway. However, further in vitro experiments are required to confirm this hypothesis.
Despite the significance of our findings, there are also limitations to this research. First, the selection of six CRC metastasis-associated targets was validated through in vitro experiments without confirmation by clinical data. Additionally, the precise regulatory relationship between ALPP and the Wnt/β-catenin signaling pathway requires further clarity, necessitating reverse validation experiments. It has been shown that the β-catenin/TCF4 complex induces epithelial mesenchymal transition (EMT) and that β-catenin undergoes obligatory nuclear allosteric translocation during this process.33 We hypothesize that this may be a potential mechanism by which ALPP plays a regulatory role, which cannot be verified by our current conclusions, and we will subsequently extract nuclear and cytoplasmic proteins separately to examine β-catenin expression. We will verify whether there is an interaction relationship between β-catenin and TCF4 through COIP experiments, and further explore the mechanism study of ALPP action in CRC. Initially, we restricted our functional assays to HCT116 and HT29 cells; although these lines are widely used, they cannot fully capture the molecular heterogeneity of colorectal cancer. To enhance the generalizability of our findings, we will expand the validation cohort to include additional genomically distinct models such as SW480. Finally, although ALPP can modulate the expression of EMT-associated proteins, EMT is a dynamic outcome influenced by interactions with the microenvironment or other cells.37 The intricate nature of tumor EMT necessitates further experiments to elucidate the direct interplay between ALPP and EMT. Our study reveals, for the first time, role of ALPP in promoting CRC cell proliferation and invasion, and thereby constitutes a preliminary report of signaling pathways associated with this process. Moreover, potential biomarkers for distinguishing between metastatic and non-metastatic CRC have also been identified.
Conclusion
In summary, ALPP acts as an oncogenic factor in colorectal cancer cell lines HT29 and HCT116, promoting cell proliferation and facilitating EMT. These effects collectively contribute to enhanced metastatic potential in CRC to the Results section. ALPP induces abnormal activation of the Wnt/β-catenin signaling pathway.
Acknowledgments
We are grateful to the Second Hospital of Hebei Medical University for providing the experimental site and to the members of the group for their support.
Funding
Key Project of the Natural Science Foundation of Hebei Province (H2020206485).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel RL, Wagle NS, Cercek A, Smith RA, Jemal A. Colorectal cancer statistics, 2023. CA Cancer J Clin. 2023;73(3):233–254. doi:10.3322/caac.21772
2. Kim BJ, Hanna MH. Colorectal cancer in young adults. J Surg Oncol. 2023;127(8):1247–1251. doi:10.1002/jso.27320
3. Deng Y, Wang L, Huang J, Ding H, Wong MCS. Associations between potential causal factors and colorectal cancer risk: a systematic review and meta-analysis of Mendelian randomization studies. J Dig Dis. 2022;23(8–9):435–445. doi:10.1111/1751-2980.13130
4. Morgan E, Arnold M, Gini A, et al. Global burden of colorectal cancer in 2020 and 2040: incidence and mortality estimates from GLOBOCAN. Gut. 2023;72(2):338–344. doi:10.1136/gutjnl-2022-327736
5. Zhao W, Dai S, Yue L, et al. Emerging mechanisms progress of colorectal cancer liver metastasis. Front Endocrinol. 2022;13:1081585. doi:10.3389/fendo.2022.1081585
6. Morris VK, Kennedy EB, Baxter NN, et al. Treatment of metastatic colorectal cancer: ASCO guideline. J Clin Oncol. 2023;41(3):678–700. doi:10.1200/jco.22.01690
7. Zhao S, Ly A, Mudd JL, et al. Characterization of cell-type specific circular RNAs associated with colorectal cancer metastasis. NAR Cancer. 2023;5(2):zcad021. doi:10.1093/narcan/zcad021
8. Tan Z, Sun W, Li Y, et al. Current progress of EMT: a new direction of targeted therapy for colorectal cancer with invasion and metastasis. Biomolecules. 2022;12(12):1723. doi:10.3390/biom12121723
9. Chi C, Hou W, Zhang Y, et al. PDHB-AS suppresses cervical cancer progression and cisplatin resistance via inhibition on Wnt/β-catenin pathway. Cell Death Dis. 2023;14(2):90. doi:10.1038/s41419-022-05547-5
10. Zhang X, Dong N, Hu X. Wnt/β-catenin Signaling Inhibitors. Curr Top Med Chem. 2023;23(10):880–896. doi:10.2174/1568026623666230303101810
11. Wang H, Zhang H, Chen Y, et al. Targeting Wnt/β-catenin signaling exacerbates ferroptosis and increases the efficacy of melanoma immunotherapy via the regulation of MITF. Cells. 2022;11(22):3580. doi:10.3390/cells11223580
12. Yao H, Ren D, Wang Y, et al. KCTD9 inhibits the Wnt/β-catenin pathway by decreasing the level of β-catenin in colorectal cancer. Cell Death Dis. 2022;13(9):761. doi:10.1038/s41419-022-05200-1
13. Wang J, Xu C, Cheng Q, et al. RNA sequencing revealed signals of evolution from gallbladder stone to gallbladder carcinoma. Front Oncol. 2020;10:823. doi:10.3389/fonc.2020.00823
14. Tyagi K, Venkatesh V. Emerging potential approaches in alkaline phosphatase (ALP) activatable cancer theranostics. RSC Med Chem. 2024;15(4):1148–1160. doi:10.1039/d3md00565h
15. Abreu RDS, Antunes D, Moreira ADS, et al. Next generation of ovarian cancer detection using aptamers. Int J Mol Sci. 2023;24(7):6315. doi:10.3390/ijms24076315
16. Li G, Hu X, Nie P, et al. Lipid-raft-targeted molecular self-assembly inactivates YAP to treat ovarian cancer. Nano Lett. 2021;21(1):747–755. doi:10.1021/acs.nanolett.0c04435
17. Liu S, Mao Q, Xue W, et al. High expression of ALPPL2 is associated with poor prognosis in gastric cancer. Hum Pathol. 2019;86:49–56. doi:10.1016/j.humpath.2018.11.019
18. Bassi G, Favalli N, Pellegrino C, et al. Specific inhibitor of placental alkaline phosphatase isolated from a DNA-encoded chemical library targets tumor of the female reproductive tract. J Med Chem. 2021;64(21):15799–15809. doi:10.1021/acs.jmedchem.1c01103
19. Reiswich V, Gorbokon N, Luebke AM, et al. Pattern of placental alkaline phosphatase (PLAP) expression in human tumors: a tissue microarray study on 12,381 tumors. J Pathol Clin Res. 2021;7(6):577–589. doi:10.1002/cjp2.237
20. Sun D, Qin Z, Xu Y, et al. The IVF-generated human embryonic microenvironment reverses progestin resistance in endometrial cancer cells by inducing cancer stem cell differentiation. Cancer Lett. 2022;526:311–321. doi:10.1016/j.canlet.2021.11.003
21. Li X, Berahovich R, Zhou H, et al. PLAP -CAR T cells mediate high specific cytotoxicity against colon cancer cells. Front Biosci. 2020;25(9):1765–1786. doi:10.2741/4877
22. Huang A, Sun Z, Hong H, et al. Novel hypoxia- and lactate metabolism-related molecular subtyping and prognostic signature for colorectal cancer. J Transl Med. 2024;22(1):587. doi:10.1186/s12967-024-05391-5
23. Zhao W, Liu M, Zhang M, et al. Effects of inflammation on the immune microenvironment in gastric cancer. Front Oncol. 2021;11:690298. doi:10.3389/fonc.2021.690298
24. Fang Y, Huang S, Han L, Wang S, Xiong B. Comprehensive analysis of peritoneal metastasis sequencing data to identify LINC00924 as a prognostic biomarker in gastric cancer. Cancer Manag Res. 2021;13:5599–5611. doi:10.2147/cmar.S318704
25. Zucha D, Kubista M, Valihrach L. Tutorial: guidelines for single-cell RT-qPCR. Cells. 2021;10(10):2607. doi:10.3390/cells10102607
26. Cervantes A, Adam R, Roselló S, et al. Metastatic colorectal cancer: ESMO clinical practice guideline for diagnosis, treatment and follow-up. Ann Oncol. 2023;34(1):10–32. doi:10.1016/j.annonc.2022.10.003
27. Zhao H, Ming T, Tang S, et al. Wnt signaling in colorectal cancer: pathogenic role and therapeutic target. Mol Cancer. 2022;21(1):144. doi:10.1186/s12943-022-01616-7
28. Babaei G, Aziz SG, Jaghi NZZ. EMT, cancer stem cells and autophagy; The three main axes of metastasis. Biomed Pharmacother. 2021;133:110909. doi:10.1016/j.biopha.2020.110909
29. Gundamaraju R, Lu W, Paul MK, et al. Autophagy and EMT in cancer and metastasis: who controls whom? Biochim Biophys Acta Mol Basis Dis. 2022;1868(9):166431. doi:10.1016/j.bbadis.2022.166431
30. Pastushenko I, Blanpain C. EMT transition states during tumor progression and metastasis. Trends Cell Biol. 2019;29(3):212–226. doi:10.1016/j.tcb.2018.12.001
31. Jiang M, Wang J, Li Y, et al. EMT and cancer stem cells: drivers of therapy resistance and promising therapeutic targets. Drug Resist Updat. 2025;83:101276. doi:10.1016/j.drup.2025.101276
32. Gurzu S, Silveanu C, Fetyko A, Butiurca V, Kovacs Z, Jung I. Systematic review of the old and new concepts in the epithelial-mesenchymal transition of colorectal cancer. World J Gastroenterol. 2016;22(30):6764–6775. doi:10.3748/wjg.v22.i30.6764
33. Zheng X, Carstens JL, Kim J, et al. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature. 2015;527(7579):525–530. doi:10.1038/nature16064
34. Aghabozorgi AS, Bahreyni A, Soleimani A, et al. Role of adenomatous polyposis coli (APC) gene mutations in the pathogenesis of colorectal cancer; current status and perspectives. Biochimie. 2019;157:64–71. doi:10.1016/j.biochi.2018.11.003
35. Li Q, Lai Q, He C, et al. RUNX1 promotes tumour metastasis by activating the Wnt/β-catenin signalling pathway and EMT in colorectal cancer. J Exp Clin Cancer Res. 2019;38(1):334. doi:10.1186/s13046-019-1330-9
36. Zhou Y, Xu J, Luo H, Meng X, Chen M, Zhu D. Wnt signaling pathway in cancer immunotherapy. Cancer Lett. 2022;525:84–96. doi:10.1016/j.canlet.2021.10.034
37. Brabletz S, Schuhwerk H, Brabletz T, Stemmler MP. Dynamic EMT: a multi-tool for tumor progression. EMBO J. 2021;40(18):e108647. doi:10.15252/embj.2021108647
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.