Back to Journals » Journal of Asthma and Allergy » Volume 19
Allergen Sensitization Profiles and Co-Sensitization Networks in Children with Allergic Rhinitis and Asthma: A Propensity Score-Matched Retrospective Study
Authors Zou P ![]() , Fan Y, Xie C, Xia W, Liu Y, Tao P, Huang Y, Wang L, Ai T
, Fan Y, Xie C, Xia W, Liu Y, Tao P, Huang Y, Wang L, Ai T
Received 29 April 2026
Accepted for publication 8 June 2026
Published 17 June 2026 Volume 2026:19 620785
DOI https://doi.org/10.2147/JAA.S620785
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Luis Garcia-Marcos
Pinli Zou, Yinghong Fan, Cheng Xie, Wanmin Xia, Yanru Liu, Pei Tao, Yijie Huang, Li Wang,* Tao Ai*
Pediatric Respiratory Medicine Department, Chengdu Women’s and Children’s Central Hospital, School of Medicine, University of Electronic Science and Technology of China, Chengdu, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Li Wang, Pediatric Respiratory Medicine Department, Chengdu Women’s and Children’s Central Hospital, School of Medicine, University of Electronic Science and Technology of China, 1617 Riyue Avenue, Qingyang District, Chengdu, 610091, People’s Republic of China, Email [email protected] Tao Ai, Pediatric Respiratory Medicine Department, Chengdu Women’s and Children’s Central Hospital, School of Medicine, University of Electronic Science and Technology of China, 1617 Riyue Avenue, Qingyang District, Chengdu, 610091, People’s Republic of China, Email [email protected]
Background: The sensitization mechanisms driving allergic airway disease progression from allergic rhinitis (AR) to asthma (AS) in children remain incompletely understood. This study investigated age-related patterns and topological network heterogeneity of allergen sensitization between these airway phenotypes.
Methods: In this retrospective cross-sectional study, 2193 children with AR or AS were enrolled. Propensity score matching (PSM) was utilized to control for age, yielding a 1:1 matched cohort of 1206 patients. The topological architecture of allergen co-sensitization was reconstructed using an Ising probability graphical model. Independent risk factors for lower airway involvement were identified via multivariable logistic regression.
Results: House dust mites mix (41%), cat dander (17%), and tree pollen mix (9.5%) dominated inhalant sensitizations. Egg white (22%), cow’s milk (20%), and lamb (19%) were the most prevalent food sensitizations. While inhalant sensitization exhibited homogenous age-dependent increases in both groups, foodborne immune tolerance showed profound heterogeneity. By adolescence (≥ 12 years), the risk of sensitization to milk and egg white was 5.64-fold (OR = 5.64, 95% CI: 1.47– 21.69, p = 0.010) and 3.40-fold (OR = 3.40, 95% CI: 1.07– 10.07, p = 0.029) higher than in the AR group. Ising network analysis revealed a highly compartmentalized modular structure in AR, whereas the AS network exhibited significant complexity and deep cross-system connectivity, with peanut acting as a topological hub bridging core indoor inhalants. Multivariable regression demonstrated cockroach (aOR: 2.70; 95% CI: 1.11– 7.57; p=0.039) and peanut (aOR: 1.86; 95% CI: 1.02– 3.47; p=0.046) sensitizations were independent risk factors for lower airway involvement.
Conclusion: Children with AS demonstrate a more complex cross-sensitization network than those with AR, rather than simply a higher burden of inhalant allergies. The delayed resolution of food sensitizations serves as a critical differentiator for the AS phenotype, highlighting the necessity of monitoring food allergies in pediatric airway disease management.
Keywords: asthma, allergic rhinitis, sensitization profile, atopic march, propensity score matching, Ising model
Introduction
Childhood airway allergic diseases, especially allergic rhinitis(AR) and bronchial asthma(AS), represent one of the most severe challenges faced by contemporary pediatrics, allergenics, and public health fields1。In the past decades, the incidence rate of AR and AS has shown an exponential upward trend due to the profound impact of global urbanization, changes in the ecological environment and changes in modern lifestyles2。Clinical and epidemiological evidence indicates that these two conditions are not isolated pathological events, but rather manifestations of highly intertwined systemic immune dysregulation3。The development of allergic diseases typically exhibits a significant age-dependent phenotypic variations—originating from impaired skin and gastrointestinal barriers in early life and progressively spreading to the upper and lower respiratory tract mucosa. This progression is academically referred to as the atopic march4。This variation further suggests that allergic rhinitis and asthma are not independent diseases, but rather heterogeneous expressions of the same allergic airway inflammation at different anatomical sites and developmental stages. However, this also poses a lingering clinical challenge: why do symptoms remain confined to the upper respiratory tract in some children, while others eventually progress to asthma?
Inhalant allergens, particularly house dust mites (HDM), are the most prevalent allergens for AS and AR, serving as a core driver of symptom exacerbation. Furthermore, the expansion of the sensitization spectrum—specifically polysensitization—is identified as a key factor in disease worsening or suboptimal control.5 However, allergen profiles vary significantly across regions, clinical phenotypes, and pediatric age groups. In northern and northwestern China, where the climate is relatively arid, outdoor pollens such as mugwort, dandelion, ragweed, and elm predominate.6 Conversely, in the warm and humid regions of southern, eastern, and southwestern China, indoor allergens like Dermatophagoides pteronyssinus, Dermatophagoides farinae, and cockroaches are dominant.7 The northeast and central regions exhibit mixed sensitization patterns, encompassing both mites and pollens from trees like plane, birch, and mugwort.6 Notably, research indicates that children with asthma have significantly higher sensitization rates to indoor allergens compared to those with AR alone. In contrast, AR patients show a higher prevalence of sensitivity to summer-autumn pollens (eg., mugwort). Furthermore, children with asthma exhibit a much higher proportion of strong positive reactions to single allergens and a significantly greater frequency of polysensitization involving four or more allergens than their AR counterparts.5,8
Moreover, foodborne allergen sensitization and the development of oral tolerance play crucial roles in the progression of pediatric allergic airway diseases. Oral tolerance is defined as an active state of immunological unresponsiveness to ingested food antigens, mediated primarily by gut-associated lymphoid tissue.9 During infancy, the early dietary introduction of common allergenic foods (eg., peanut, egg, and cow’s milk) facilitates the establishment of oral tolerance and significantly reduces the risk of food allergies. Importantly, this early intervention may disrupt the atopic march, thereby mitigating the subsequent progression to asthma and allergic rhinitis.10 Although previous research has well documented the impact of food allergy on the atopic march, which was particularly focusing on the progression from early-childhood atopic dermatitis to school-age atopic disorders,11 the specific role of food sensitization in the direct trajectory from allergic rhinitis to asthma remains incompletely understood.
Notably, as children represent a population with a continuously maturing immune system, age serves as a critical confounding variable, leading to substantial variations in allergen profiles across different developmental stages. Previous studies have consistently shown that the sensitization rate for inhalant allergens tends to increase with age, whereas that for food allergens progressively declines.12–14 Consequently, comparative analyses of allergen profiles in pediatric AR and AS must account for potential bias introduced by age shifts. Regrettably, most existing comparisons between AR and AS only report crude intergroup differences without rigorous age-matching, leaving it unclear whether the influence of age-related bias has been effectively eliminated.
To resolve the fog of age-related confounding and reveal the authentic differences in immune sensitization between AR and AS, this study performed an in-depth analysis of a large real-world cohort comprising over 2000 allergic children. We employed a rigorous 1:1 Propensity Score Matching (PSM) to equalize the age distribution across both groups. Within this purified matched cohort, we subsequently introduced the Ising Model,15 specifically designed for binary data analysis. By constructing allergen topological networks for the AR and AS groups, this study aims to address a fundamental question: once the natural influence of chronological growth is decoupled, do upper and lower respiratory allergies in children still exhibit distinct systemic co-sensitization architectures? Furthermore, what role do the cross-boundary links between food and inhalant allergens play within these frameworks?
Methods
Study Design and Population
Pediatric patients (aged 1–17 years) who were treated at Chengdu Women’s and Children’s Central Hospital between January 2025 and March 2026 and underwent specific allergen screening were retrospectively included in this study. The cohort was established by systematically querying the hospital’s electronic medical record system. De-identified clinical data, including demographic characteristics, medical history, and specific allergen screening results, were retrieved and exported from the hospital’s database. The criteria for phenotype classification were strictly adhered to according to international standards. AR was diagnosed based on the Chinese Guideline for Diagnosis and Treatment of Allergic Rhinitis (2022, revision),16 wherein patients must exhibit typical nasal symptoms such as rhinorrhea, sneezing, nasal congestion, or nasal itching, and where clinical examination or nasal endoscopy indicates allergic mucosal inflammation. AS was diagnosed referencing the Global Initiative for Asthma (GINA) report17, with recurrent episodes of wheezing, dyspnea, or cough accompanied by reversible airflow limitation being the core diagnostic criterion.
Furthermore, cases presenting with severe congenital airway anomalies, autoimmune diseases, hereditary immunodeficiency, or severely incomplete clinical data were systematically excluded. The final cohort was thus comprised of 2193 confirmed pediatric patients, with 1430 belonging to the AR group and 763 to the AS group.
Allergen Sensitization Profiling
Peripheral blood samples were processed by centrifugation, and serum specific immunoglobulin E (sIgE) levels were quantitatively measured using the EUROIMMUN automated immunoblotting system via the Western blot method.18 The screening panel comprised 18 core allergens with the highest epidemiological representativeness in the region. These included nine inhalant allergens: Artemisia (mugwort), HDM mix (Dermatophagoides pteronyssinus and Dermatophagoides farinae), dog dander, Humulus scandens, cat dander, mold mix (Penicillium notatum, Alternaria alternata, Aspergillus fumigatus, and Cladosporium herbarum), ragweed, tree pollen mix (willow, poplar, and elm), and cockroaches; and nine food allergens: sea fish mix (cod, lobster, and scallop), peanuts, soybeans, egg white, milk, beef, shrimp, crab, and lamb. Based on sIgE titers, results were categorized into six grades: Grade 0 (<0.35 kU/L), Grade 1 (0.35−0.7 kU/L), Grade 2 (0.7−3.5 kU/L), Grade 3 (3.5−17.5 kU/L), Grade 4 (17.5−50 kU/L), Grade 5 (50−100 kU/L), and Grade 6 (>100 kU/L). Grades 1–6 were defined as allergen-positive, grades ≥3 were considered strongly positive, and positivity to ≥2 allergens was defined as polysensitization. Importantly, given the retrospective nature of this cohort, positivity to foodborne allergens specifically indicates food sensitization strictly defined by serological testing, rather than clinically confirmed food allergies, as gold-standard oral food challenges (OFC) were not universally recorded in the electronic medical records.
Statistical Analysis
All data cleaning, matching, and visualization were performed using R software (version 4.5.0), primarily utilizing the MatchIt, bootnet (with IsingFit algorithm), tidyverse, gtsummary, and qgraph packages.
Propensity score matching (PSM) was executed to balance age distribution between groups, using a 1:1 nearest-neighbor algorithm without replacement and a caliper width of 0.05. The balance was validated using independent samples t-tests. In the matched cohort, categorical variables were presented as frequencies (percentages), while continuous variables were expressed as mean ± SD. Differences in allergen sensitization rates between groups were assessed using univariate logistic regression, with results reported as odds ratios (ORs) and 95% confidence intervals (CIs). Statistical significance was defined as a two-sided P<0.05. Given the multiple comparisons performed across various allergens and age strata, raw P-values in these comparative analyses were adjusted using the false discovery rate method via the Benjamini-Hochberg procedure to control for false positives.
Topological networks were constructed via the IsingFit algorithm based on the eLasso method.19 To facilitate rigorous visual comparison between the AR and AS groups, node coordinates were constrained to a fixed average layout.
Ethical Approval and Funding Support
This study protocol received approval from the Ethics Committee of Chengdu Women’s and Children’s Central Hospital (Approval No. 2025–112-2). Due to the retrospective observational nature of this research, informed consent was waived. Patient data confidentiality was strictly maintained, and the study was conducted in accordance with the Declaration of Helsinki.
This work was supported by the Medical Research Project of Chengdu Health Commission (Grant No. [2025170]). The funding body played no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Results
Baseline Demographic Characteristics and Sensitization Burden
This study included a total of 2193 pediatric patients with complete quantitative sIgE data, consisting of 763 cases in the AS group and 1430 cases in the AR group. As summarized in Table 1, no significant difference was observed in gender distribution between the two groups (proportion of boys: 63% vs. 63%, p =0.837). The mean age of the AS group was significantly lower than that of the AR group (4.5 ± 3.1 years vs. 7.1 ± 3.2 years, p <0.001). Notably, despite the differing clinical phenotypes, there were no statistically significant differences in the overall atopy rate (70% vs. 73%, p =0.139) or the polysensitization burden (median number of positive allergens: 1 [IQR: 0–3] for both groups, p =0.806).
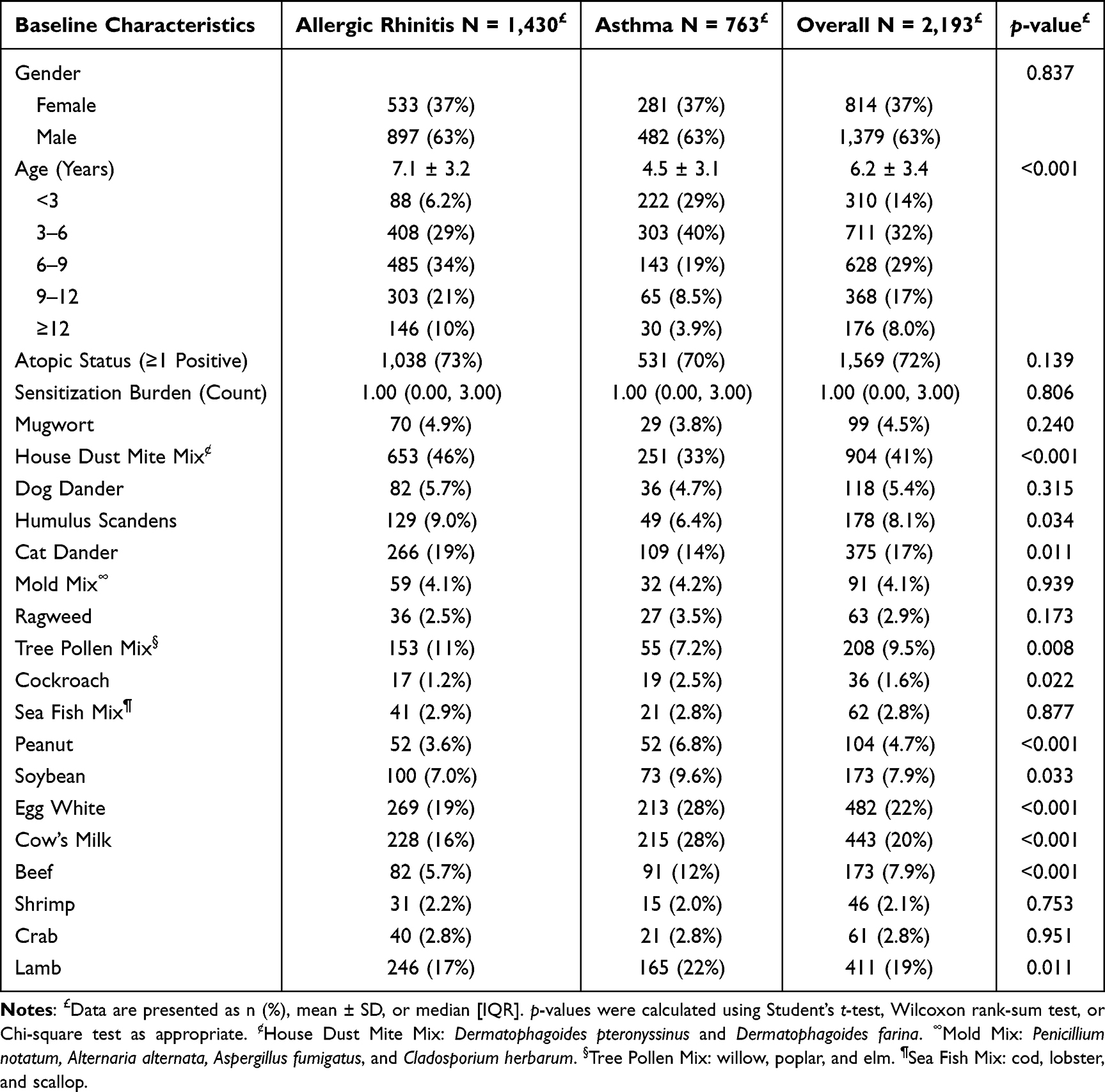 |
Table 1 Baseline Characteristics and Sensitization Profiles of the Primary Cohort |
Among the 2193 included pediatric patients, the regional allergen profile in Southwestern China was characterized by a profound dominance of indoor inhalants. Specifically, within the inhalant spectrum, house dust mites mix ranked highest in prevalence (41%, n =904), followed by cat dander (17%, n =375) and tree pollen mix (9.5%, n=208). Regarding food-borne exposures, egg white (22%, n = 482), cow’s milk (20%, n = 443), and lamb (19%, n = 411) exhibited the highest baseline positivity rates.
Divergence of Sensitization Profiles Across Age Strata
In the full unmatched cohort (N = 2193), the allergen sensitization spectra exhibited highly characteristic and oppositely directed age-related distribution patterns. Overall, the age-related trends of environmental inhalant and food allergens were not parallel, but demonstrated a crossover (Figure 1).
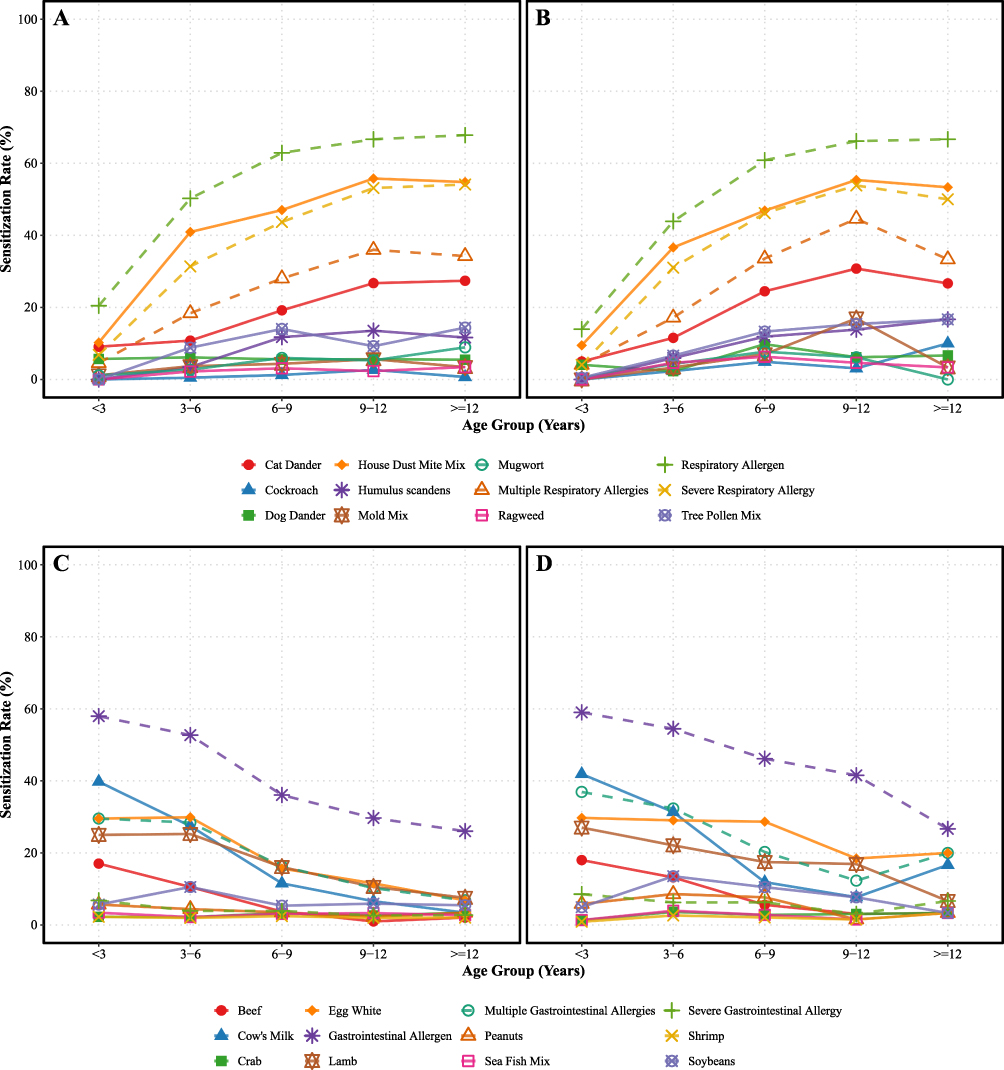 |
Figure 1 Age-related trends of specific allergen sensitization rates across pediatric age groups. The cohort is stratified into five predefined age intervals (<3, 3–6, 6–9, 9–12, and ≥12 years). The four quadrants delineate distinct profiles based on clinical phenotype and allergen category: (A) Inhalant allergens in Allergic Rhinitis (AR); (B) Inhalant allergens in Allergic Asthma (AS); (C) Foodborne allergens in AR; and (D) Foodborne allergens in AS. Component definitions for mixture allergens are specified as follows: House Dust Mite Mix comprises Dermatophagoides pteronyssinus and Dermatophagoides farinae; Mold Mix comprises Penicillium notatum, Alternaria alternata, Aspergillus fumigatus, and Tree Pollen Mix comprises willow, poplar, and elm; and Sea Fish Mix comprises cod, lobster, and scallop. |
Indoor inhalant allergens, represented by HDM mix, displayed highly overlapping, steeply ascending sensitization curves in both groups. In the early-life window (< 3 years), sensitization rates were comparably low between the AR and AS groups (10.2% vs. 9.5%; OR = 0.92, 95% CI: 0.40–2.09, p = 0.837). Positive rates increased exponentially with age, peaking at 9–12 years (55.8% vs. 55.4%; OR = 0.98; 95% CI: 0.57-1.69, p = 0.954). Age-stratified logistic regression analyses confirmed that the risk of HDM mix sensitization remained consistently equivalent across all five age strata from infancy to adolescence (Table S1).
In stark contrast to the age-related increase of inhalant sensitization, food allergens exhibited a typical early-peak followed by a lower prevalence in older strata, a pattern consistent with the acquisition of oral tolerance (Figure 1). However, the two groups exhibited marked longitudinal heterogeneity in the establishment of tolerance. During infancy (< 3 years), cow’s milk and egg white constituted the predominant sensitizing drivers, with comparable baseline rates between groups (milk: OR = 1.09, p = 0.733; egg: OR = 1.01, p = 0.974). In the AR group, classic oral tolerance was established, with sensitization rates declining sharply to 3.4% and 6.8% for cow’s milk and egg white, respectively, by adolescence (≥ 12 years). In the AS group, however, a clear delay in tolerance acquisition was evident: at ≥ 12 years, the risk of sensitization to milk and egg white was 5.64-fold (OR = 5.64, 95% CI: 1.47–21.7, p = 0.010) and 3.40-fold (OR = 3.40, 95% CI: 1.07–10.1, p = 0.029) higher than in the AR group.
Furthermore, during the school-age period (6–9 years), the high-risk driving effect of food sensitization on lower-airway involvement was particularly pronounced. The AS group showed significantly higher peanut sensitization (OR = 2.29, 95% CI: 1.02–4.96, p = 0.038) and egg white (OR = 2.16, 95% CI: 1.39–3.34, p < 0.001) than the AR group.
To dissect the age-related variation of sensitization, we analyzed the positivity rates across five age strata (Supplementary Figure S1). The trends of inhalant allergens, like HDM mix and animal dander, exhibited a homogenous, age-dependent continuous escalation in both the AR and AS cohorts. Conversely, a profound divergence was observed in foodborne allergens. While the AR group demonstrated a typical progressive acquisition of oral tolerance, characterized by a sharp decline in cow’s milk and egg white sensitization with age, the AS cohort exhibited a prolonged persistence of food sensitization into adolescence, highlighting a significant developmental tolerance gap.
Propensity Score Matching and Topological Heterogeneity of Allergen Co-Sensitization Networks
We further investigated the difference in absolute intensity of humoral responses between the AR and AS groups. When restricting the analysis to sensitized individuals (sIgE > 0.35 kU/L), the quantitative titer profiles for the majority of inhalant allergens, including HDM mix and cat dander, showed no significant differences between the AR and AS phenotypes (Supplementary Figure S2). Notably, cow’s milk maintained a significantly higher quantitative titer in the AS group (p < 0.01).
To control for chronological age, we applied 1:1 propensity score matching (PSM) to the initial cohort. This yielded a subcohort of 1206 children (603 in the AS group and 603 in the AR group). Post-matching, the age distribution and other demographic covariates were balanced between the two groups (p = 0.990; Table 2).
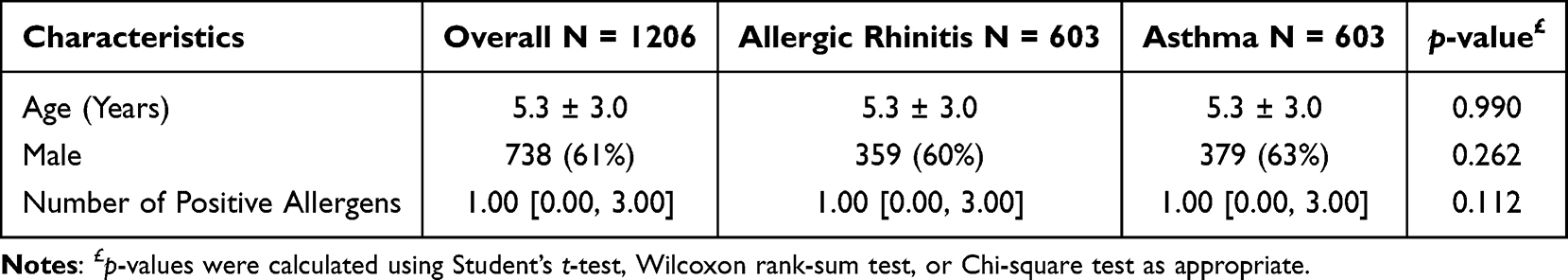 |
Table 2 Baseline Characteristics of the Propensity Score-Matched Cohort |
Based on this matched cohort, we constructed co-sensitization topological networks using the Ising model (Figure 2), where nodes represent specific allergens and edges denote regularized conditional dependence. In the AR group, the network displayed modular segregation: food and inhalant allergens formed intra-cluster connections but remained largely isolated from each other. Limited bridging edges existed between egg white and cat epithelium, and between peanut and dog epithelium. Plant-derived inhalants (ragweed, mugwort, Humulus scandens, and tree pollen mix) constituted a distinct module with strong inter-connections.
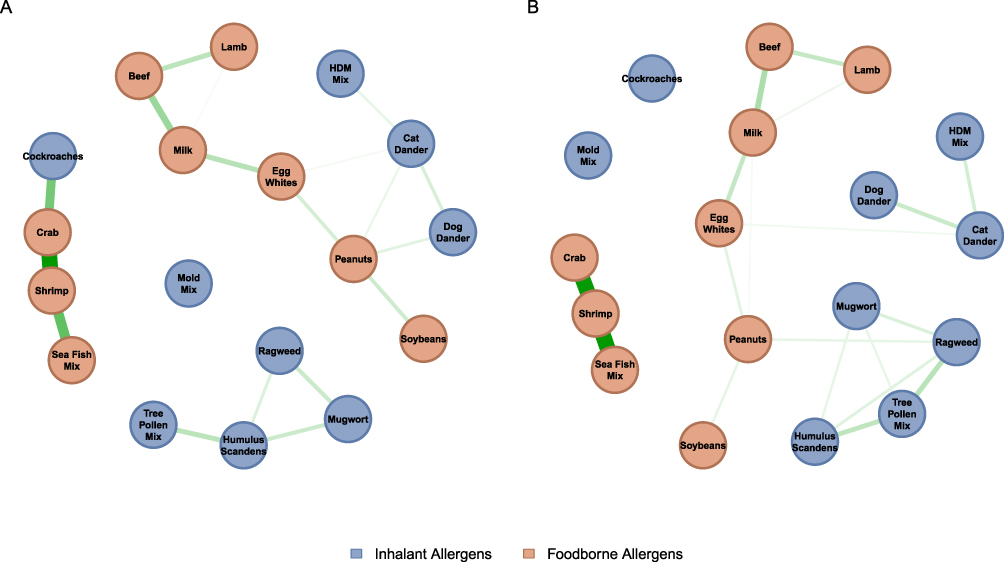 |
Figure 2 Topological networks of allergen co-sensitization in the Asthma and Allergic Rhinitis cohorts. (A) Topological network in allergic rhinitis group by Ising model. (B) Topological network in asthma group by Ising model. Nodes represent specific allergens, and the thickness of the edges indicates the strength of the partial correlations (co-sensitization). Notes: House Dust Mite Mix includes Dermatophagoides pteronyssinus and Dermatophagoides farinae; Mold Mix includes Penicillium notatum, Alternaria alternata, Aspergillus fumigatus, and Cladosporium herbarum; Tree Pollen Mix includes willow, poplar, and elm; Sea Fish Mix includes cod, lobster, and scallop. |
In contrast, the AS cohort exhibited an increase in both overall network density and topological complexity. The boundary between food and inhalant allergens was blurred, with peanut emerging as a central hub bridging indoor and outdoor (plant-derived) inhalant allergens.
In both groups, mold mix formed an independent sensitization cluster, and the seafood cluster (shrimp, crab, sea fish mix) exhibited tight internal co-sensitization. The seafood cluster in the AR group maintained a strong connection with cockroach, a linkage absent in the AS network. Despite the high prevalence of HDM mix sensitization in both cohorts, this allergen occupied a peripheral position in both networks, showing only a single strong edge to cat dander.
Independent Risk Factors for AR and AS Phenotypes
After balancing baseline confounders via PSM (n=1206), a logistic regression model was employed to quantify the independent sensitization risk factors associated with the AS phenotype compared to isolated AR. Table 3 revealed that high-frequency indoor environmental antigens, including HDM mix (OR = 0.92, 95% CI: 0.73–1.16, p = 0.481), cat dander (OR = 1.24, 95% CI: 0.91–1.70, p = 0.177), and dog dander (OR = 0.86, 95% CI: 0.52–1.39, p = 0.536), showed no statistically significant differences in sensitization rates between the two groups.
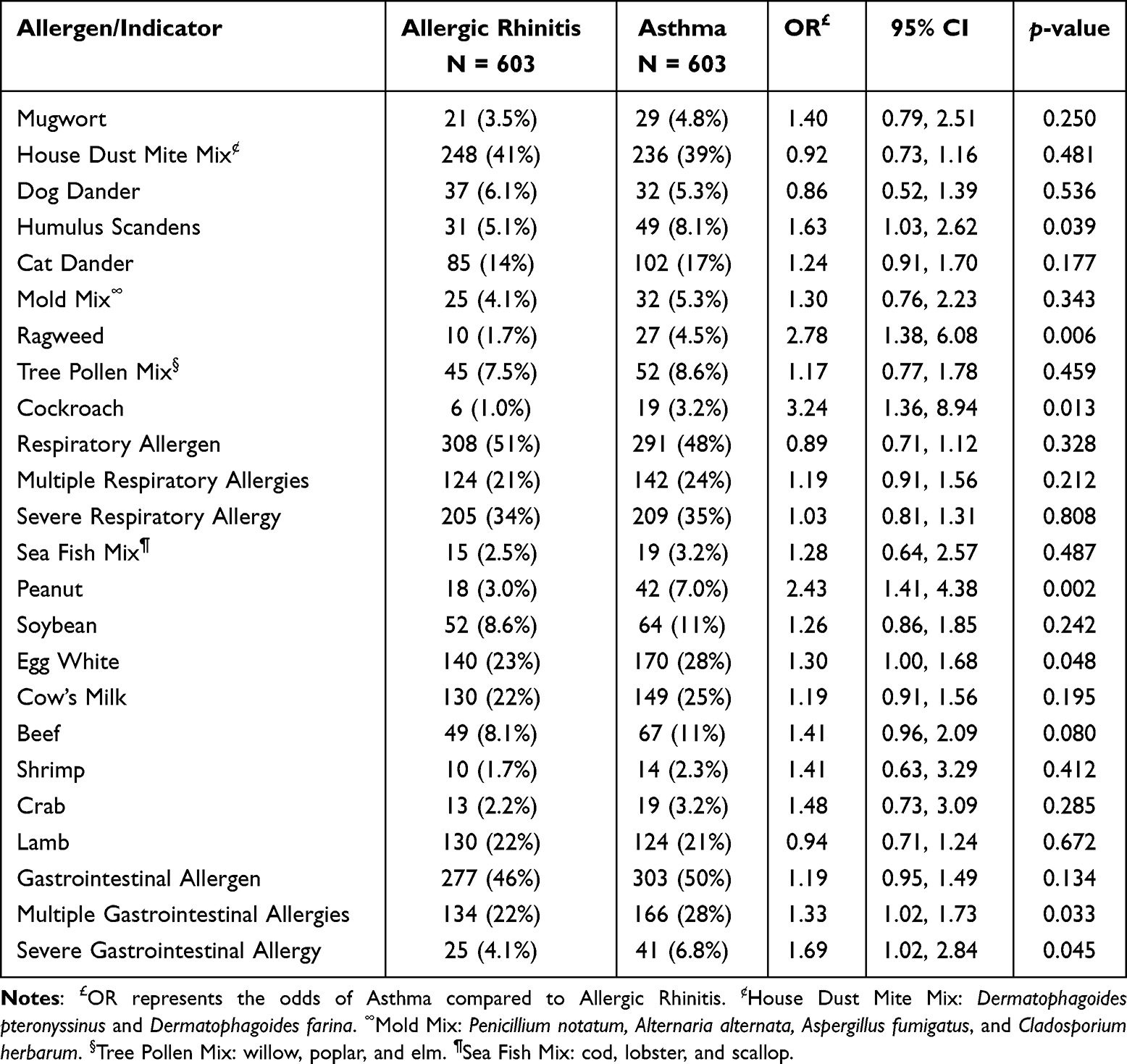 |
Table 3 Univariate Logistic Regression Analysis of Allergen Sensitizations Associated with Asthma versus Allergic Rhinitis in the Propensity Score-Matched Cohort |
In contrast, specific atypical indoor and outdoor allergens emerged as risk factors for asthma. Children sensitized to cockroaches had a higher relative risk of developing AS compared to those with AR alone (OR = 3.24, 95% CI: 1.36–8.94, p = 0.013). Similarly, autumn weed pollens, including ragweed (OR = 2.78, 95% CI: 1.38–6.08, p = 0.006) and Humulus scandens (OR = 1.63, 95% CI: 1.03–2.62, p = 0.039), significantly increased the risk of AS. Among food allergens, peanut sensitization was identified as a significant risk factor for asthma (OR = 2.43, 95% CI: 1.41–4.38, p = 0.002).
Regarding the dimension of sensitization burden, children with polysensitization to food allergens (multiple food sensitization: OR = 1.33, 95% CI: 1.02–1.73, p = 0.033) or those exhibiting strong positive reactions (severe food sensitization: OR = 1.69, 95% CI: 1.02–2.84, p = 0.045) showed a significantly elevated risk of lower airway involvement.
To identify the independent predictors of the asthma phenotype, we constructed a multivariable logistic regression model incorporating all allergens that reached significance in the univariate analysis. After adjustment for potential co-sensitization, sensitization to cockroaches (adjusted OR = 2.70, 95% CI: 1.11-7.57, p = 0.039) and peanuts (adjusted OR = 1.86, 95% CI: 1.02-3.47, p = 0.046) emerged as the most significant independent risk factors for lower airway involvement within the matched cohort (Figure 3). Despite the high prevalence of HDM mix sensitization in both groups, it did not show a significant predictor in the univariate logistic regression model.
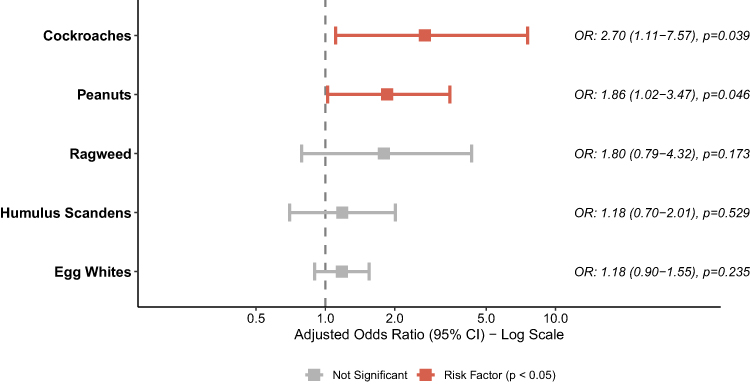 |
Figure 3 Forest plot of multivariable logistic regression analysis assessing independent allergen sensitizations distinguishing Asthma from Allergic Rhinitis. The forest plot presents the adjusted Odds Ratios (aOR) and 95% Confidence Intervals (CI) on a log scale. Red squares (aOR > 1) denote significant risk factors for AS, while nonsignificant associations are in gray. Statistical significance was defined as P < 0.05. |
Discussion
The classical theory of the atopic march typically depicts a linear trajectory of allergic development, beginning with food allergies in early life and gradually transitioning to inhalant allergies during the preschool and school-age years.20 However, our age-stratified data and propensity-matched analysis suggest that when assessing the risk of lower airway disease, primary focus should be directed toward the establishment of food immune tolerance. Our observations reveal that the critical clue distinguishing asthma from rhinitis is not merely whether food sensitization occurred in early life, as both groups exhibited similar sensitization rates (approximately 30–40%) to cow’s milk and egg white before three years old, but rather whether this sensitization state was effectively resolved in subsequent life stages.
In the AR group, the positive rates of these foundational food allergens exhibited a clear natural resolution prior to adolescence, consistent with the typical establishment of oral mucosal tolerance. Conversely, this tolerance process was significantly delayed in the AS cohort; even after entering adolescence (≥12 years), the AS group exhibited significantly higher odds of sensitization to specific food allergens, such as cow’s milk and egg white, compared to those with AR alone (OR = 5.64 and 3.40, respectively; both p < 0.05). Furthermore, in the age-matched multivariate analysis, peanut sensitization remained robustly associated with the asthma phenotype as an independent risk factor.
These findings strongly corroborate recent conclusions from large-scale prospective birth cohorts, providing compelling converging evidence. For instance, the six-year follow-up report from the authoritative Australian HealthNuts study in 2023 explicitly stated that persistent food sensitization during infancy—whether involving unresolved reactions to egg white or peanut—is directly linked to significantly reduced lung function and a markedly elevated risk of confirmed asthma at age six.21
Beyond the temporal persistence of early-life sensitization, our evaluation of sensitization profiles provides further systemic insights. Our findings suggest that the clinical differentiation between upper and lower airway phenotypes (AR vs. AS) is not primarily dictated by the mere magnitude of sIgE production, given the broadly non-significant difference in inhalant titer intensities between the two groups. Instead, network topology analysis reveals that while AR and AS belong to the same respiratory continuum,22 their underlying systemic sensitization architectures exhibit notable heterogeneity. Furthermore, despite its high prevalence, the consistently peripheral positioning of HDM mix in both networks indicates that HDM mix sensitization possesses a relatively independent profile within the overall pediatric co-sensitization architecture.
Building upon this topological framework, our logistic regression results suggest that while core inhalants such as HDM mix, cat and dog dander form the foundation of airway involvement in atopic children, they do not serve as specific differentiating drivers determining whether the disease progresses to asthma. Instead, atypical allergens characterized by high allergenicity and potent protease activity (eg., cockroaches and weed pollens) demonstrated a stronger predilection for the lower airway. More strikingly, food allergens played a non-negligible systemic role in driving the asthma phenotype. Echoing the central position of peanut in our topological network, peanut sensitization, alongside the increased depth and breadth of overall food sensitization, provided more distinct risk factors for lower airway involvement than ubiquitous indoor allergens. Our cross-sectional findings regarding the complex network architecture in age strata strongly align with the long-term sensitization structures reported in a recent longitudinal study by Cucco et al23 Their dimensionality reduction mapping, generated via manifold learning and graph embedding techniques, demonstrated that throughout childhood, component-resolved IgE sensitization networks in asthmatic patients consistently display a higher degree of complexity and connectivity compared to non-asthmatic children.
These topological findings provide compelling clinical epidemiological evidence supporting the gut-lung axis theory. Embryologically, both the gastrointestinal and respiratory tracts originate from the primitive foregut and share a highly homologous immune network of mucosa-associated lymphoid tissue.24 Frontier immunological research indicates that the disruption of the intestinal mucosal barrier not only triggers local inflammation but also drives activated Th2 cells, innate lymphoid cells, and inflammatory mediators to migrate to the respiratory mucosa via lymphatic and systemic circulation, thereby exacerbating type 2(T2) inflammation in the lower airways.25 Consequently, for pediatric patients exhibiting unresolved food allergies or persistent sensitization to highly allergenic antigens such as peanuts, clinicians should not focus solely on gastrointestinal or dermatological symptoms. Instead, they must maintain high clinical vigilance regarding the potential risk of atopic march to the lower airways, initiating systematic evaluations of lung function and lower airway inflammation at the earliest opportunity.
Concurrently, through rigorously matched multivariable regression analysis, this study precisely identified cockroach and peanut sensitizations as independent risk factors triggering lower airway allergic inflammation, which is consistent with previous reports.26 The distinct pathogenic roles of these allergens stem from different but synergistic mechanistic pathways. On one hand, cockroach frass is highly enriched with active endogenous serine proteases capable of directly cleaving tight junction proteins between airway epithelial cells, leading to physical barrier leakage.27 Such impairment of barrier function allows inhaled antigens to directly interface with submucosal immune cells, thereby driving intense eosinophilic airway inflammation.28 On the other hand, peanut sensitization similarly exhibited a higher prevalence in the AS cohort and maintained an independent association in the multivariable analysis, suggesting that its contribution to lower airway involvement is more likely characterized by systemic immune dysregulation.21,29 Distinct from the direct disruption of the airway epithelial barrier by inhalant allergens such as cockroaches, peanut sensitization more likely reflects a failure of oral tolerance mechanisms, resulting in a systemic inflammatory state dominated by T2 immune responses. Consequently, it partially amplifies the inflammatory effects induced by environmental allergens, ultimately culminating in a synergistic pathological cascade that promotes lower airway inflammation and airway remodeling.24
However, this study has several limitations. First, although PSM was utilized to simulate cross-age sensitization trends, the design remains inherently cross-sectional and cannot entirely substitute for a true prospective longitudinal birth cohort in establishing causal relationships. Second, foodborne allergen reactivity in our cohort was defined based on sIgE sensitization rather than clinically confirmed food allergy via oral food challenges. While sIgE serves as a robust and standard epidemiological marker for assessing atopic status in large-scale retrospective studies, asymptomatic sensitization does not always perfectly translate to clinical allergic symptoms. Future prospective studies incorporating OFC are warranted to validate the clinical impact of these network topologies. In addition, the study lacks direct laboratory metrics assessing epithelial barrier function; thus, the discussion regarding barrier disruption is largely extrapolated from established theoretical frameworks. Furthermore, as a single-center study, the observed sensitization profiles may be specifically influenced by the unique vegetation and environmental exposures of the Chengdu region. Future research should incorporate multicenter longitudinal follow-ups, integrating respiratory microbiome analyses, metabolomics, and epithelial function assessments to further elucidate the underlying molecular mechanisms driving the transition of sensitization networks from a modular to a systemic architecture.
Conclusion
In conclusion, utilizing a large-scale pediatric allergic airway cohort, this study explored the age-related differences and network characteristics of sensitization profiles in AR and AS in Southwestern China, effectively eliminating age-related confounding through propensity score matching and an Ising probabilistic graphical model. Our principal findings are as follows: First, high-frequency indoor inhalants (eg., HDM mix, cat and dog dander) exhibited highly consistent exposure and sensitization trends across both AR and AS phenotypes; in the present matched cohort, they did not emerge as independent risk factors differentiating asthma from isolated allergic rhinitis. Second, network topology analysis revealed that, in contrast to the highly compartmentalized, modular sensitization structure observed in the AR group, the underlying sensitization architecture in the AS group was significantly more complex, characterized by deep cross-system connectivity between food and inhalant allergens. Third, persistent food sensitization—specifically, delayed oral tolerance to cow’s milk and egg white—along with sensitization to specific high-risk antigens (such as peanut and cockroach), was significantly associated with an elevated risk of lower airway involvement.
Disclosure
The authors declare that they have no conflicts of interest related to the publication of this manuscript.
References
1. Pham TT, Akinbolagbe Y, Nguyen TH, et al. Urbanization and childhood asthma. Pediatr Allergy Immunol. 2025;36(7):e70137. doi:10.1111/pai.70137
2. Kanannejad Z, Taylor WR, Mohkam M, Ghatee MA. Urban lifestyle and climate-driven environmental exposures: immunological consequences for pediatric respiratory allergies. Immun Inflamm Dis. 2025;13(8):e70248. doi:10.1002/iid3.70248
3. Giombi F, Pace GM, Pirola F, et al. Airways type-2 related disorders: multiorgan, systemic or syndemic disease? Int J Mol Sci. 2024;25(2):730. doi:10.3390/ijms25020730
4. de Las Vecillas L, Quirce S. The multiple trajectories of the allergic march. J Investig Allergol Clin Immunol. 2024;34(2):75–12. doi:10.18176/jiaci.0983
5. Xu Q, Zhao H, Dong Y, et al. Polysensitization patterns and age-dependent allergen profiles in children from suzhou, china: a component-resolved cross-sectional study. J Asthma Allergy. 2026;19:556612. doi:10.2147/JAA.S556612
6. Huang Z, Li A, Zhu H, et al. Multicenter study of seasonal and regional airborne allergens in Chinese preschoolers with allergic rhinitis. Sci Rep. 2024;14(1):4754. doi:10.1038/s41598-024-54574-z
7. Chen Y, Zhou X, Wang Y, et al. Characterising the allergen landscape in paediatric allergic rhinitis and/or asthma. BMJ Open Respir Res. 2026;13.
8. Guan K, Zhu W, Sha L, et al. Prevalence of sensitization to aeroallergens in greater Beijing region children with respiratory allergy. Front Pediatr. 2022;10:848357. doi:10.3389/fped.2022.848357
9. Foong RX, Santos AF. Oral tolerance induction-opportunities and mechanisms. Foods. 2022;11.
10. Trogen B, Jacobs S, Nowak-Wegrzyn A. Early introduction of allergenic foods and the prevention of food allergy. Nutrients. 2022;14.
11. Zeiger RS, Schatz M, Zhou B, et al. Impact of food allergy on the atopic march progression from atopic dermatitis in early childhood to other atopic disorders at school age. J Allergy Clin Immunol Pract. 2025;13(8):1991–2003.e9. doi:10.1016/j.jaip.2025.05.001
12. Hu Z, Xue J, Pan M, et al. Prevalence of allergen sensitization among children with allergic rhinitis in Changzhou, China: a retrospective observational study. BMC Pediatric. 2023;23(1):466. doi:10.1186/s12887-023-04291-9
13. Zhao Z, Chen L, Huang C, et al. Allergen sensitization patterns in children with allergic rhinitis: insights from a four-year retrospective study in Shenzhen, China. BMC Pediatr. 2025;25(1):544. doi:10.1186/s12887-025-05885-1
14. Tsai Y-C, Huang Y-L, Hsieh P-H, et al. Differential contributions of inhalant and food allergen sensitization to atopic phenotype asthma prevalence among children across age groups. Pediatr Crit Care Med. 2025;9(3):84–90. doi:10.4103/prcm.prcm_10_25
15. Finnemann A, Borsboom D, Epskamp S, et al. The theoretical and statistical ising model: a practical guide in R. Psych. 2021;3(4):593–617. doi:10.3390/psych3040039
16. Subspecialty Group of Rhinology. E.B.o.C.J.o.O.H.a.N.S. and S.o.O.H.a.N.S. Subspecialty Group of Rhinology, Chinese Medical Association, Chinese guideline for diagnosis and treatment of allergic rhinitis (2022, revision). Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2022;57(2):106–129. doi:10.3760/cma.j.cn115330-20211228-00828
17. Asthma, G.I.f. Global Strategy for Asthma Management and Prevention (2024 Update). Global Initiative for Asthma; 2024.
18. Lazova S, Baleva M, Priftis S, et al. Atopic status in children with asthma and respiratory allergies—comparative analysis of total IgE, ImmunoCAP Phadiatop/fx5 and euroimmun pediatric immunoblot. Sinusitis. 2022;6(1):1–14. doi:10.3390/sinusitis6010001
19. Zhang Z. Propensity score method: a non-parametric technique to reduce model dependence. Ann Transl Med. 2017;5(1):7. doi:10.21037/atm.2016.08.57
20. Gabryszewski SJ, Hill DA. One march, many paths: insights into allergic march trajectories. Ann Allergy Asthma Immunol. 2021;127(3):293–300. doi:10.1016/j.anai.2021.04.036
21. Peters RL, Soriano VX, Lycett K, et al. Infant food allergy phenotypes and association with lung function deficits and asthma at age 6 years: a population-based, prospective cohort study in Australia. Lancet Child Adolesc Health. 2023;7(9):636–647. doi:10.1016/S2352-4642(23)00133-5
22. Brożek JL, Bousquet J, Agache I, et al. Allergic Rhinitis and its Impact on Asthma (ARIA) guidelines-2016 revision. J Allergy Clin Immunol. 2017;140(4):950–958. doi:10.1016/j.jaci.2017.03.050
23. Cucco A, Simpson A, Murray C, et al. Exploring the temporal interplay of allergic sensitization and asthma via graph and manifold analysis. J Allergy Clin Immunol. 2025;155(6):1836–1845. doi:10.1016/j.jaci.2025.02.012
24. Dora D, Szőcs E, Soós Á, et al. From bench to bedside: an interdisciplinary journey through the gut-lung axis with insights into lung cancer and immunotherapy. Front Immunol. 2024;15–2024.
25. Parrón-Ballesteros J, Gordo RG, López-Rodríguez JC, et al. Beyond allergic progression: from molecules to microbes as barrier modulators in the gut-lung axis functionality. Front Allergy. 2023;4:1093800. doi:10.3389/falgy.2023.1093800
26. Zoratti E, Wood R, Pomés A, et al. A pediatric randomized, controlled trial of German cockroach subcutaneous immunotherapy. J Allergy Clin Immunol. 2024;154(3):735–744.e10. doi:10.1016/j.jaci.2024.04.022
27. Ouyang X, Reihill JA, Douglas LEJ, et al. Airborne indoor allergen serine proteases and their contribution to sensitisation and activation of innate immunity in allergic airway disease. Eur Respir Rev. 2024;33(172):230126. doi:10.1183/16000617.0126-2023
28. Lu HF, Zhou YC, Yang LT, et al. Involvement and repair of epithelial barrier dysfunction in allergic diseases. Front Immunol. 2024;15:1348272. doi:10.3389/fimmu.2024.1348272
29. Hamaguchi S, Umezawa K, Toyokuni K, et al. Differential impact of allergen sensitization on asthma development in high-risk japanese infants with atopic dermatitis: the possible role of peanut sensitization. Jma J. 2026;9(1):217–224. doi:10.31662/jmaj.2025-0301
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Specific IgE Response and Omalizumab Responsiveness in Severe Allergic Asthma
Tajiri T, Suzuki M, Kutsuna T, Nishiyama H, Ito K, Takeda N, Fukumitsu K, Kanemitsu Y, Fukuda S, Umemura T, Ohkubo H, Maeno K, Ito Y, Oguri T, Takemura M, Yoshikawa K, Niimi A
Journal of Asthma and Allergy 2023, 16:149-157
Published Date: 22 January 2023
Elevated Saliva Pepsin Concentration as a Risk Factor for Asthma in Children with Allergic Rhinitis: A Preliminary Study
Sui H, Shen H, Zhang C, Wang M, Zhen Z, Zhang J
Journal of Asthma and Allergy 2024, 17:391-397
Published Date: 22 April 2024
Association of Mite Molecular Sensitization Profiles with Respiratory Allergies and Asthma Control in Children from East China
He J, Lin N, Jin T, Lin M, Huang Z, Li S, Liu J, Su L, Ye X, Wu L, Song Z, Xu H, Chen Z
Journal of Asthma and Allergy 2024, 17:965-975
Published Date: 7 October 2024
Knowledge Mapping of COVID-19 and Asthma/Allergic Rhinitis: A Visual and Bibliometric Analysis
Chen Y, Song C, Wang J, Cao Y, Lu Y, Han X
Journal of Asthma and Allergy 2025, 18:705-721
Published Date: 6 May 2025
Dupilumab is Effective in Young Children with Atopic Dermatitis Regardless of Type 2 Comorbidities
Boguniewicz M, Sher LD, Paller AS, Arkwright PD, Yoshihara S, Smith Begolka W, Chen Z, Shah P, Rodríguez Marco A
Journal of Asthma and Allergy 2026, 19:590176
Published Date: 19 May 2026