Back to Journals » The Application of Clinical Genetics » Volume 13
Alkaptonuria: Current Perspectives
Authors Zatkova A, Ranganath L, Kadasi L
Received 30 September 2019
Accepted for publication 4 December 2019
Published 23 January 2020 Volume 2020:13 Pages 37—47
DOI https://doi.org/10.2147/TACG.S186773
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Martin Maurer
Andrea Zatkova,1 Lakshminarayan Ranganath,2 Ludevit Kadasi1,3
1Department of Human Genetics, Biomedical Research Center, Slovak Academy of Sciences, Institute of Clinical and Translational Research, Bratislava, Slovakia; 2National Alkaptonuria Centre, Royal Liverpool University Hospital, Liverpool, UK; 3Faculty of Natural Sciences, Department of Molecular Biology, Comenius University, Bratislava, Slovakia
Correspondence: Ludevit Kadasi
Department of Molecular Biology, Faculty of Natural Sciences, Comenius University Address 6 Ilkovicova, Bratislava 84215, Slovakia
Tel +421 905 659 880
Email [email protected]
Abstract: The last 15 years have been the most fruitful in the history of research on the metabolic disorder alkaptonuria (AKU). AKU is caused by a deficiency of homogentisate dioxygenase (HGD), the enzyme involved in metabolism of tyrosine, and is characterized by the presence of dark ochronotic pigment in the connective tissue that is formed, due to high levels of circulating homogentisic acid. Almost 120 years ago, Sir Archibald Garrod used AKU to illustrate the concept of Mendelian inheritance in man. In January 2019, the phase III clinical study SONIA 2 was completed, which tested the effectiveness and safety of nitisinone in the treatment of AKU. Results were positive, and they will serve as the basis for the application for registration of nitisinone for treatment of AKU at the European Medicines Agency. Therefore, AKU might become a rare disease for which a cure will be found by 2020. We understand the natural history of the disease and the process of ochronosis much more, but at the same time there are still unanswered questions. One of them is the issue of the factors influencing the varying severity of the disease, since our recent genotype–phenotype study did not show that differences in residual homogentisic acid activity caused by the different mutations was responsible. Although nitisinone has proved to arrest the process of ochronosis, it has some unwanted effects and does not cure the disease completely. As such, enzyme replacement or gene therapy might become a new focus of AKU research, for which a novel suitable mouse model of AKU is available already. We believe that the story of AKU is also a story of effective collaboration between scientists and patients that might serve as an example for other rare diseases.
Keywords: alkaptonuria, rare disease, ochronosis, nitisinone, ochronotic pigment
Introduction
Alkaptonuria (AKU; OMIM 203500) is a classic example of a rare monogenic autosomal-recessive disease, characterized by high circulating homogentisic acid (HGA).1 In AKU, mutations of a gene coding for homogentisate dioxygenase (HGD; EC 1.13.11.5)2 cause a block in metabolism of tyrosine3 and accumulation of HGA, leading to a painful multisystemic disease (Figure 1). Normally from birth, homogentisic aciduria can be observed in patients, which manifests by darkening of the urine upon standing. Other related symptoms, such as ochronosis (pigmentation) of collagenous tissue and severe ochronotic osteoarthropathy in weight-bearing joints and the vertebral column,4 show delayed manifestations, usually from around 30 years of age. Ochronosis can occur in the eyes, ears, skin, and cardiac valves. Sometimes, black kidneys and prostate stones are found, and fractures as well as muscle and tendon ruptures are not rare. Disease severity varies among individual patients, even between siblings, and increases with age because of ongoing HGA accumulation. Usually, life span is not shortened in AKU, but quality of life is severely affected, mainly due to painful destruction of the joints, as their mechanical properties are altered due to ochronosis.
 |
Figure 1 Phenylalanine/tyrosine metabolism in brief and main AKU clinical symptoms. Notes: Metabolic block in alkaptonuria (AKU) affects homogentisic acid dioxygenase (HGD) and causes its accumulation, while nitisinone blocks 4-hydroxyphenylpyruvic acid dioxygenase and impedes formation of HGA. Ochronotic pigment is formed in connective tissue from benzoquinone acetic acid intermediate product. Main AKU clinical symptoms are homogentisic aciduria (urine gets dark upon standing, due to the presence of HGA) and ochronosis (dark pigmentation) of ear, eye, and joints. |
Role of Patient Organizations
Many people diagnosed with AKU have difficulty accessing efficient and effective care. This is mainly due to a lack of local expertise and knowledge when dealing with such a rare disease, and might result in a failure to detect complications of AKU. In 2003, the AKU Society (www.akusociety.org) was established, the first AKU-patient organization in the world, with the aim of providing support and information to patients and families, connecting AKU patients across the UK (and internationally), and promoting medical research, including for potential treatments. It took 10 years of tireless campaigning, funding research projects, and building partnerships, before (with the help of the AKU Society) the DevelopAKUre consortium was created, and a series of major international clinical trials started in Europe (Table 1). At the same time, the Robert Gregory National AKU Centre (NAC) was established in Liverpool (based at the Royal Liverpool University Hospital), named after the patient and cofounder of the AKU Society. The NAC has since then provided assessments and treatment for people with AKU living in England and Scotland. Subsequently, several sister AKU societies have been set up to raise awareness of AKU (AIMAKU-Italy, DSAKU-Germany, ALCAP-France, AKU Society Netherlands, AKU Society of Asia-Pacific, AKU Society North America, Black Bone Club [Slovakia], Jordanian AKU Society, AKU Society Brasil, and AKU Society India). They are run by patients and patient representatives who really understand AKU and the impact the disease can have on patients’ lives.
 |
Table 1 Summary of the Clinical Studies SONIA 1, SONIA 2, and SOFIA, Performed within the DevelopAKUre Project |
Is There a Cure for AKU?
So far, no approved specific cure exists for this disorder. Physiotherapy helps patients to maintain mobility and reduce the rate of musculoskeletal damage, pain is managed by strong painkillers, and joint-replacement surgery is used in advanced stages.5 However, it was shown recently that nitisinone, a potent inhibitor of 4-hydroxyphenylpyruvate dioxygenase (EC 1.13.11.27; Swedish Orphan Biovitrum), the enzyme that produces HGA, can arrest ochronosis and decrease the rate of progression of AKU (Figure 1).6 Nitisinone is already US Food and Drug Administration–approved for tyrosinemia type 1, while recently the EC-funded Seventh Framework Programme DevelopAKUre (2012–2019) tested its possible application in AKU (Table 1). The first of three DevelopAKUre studies, SONIA 1 confirmed that nitisinone decreases urine HGA in a dose-dependent and concentration-dependent manner.7,8 NAC patients from England and Scotland have access to nitisinone off-label (2 mg dose), and several important observations have been achieved over a 3-year period.6 In addition to decreasing urine HGA by >80%, 2 mg nitisinone daily inhibited ochronosis and significantly slowed the progression of AKU. The SONIA 2 long-term phase III clinical study (DevelopAKUre) aimed to assess the efficacy of nitisinone treatment on clinical outcomes, biochemical markers, and safety. Since the SONIA 2 preliminary results were positive, they will serve as a basis for the application for registration of nitisinone for AKU at the European Medicines Agency. The year 2020 is likely to be a breakthrough one for AKU patients.
The main side effect of nitisinone administration is tyrosinemia, which might cause corneal keratopathy and/or corneal opacity in the eyes of AKU patients. Although keratopathy and symptoms disappear as soon as 2 weeks after cessation of nitisinone,9,10 the use of the drug will probably require dietary monitoring and regular slit-lamp examinations, since keratopathy can be silent.6 Recently, it was shown that tyrosine/phenylalanine dietary restriction significantly reduced nitisinone-induced tyrosinemia in mice. Phenylalanine restriction alone proved ineffective. Similarly, protein restriction significantly reduced circulating tyrosine in AKU patients.11
The benefit of nitisinone for individuals with already-established pigmentation and tissue impregnation with already-advanced ochronosis with cross-linked HGA is most likely limited, requiring an early start to prevent morbidity. The search for alternative treatments, such as enzyme-replacement or gene therapy, expected to decrease HGA without causing tyrosinemia, represent new challenges in AKU. In any case, in order to prevent the damage caused by accumulated HGA or pigment, every therapy will need to be started early in life, as the disease is irreversible.
Natural History of AKU
Thanks to recent interest in this metabolic disorder, we are also learning a lot about the natural history of AKU. The SOFIA study, the last part of the DevelopAKUre project (Table 1), evaluated at what age the first signs of ochronosis begin and whether they present (also at microscopic level) before the onset of visible clinical symptoms of AKU, such as joint pain. Thirty patients from different age-groups (from 16 years) were recruited and several outcomes evaluated: ochronosis on ear biopsy, visible ochronosis quantification in eyes and ears, MRI examination, deviation in gait, modified questionnaire Alkaptonuria Severity Score Index (qAKUSSI), circulating and urine HGA, biomarkers of inflammation, cartilage damage, bone and other tissue, and quality of life.12
Pigmentation within ear biopsy was detected in a 20-year-old female, providing evidence that ochronosis can start in AKU patients before the age of 20 years. Eye ochronosis increased with age, and was first detected at age 22 years. Externally visible ear ochronosis was detected only after age 34 years.12 TIM, a marker of MMP-mediated titin degradation reflecting a remodeling of the cardiac sarcomere, could be an indirect marker of ochronosis of the cardiac valve, since it was increased with age in AKU patients, but not in controls. Interestingly, the inflammation biomarkers SAA and serum protein thiols increased with age, with no significant difference between AKU patients and controls.12 Despite the lack of change in the spine and joints at ages <30 years (as MRI analysis showed), gait analysis was abnormal early on, since young patients had mean-deviation profiles indicating that the gait had been affected at an early age.12
Analysis also showed that quality of life appears to deteriorate seriously from the third decade. As AKU patients get older, they score more highly on the qAKUSSI, regardless of sex, reflecting a worsening disease burden. It can be concluded that ochronosis starts at an early age, before adulthood, but a pediatric study is needed to assess the earliest age that it may start. Recently, the AKU Society was awarded a grant from the Childwick Trust that will fund a study into children with AKU: SOFIA–Paediatric. This new study will focus on children aged <16 years to find out the exact age at which damage starts, and thus to know the best age at which nitisinone therapy can be started.
Genetics of AKU and the HGD Mutation Database
AKU is very rare in most ethnic groups (one in 250,000–1,000,000), but in some countries, such as Slovakia and recently Jordan and India, it exhibits increased prevalence.13–16 Thanks to the work of patient organizations, effectively coordinated by the AKU Society, many new patients have been discovered. A new overview map has recently been created that reports 1,233 AKU patients worldwide (Figure 2, courtesy of AKU Society, Ciaran Scott). AKU patients are homozygous or compound heterozygous for pathogenic variants of the HGD gene14,17 (3q13.33), which is a single-copy gene composed of 14 exons.2,18,19 Mutation analysis in AKU is usually performed using DNA sequencing and multiplex ligation-dependent probe–amplification analysis, since recently several larger-scale deletions were discovered in AKU.20 The HGD Mutation Database was created (http://hgddatabase.cvtisr.sk),21 which summarizes results of mutation analysis from approximately 530 AKU patients reported so far. At present, 212 HGD gene variants have been reported. In addition to AKU-causing mutations, the ApreciseKUre database (http://www.bio.unisi.it/aku-db) facilitates collection and analysis of clinical and biochemical patients’ data, with an aim to share the results among registered users.22
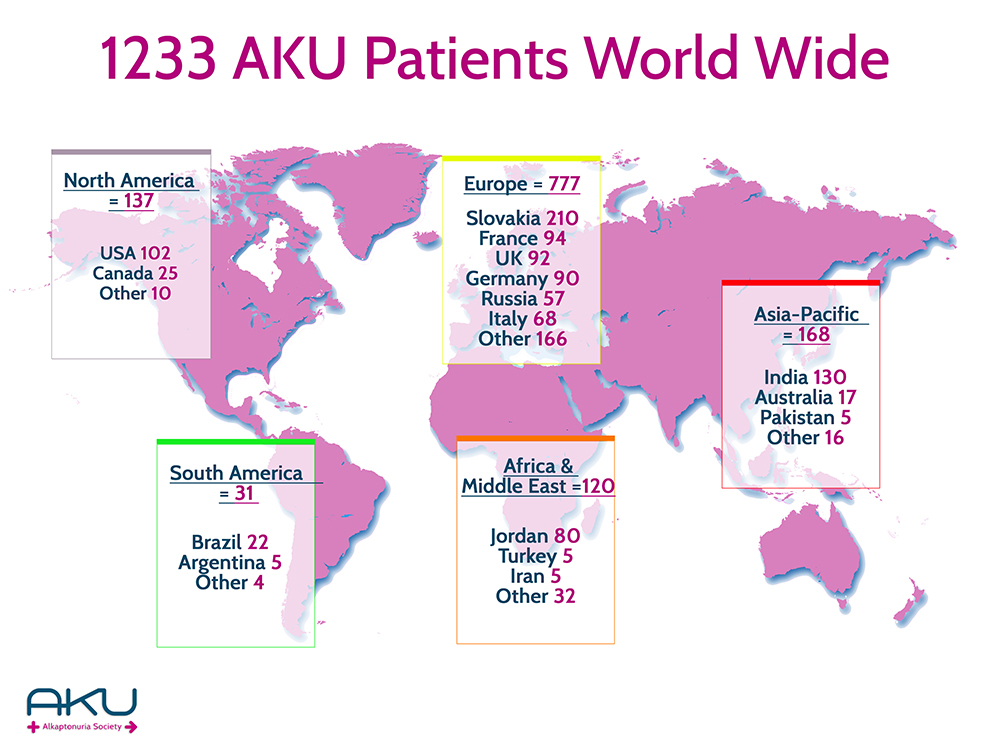 |
Figure 2 Overview of the number of patients with alkaptonuria reported worldwide. Note: Courtesy of AKU Society (www.akusociety.org), Ciarán Scott. |
The protomer of HGD is composed of 445 amino acids (NP_000178.2) and expressed mainly in kidneys and liver,2 but some expression has also been reported in the prostate, small intestine, colon,2 chondrocytes, synoviocytes, osteoblasts,23 and brain.24 However, recent detailed analysis in a new mouse model confirmed expression only in the liver and kidney cortex.25 This model included a β-galactosidase (lacZ) gene trap within the Hgd gene locus that has enabled precise localization of HGD expression. The active form of the HGD enzyme is a highly complex and dynamic hexamer composed of two disk-like trimers.26 The HGD protein has a low tolerance to mutation, and missense variants represent approximately 65% of all known AKU substitutions27,28 (Figure 3).
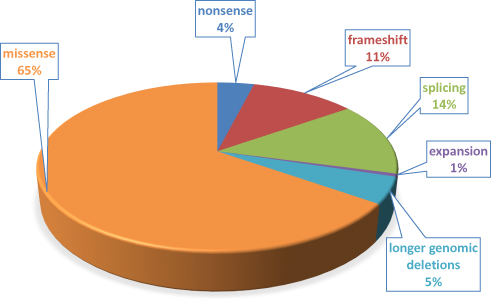 |
Figure 3 Proportion of different mutation types of 178 HGD gene variants affecting HGD function identified in about 530 AKU patients. Note: Data from HGD Mutation Database (http://hgddatabase.cvtisr.sk). |
Analysis of evolutionary conservation (Shannon entropy) showed that AKU variants were located at more conserved residue positions.20 Evaluation of the structural consequences of all HGD missense variants indicated that AKU-causing changes have an effect on protein folding and stability or interactions with other protomers or substrate. In other words, they decrease stability of individual protomers, disrupt protomer–protomer interactions, or modify residues in the active-site region.20,27 When a novel mutation is identified, it is important to distinguish causal AKU missense variants from nonpathogenic ones. In the case of AKU, mCSM-Stability, mCSM-PPI and mCSM-Lig predictions were shown to be the most effective tool to identify changes that could compromise enzyme activity.20
Frequently, AKU patients carry compound heterozygotes for two HGD gene variants. In such cases, it is difficult to estimate what the role of each missense variant is, since the hexamer could be formed either with monomers all affected by the same variant (homo-oligomer) or by two different ones (hetero-oligomer).29 Variants affecting two different regions could have additive destructive effect. Instead, the effects could partially compensate for those that belong to the same region. No tools to evaluate such events are available so far.
Nevertheless, with prospective development of novel treatment strategies for selected variants in AKU, it is important to be able to evaluate the effect of each variant on the HGD protein structure and function, as well as to estimate its possible residual activity. Missense variants in particular seem to be promising, since targeting HGD with pharmacological chaperones (small molecules helping structural stability) has predicted a total or partial rescue of enzyme activity.30
Genotype–Phenotype Correlations
Recently, we performed a detailed functional analysis of the three missense variants G161R, M368V, and A122V, and showed that all three mutants had significantly reduced activity compared to the wild-type enzyme, ranging from <1% of wild-type activity for G161R to 31% and 34% for A122V and M368V, respectively.20 This result confirmed previous in vitro studies on mutants that also showed different levels of residual activity.31 It can be speculated that different residual catalytic activity of the HGD proteins carrying different variants can manifest in different amounts of unmetabolized HGA. This in turn could lead to differences in serum and urine levels of HGA, and consequently differences in disease severity.
In order to follow on from this concept, for the three afore mentioned HGD variants and for the first time in AKU, we also performed a genotype–phenotype correlation analysis in 33 patients from the SONIA 2 clinical study. We saw that there was a small but statistically significant difference in urinary HGA excretion, corrected for dietary protein intake, in patients who carried variants with 1% and >30% residual HGD activity.20 However, serum levels or absolute urinary excretion of HGA showed no difference. Similarly, there were no differences in the tested AKU symptoms. This indicates that there is no direct effect of HGD mutation type on the variability of the AKU phenotype. Since high intra- and interindividual variability in urine HGA levels were also observed, we believe that mainly the dietary protein intake (ie, of tyrosine) and also the patient’s renal function are essential for determination of circulating concentrations of HGA, as well as for amounts of HGA excreted in urine.
In some AKU families with several affected siblings, considerable variability in the age of onset and severity is observed, despite the same genetic HGD mutations and similar diet (protein intake). As such, possibly other factors — genetic or biomechanical — are more important than differences in HGD variants in determining amounts of HGA that accumulate in the body of AKU patients, influencing a predisposition to connective-tissue damage or accelerating pigmentation.
The analysis of AKU siblings might be a powerful tool to help to identify such factors. DNA analysis of the whole genome or exomes is becoming a common and effective approach for in-depth phenotyping (molecular characterization) of the disease, as well as in pharmacogenomic prediction of the patient’s response to planned medical therapy. Detailed evaluation and comparison of clinical and genomic data of sibling patient pairs can be a key to understanding its variability. A disease severity index for AKU32 (AKUSSI) and other clinical and biochemical features can serve for careful monitoring of disease status, and the influence of non-AKU genes can be evaluated, including those responsible for aging and osteoarthritis susceptibility.
Studies in Animal Models
Studies in model organisms, especially AKU mice, have proved essential in studying AKU pathophysiology and testing potential treatments. The first mouse model was created by ethylnitrosourea (ENU) mutagenesis at the Pasteur Institute in Paris by Montagutelli et al,33 where it was identified thanks to the dark cage bedding due to elevated HGA. After identification of the murine hgd gene, the AKU mutation in this model was identified.34,35 Despite the elevated urine HGA, ochronosis was not observed in these animals. Microscopic pigmentation synonymous with human ochronosis was observed later on in a crossbred murine model of AKU, Hgd–/–Fah–/–, derived from the original ENU-mutagenesis mouse.36
In 2014 a consistent murine model of AKU was described in the BALB/c background, Hgd–/–, with no confounding hereditary tyrosinemia type I pathology.37 These animals demonstrate that ochronosis can be observed from early in life and increase linearly, while mean lifetime plasma-HGA levels remain relatively constant. Interestingly, end-stage pigmentation of the calcified and the hyaline cartilage, characteristic of AKU in humans, was not observed in mice. The shorter rodent life span can explain this fact, together with reduced joint-loading and/or faster cellular turnover that clears HGA-adducted proteins before polymerization.37 It was also shown that lifetime AKU mouse nitisinone treatment resulted in an 88% plasma-HGA reduction and completely prevented chondron pigmentation, confirming that if nitisinone is started early, it can arrest the progress of ochronosis.37,38 The results obtained with this model have encouraged human clinical trials (DevelopAKUre) and the off-label use of nitisinone.
ENU mutagenesis is not a targeted approach. Other hidden and uncharacterized mutations could potentially exist in models derived from ENU-mutagenesis mice that could also affect the phenotype. A new targeted mouse model of AKU (Hgd tm1a–/–) in the C57BL/6 background has been generated recently, using a mutant knockout-first Hgd allele obtained from the UC Davis Knockout Mouse Project Repository (https://www.komp.org).25 Urine HGA was elevated 100,000-fold (99,575 µmol/L) and plasma HGA elevated about 100-fold (100.5 µmol/L) in this model compared to C57BL/6 wild type (urine 0.8 µmol/L, plasma 1.7 µmol/L). Examination of knee joints in AKU mice showed ochronosis in calcified articular cartilage, first appearing at 9 weeks. Initially, dark pigment was localized to the pericellular matrix of chondrocytes (9–11 weeks) and was very infrequent, but at 20 and 40 weeks the number and intensity of pigmented chondrons was increased and showed advancement to the intracellular compartment. However, also in these animals, at 40 weeks ochronosis was still confined to calcified cartilage. Therefore, ochronosis in the mouse appears to represent the early stages of human joint pathophysiology in AKU.
The lacZ reporter enables both temporal and spatial histological localization of HGD, and it has been shown that in adults HGD is present in hepatocytes (throughout the liver parenchyma) and in proximal convoluted cells of the kidney cortex. Staining in embryos showed the first hepatic and kidney HGD expression at E12.5 and E15.5, respectively.25 All other embryonic tissue examined in frozen sections was negative.
Using Flp–Cre recombination, a double-transgenic conditional Hgd-knockout mouse was generated from this knockout-first Hgd tm1a allele. The use of Mx–1 Cre recombinase (MxCre) allows inducible and liver-specific knockout of Hgd mRNA by removal of the sixth Hgd exon in the conditional mouse model (Hgd tm1d MxCre±).25 Such knockout was sustained to 20 weeks post polyinosinic:polycytidylic acid injection (pIpC) and resulted in an approximate 80% reduction in liver Hgd mRNA, which subsequently caused plasma HGA to increase to a level comparable with AKU controls. Urinary HGA was increased too, but not to the level of AKU controls. This might have been caused by the fact that kidney HGD expression was maintained at the wild-type level, which on the other hand did not have an impact on plasma HGA, thus showing that kidney HGD is unlikely to rescue the AKU phenotype. Taken together, these results suggest that it is hepatic HGD that is crucial for HGA metabolism and the liver might be a suitable target for future AKU enzyme/gene therapy.25
In addition, the authors tried to answer an important question of how much liver HGD mRNA is required to rescue the phenotype. For this purpose, a short-term dose–response study was performed to estimate how much liver HGD mRNA would lower circulating HGA. A dose response was observed in both plasma- and urine-HGA levels,25 especially in lower pIpC–concentration groups (0.33–0.01 µg/g), but was not visible with higher concentrations (3.33 µg/g and 1 µg/g), most likely because the HGD mRNA:HGD protein ratio was not linear. However, the authors suggested that the minimum level of liver HGD mRNA required to eliminate circulating HGA and thus to rescue AKU must fall between 26% and 43%. This estimate corresponds well with the genotype–phenotype correlation study mentioned earlier, where we could see that 31%–33% residual activity of HGD in vitro did not reflect potential lower phenotype severity in patient carrying such mutations compared with those with only 1% residual activity.20
Another important finding for future gene/enzyme replacement therapy is that circulating HGA can reenter HGD-expressing cells to be metabolized by intracellular HGD. Therefore, it should not be necessary to repair 100% of liver cells in order to eliminate elevated circulating HGA.25
It is clear now that HGD is important very early in life, since HGD expression begins during embryonic development, and plasma HGA, measured at day 1 in Hgd tm1a–/– mice, shows levels threefold (308.8 umol/L) that of adult mice.25 This might suggest that any therapeutic strategy should ideally begin in early life or even at birth.25
Understanding the Process of Ochronosis and Specific Features of Ochronotic Cartilage
At some point in life of AKU patients, increased HGA levels in plasma and extracellular fluid start the process of ochronosis and the deposition of polymers of HGA as pigment in connective tissue, including cartilage, heart valves, and sclera. Many aspects of ochronosis have been elucidated (summarized recently by Ranganath et al39); however, we still do not understand fully the molecular mechanism of ochronotic pigmentation or the factors that influence its onset time and progress in general. This is complicated by the fact that AKU is not a local disease, as it is a complex inflammatory multisystemic disease involving many different organs.40 Although symptoms manifest clinically in joints and peripheral tissue, tyrosine catabolism and production of HGA, metabolites responsible for the pathology, occur mainly in the liver, with a minor contribution from the kidneys. Results of HGA measurements in 225 AKU patients from four recent clinical studies are summarized in Table 2.41 Generally, there was large variation in plasma- and urine-HGA concentrations among alkaptonuric patients. In serum, values were 7.6–82.4 µmol/L, and in urine 5,175–84,320 µmol/day, while normal urine values were 1.94±0.12 µmol/day or lower.41 As mentioned earlier, in patients this variability does not reflect the differences in residual activity of the HGD enzyme due to different mutations.20 It is believed that the variability depends mainly on daily intake of the proteins (in the diet) and on the effectiveness of renal excretion of accumulated HGA. This indicates that values actually can vary even in the same patient.
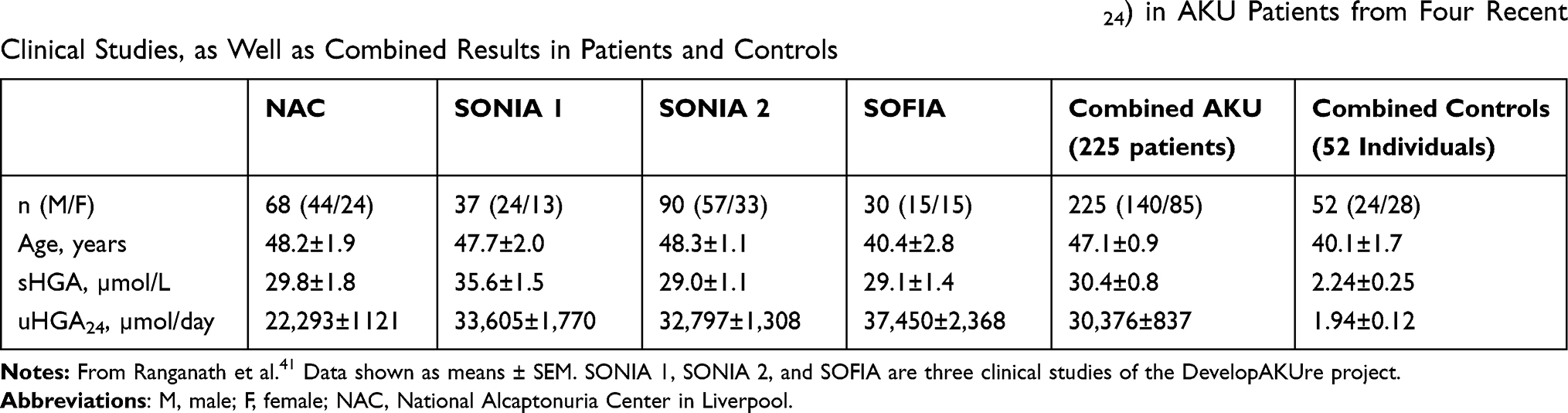 |
Table 2 Demographic Data, Serum HGA (sHGA) and Daily Urine HGA Excretion (uHGA24) in AKU Patients from Four Recent Clinical Studies, as Well as Combined Results in Patients and Controls |
The importance of kidney function manifested in the case of a 46-year-old man with AKU and diabetic renal failure. His plasma-HGA concentration was extremely high and his ochronosis progressed much more rapidly than that of his two AKU siblings. After renal transplantation, however, his plasma HGA normalized and HGA urinary excretion decreased substantially.42
The aging kidney manifests decreased excretion, which might lead to rising plasma-HGA concentration in AKU. Interestingly, this coincides with the observation of evident pathological, ochronotic changes that usually occur in humans in the beginning of the fourth decade. However, the process of changes in tissue at cellular/microscopic level starts earlier. At which exact age is not known yet, but the SOFIA study indicates it is before the age of 20 years.12
It has been shown that ochronosis localizes to areas of high mechanical loading, such as the large weight-bearing joints, but including also the aortic and mitral valves. It seems that both mechanical and biochemical matrix (extracellular matrix [ECM]) degradation contribute to the complex lesions in AKU cartilage.
Generally, initial pigmentation tends to be focal and is located in individual chondrocytes and their territorial matrix in calcified cartilage. In following stages, it proliferates throughout the hyaline cartilage in either granular or homogeneous conglomerates.43
It has been shown that the ECM is initially resistant to pigmentation, indicating that there are some protective mechanisms. ECM components are assembled into complex structures associated with fibrous proteins, proteoglycans, and glycosaminoglycans (GAGs). Following breakdown or loss of these constituents, pigment can access binding sites, which have been shown to be associated with the ultrastructural periodicity of collagen fibrils.44 This has also been described as the “exposed-collagen hypothesis”.45 It is not clear yet whether HGA, its oxidized intermediate benzoquinoneacetic acid,or directly ochronotic pigment is bound to collagenous matrices.39 Depletion of glycosaminoglycans not only triggers but also accelerates and amplifies degeneration in AKU cartilage.46
A model of ochronosis progression in articular cartilage has been proposed.39,47 Ochronosis starts in calcified cartilage with the deposition of pigment in individual chondrocytes and their territorial matrix. Pigmentation causes focal increases in stiffness, and load distribution is changed. Subsequently, ochronosis also spreads toward calcified matrix chondrons, and then proliferates throughout the hyaline cartilage. Ochronotic cartilage shields are not able to provide underlying bone with the necessary protection from normal mechanical loading, and aggressive resorption of the subchondral plate starts, which involves calcified cartilage and bone. The pigmented shell of the remaining articular cartilage is rather stiff and fails catastrophically. Pigmented cartilage impacts on the underlying trabecular bone, is embedded in the marrow space, and its fragments are present in synovial tissue.47 Ochronosis thus causes painful destruction of large weight-bearing joints and fusion of the vertebrae. The whole process is accompanied by chronic low-grade inflammation and amyloidosis.48,49
Once ochronosis has begun, its progress is fast and most likely not fully reversible, despite some indications of some reversal of pigment by unknown mechanisms.6 If there are some reparative responses, they are not sufficient to restore tissue functionality.
Brunetti et al50 showed recently that bone impairment can be attributed to high osteoclastic activity in AKU. Elevated levels of RANKL and reduced amounts of osteoprotegerin were observed in sera of AKU patients.50 In brief, RANKL binds to RANK receptors and promotes osteoclast precursor differentiation and fusion, as well as stimulates mature osteoclasts to resorb bone. On the contrary, osteoprotegerin competes with RANKL in binding to RANK, preventing a pro-osteoclastogenic effect. Interestingly, spontaneous osteoclastogenesis and increased percentage of osteoclast precursors have also been found in AKU children/young AKU patients,50 confirming the need for early intervention in AKU.
Characteristics of AKU cartilage were recently studied using thermal (thermogravimetry and differential scanning calorimetry) and rheological analysis.51 Drastic changes in thermal and rheological parameters were found, indicating that AKU tissue is not able to enhance chondrocyte metabolism and produce proteoglycans. Consequently, its stiffness drastically rises while dissipative and lubricant roles diminish, manifested also as destruction of the cell-matrix microstructure and disruption of the superficial layer. HGA-treated healthy cartilage manifested the same features, indicating that HGA is actually the toxic agent responsible for alterations in cartilage in AKU.51
There have been some specific features of AKU cartilage described recently. In normal cartilage, collagen fibers were cross-linked covalently to other-type collagen molecules, especially to homotypic polymers of type II collagen, whereas in AKU cartilage high amounts of type III collagen were present.52 The normal type II collagen was the main collagen in healthy controls, while some type III was present in osteoarthritis too. This might indicate that in this case there is also a repair process of matrix damage, but to a lesser extent than in AKU. Millucci et al52 suggested that matrix damage due to HGA might trigger a chondrocyte-healing response, in which type III collagen synthesized in mature articular cartilage plays an important role. It acts as a covalent modifier that may add cohesion to a weakened collagen type II fibril network.
AKU is one of the examples that confirms that the research on extreme phenotypes in monogenic diseases can help to elucidate the molecular pathogenesis of more common disorders. Investigating ochronosis has already provided important insights into the pathogenesis of osteoarthritis. Several previously unrecognized pathophysiological features of the human osteoarthritis phenotype have already been identified from studies of AKU. As an example can serve the aforementioned complete resorption of the subchondral plate,43 as well as trabecular excrescences, which are novel microanatomical structures that were first found abundant in AKU and subsequently at a lower frequency in osteoarthritis.53
Acknowledgment
The European Commission Seventh Framework Program funding granted in 2012 supported the SONIA 1, SONIA 2, and SOFIA studies (DevelopAKUre, project 304985).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Garrod AE. Croonian lectures on inborn errors of metabolism, lecture II: alkaptonuria. Lancet. 1908;2:73–79.
2. Fernández-Cañón JM, Granadino B, Beltrán-valero de Bernabé D, et al. The molecular basis of alkaptonuria. Nat Genet. 1996;14(1):19–24. doi:10.1038/ng0996-19
3. La Du BN, Zannoni VG, Laster L, Seegmiller JE. The nature of the defect in tyrosine metabolism in alcaptonuria. J Biol Chem. 1958;230(1):251–260.
4. Zannoni VG, Lomtevas N, Goldfinger S. Oxidation of homogentisic acid to ochronotic pigment in connective tissue. Biochim Biophys Acta. 1969;177(1):94–105. doi:10.1016/0304-4165(69)90068-3
5. Ranganath LR, Jarvis JC, Gallagher JA. Recent advances in management of alkaptonuria (invited review; best practice article). J Clin Pathol. 2013;66(5):367–373. doi:10.1136/jclinpath-2012-200877
6. Ranganath LR, Khedr M, Milan AM, et al. Nitisinone arrests ochronosis and decreases rate of progression of Alkaptonuria: evaluation of the effect of nitisinone in the United Kingdom National Alkaptonuria Centre. Mol Genet Metab. 2018;125(1–2):127–134. doi:10.1016/j.ymgme.2018.07.011
7. Ranganath LR, Milan AM, Hughes AT, et al. Suitability Of Nitisinone In Alkaptonuria 1 (SONIA 1): an international, multicentre, randomised, open-label, no-treatment controlled, parallel-group, dose-response study to investigate the effect of once daily nitisinone on 24-h urinary homogentisic acid excretion in patients with alkaptonuria after 4 weeks of treatment. Ann Rheum Dis. 2016;75(2):362–367. doi:10.1136/annrheumdis-2014-206033
8. Olsson B, Cox TF, Psarelli EE, et al. Relationship between serum concentrations of nitisinone and its effect on homogentisic acid and tyrosine in patients with alkaptonuria. JIMD Rep. 2015;24:21–27.
9. Hughes AT, Milan AM, Davison AS, et al. Serum markers in alkaptonuria: simultaneous analysis of homogentisic acid, tyrosine and nitisinone by liquid chromatography tandem mass spectrometry. Ann Clin Biochem. 2015;52(Pt 5):597–605. doi:10.1177/0004563215571969
10. Hughes AT, Milan AM, Christensen P, et al. Urine homogentisic acid and tyrosine: simultaneous analysis by liquid chromatography tandem mass spectrometry. J Chromatogr B Analyt Technol Biomed Life Sci. 2014;963:106–112. doi:10.1016/j.jchromb.2014.06.002
11. Hughes JH, Wilson PJM, Sutherland H, et al. Dietary restriction of tyrosine and phenylalanine lowers tyrosinaemia associated with nitisinone therapy of alkaptonuria. J Inherit Metab Dis. 2019. doi:10.1002/jimd.12172
12. Cox TF, Psarelli EE, Taylor S, et al. Subclinical Ochronosis Features In Alkaptonuria: a Cross-Sectional Study. BMJ Innovations. 2019. doi:10.1136/bmjinnov-2018-000324
13. Al-Sbou M, Mwafi N. Nine cases of Alkaptonuria in one family in southern Jordan. Rheumatol Int. 2012;32(3):621–625. doi:10.1007/s00296-010-1701-1
14. Janocha S, Wolz W, Srsen S, et al. The human gene for alkaptonuria (AKU) maps to chromosome 3q. Genomics. 1994;19(1):5–8. doi:10.1006/geno.1994.1003
15. Sakthivel S, Zatkova A, Nemethova M, Surovy M, Kadasi L, Saravanan MP. Mutation screening of the HGD gene identifies a novel alkaptonuria mutation with significant founder effect and high prevalence. Ann Hum Genet. 2014;78(3):155–164. doi:10.1111/ahg.12055
16. Srsen S, Varga F. Screening for alkaptonuria in the newborn in Slovakia. Lancet. 1978;2(8089):576. doi:10.1016/S0140-6736(78)92910-0
17. Pollak MR, Chou YH, Cerda JJ, et al. Homozygosity mapping of the gene for alkaptonuria to chromosome 3q2. Nat Genet. 1993;5(2):201–204. doi:10.1038/ng1093-201
18. Fernández-Cañón JM, Peñalva MA. Molecular characterization of a gene encoding a homogentisate dioxygenase from Aspergillus nidulans and identification of its human and plant homologues. J Biol Chem. 1995;270(36):21199–21205. doi:10.1074/jbc.270.36.21199
19. Granadino B, Beltrán-valero de Bernabé D, Fernández-Cañón JM, Peñalva MA, Rodríguez de Córdoba S. The human homogentisate 1,2-dioxygenase (HGO) gene. Genomics. 1997;43(2):115–122. doi:10.1006/geno.1997.4805
20. Ascher DB, Spiga O, Sekelska M, et al. Homogentisate 1,2-dioxygenase (HGD) gene variants, their analysis and genotype-phenotype correlations in the largest cohort of patients with AKU. Eur J Hum Genet. 2019;27(6):888–902. doi:10.1038/s41431-019-0354-0
21. Zatkova A, Sedlackova T, Radvansky J, et al. Identification of 11 novel homogentisate 1,2 dioxygenase variants in alkaptonuria patients and establishment of a novel LOVD-based HGD mutation database. JIMD Rep. 2012;4:55–65.
22. Spiga O, Cicaloni V, Bernini A, Zatkova A, Santucci A. ApreciseKUre: an approach of precision medicine in a rare disease. BMC Med Inform Decis Mak. 2017;17(1):42. doi:10.1186/s12911-017-0438-0
23. Laschi M, Tinti L, Braconi D, et al. Homogentisate 1,2 dioxygenase is expressed in human osteoarticular cells: implications in alkaptonuria. J Cell Physiol. 2012;227(9):3254–3257. doi:10.1002/jcp.24018
24. Bernardini G, Laschi M, Geminiani M, et al. Homogentisate 1,2 dioxygenase is expressed in brain: implications in alkaptonuria. J Inherit Metab Dis. 2015;38(5):807–814. doi:10.1007/s10545-015-9829-5
25. Hughes JH, Liu K, Plagge A, et al. Conditional targeting in mice reveals that hepatic homogentisate 1,2-dioxygenase 1 activity is essential in reducing circulating homogentisic acid and for effective therapy 2 in the genetic disease alkaptonuria. Hum Mol Genet. 2019. doi:10.1093/hmg/ddz234
26. Titus GP, Mueller HA, Burgner J, Rodríguez de Córdoba S, Peñalva MA, Timm DE. Crystal structure of human homogentisate dioxygenase. Nat Struct Biol. 2000;7(7):542–546. doi:10.1038/76756
27. Nemethova M, Radvanszky J, Kadasi L, et al. Twelve novel HGD gene variants identified in 99 alkaptonuria patients: focus on ‘black bone disease’ in Italy. Eur J Hum Genet. 2016;24(1):66–72. doi:10.1038/ejhg.2015.60
28. Zatkova A. An update on molecular genetics of Alkaptonuria (AKU). J Inherit Metab Dis. 2011;34(6):1127–1136. doi:10.1007/s10545-011-9363-z
29. Gupta V, Kalaiarasan P, Faheem M, Singh N, Iqbal MA, Bamezai RN. Dominant negative mutations affect oligomerization of human pyruvate kinase M2 isozyme and promote cellular growth and polyploidy. J Biol Chem. 2010;285(22):16864–16873. doi:10.1074/jbc.M109.065029
30. Bernini A, Galderisi S, Spiga O, et al. Toward a generalized computational workflow for exploiting transient pockets as new targets for small molecule stabilizers: application to the homogentisate 1,2-dioxygenase mutants at the base of rare disease Alkaptonuria. Comput Biol Chem. 2017;70:133–141. doi:10.1016/j.compbiolchem.2017.08.008
31. Rodriguez JM, Timm DE, Titus GP, et al. Structural and functional analysis of mutations in alkaptonuria. Hum Mol Genet. 2000;9(15):2341–2350. doi:10.1093/oxfordjournals.hmg.a018927
32. Langford B, Besford M, Hall A, et al. Alkaptonuria severity score index revisited: analysing the AKUSSI and its subcomponent features. JIMD Rep. 2018;41:53–62.
33. Montagutelli X, Lalouette A, Coude M, Kamoun P, Forest M, Guenet JL. aku, a mutation of the mouse homologous to human alkaptonuria, maps to chromosome 16. Genomics. 1994;19(1):9–11. doi:10.1006/geno.1994.1004
34. Manning K, Fernández-Cañón JM, Montagutelli X, Grompe M. Identification of the mutation in the alkaptonuria mouse model. Mutations in brief no. 216. Online. Hum Mutat. 1999;13(2):171. doi:10.1002/(SICI)1098-1004(1999)13:2<171::AID-HUMU15>3.0.CO;2-W
35. Schmidt SR, Gehrig A, Koehler MR, Schmid M, Muller CR, Kress W. Cloning of the homogentisate 1,2-dioxygenase gene, the key enzyme of alkaptonuria in mouse. Mamm Genome. 1997;8(3):168–171. doi:10.1007/s003359900383
36. Taylor AM, Preston AJ, Paulk NK, et al. Ochronosis in a murine model of alkaptonuria is synonymous to that in the human condition. Osteoarthritis Cartilage. 2012;20(8):880–886. doi:10.1016/j.joca.2012.04.013
37. Preston AJ, Keenan CM, Sutherland H, et al. Ochronotic osteoarthropathy in a mouse model of alkaptonuria, and its inhibition by nitisinone. Ann Rheum Dis. 2014;73(1):284–289. doi:10.1136/annrheumdis-2012-202878
38. Keenan CM, Preston AJ, Sutherland H, et al. Nitisinone arrests but does not reverse ochronosis in Alkaptonuric mice. JIMD Rep. 2015;24:45–50.
39. Ranganath LR, Norman BP, Gallagher JA. Ochronotic pigmentation is caused by homogentisic acid and is the key event in alkaptonuria leading to the destructive consequences of the disease-A review. J Inherit Metab Dis. 2019;42(5):776–792. doi:10.1002/jimd.v42.5
40. Helliwell TR, Gallagher JA, Ranganath L. Alkaptonuria–a review of surgical and autopsy pathology. Histopathology. 2008;53(5):503–512. doi:10.1111/j.1365-2559.2008.03000.x
41. Ranganath LR, Milan AM, Hughes AT, et al. Homogentisic acid is not only eliminated by glomerular filtration and tubular secretion but also produced in the kidney in alkaptonuria. JIMD. 2019. doi:10.1002/jimd.12181
42. Introne WJ, Phornphutkul C, Bernardini I, McLaughlin K, Fitzpatrick D, Gahl WA. Exacerbation of the ochronosis of alkaptonuria due to renal insufficiency and improvement after renal transplantation. Mol Genet Metab. 2002;77(1–2):136–142. doi:10.1016/S1096-7192(02)00121-X
43. Taylor AM, Boyde A, Wilson PJ, et al. The role of calcified cartilage and subchondral bone in the initiation and progression of ochronotic arthropathy in alkaptonuria. Arthritis Rheum. 2011;63(12):3887–3896. doi:10.1002/art.30606
44. Taylor AM, Wlodarski B, Prior IA, et al. Ultrastructural examination of tissue in a patient with alkaptonuric arthropathy reveals a distinct pattern of binding of ochronotic pigment. Rheumatology (Oxford). 2010;49(7):1412–1414. doi:10.1093/rheumatology/keq027
45. Gallagher JA, Dillon JP, Sireau N, Timmis O, Ranganath LR. Alkaptonuria: an example of a “fundamental disease”–A rare disease with important lessons for more common disorders. Semin Cell Dev Biol. 2016;52:53–57. doi:10.1016/j.semcdb.2016.02.020
46. Taylor AM, Hsueh MF, Ranganath LR, et al. Cartilage biomarkers in the osteoarthropathy of alkaptonuria reveal low turnover and accelerated ageing. Rheumatology (Oxford). 2017;56(1):156–164. doi:10.1093/rheumatology/kew355
47. Gallagher JA, Taylor AM, Boyde A, Jarvis JC, Ranganath LR. Recent advances in understanding the pathogenesis of ochronosis. Reumatologia. 2012;50(4):316–323.
48. Millucci L, Braconi D, Bernardini G, et al. Amyloidosis in alkaptonuria. J Inherit Metab Dis. 2015;38(5):797–805. doi:10.1007/s10545-015-9842-8
49. Millucci L, Spreafico A, Tinti L, et al. Alkaptonuria is a novel human secondary amyloidogenic disease. Biochim Biophys Acta. 2012;1822(11):1682–1691. doi:10.1016/j.bbadis.2012.07.011
50. Brunetti G, Tummolo A, D’Amato G, et al. Mechanisms of Enhanced Osteoclastogenesis in Alkaptonuria. Am J Pathol. 2018;188(4):1059–1068. doi:10.1016/j.ajpath.2017.12.008
51. Bernardini G, Leone G, Millucci L, et al. Homogentisic acid induces morphological and mechanical aberration of ochronotic cartilage in alkaptonuria. J Cell Physiol. 2019;234(5):6696–6708. doi:10.1002/jcp.27416
52. Millucci L, Bernardini G, Spreafico A, et al. Histological and ultrastructural characterization of Alkaptonuric tissues. Calcif Tissue Int. 2017;101(1):50–64. doi:10.1007/s00223-017-0260-9
53. Taylor AM, Boyde A, Davidson JS, Jarvis JC, Ranganath LR, Gallagher JA. Identification of trabecular excrescences, novel microanatomical structures, present in bone in osteoarthropathies. Eur Cell Mater. 2012;23:
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.