Back to Journals » International Journal of Nanomedicine » Volume 17
Albumins as Extracellular Protein Nanoparticles Collaborate with Plasma Ions to Control Biological Osmotic Pressure
Authors Zheng Z, Wang Y ![]() , Li M, Li D
, Li M, Li D ![]() , Nie A, Chen M, Ruan Q, Guo Y, Guo J
, Nie A, Chen M, Ruan Q, Guo Y, Guo J ![]()
Received 23 July 2022
Accepted for publication 3 October 2022
Published 11 October 2022 Volume 2022:17 Pages 4743—4756
DOI https://doi.org/10.2147/IJN.S383530
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Anderson Oliveira Lobo
Zihui Zheng,1,2,* Yuanyuan Wang,1,2,* Meng Li,3,* Dongfang Li,1,2 Aobo Nie,1,2 Miao Chen,1,2 Qinli Ruan,1,2 Yichen Guo,4 Jun Guo1,2
1School of Medicine & Holistic Integrative Medicine, Nanjing University of Chinese Medicine, Nanjing, Jiangsu, People’s Republic of China; 2Key Laboratory of Drug Target and Drug for Degenerative Disease, Nanjing University of Chinese Medicine, Nanjing, Jiangsu, People’s Republic of China; 3Children’s Hospital of Nanjing Medical University, Nanjing, Jiangsu, People’s Republic of China; 4Biomedical Engineering, University of Alabama at Birmingham School of Medicine, Birmingham, AL, USA
*These authors contributed equally to this work
Correspondence: Jun Guo, School of Medicine & Holistic Integrative Medicine, Nanjing University of Chinese Medicine, Nanjing, 210023, Jiangsu, People’s Republic of China, Tel +86 13813909055, Email [email protected] Yichen Guo Biomedical Engineering, University of Alabama at Birmingham School of Medicine, Birmingham, AL, USA, Email [email protected]
Introduction: Plasma albumins as protein nanoparticles (PNs) exert essential functions in the control of biological osmotic pressure (OP), being involved in regulating water metabolism, cell morphology and cell tension. Understanding how plasma albumins and different electrolytes co-determine biological OP effects is crucial for correct interpretation of hemodynamic disorders, and practical treatment of hypo/hyper-proteinemia.
Methods: Optical measurement based on intermediate filament (IF) tension probe was used for real-time evaluation of transmembrane osmotic effects in live cells. Ion fluorescent probes were employed to evaluate intracellular ion levels, and a current clamp was used to measure membrane potential, thus exploring association of electrochemical and osmotic effects.
Results: Albumins are involved in regulation of intracellular osmolarity by a quantitative relationship. Extracellular PNs can alter membrane potentials by adsorbing counterions, induce production of intracellular PNs and further control the opening of ion channels and ion flow, contributing to electrochemical and osmotic re-equilibrium. Furthermore, various ions interplay with extracellular PNs, showing different osmotic effects: increased levels of calcium ions result in a hypotonic effect, whereas potassium ions induce hyper-osmolarity.
Conclusion: Extracellular PNs and Ca2+/K+ display antagonistic or synergetic effects in regulating biological OP. Live cells can spontaneously regulate osmotic effects through changing membrane potential and controlling intracellular ion content. Various plasma components need to be comprehensively analyzed, further developing a diagnostic index that considers the biological OP effects of various blood components and improves the evaluation of symptoms and diseases, such as calcium/potassium-hemodynamic disorders and edema.
Keywords: biological OP, protein nanoparticle-ion interplay, albumin, electrochemical and osmotic re-equilibrium, intermediate filament tension
Graphical Abstract:
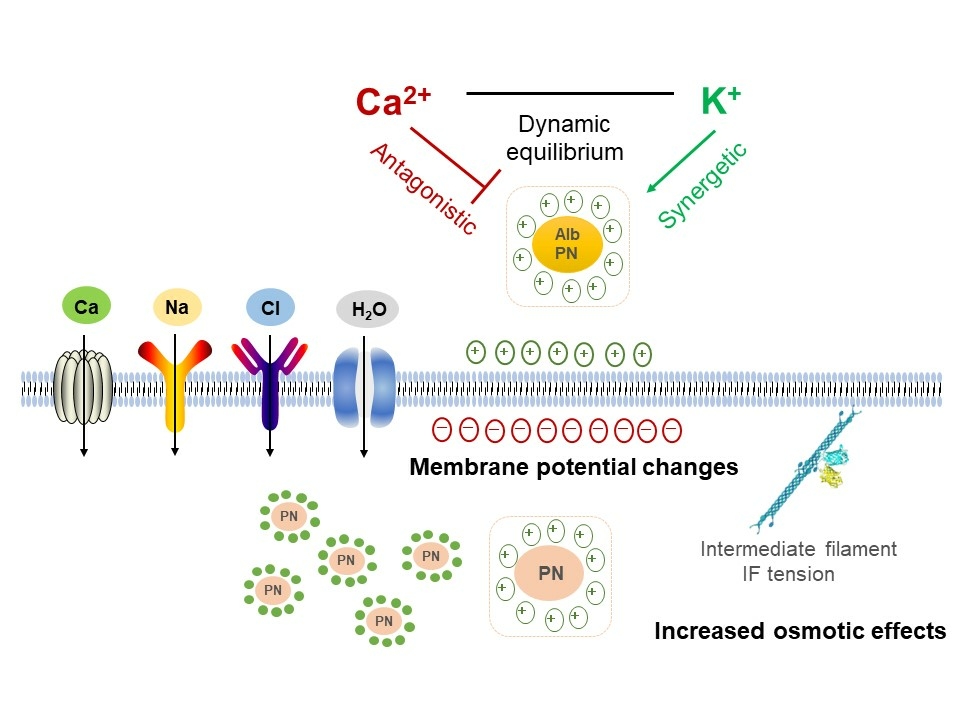
Introduction
Plasma albumins and electrolytes play essential roles in regulating osmotic pressure (OP) gradients in blood plasma, interstitial fluid, and intracellular fluid, physiological concentrations of which are maintained within narrow ranges.1–7 Maintenance of body-fluid homeostasis relies on modulating OP between the two sides of a cell membrane. According to the Van’t Hoff theory, OP values are determined by the amount of solution particles, including ions and colloid. Ion- and colloid-induced OP can be, respectively, measured by the freezing point osmometer and colloid osmometer.8–10 However, colloid components are considered to occupy a smaller percent of plasma components than ion particles; and it is assumed that colloid osmotic pressure (COP) values are not comparable to ion-induced OP values and have lesser effects.4,8 However, it raises the question of physical isotonicity dependent on semipermeable membranes whether presents “biological osmotic effects” in the body. Medical research has shown that plasma-protein-induced COP plays a crucial role that may underly undiscovered mechanical mechanisms in the body and that are different from single-ion-induced OP.
Recent studies have demonstrated that intracellular protein nanoparticles (PNs) exert amplified osmotic effects in live cells,11–14 termed PN-induced osmotic pressure (PN-OP). Specifically, PNs carrying negative charges adsorb cations and induce changes in free ion levels, as theoretically supported by the Gibbs-Donnan effect.15,16 Although this theory is still disputed due to serum albumins having little effect on blood OP, it provides the possibility that albumins could interplay with various ions to regulate biological OP.17,18 The theory of electric double layer could provide a reasonable explanation. High-valence ions, such as Ca2+, can be adsorbed by PNs and replace low-valence ions (Na+, K+), contributing to the decreasing number of ions in the two electric layers and compression of the diffused layer.19,20 PN-ion interplay induced biological OP effects are closely associated with symptoms and pathogenesis. For example, inflammasomes facilitate astrocytic swelling during pyroptosis,14 and PNs derived from cytoskeletal depolymerization and inflammasome induce pulmonary edema.21 Therefore, biological OP effects on transmembrane water-ion flux are closely associated with the opening of ion channels, whose dynamic equilibrium highly depends on PN and ion features.
Albumin acts as a plasma buffer, low levels of which are closely associated with decreased plasma OP, increased capillary filtration, and edema.3 Hypoalbuminemia is widely used as a predictive indicator of edema morbidity.22,23 However, human albumin supplementation to treat hypoalbuminemia is a controversial issue, possibly because hypoalbuminemia is often simply an epiphenomenon of another condition.24–29 Notably, abnormal albumin levels can be accompanied by pathophysiological alteration of electrolytes,17,18 suggesting that different plasma ions collaborate with albumin particles to play the amplified roles in control of biological OP, thus critically involved in biophysical regulation of cellular function and hemodynamic disorders.
Osmotic effects of the PN-ion interplay are difficult to evaluate in real-time using osmometry based on the concentration of plasma proteins and electrolytes. Thus, it is necessary to establish a biocompatible sensor for real-time monitoring and analysis of transmembrane osmotic effects in PN-ion mixed systems for live cells. In the present study, we used a fluorescence resonance energy transfer (FRET)-based intermediate filament (IF) tension probe to realize real-time evaluation of transmembrane OP gradients. Using this probe, we have illustrated synergetic or antagonistic OP regulation by PNs (albumins) and multiple electrolytes, and their stress-compensatory osmolarity control in the human body. The Gibbs-Donnan effect and the Van’t Hoff theory partly explain the mechanisms underlying the PN-OP effects under different ionic circumstances, in which calcium and potassium ions play crucial roles. Particularly, PN induced electrochemical and osmotic alterations regulate water-ion flux. These provide reasonable interpretations of osmotic effects in regulating water metabolism, cell swelling, and pathological morphogenesis. Our findings will provide better understanding of how plasma albumins and electrolytes co-determine hemodynamic disorders as well as how the body self-compensates to achieve osmotic equilibrium, which is crucial for the correct interpretation of hemodynamic clinical symptoms and providing practical albumin treatments corresponding to different electrolytic disorders.
Materials and Methods
Cell Culture and Reagents
Astrocytes of the U87 cell line were purchased from the American Type Culture Collection (ATCC, Manassas, USA). U87 cells were cultured in Dulbecco’s modified Eagle’s Medium (Invitrogen, New York, USA) containing 10% fetal bovine serum (GIBCO BRL, Grand Island, NY, USA) and 1% penicillin-streptomycin (Hyclone, South Logan, UT, USA) maintained at 37°C under 5% CO2. Modified Hanks’ balanced salt solution (MHBSS) was prepared to simulate blood plasma, to which was added albumins (Bovine Serum Albumin Fraction V, BioFRoxx, Einhausen, Germany) and Ca2+ (concentrations to 2.4 mmol/L). The electrolyte concentrations were adjusted using CaCl2, MgCl2, NaCl and KCl (Solarbio, Beijing, China). The pH and OP were maintained at 7.4 and 300 ± 10 mOsmol/kg. MHBSS components are shown in the Supplementary Materials And Methods Tables S1 and S2.
Probe Construction
Based on the FRET principle, a glial fibrillary acidic protein (GFAP) fluorescent tension probe was constructed using a cloning kit and restriction enzyme cloning technology.13,30 We constructed fluorescent sensors with circularly permutated cpVenus and cpCerulean (cpVenus–7aa–cpCerulean [cpstFRET]).
cpstFRET, FRET-AB, and FRAP Analyses
The FRET effectiveness was determined by the dipole angle between donor/eCFP and acceptor/eYFP.13,31–33 The donor and acceptor were tested using an argon laser at 458 and 514 nm, respectively. The CFP/FRET ratio was calculated using the relationship 1/R=ICerulean donor/IVenus acceptor. Acceptor photobleaching FRET (FRET-AB) and fluorescence recovery after photobleaching (FRAP) experiments were performed. The overall FRET efficiency was obtained based on the fluorescence intensities of the donor and acceptor before and after bleaching.
Measurement of Cytoplasmic OP and Protein Particle Counts
Cells under treatment were digested and centrifuged to obtain cell supernatants. The cytoplasmic OP was assessed using an Osmomat 3000 Freezing Point Osmometer (Gonotec, Berlin, Germany). The number of cytoplasmic nanoparticles (kilocycles per second, Kcps) was determined using a Nanosight NS300 (Malvern Instruments, Malvern, UK).
Calcium and Chloride Fluorescence Imaging
Fluo-4 AM and N-6-methoxyquinolyl acetoethyl ester (MQAE) fluorescent probes were used to perform Ca2+ and Cl− imaging, respectively.34,35 Cells were loaded with the fluorescence probe and detected with Thunder Imager (SP8; Leica). Fluo-4 was detected with excitation at 494 nm and emission at 516 nm, and MQAE was detected with excitation at 355 nm and emission at 460 nm.
Electrophysiology
The patch clamp technique was employed to record the membrane potentials of U87 cells. Cells were plated onto 9 mm glass coverslips at a suitable density, mounted in a chamber (Warner Instruments, Hamden, CT; U.S.A.), placed onto the stage of an inverted microscope, and perfused with the required solution. Measurements were obtained at room temperature in whole-cell configuration.21
For measurement, the cells were perfused with a solution containing (in mM): 144 NaCl, 4.3 KCl, 2.5 CaCl2, 1.1 MgCl2, 10 glucose, and 10 HEPES (pH 7.3), and mannitol was used to adjust the osmolality to 300–310 mosmol/kg. The concentrations of Ca2+, K+ and albumin were adjusted according to the experimental requirements. The patch electrodes were pulled from borosilicate capillary glass (1.5 mm outer diameter; 0.86 mm inner diameter; Sutter Instruments, USA) with a vertical two-stage pipette puller (P-1000; Narishige). Measurements were performed with glass pipettes (between 3 and 6 MΩ) filled with an intracellular solution comprising (in mM): 140 KCl, 2 MgCl2, 2 Na2ATP, 1 CaCl2, 5 Ethylene Glycol Tetraacetic Acid (EGTA), 5 glucose (pH 7.3, 290–300 mosmol/kg). All measurements were performed using a Multiclamp 700B amplifier at a data-sampling rate of 2 kHz with a Digidata 1550B Digitizer controlled using PClamp 10.6 software (Molecular Devices, Sunnyvale, CA, U.S.A.).
Statistical Analyses
FRET ratios were determined using Image J image analysis software, and the results of ion imaging were analyzed semi-quantitatively. Graph Pad Prism 7 software was used for statistical processing. Each experiment was repeated at least three times, and the data are expressed as Means ± standard error of mean (SEM). Unpaired t-test (two-tailed P value) was used to determine the statistical significance, with P<0.05 taken as statistically significant.
More detailed methods can be found in the Supplementary Materials and Methods.
Results
Osmotic Effects in Live Cells Result from Transmembrane OP Gradients and Intracellular PN Amount
Osmotic changes are closely associated with cell volume and IF tension, which requires cytoskeletal tractive force against outward osmotic force.14 Thus, to quantitatively analyze osmotic effects on IF tension, a glial fibrillary acidic protein (GFAP) FRET-based tension probe was deployed in astrocytes to provide real-time IF tension measurements based on optical variation.13 Astrocytes were selected based on their sensitivity to OP and the accompanying volume change. They were individually exposed to Hanks’ balanced salt solutions (HBSSs) having different NaCl concentrations corresponding to OP gradients of 150–450 mOsmol/kg. The GFAP tension remains stable under isotonic conditions (300 mOsmol/kg). However, it increases under extracellular hypoosmolarity and decreases under hyperosmotic conditions within 15 min (Figure 1A and B). This indicates that transmembrane OP differences are strongly correlated with IF tension, and a linear correlation between GFAP tension and NaCl concentration is observed (R2 = 0.9512, Figure 1D). Consistently, the correlation between GFAP tension and osmotic gradient is also observed when mannitol is used in the HBSS instead of NaCl (R2 = 0.9942, Figure 1C and E).
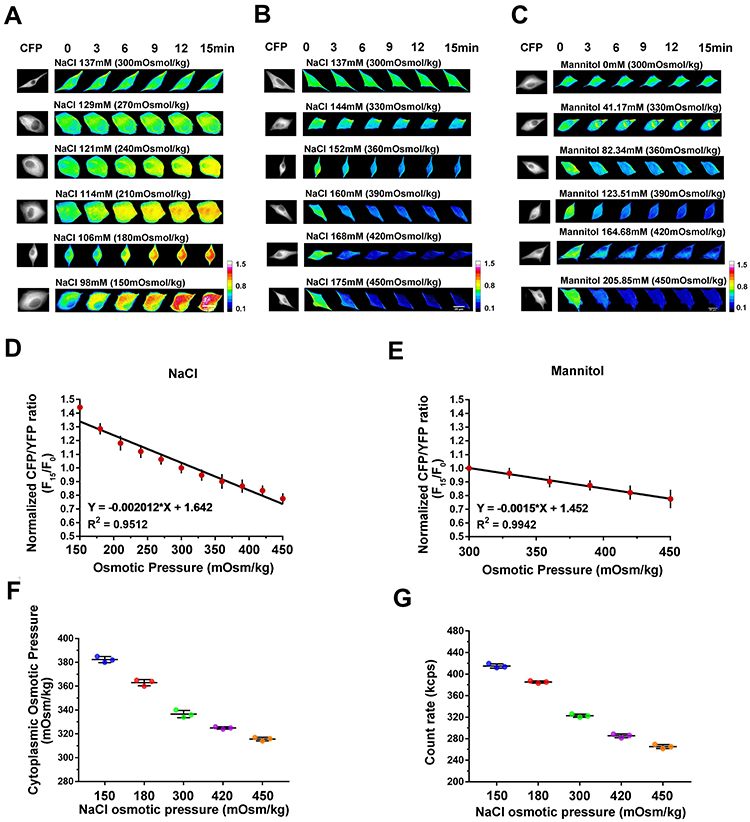 |
Figure 1 Relationships between extracellular OP gradient, GFAP tension, and intracellular OP. (A and B) A GFAP cpstFRET probe module was used to measure changes in IF tension in U87 cells. Representative images of normalized Cyan Fluorescence Protein (CFP)/FRET ratios of GFAP tension subjected to the NaCl solutions with different concentrations, including extracellular hypoosmolality (less than 300 mOsmol/kg) and hyperosmolality (higher than 300 mOsmol/kg) are shown. (C) Representative images of normalized CFP/FRET ratios of GFAP tension subjected to hyperosmotic mannitol solutions. Scale bar, 10 μm. (D) Linear correlation between GFAP tension and NaCl concentration. (E) Linear correlation between GFAP tension and mannitol concentration. (F) Cytoplasmic OP values as measured using a freezing point osmometer. (G) Intracellular PN count rates under different treatments. Data presented as mean ± SEM. |
As osmotic effects on live cells are determined by transmembrane oncotic pressure gradients rather than extracellular OP, we measured the cytoplasmic OP using a freezing point osmometer. Extracellular hypoosmotic stimuli promote an increase in cytoplasmic OP (Figure 1F), contributing to enhanced transmembrane OP gradients and stronger osmotic effects on cells. Furthermore, PNs are produced in cells under hypotonic stress, and the increased number of intracellular PNs along with the decreased extracellular OP (Figure 1G) impacts the oncotic pressure in the cells. These data demonstrate the generation of transmembrane OP gradients resulting from alterations of intracellular OP as well as PN amount. Transmembrane differences in OP induced by intracellular PNs determine the osmotic effects on live cells, which can be evaluated in real-time by monitoring IF tension.
Quantitative Analysis of IF Tension Regulated by Extracellular PNs and Different Ionic Components
Ions and PNs are predominant components of plasma and intracellular fluid in the human body. According to the Van’t Hoff theory, ions rather than PNs (or colloids) are the dominating factor controlling OP in the living body. However, the Gibbs-Donnan effect provides an alternative explanation whereby intracellular PNs play a vital role, as clarified in our recent study.13,14 To investigate whether extracellular PNs share similar mechanisms underlying the osmotic effects for live cells, astrocytes were treated with isotonic albumin in Modified Hanks’ balanced salt solution (MHBSS), which contains albumins in different concentrations. When extracellular albumins are present at physiological concentration (50 mg/mL), IF tension remains unchanged. However, hypoalbumins (albumins at 10 and 30 mg/mL) induce a reduction of IF tension, whereas hyperalbumins (albumins at 70 and 100 mg/mL) cause a tension increase (Figure 2A). A parabolic equation for the correlation between IF tension and extracellular PN concentrations was determined. Importantly, this positive correlation is linear for PN concentration and IF tensions (10–100 mg/mL, Figure 2B).
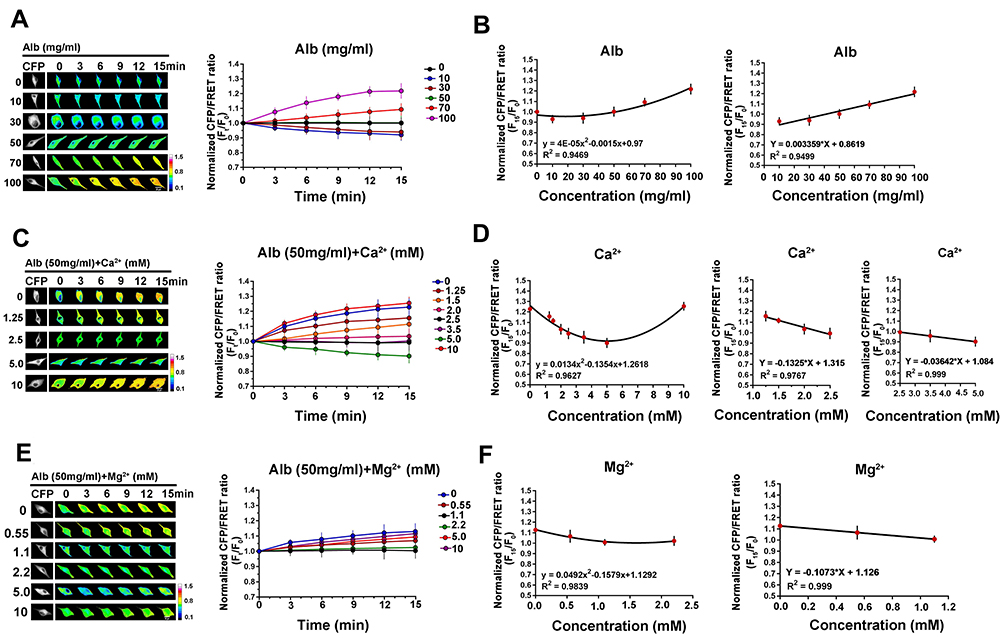 |
Figure 2 Quantitative analysis of IF tension regulated by albumin and divalent cations. (A) Representative images and mean values of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic MHBSS solutions with different albumin concentrations (0, 10, 30, 50, 70 and 100 mg/mL). (B) Relationship between GFAP tension and albumin concentration, showing the parabolic correlation of the experimental concentrations and the linear correlation in the pathophysiologic ranges (10–100 mg/mL albumin). (C) Representative images and mean values of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic solutions containing 50 mg/mL albumin (the normal physiological concentration) and different Ca2+ concentrations (mM as number indicators). (D) Relationship between GFAP tension and Ca2+ concentration in different experimental and pathophysiologic ranges. (E) Representative images and mean values of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic MHBSS solutions containing 50 mg/mL albumin and different Mg2+ concentrations (mM as number indicators). (F) Relationship between GFAP tension and Mg2+ concentration in different ranges. Scale bar, 10 μm. Data presented as mean ± SEM. |
Under physiological conditions, extracellular ion profiles remain in homeostasis. However, pathological data reveal hypo- or hyperalbuminemia accompanying varying plasma ion levels. According to the Gibbs-Donnan effect, PNs can regulate OP through ion adsorption. Thus, to determine the effects that interactions of different ions with extracellular PNs on osmotic regulation, cells were treated with electrolytes containing different biologically relevant cations at differing concentrations along with 50 mg/mL albumin at a constant extracellular OP of 300 mOsmol/kg. Ca2+ at typical physiological concentration (2.5 mM) has no effect on IF tension or isotonic equilibrium. However, abnormal Ca2+ levels cause IF tension changes (Figure 2C) due to transmembrane disequilibrium. IF tension and extracellular Ca2+ concentration is closely associated in the curvilinear relationship, determined by 0–10 mM of Ca2+. For the two pathophysiologic ranges of plasma Ca2+ concentration, ie, hypocalcemia (<2.5 mM) and hypercalcemia (2.5–5 mM), the negative correlation between IF tension and Ca2+ concentration is linear (Figure 2D). These data suggest that hypocalcemia can result in increased IF tension, corresponding to hyperosmolarity. The other divalent cation found in plasma, Mg2+, induces slight changes in IF tension under the pathophysiologic concentrations (other than 1.1 mM, Figure 2E and F).
In terms of monovalent cations, changes in extracellular K+ and Na+ levels also strongly impact cell IF tension. K+ at its typical physiological level (4.3mM) mixed with 50 mg/mL albumin presents an isotonic effect. However, lower K+ concentrations (0 and 2.15 mM) contribute to decreased IF tension, representing the hypotonic effects on live cells, while abnormally high K+ concentrations (5, 8.6, and 10 mM) increase IF tension in response to the hypertonic effects (Figure 3A). K+ concentration and IF tension present a linear correlation (R2 = 0.9735, Figure 3B). In addition, extracellular Na+ concentration negatively correlates with IF tension (R2 = 0.9591, Figure 3C and D). The univariate data reveal that IF tension, extracellular PN concentration and electrolyte concentration are strongly correlated, as shown by the linear regression equations and R2 values in Tables S3 and S4.
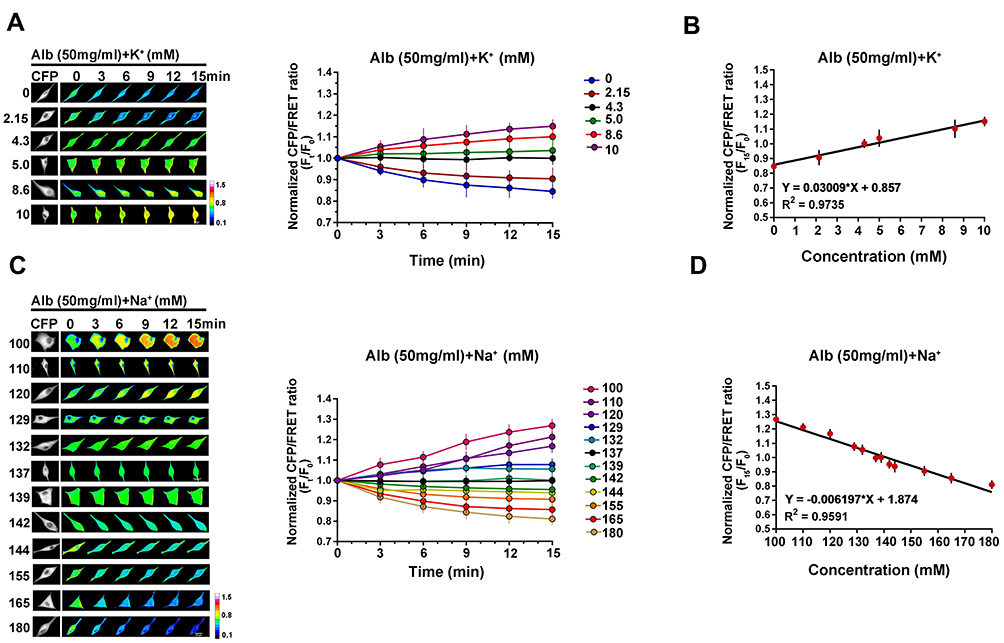 |
Figure 3 Quantitative analysis of IF tension regulated by monovalent cations. (A) Representative images and mean values of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic MHBSS solutions containing 50 mg/mL albumin and different K+ concentrations (mM as number indicators). (B) Linear relationship between GFAP tension and K+ concentration. (C) Representative images and mean values of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic MHBSS solutions containing 50 mg/mL albumin and different Na+ concentrations (mM as number indicators). (D) Linear relationship between GFAP tension and Na+ concentration. Scale bar, 10 μm. Data presented as mean ± SEM. |
Pathological Concentrations of Plasma Components Induce Transmembrane OP Gradients and Production of Intracellular PNs
As IF tension is closely related to transmembrane OP gradients, we sought to determine whether changes in extracellular components also affect cytoplasmic OP. When the albumin, Ca2+, and K+ concentrations of the simulated plasma solutions change, the cytoplasmic OP gradient changes (Figure 4A–C) in a manner consistent with that of IF tension. Previous reports have indicated that cytoplasmic OP may be regulated by intracellular PNs.12–14 Indeed, the amounts of cytoplasmic PNs correlate closely with IF tension and cytoplasmic OP (Figure 4D–F), indicating that extracellular PN and ion-induced transmembrane osmotic gradients could be caused by changes in the number of intracellular PNs.
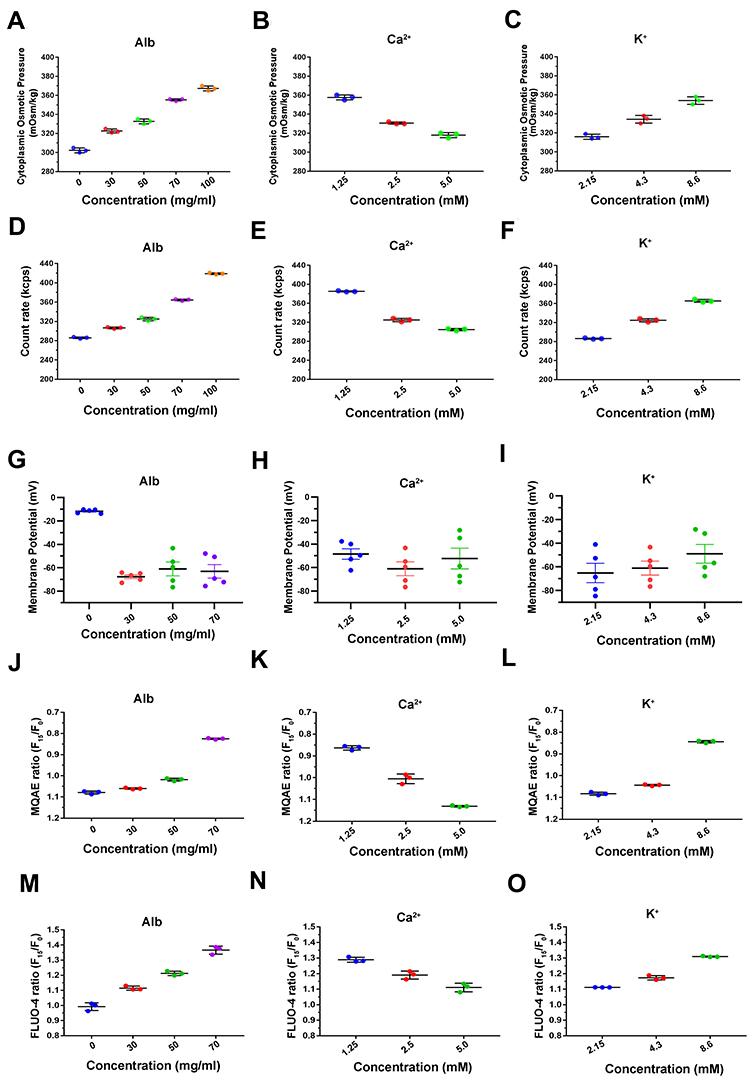 |
Figure 4 Osmotic effects for live cells derived from changes in intracellular OP, PN count, membrane potential and intracellular ion levels. The cytoplasmic OP values in response to extracellular variations of albumin (A), 50 mg/mL albumin and Ca2+ (B), 50 mg/mL albumin and K+ (C) as determined using a freezing point osmometer. Intracellular PN counts under the effects of different extracellular components, including albumin (D), 50 mg/mL albumin and Ca2+ (E), 50 mg/mL albumin and K+ (F). Membrane potential was measured by the current-clamp method under treatments with albumin (G), 50 mg/mL albumin and Ca2+ (H), and 50 mg/mL albumin and K+ (I). Chloridion imaging presents the relative MQAE fluorescence intensity ratio of 15 min (F15/F0) under different concentrations of extracellular albumin (J), 50 mg/mL albumin and Ca2+ (K), 50 mg/mL albumin and K+ (L). An increase in intracellular Cl− levels leads to a decrease in MQAE fluorescence value. Calcium imaging presents the relative FLUO-4 fluorescence intensity ratio of 15 min (F15/F0) for cells under different concentrations of extracellular albumin (M), 50 mg/mL albumin and Ca2+ (N), 50 mg/mL albumin and K+ (O). Data presented as mean ± SEM. |
Extracellular PNs Regulate Osmotic Effects by Inducing Changes in Membrane Potential
According to the Donnan effect, intracellular PNs can adsorb counter ions, leading to osmotic disequilibrium.13–15 We speculate that PNs induced by extracellular stimuli adsorb cations, disrupting the electrochemical equilibrium near membranes. To test this speculation, a current clamp was used to measure membrane potential. The data indicate that extracellular albumins cause membrane hyperpolarization independent of solute concentration (Figure 4G–I, Figure S1A–C). These results suggest that extracellular albumins change membrane potential, affecting cytoplasimc OP and IF tension.
Extracellular Components Regulate PN-OP Associated with Intracellular Ion Levels
Intracellular ion levels were also determined using chloridion and calcium imaging to clarify any effects they may have alongside those of extracellular ions. The results show that the intracellular Cl− and Ca2+ contents are positively correlated with extracellular albumin concentration (Figure 4J and M, Figure S2A and D). However, extracellular hyper-calcium inhibits cytoplasmic levels, while hypo-calcium promotes Cl− and Ca2+accumulation in cells (Figure 4K and N, Figure S2B and E). Intracellular Cl− and Ca2+ levels are positively correlated with extracellular K+ concentration (Figure 4L and O, Figure S2C and F).
Overall, changes in extracellular components affect the accumulation of various ions in cells, which is related to transmembrane OP gradients. When extracellular PNs are present, alterations in extracellular ion components affect transmembrane PN-OP, potentially due to changes in membrane potential and ion influx. Notably, Ca2+ and K+ appear to exert opposite effects on PN-OP regulation.
Extracellular PNs, Ca2+ and K+ Synergistically Regulate Transmembrane Osmotic Equilibria
As the above data appear to reveal contrary influences on transmembrane osmotic effects, we investigated the synergetic or antagonistic effects of extracellular albumins and Ca2+ on the regulation of PN-OP. Cells were treated with hypoalbuminous (30 mg/mL) and hyperalbuminous (70 mg/mL) solutions also containing Ca2+ at different concentrations (1.25, 2.5, and 5 mM). The IF tension data indicate two conditions of osmotic equilibrium, where isotonic IF tension is established within 15 mins. These conditions are 30 mg/mL albumin with 1.25 mM Ca2+ and 70 mg/mL albumin with 5 mM Ca2+ (Figure 5A). Compared with plasma component levels under normal physiological conditions (50 mg/mL albumins with 2.5 mM Ca2+), these two osmotically equilibrated states are similar in terms of cytoplasmic OP and intracellular PN count (Figure 5B and C), accompanied by membrane hyperpolarization (Figure 5D and E, Figure S1D and E). The intracellular Cl− and Ca2+ contents at the two equilibrium states seemed to change the least (Figure 5F–I). Likewise, hypo/hyper-albumins and K+ concentrations present two states of osmotic equilibrium: 30 mg/mL albumins with 8.6 mM K+ and 70 mg/mL albumins with 2.15 mM K+ (Figure 6A), which is correlated with cytoplasmic OP, PN count, membrane potential, and the intracellular Cl−/Ca2+ content data (Figure 6B–I, Figure S1F and G). These results suggest that hypoalbumins mixed with hypocalcium or hyperpotassium can provide osmotic equilibrium, whereas hyperalbumin mixed with hypercalcium or hypopotassium maintain the transmembrane isotonic states.
 |
Figure 5 Albumin and Ca2+ co-regulate osmotic effects. (A) Representative images and mean values of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic solutions with hyper/hypo-albumin and Ca2+. Scale bar, 10 μm. (B) Cytoplasmic OP values as measured using a freezing point osmometer. (C) Intracellular PN counts, ns, not significant. (D and E) Membrane potentials using the current-clamp method under different conditions. (F and G) Chloridion imaging micrographs and traces of relative MQAE fluorescence intensity ratio of 15 min (F15/F0). An increase in intracellular Cl− level leads to a decrease of MQAE fluorescence value. (H and I) Calcium imaging micrographs and traces of relative FLUO-4 fluorescence intensity ratio of 15 min (F15/F0). Scale bar, 100 μm. Data presented as mean ± SEM. |
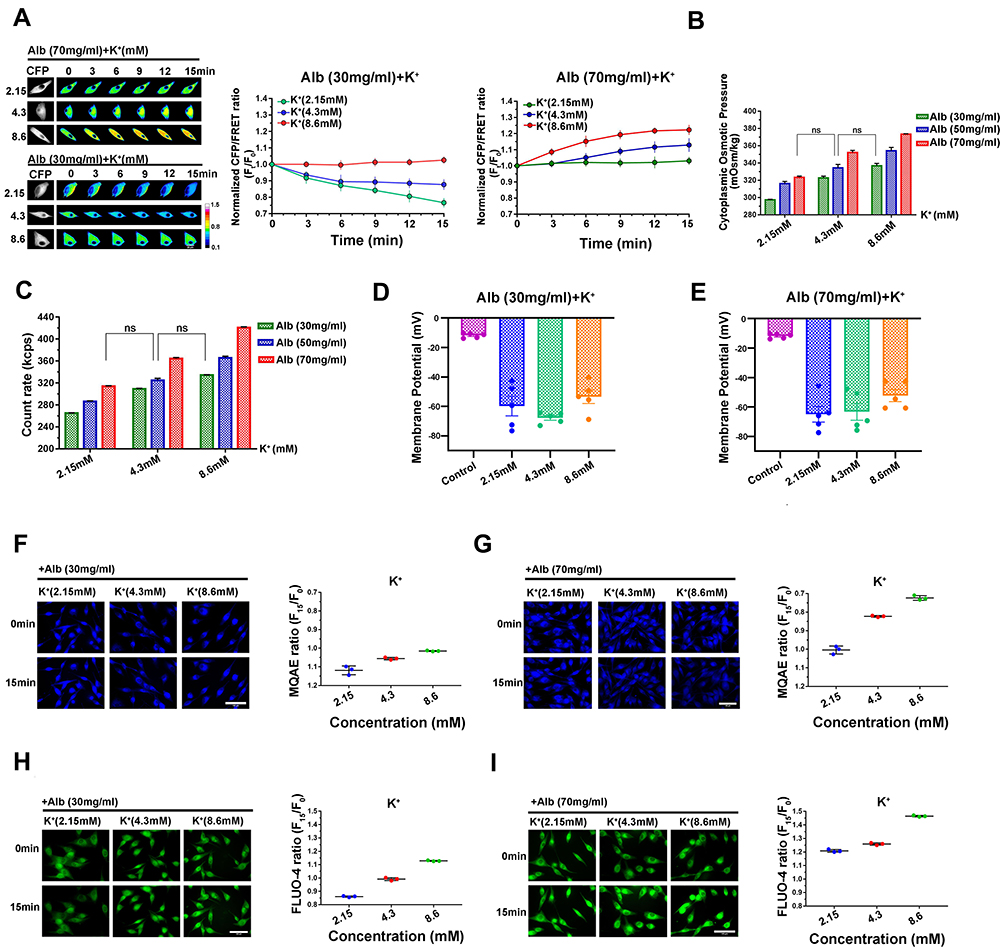 |
Figure 6 Albumin and K+ synergistically regulate transmembrane osmotic effects. (A) Representative images and mean values of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic solutions with different albumin and K+ concentrations. Scale bar, 10 μm. (B) Cytoplasmic OP values. (C) Intracellular PN count, ns, not significant. (D and E) Membrane potentials using the current-clamp method under different conditions. (F and G) Chloridion imaging micrographs and traces of relative MQAE fluorescence intensity ratio of 15 min (F15/F0). (H and I) Calcium imaging micrographs and traces of relative FLUO-4 fluorescence intensity ratio of 15 min (F15/F0). Scale bar, 100 μm. Data presented as mean ± SEM. |
We speculated that hypoalbuminemia-induced osmotic disequilibrium may be alleviated by adjusting extracellular Ca2+ and K+ levels. To assess this speculation, hypoalbumin (30 mg/mL) or physiologically normal albumin (50 mg/mL) was mixed with different concentrations of Ca2+ and K+ and the IF tension measured. Again, the tension results present two osmotically equilibrated conditions, ie, 50 mg/mL albumin with 1.25 mM Ca2+ and 2.15 mM K+, and 50 mg/mL albumin with 5 mM Ca2+ and 8.6 mM K+ (Figure 7). Overall, extracellular PNs, Ca2+, and K+ enable the collaborated regulation of transmembrane osmotic effects for cells. Ca2+, and K+ appear the opposite effects with albumins, possibly by adjusting membrane potentials and the amounts of intracellular PNs and ions, re-achieving transmembrane osmotic equilibrium.
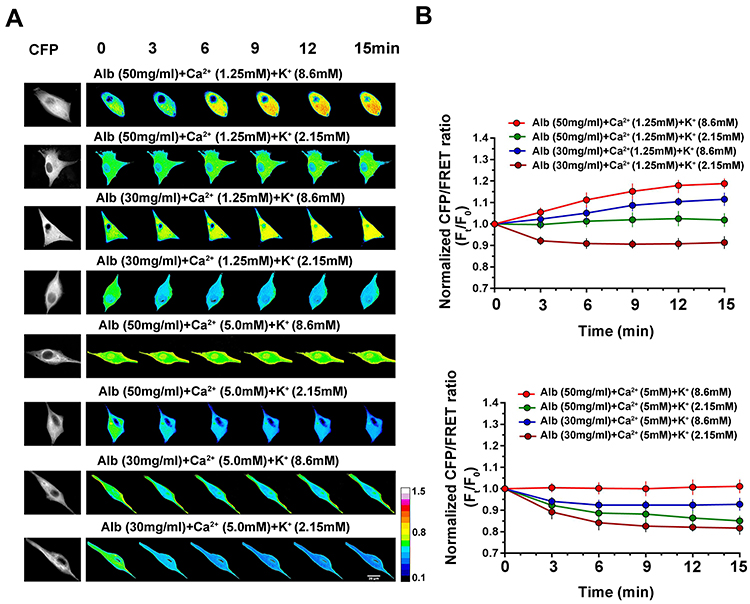 |
Figure 7 Synergistical effects of albumin, Ca2+, and K+ on IF tension. Representative images (A) and mean values (B) of normalized CFP/FRET ratios of GFAP tension in cells subjected to isotonic solutions with different albumin, Ca2+, and K+ concentrations. Scale bar, 10 μm. Data presented as mean ± SEM. |
Discussion
Previous studies have reported that physical OP can be determined from solution particles across semipermeable membrane. However, a new viewpoint of “biological OP” presents the interplay between protein nanoparticles and different ions to control osmotic effects in the body, dependent on changes in membrane potential and sensed by cell-stress-compensatory osmolarity. Changes in extracellular PN or ion component can induce corresponding cytoplasmic OP adjustment. Therefore, PNs crucially determine the osmotic effects in the body, reaching a dynamic osmotic equilibrium.
Albumins, as extracellular PNs, enable enhancement of transmembrane OP gradients accompanied by increasing IF tensions. Our results revealed that 300 mOsm/kg of ionic circumstance show the transmembrane isotonic equilibrium; however, only the presence of extracellular PNs (albumins) triggers the production of intracellular PNs and an increase of cytoplasmic OP. The amplified roles of PNs in regulating biological OP can be partly explained by the Gibbs-Donnan effect.15,16,36–38 PNs carry negative charges and adsorb cations to form the colloid-ion compassing group, changing the levels of free ions near cell membranes and membrane potential, which is closely associated with voltage-dependent ion channels, and constituting a vital mechanism underlying PN-OP in the body.
In the other case, abnormal contents of albumin, Ca2+ and K+ induce production of intracellular PNs, further driving the cytoplasmic PN-ion adsorbing. More importantly, extracellular albumins and intracellular PNs collaboratively contribute to electrochemical changes and the resultant ion rearrangement across membranes. The consequent water flow to modulate the intracellular ion concentrations, resulting in change of cell volume and osmotic potential. This is supported by the data that albumin and various cations exhibit linear correlations between their concentrations and cell IF tension, where albumin and K+ present positive slopes and Ca2+, Mg2+, and Na+ present negative slopes. At the same time, intracellular Cl− and Ca2+ levels are consistent with PN-OP effects. Therefore, abnormal levels of PNs and ions trigger proportional changes of transmembrane ion flow, depending on electrochemical-osmotic re-equilibrium.
Notably, extracellular PNs and different ionic compositions exhibit synergistic and antagonistic regulation of biological OP, achieving intero-sensory and stress-compensatory osmolarity control in the body. Consistently, the statistical analysis of clinical serum samples reveals hypoalbuminemia along with low Ca2+ plasma levels,22,39 possibly as a self-rescue measure to prevent osmotic disequilibrium in the body after the occurrence of edema. Likewise, clinical treatment of hyperpotassemia is performed with intravenous calcium injection, which is supported by the findings of the present study, ie, that increased extracellular Ca2+ levels are antagonized with hyperpotassemia to recover the isotonic state (Table S5). Changes in osmotic effects can affect neurohormones and nervous activity and are associated closely with water excretion and thirst. The extracellular hypertonic effect triggers vasopressin release and the resultant promotion of water permeability and reabsorption to regulate the osmotic gradients. In turn, water intake decreases OP and vasopressin release.40,41 Thirst and vasopressin act as osmotic responses in the intero-sensory control of body water homeostasis in the plasma, interstitial fluid, and intracellular fluid. Therefore, various ionic components are involved in regulating PN-adsorbing properties and consequent control of the activity of ion channels and biological OP, the mechanisms of which require further investigation.
The conventional method of OP measurement using an osmometer neglects the interplay of PNs and various ions in regulating transmembrane OP gradients. Thus, a reasonable system for real-time monitoring of osmotic effects in live cells is necessary. In the present study, a FRET-based GFAP tension probe was used for real-time measurement of IF tension, which is a sensitive and effective sensor for transmembrane OP gradients and a valid evaluation approach for live cells. In addition, multiple biochemical indexes of albumin and various ions undergo pathophysiologic changes, pathological symptoms do not necessarily appear due to synergy and antagonism-induced compensation and homeostasis in the human body (Figure 7). Therefore, indexes based on single plasma component are not sufficient criteria for blood disease diagnosis, especially those that are subject to transmembrane OP gradients in the body. Various components should be comprehensively considered if we are to develop new clinical diagnostic indexes or elucidate the mechanisms by which PN-OP regulates water flux in living systems.
Conclusion
In summary, biological OP is an underlying mechanism of PN-ion homeostasis in the body. The present study proposes that extracellular PN that adsorbs counterions is a crucial factor regulating biological OP. Particularly, Ca2+ and K+ exhibit the antagonistic and synergistic effects with extracellular PNs in regulating biological OP. Live cells can spontaneously regulate osmotic effects through changing membrane potential, controlling the ion channels and ion concentrations, switching between the hyper- and hypo-OP circumstances, regulating water flux, and alteration of cell volume and cytoskeleton structural tensions, eventually achieving new homeostasis. These factors suggest that asymptomatic albumin-ion disorders possibly develop into diseases, but are difficult to diagnose early. In our study, the linear relationships between the concentrations of various plasma components and transmembrane osmotic states could be comprehensively analyzed, further establishing a new appraisal index for osmotic equilibrium. Through analysis of clinical biochemical indexes and the incidence of edema, this new comprehensive index may be used to predict edema occurrence and further our understanding of the pathophysiologic variations of edema-related ions in the blood. In addition, we propose that certain PNs in the body, like albumin, inflammasome and amyloid β-protein,14,42 may also be involved in regulating biological OP, the mechanisms of which, and their relationship to underlying neurodegenerative diseases, should be further investigated.
Acknowledgments
This investigation was supported by the grants from National Natural Science Foundation of China (No. 82273908 and 82073826), Key Program of Natural Science Foundation of Jiangsu Province (No.19KJA320003) and Natural Science Foundation of Jiangsu Province (No. BK20200844).
Disclosure
Dr Jun Guo reports a patent 2021103033666 issued to Nanjing University of Chinese Medicine. The author reports no other conflicts of interest in this work.
References
1. Lopez MJ, Hall CA. Physiology, osmosis. StatPearls Publishing; 2022.
2. Bowler MG. The physics of osmotic pressure. Eur J Phys. 2017;38(5):055102. doi:10.1088/1361-6404/aa7fd3
3. Darwish A, Lui F. Physiology, colloid osmotic pressure. StatPearls Publishing; 2019.
4. Potts W, Parry G. Osmotic and ionic regulation in animals: International Series of Monographs on Pure and Applied Biology. Amsterdam: Elsevier; 2016.
5. Shorthouse D, Riedel A, Kerr E, et al. Exploring the role of stromal osmoregulation in cancer and disease using executable modelling. Nat Commun. 2018;9(1):3011.
6. Sterns RH. Treatment of severe hyponatremia. Clin J Am Soc Nephrol. 2018;13(4):641–649.
7. Tan AH, Lim SY, Ng RX. Osmotic demyelination syndrome with evolving movement disorders. JAMA Neurol. 2018;75(7):888–889.
8. Liu S, Yang H, Bian Z, et al. Regulation on mechanical properties of spherically cellular fruits under osmotic stress. J Mech Phys Solids. 2019;127:182–190.
9. Lu Y, Wang L, Chen D, et al. Determination of the concentration and the average number of gold atoms in a gold nanoparticle by osmotic pressure. Langmuir. 2012;28(25):9282–9287.
10. Mitchison TJ. Colloid osmotic parameterization and measurement of subcellular crowding. Mol Biol Cell. 2019;30(2):173–180. doi:10.1091/mbc.E18-09-0549
11. Guo Y, Wang Y, Ge Y, et al. Analysis of subcellular structural tension in axonal growth of neurons. Rev Neurosci. 2018;29(2):125–137. doi:10.1515/revneuro-2017-0047
12. Li C, Chen L, Wang Y, et al. Protein nanoparticle-related osmotic pressure modifies nonselective permeability of the blood-brain barrier by increasing membrane fluidity. Int J Nanomedicine. 2021;16:1663–1680. doi:10.2147/IJN.S291286
13. Zhang J, Wang Y, Zheng Z, et al. Intracellular ion and protein nanoparticle-induced osmotic pressure modify astrocyte swelling and brain edema in response to glutamate stimuli. Redox Biol. 2019;21:101112. doi:10.1016/j.redox.2019.101112
14. Zheng Z, Wang T, Chen J, et al. Inflammasome-induced osmotic pressure and the mechanical mechanisms underlying astrocytic swelling and membrane blebbing in pyroptosis. Front Immunol. 2021;12:688674. doi:10.3389/fimmu.2021.688674
15. Lang GE, Stewart PS, Vella D, et al. Is the Donnan effect sufficient to explain swelling in brain tissue slices? J R Soc Interface. 2014;11(96):20140123. doi:10.1098/rsif.2014.0123
16. Yu Z, Moomaw JF, Thyagarajapuram NR, et al. A mechanistic model to account for the Donnan and volume exclusion effects in ultrafiltration/diafiltration process of protein formulations. Biotechnol Prog. 2021;37(2):e3106. doi:10.1002/btpr.3106
17. Garces AH, Ang JE, Ameratunga M, et al. A study of 1088 consecutive cases of electrolyte abnormalities in oncology Phase I trials. Eur J Cancer. 2018;104:32–38. doi:10.1016/j.ejca.2018.08.019
18. Nguyen MK, Ornekian V, Kao L, et al. Defining the role of albumin infusion in cirrhosis-associated hyponatremia. Am J Physiol Gastrointest Liver Physiol. 2014;307(2):G229–G232. doi:10.1152/ajpgi.00424.2013
19. López-García JJ, Horno J, Grosse C. Influence of the dielectrophoretic force in mixed electrical double layers. J Colloid Interface Sci. 2013;405:336–343. doi:10.1016/j.jcis.2013.05.036
20. Bohinc K, Bossa GV, May S. Incorporation of ion and solvent structure into mean-field modeling of the electric double layer. Adv Colloid Interface Sci. 2017;249:220–233.
21. Qian Z, Wang Q, Qiu Z, et al. Protein nanoparticle-induced osmotic pressure gradients modify pulmonary edema through hyperpermeability in acute respiratory distress syndrome. J Nanobiotechnology. 2022;20(1):314.
22. Dalemo S, Bostrom KB, Hjerpe P. Plasma albumin and calcium concentrations, and long-term mortality in primary health care patients in Sweden. Scand J Prim Health Care. 2020;38(4):430–438.
23. Morotti A, Marini S, Lena UK, et al. Significance of admission hypoalbuminemia in acute intracerebral hemorrhage. J Neurol. 2017;264(5):905–911.
24. Conner BJ. Treating Hypoalbuminemia. Vet Clin North Am Small Anim Pract. 2017;47(2):451–459.
25. Fernandez J, Claria J, Amoros A, et al. Effects of albumin treatment on systemic and portal hemodynamics and systemic inflammation in patients with decompensated cirrhosis. Gastroenterology. 2019;157(1):149–162.
26. Kim S, McClave SA, Martindale RG, et al. Hypoalbuminemia and clinical outcomes: What is the mechanism behind the relationship? Am Surg. 2017;83(11):1220–1227.
27. Shalish W, Olivier F, Aly H, et al. Uses and misuses of albumin during resuscitation and in the neonatal intensive care unit. Semin Fetal Neonatal Med. 2017;22(5):328–335.
28. Taguchi K, Chuang VT, Maruyama T, et al. Pharmaceutical aspects of the recombinant human serum albumin dimer: structural characteristics, biological properties, and medical applications. J Pharm Sci. 2012;101(9):3033–3046.
29. Vincent JL, Russell JA, Jacob M, et al. Albumin administration in the acutely ill: what is new and where next? Crit Care. 2014;18(4):231.
30. Yang C, Zhang X, Guo Y, et al. Mechanical dynamics in live cells and fluorescence-based force/tension sensors. Biochim Biophys Acta. 2015;1853(8):1889–1904.
31. Chen T, Guo Y, Shan J, et al. Vector analysis of cytoskeletal structural tension and the mechanisms that underpin spectrin-related forces in pyroptosis. Antioxid Redox Signal. 2019;30(12):1503–1520.
32. Wang Y, Zhang X, Tian J, et al. Talin promotes integrin activation accompanied by generation of tension in talin and an increase in osmotic pressure in neurite outgrowth. FASEB J. 2019;33(5):6311–6326.
33. Zhang X, Li G, Guo Y, et al. Regulation of ezrin tension by S-nitrosylation mediates non-small cell lung cancer invasion and metastasis. Theranostics. 2019;9(9):2555–2571.
34. Chen X, He WT, Hu L, et al. Pyroptosis is driven by non-selective gasdermin-D pore and its morphology is different from MLKL channel-mediated necroptosis. Cell Res. 2016;26(9):1007–1020.
35. Rungta RL, Choi HB, Tyson JR, et al. The cellular mechanisms of neuronal swelling underlying cytotoxic edema. Cell. 2015;161(3):610–621.
36. Bolton GR, Boesch AW, Basha J, et al. Effect of protein and solution properties on the Donnan effect during the ultrafiltration of proteins. Biotechnol Prog. 2011;27(1):140–152.
37. Nguyen MK, Kurtz I. Determinants of plasma water sodium concentration as reflected in the Edelman equation: role of osmotic and Gibbs-Donnan equilibrium. Am J Physiol Renal Physiol. 2004;286(5):F828–F837.
38. Nguyen MK, Kurtz I. Quantitative interrelationship between Gibbs-Donnan equilibrium, osmolality of body fluid compartments, and plasma water sodium concentration. J Appl Physiol. 2006;100(4):1293–1300.
39. El-Kurdi B, Khatua B, Rood C, et al. Mortality from Coronavirus disease 2019 increases with unsaturated fat and may be reduced by early calcium and albumin supplementation. Gastroenterology. 2020;159(3):1015–1018.
40. Bankir L, Bichet DG, Morgenthaler NG. Vasopressin: physiology, assessment and osmosensation. J Intern Med. 2017;282(4):284–297.
41. Bichet DG. Vasopressin and the regulation of thirst. Ann Nutr Metab. 2018;72(Suppl 2):3–7.
42. Boeynaems S, Alberti S, Fawzi NL, et al. Protein phase separation: a new phase in cell biology. Trends Cell Biol. 2018;28(6):420–435.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.