Back to Journals » Breast Cancer: Targets and Therapy » Volume 18
AIM2 Lactylation Regulates Radiotherapy Sensitivity in Triple-Negative Breast Cancer
Authors Chen J, Zhao Z, Xing Y, Li X, Liu D, Liu L, Yang M, Zhou H, Wang J, Wu Q ![]()
Received 30 March 2026
Accepted for publication 26 June 2026
Published 9 July 2026 Volume 2026:18 613159
DOI https://doi.org/10.2147/BCTT.S613159
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Jianli Chen,1,2,* Ziming Zhao,1,* Yangyang Xing,3 Xiaorui Li,4 Di Liu,4 Lijia Liu,4 Mingyu Yang,4 Hua Zhou,2,5 Jue Wang,1 Qibiao Wu1,2
1Faculty of Chinese Medicine and State Key Laboratory of Mechanism and Quality of Chinese Medicine, Macau University of Science and Technology, Macau, People’s Republic of China; 2Zhuhai M.U.S.T. Science and Technology Research Institute, Guangdong-Macao In-Depth Cooperation Zone in Hengqin, Zhuhai, Guangdong, People’s Republic of China; 3The Macau Holy House of Mercy, Macau, People’s Republic of China; 4Radiotherapy Ward 3, The First Affiliated Hospital of Xinxiang Medical University, Xinxiang, Henan, People’s Republic of China; 5State Key Laboratory of Traditional Chinese Medicine Syndrome, Guangdong Provincial Hospital of Chinese Medicine, Guangdong Provincial Academy of Chinese Medical Sciences, The Second Affiliated Hospital of Guangzhou University of Chinese Medicine, Guangzhou, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qibiao Wu, Jue Wang, Faculty of Chinese Medicine and State Key Laboratory of Mechanism and Quality of Chinese Medicine, Macau University of Science and Technology, Avenida Wai Long, Taipa, Macau, 999078, People’s Republic of China, Email [email protected]; [email protected]
Background: Triple-negative breast cancer (TNBC) is an aggressive breast cancer subtype with limited targeted treatment options. Radiotherapy remains an important therapeutic approach for TNBC, but radioresistance markedly limits its efficacy. Absent in melanoma 2 (AIM2) has been increasingly implicated in tumor biology and innate immune signaling; however, its role in TNBC radioresistance and its regulation by the lactate-rich tumor microenvironment remain unclear.
Methods: Radiation-resistant TNBC cell models were established from BT-549 and MDA-MB-231 cells by repeated fractional irradiation. AIM2 knockdown and IRF3 overexpression were achieved by lentiviral transduction. Cell proliferation, colony formation, apoptosis, protein expression, AIM2 lactylation, subcellular localization, and AIM2–IRF3 interaction were evaluated using CCK-8 assay, colony formation assay, flow cytometry, RT-qPCR, Western blotting, co-immunoprecipitation, and immunofluorescence. Bioinformatics analyses were performed to assess the potential prognostic relevance of AIM2. In vitro experiments were performed with at least three independent biological replicates.
Results: Radiation treatment increased AIM2 expression in TNBC cells. AIM2 knockdown reduced colony formation and promoted apoptosis in radioresistant TNBC cells. High-lactate conditions decreased AIM2 expression, increased AIM2 lactylation, and promoted AIM2 nuclear translocation. IRF3 overexpression partially reversed the effects of AIM2 knockdown on colony formation and apoptosis, whereas lactate treatment weakened the interaction between AIM2 and IRF3. Bioinformatics analyses indicated that elevated AIM2 expression was associated with poorer prognosis.
Conclusion: These findings suggest that AIM2 may be involved in the regulation of TNBC radioresistance, potentially through its interaction with IRF3. High-lactate conditions may affect this regulatory axis by enhancing AIM2 lactylation. This study provides preliminary evidence for a potential mechanism linking lactate-mediated post-translational modification with radiosensitivity regulation in TNBC; however, further mechanistic, in vivo, and clinical validation is required.
Keywords: triple-negative breast cancer, radiotherapy resistance, AIM2, lactylation, IRF3, signal transduction
Introduction
Triple-negative breast cancer (TNBC) is an aggressive and heterogeneous subtype of breast cancer defined by the absence of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) expression.1 Because of the lack of these therapeutic receptors, TNBC has limited options for endocrine therapy or HER2-targeted therapy, and systemic treatment still relies heavily on chemotherapy, immunotherapy, antibody–drug conjugates, and local treatments such as surgery and radiotherapy.1,2 Radiotherapy is an important component of TNBC management, but intrinsic or acquired radioresistance can reduce treatment efficacy and contribute to recurrence or treatment failure.3
TNBC radioresistance is a multifactorial process involving enhanced DNA damage repair, apoptosis resistance, oxidative stress adaptation, metabolic reprogramming, epigenetic regulation, inflammatory signaling, and immune escape.4,5 Among these mechanisms, metabolic reprogramming has attracted increasing attention because TNBC cells frequently exhibit altered glucose metabolism, lactate accumulation, and metabolic crosstalk with the tumor microenvironment.6 Metabolic remodeling can also shape antitumor immunity by affecting immune-cell function, stromal interactions, and immune escape within the TNBC microenvironment.6 Therefore, understanding how metabolic changes regulate radiotherapy response may help clarify the biological basis of TNBC radioresistance.
Absent in melanoma 2 (AIM2) is a cytosolic DNA sensor that recognizes double-stranded DNA and participates in inflammasome activation, inflammatory cytokine maturation, and programmed cell death.7 AIM2 has context-dependent roles in cancer, acting as either a tumor suppressor or tumor-promoting factor depending on cancer type, cellular context, and tumor microenvironment.7,8 In breast cancer, earlier studies suggested that AIM2 may suppress cell proliferation and tumor growth, while more recent evidence indicates that AIM2-related inflammasome or pyroptosis pathways may also participate in breast cancer cell death regulation.9,10 However, the role of AIM2 in TNBC remains incompletely defined, especially in the context of radiotherapy response.
Radiotherapy induces DNA damage, oxidative stress, and inflammatory responses, all of which may be functionally connected with DNA-sensing pathways and inflammasome activation.5,11 A previous study showed that AIM2 can sense radiation-induced DNA damage and mediate inflammasome-associated cell death and tissue injury, supporting a potential relationship between AIM2 signaling and radiation response.11 In tumor cells, AIM2 has also been reported to promote irradiation resistance, migration, and PD-L1 expression in oral squamous cell carcinoma, suggesting that its role in radioresponse may be cancer-context dependent.12 Nevertheless, whether AIM2 contributes to TNBC radioresistance and how it is regulated by the metabolic tumor microenvironment remain unclear.
Lactate is a major metabolic byproduct of glycolysis and is increasingly recognized as an active signaling molecule in cancer rather than merely a metabolic waste product.13,14 Lactate can regulate tumor progression, immune suppression, and treatment resistance through metabolic and epigenetic mechanisms.13,14 Lactylation, a lactate-derived post-translational modification of lysine residues, links lactate accumulation with protein function and gene regulation.14,15 Recent studies have shown that lactylation can regulate cancer progression, immune remodeling, and therapy resistance, including in breast cancer and TNBC-related metabolic contexts.14,16 However, whether AIM2 undergoes lactylation in TNBC cells and whether lactate-mediated AIM2 lactylation participates in radiotherapy resistance remain unknown.
Interferon regulatory factor 3 (IRF3) is a key transcription factor downstream of innate DNA-sensing pathways and is closely linked to type I interferon signaling, inflammatory regulation, and cellular stress responses.17,18 Radiation-induced DNA damage can activate innate immune signaling and type I interferon responses, suggesting that IRF3-related signaling may be involved in the cellular response to radiotherapy.17 Given the potential functional connection between DNA sensing, AIM2-related signaling, lactate-induced post-translational modification, and IRF3-mediated transcriptional responses, the AIM2–IRF3 axis may represent a biologically relevant pathway in TNBC radioresistance. However, whether high-lactate conditions affect AIM2 function and its interaction with IRF3 has not been fully investigated.
In this study, we established radioresistant TNBC cell models using BT-549 and MDA-MB-231 cells and investigated the role of AIM2 in TNBC radiotherapy response. We further examined whether a lactate-rich environment affects AIM2 expression, lactylation, subcellular localization, and interaction with IRF3. Our study aimed to explore whether lactate-mediated AIM2 lactylation may be involved in the regulation of TNBC radiosensitivity and to provide preliminary evidence for a potential metabolic–innate immune regulatory mechanism in TNBC radioresistance.
Materials and Methods
Cell Culture
All cell lines were obtained from Wuhan Pulunsi Life Science and Technology Co., Ltd. and cultured according to the supplier’s instructions. BT-549 cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS), penicillin (100 units/mL), streptomycin (50 units/mL), and 10 µg/mL insulin. MDA-MB-231 cells were maintained in DMEM supplemented with 10% FBS and the same antibiotics. Both cell lines were maintained at 37°C in a humidified incubator with 5% CO2. To generate radioresistant cell lines, BT-549 and MDA-MB-231 cells in the logarithmic growth phase were exposed to repeated fractional irradiation at 2 Gy per fraction, five fractions per week, for six consecutive weeks, reaching a cumulative dose of 60 Gy. During the irradiation process, cells were passaged when they reached approximately 80% confluence. After completion of irradiation, the surviving cells were continuously cultured in complete medium and designated as radioresistant cells, namely BT-549-RR and MDA-MB-231-RR. For subsequent experiments, cells in the logarithmic growth phase were digested with 0.25% trypsin, neutralized with complete medium, and seeded into appropriate culture plates according to the requirements of each assay.
Lentiviral Transduction
In this study, lentiviral vectors were constructed for AIM2 knockdown (sh-AIM2) and IRF3 overexpression (OE-IRF3). The sh-AIM2 vector was constructed using the pLKO.1 backbone, while the OE-IRF3 vector was constructed using the pLV-Puro backbone. Both vectors were synthesized by Suzhou Jima Gene Co., Ltd. AIM2 gene sequences were obtained from NCBI, and specific shRNAs targeting AIM2 were designed, synthesized, and cloned into the pLKO.1 vector to generate the sh-AIM2 plasmid. A non-targeting shRNA sequence was used as the negative control (sh-NC), and the shRNA sequences are listed in Table 1. Following plasmid construction, a lentiviral packaging system was employed to package the vectors into lentiviruses, and virus stocks were harvested through precipitation and concentration steps. Enzymatic digestion and sequencing confirmation were performed by Shanghai Shengyuan Biological Engineering Co., Ltd.
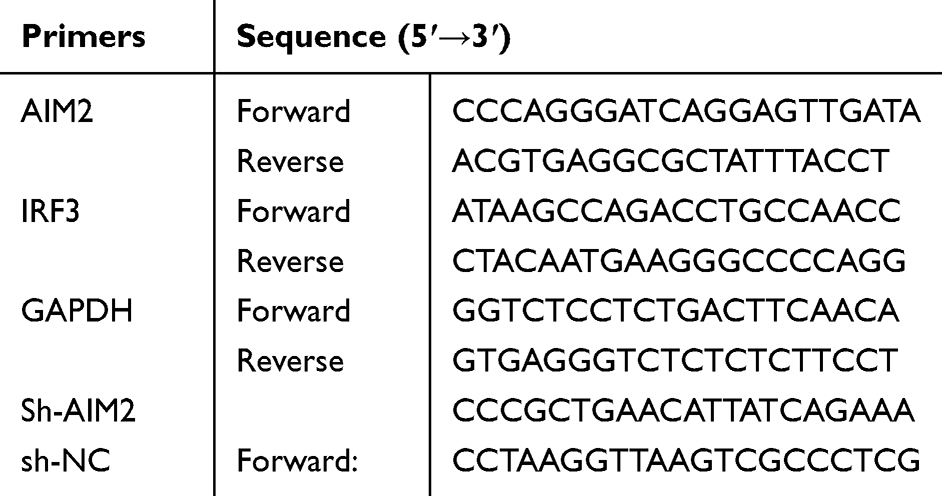 |
Table 1 Sequences of the Sh-RNAs |
Prior to lentiviral transduction, cells were seeded into six-well plates and cultured to approximately 50–70% confluence. Cells were then infected with sh-AIM2, sh-NC, OE-IRF3, or OE-NC lentivirus at an MOI of 100 in the presence of 5 μg/mL polybrene. After 24 h of infection, the medium was replaced with fresh complete medium. Stable cells were selected using 5 μg/mL puromycin for 7 days. The knockdown or overexpression efficiency was verified by RT-qPCR and Western blot analysis before subsequent experiments.
RT-qPCR
To evaluate the mRNA levels of AIM2 and IRF3, total RNA was extracted from cells in the logarithmic growth phase. Briefly, cells were lysed using Buffer RL, and RNA was purified using FastPure RNA Columns III, followed by washing with absolute ethanol and elution in RNase-free ddH2O. RNA concentration and purity were measured using a Nano 600 spectrophotometer. Total RNA was then reverse transcribed into cDNA using gDNA wiper mix and reverse transcriptase at 37°C for 15 min according to the manufacturer’s instructions. Real-time quantitative PCR (RT-qPCR) was subsequently performed. The amplification program was as follows: 95°C for 3 min, followed by 40 cycles of 95°C for 15s and 60°C for 30s. The relative mRNA expression levels of AIM2 and IRF3 were calculated using the 2−ΔΔCt method, with GAPDH as the internal control. The primer sequences used for RT-qPCR are listed in Table 1.
Western Blot
Equal amounts of protein were mixed with loading buffer, denatured, separated by SDS-PAGE using a 10% separation gel and a 5% stacking gel, and then transferred onto PVDF membranes at 300 mA for 90 min. After blocking with 5% non-fat milk or 5% BSA for 1 h at room temperature, the membranes were incubated with primary antibodies overnight at 4°C, followed by incubation with the corresponding secondary antibodies for 1 h at room temperature. Protein bands were visualized using an enhanced chemiluminescence detection system and captured with an imaging instrument. Protein expression levels were analyzed using ImageJ software, and relative expression levels were calculated using the corresponding internal control. Detailed antibody information is provided in Table 2.
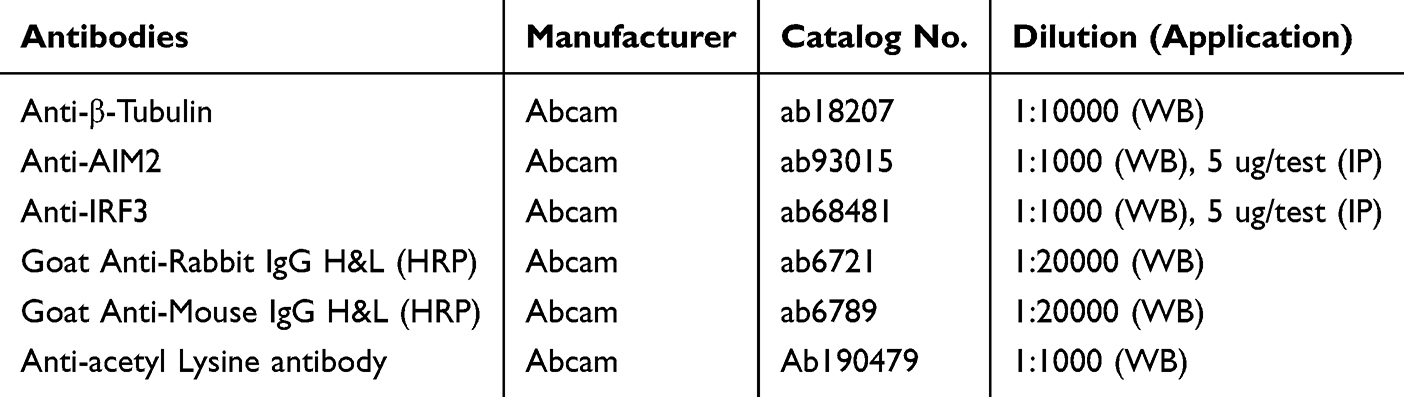 |
Table 2 Dilutions of the Antibodies Used for Western Blotting |
Colony Formation Assay
Cells in the logarithmic growth phase were seeded into six-well plates at a density of 1000 cells per well. After the indicated treatments, cells were cultured in an incubator for 14 days until visible colonies formed. After removing the culture medium, the cells were washed with PBS and fixed with 4% paraformaldehyde for 20 minutes. The paraformaldehyde was then removed, and the cells were stained with 0.2% crystal violet for 5 minutes. After staining, the cells were rinsed with water, air-dried, and photographed. The colony formation rate was calculated as the number of colonies divided by the number of seeded cells, multiplied by 100%.
CCK-8 Detection
Cells in the logarithmic growth phase were seeded into 96-well plates at a density of 2×105 cells per well and allowed to adhere overnight. After treatment with lactate at the concentrations indicated in the corresponding figure, 10 μL of CCK-8 reagent was added to each well, followed by incubation at 37°C for 1 h. The absorbance at 450 nm was measured using a microplate reader. Cell viability was calculated relative to the control group. All experiments were performed with three independent biological replicates.
Flow Cytometry
Cells in the logarithmic growth phase were seeded into six-well plates at a density of 1×106 cells per well and treated as indicated for 24 h. After treatment, the cells were harvested, washed with PBS, and centrifuged to remove the supernatant. The cell pellets were then resuspended in binding buffer, followed by staining with Annexin V and propidium iodide (PI) according to the manufacturer’s instructions. After incubation for 15 min at room temperature in the dark, the stained cells were analyzed by flow cytometry within 1 h.
Coimmunoprecipitation Assay
Preparation of cell lysate: When the degree of cell fusion reached 80%, drug treatment was added. After 24 hours, the cells were collected, washed with precooled PBS, lysed with RIPA containing PMSF, centrifuged at 4°C, and the supernatant was collected and frozen at −80°C. Before clearance of the cell lysate, protein A and protein G agarose were added to the cell lysate, which was incubated at 4°C and centrifuged, after which the supernatant was retained. For antigen‒antibody binding, the precleared lysates were incubated overnight at 4°C with anti-AIM2 antibody, anti-IRF3 antibody, or control IgG. Immunocomplex precipitation: Protein A/G beads were then added and incubated at 4°C for 2 h to precipitate the immune complexes, the mixture was centrifuged, the precipitate was washed, the mixture was resuspended in SDS‒PAGE electrophoresis buffer, and the CoIP samples were prepared after boiling in a water bath. For electrophoresis and membrane transfer, the samples were subjected to SDS-PAGE, transferred to a PVDF membrane, and washed with TBST. Development: After blocking, the membranes were incubated with the indicated primary antibodies overnight at 4°C, followed by incubation with the corresponding secondary antibodies, chemiluminescence detection was performed, and ImageJ software was used to analyze the relative protein expression levels. The specific antibody dilution ratios are shown in Table 2.
Immunofluorescence
Cells were seeded at a density of 1×106 per well in a 6-well plate. After the indicated treatments, the culture medium was removed, and the cells were gently washed with PBS. Cells were then fixed with 4% paraformaldehyde for 20 minutes, followed by another PBS wash. Membrane permeability was increased using 0.25% Triton X-100, after which the cells were washed again with PBS. To reduce nonspecific binding, cells were blocked with goat serum for 30 minutes. The blocking solution was then discarded, and a 1:200 dilution of AIM2 primary antibody was added. Samples were incubated overnight at 4°C. The next day, cells were washed with TBST, and a 1:200 dilution of the secondary antibody was added, followed by a 1-hour incubation at room temperature in the dark. After washing with TBST, DAPI staining was performed to label the nuclei. Samples were rinsed again with TBST and sealed with Fluoroshield mounting medium containing DAPI. Images were captured using a fluorescence microscope, and ImageJ software was used to analyze AIM2 fluorescence intensity to evaluate expression levels.
Bioinformatics Analysis
Breast cancer-related gene expression datasets, including GSE36295, GSE25066, and GSE246022, were obtained from the Gene Expression Omnibus (GEO) database. After data preprocessing and normalization, differentially expressed genes were screened between the indicated comparison groups. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed to explore the potential biological functions and signaling pathways associated with the differentially expressed genes.
Protein–protein interaction (PPI) analysis was performed to investigate potential interactions among candidate genes, and the interaction network was visualized using Cytoscape software. AIM2-related candidate genes and IRF3-associated interaction networks were further analyzed to explore potential regulatory relationships involved in TNBC radioresistance.
For prognostic analysis, clinical and gene expression data were integrated where available. Weighted gene co-expression network analysis (WGCNA) was used to identify modules associated with clinical features. LASSO-Cox regression analysis was then performed to screen prognosis-related genes and construct a prognostic model. Kaplan–Meier survival analysis was used to compare survival differences between high- and low-risk or high- and low-expression groups. Receiver operating characteristic (ROC) curve analysis was performed to evaluate the predictive performance of the prognostic model.
Data Analysis
Experimental data were analyzed and plotted using GraphPad Prism 9.4.0. Images were organized using Adobe Illustrator 2023. Data are presented as mean ± SD. Unless otherwise stated, all in vitro experiments were performed with at least three independent biological replicates. Comparisons between two groups were performed using Student’s t-test, while comparisons among multiple groups were performed using one-way analysis of variance (ANOVA) followed by appropriate post hoc tests. A P value < 0.05 was considered statistically significant.
Results
AIM2 Is Associated with Radiation Response in TNBC Cells
To investigate whether AIM2 expression is affected by radiation exposure, BT-549 and MDA-MB-231 cells were treated with 4 Gy radiation or left untreated as controls. RT-qPCR analysis showed that AIM2 mRNA levels were increased after 4 Gy radiation treatment in both cell lines (Figure 1A). Consistently, Western blot analysis showed increased AIM2 protein expression in BT-549 and MDA-MB-231 cells following 4 Gy radiation exposure (Figure 1B).
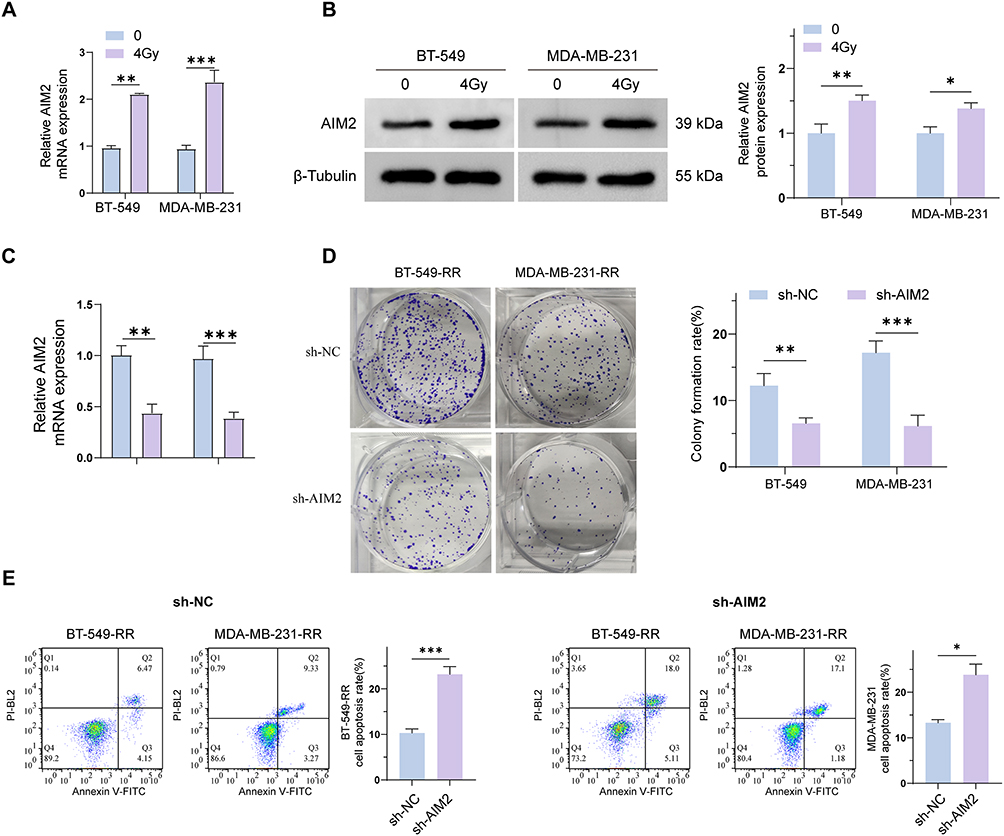 |
Figure 1 Differential expression of AIM2 in TNBC cells and radioresistant cells. (A) mRNA level of AIM2 in BT-549 and MDA-MB-231 cells after 4Gy radiation treatment. (B) Protein levels of AIM2 in BT-549 and MDA-MB-231 cells after 4 Gy irradiation. (C) The mRNA levels of AIM2 in BT-549-RR and MDA-MB-231-RR cells transfected with sh-AIM2 were compared with those in the sh-NC control group. (D) Colony formation assay of BT-549-RR and MDA-MB-231-RR cells transfected with sh-AIM2 compared with those in the sh-NC control group. (E) Apoptosis of BT-549-RR and MDA-MB-231-RR cells transfected with shAIM2 compared with those in the sh-NC control group. (*P < 0.05; **P < 0.01; ***P < 0.001). |
To further explore the potential role of AIM2 in radiation-resistant TNBC cells, AIM2 was knocked down in BT-549-RR and MDA-MB-231-RR cells using sh-AIM2, with sh-NC as the negative control. RT-qPCR confirmed that AIM2 expression was significantly reduced in the sh-AIM2 group compared with the sh-NC group (Figure 1C). Colony formation assays showed that AIM2 knockdown decreased the colony formation ability of radioresistant TNBC cells (Figure 1D). In addition, Annexin V/PI flow cytometry showed that AIM2 knockdown increased the apoptotic rate compared with the sh-NC group (Figure 1E).
Together, these results indicate that AIM2 expression is induced by radiation exposure in TNBC cells and that AIM2 knockdown suppresses colony formation while promoting apoptosis in radioresistant TNBC cells. These findings suggest that AIM2 may be involved in the regulation of radiation response in TNBC cells.
High-Lactate Conditions Affect AIM2 Expression, Lactylation, and Localization in Radioresistant TNBC Cells
We evaluated the effects of high-lactate conditions on AIM2 expression and cellular phenotypes in radioresistant TNBC cells. Treatment with increasing concentrations of exogenous lactate (LA) increased cell viability (Figure 2A), whereas the apoptotic rate was decreased (Figure 2B), suggesting that lactate may promote the survival of radioresistant TNBC cells.
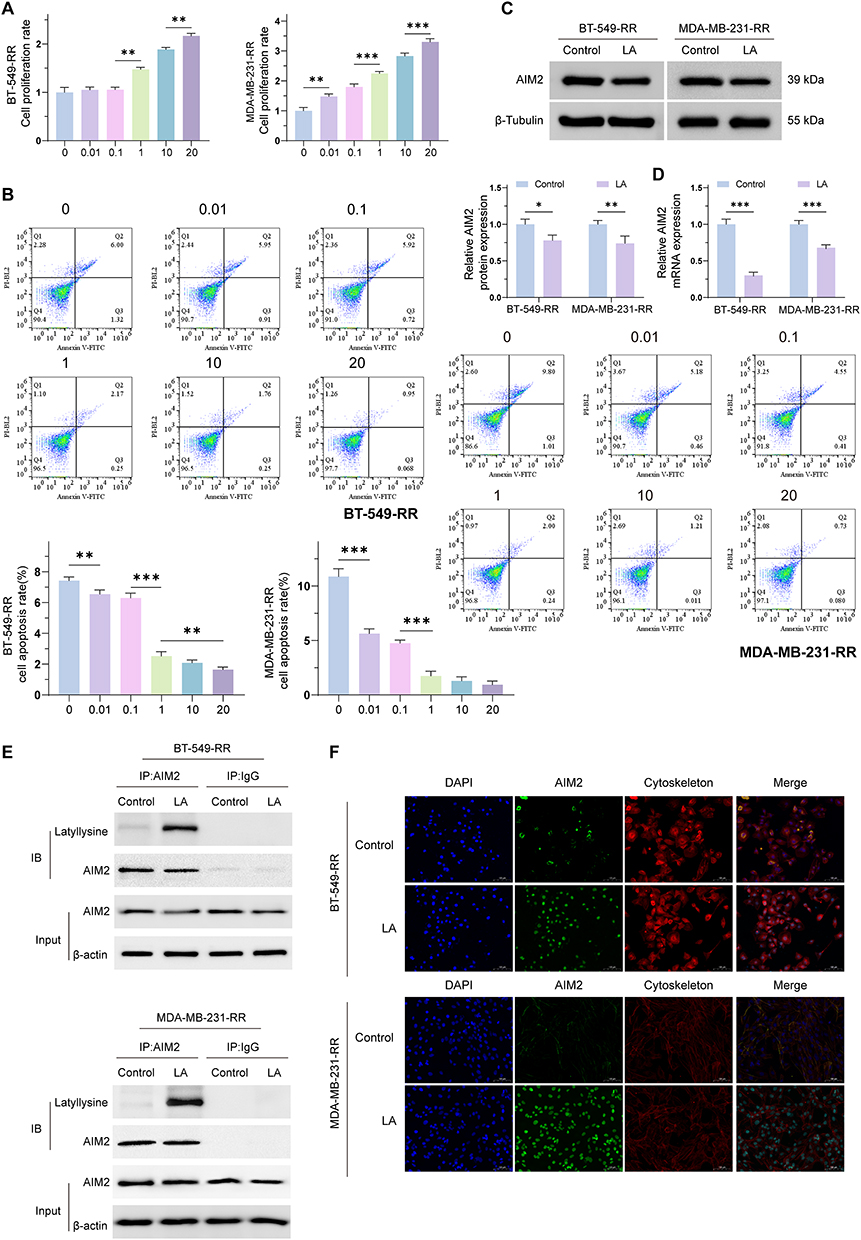 |
Figure 2 Investigating the effect of a high lactate environment on AIM2 expression in radiation-resistant cells. (A) Proliferation ability of BT-549-RR and MDA-MB-231-RR cells after treatment with different concentrations of exogenous lactate (LA). The results showed that with increasing LA concentration, the cell proliferation ability gradually increased. (B) Flow cytometry analysis revealed that under different concentrations of LA, the apoptosis rates of BT-549-RR and MDA-MB-231-RR cells gradually decreased. (C) Western blot detection of AIM2 protein levels in BT-549-RR and MDA-MB-231-RR cells after treatment with 20 mM LA for 24 h. (D) RT-qPCR detection of AIM2 mRNA levels in BT-549-RR and MDA-MB-231-RR cells after treatment with 20 mM LA for 24 h. (E) Coimmunoprecipitation (CoIP) detection revealed a significant increase in AIM2 lactylation levels in BT-549-RR and MDA-MB-231-RR cells treated with 20 mM LA. (F) Immunofluorescence localization revealed that AIM2 was localized in both the cytoplasm and nucleus under high lactate conditions and that LA treatment appeared to promote AIM2 nuclear translocation. (*P < 0.05; **P < 0.01; ***P < 0.001). |
To further investigate the effect of lactate on AIM2 expression, BT-549-RR and MDA-MB-231-RR cells were treated with 20 mM LA for 24 h. Western blot analysis showed that LA treatment decreased AIM2 protein expression compared with the untreated control group (Figure 2C). Consistently, RT-qPCR analysis showed reduced AIM2 mRNA expression after LA treatment (Figure 2D). These findings suggest that AIM2 expression is downregulated under high-lactate conditions.
Furthermore, co-immunoprecipitation (Co-IP) analysis showed that LA treatment increased the lactylation level of AIM2 (Figure 2E). Immunofluorescence analysis showed that AIM2 was distributed in both the cytoplasm and nucleus, and LA treatment appeared to promote AIM2 nuclear translocation (Figure 2F). Taken together, these results suggest that high-lactate conditions may affect AIM2 expression, lactylation, and subcellular localization in radioresistant TNBC cells.
Bioinformatics Analysis Identifies Potential AIM2- and IRF3-Related Effectors in TNBC Radioresistance
To explore potential AIM2-related effectors involved in TNBC radioresistance, bioinformatics analyses were performed. STRING analysis suggested a potential interaction between AIM2 and IRF3, indicating that IRF3 may be a candidate molecule associated with AIM2-related regulation (Figure 3A–C). In addition, four candidate proteins were predicted to have potential regulatory relationships with IRF3 (Figure 3D). These predicted interactions provide a basis for further experimental validation.
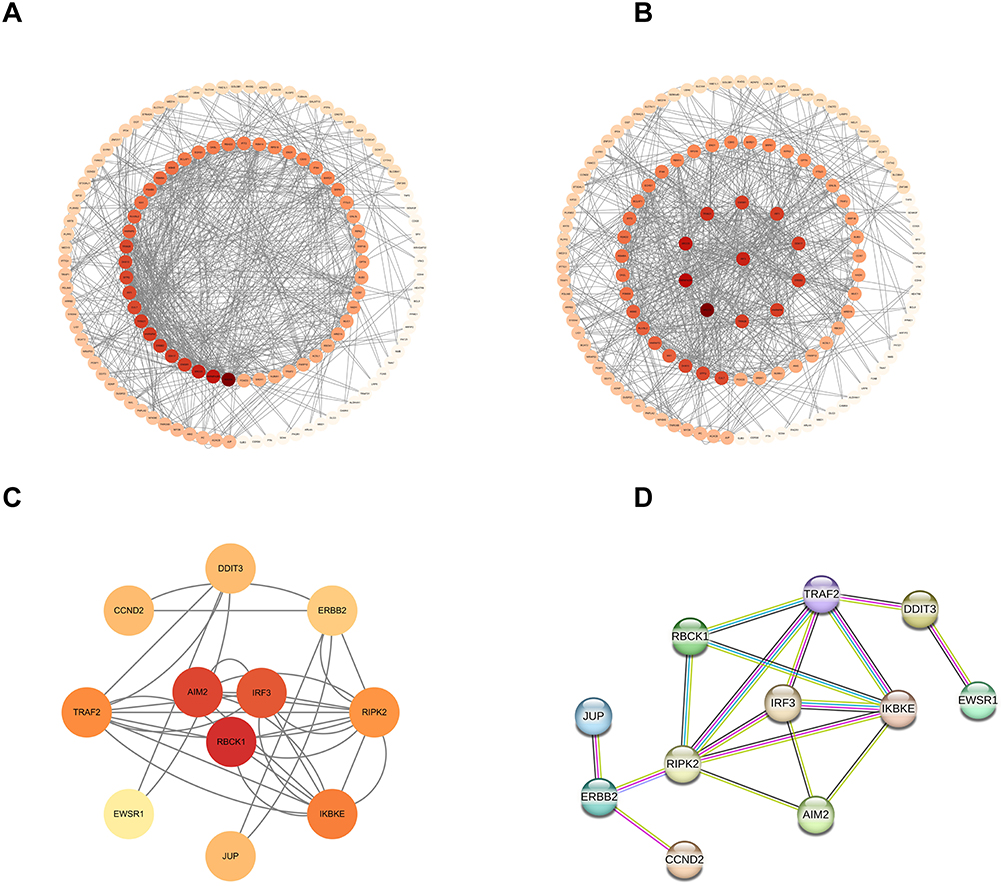 |
Figure 3 IRF3 and AIM2 potentially interact. (A–C) IRF3 and AIM2 potentially interact. (D) Four additional genes surrounding IRF3 that potentially interact with IRF3. |
LASSO-Cox regression analysis including five candidate genes showed that AIM2 was associated with the prognostic risk model (Figure 4A). As AIM2 expression increased, the patient risk score and number of deaths showed an increasing trend. ROC curve analysis showed that the 1-year, 3-year, and 5-year predictive curves were above the diagonal reference line, suggesting potential predictive performance of the model (Figure 4B–D). Kaplan–Meier survival analysis further indicated that patients with higher AIM2 expression had a lower survival probability than those with lower AIM2 expression (Figure 4E). These findings suggest that AIM2 may be associated with prognosis and may have potential relevance in TNBC radioresistance.
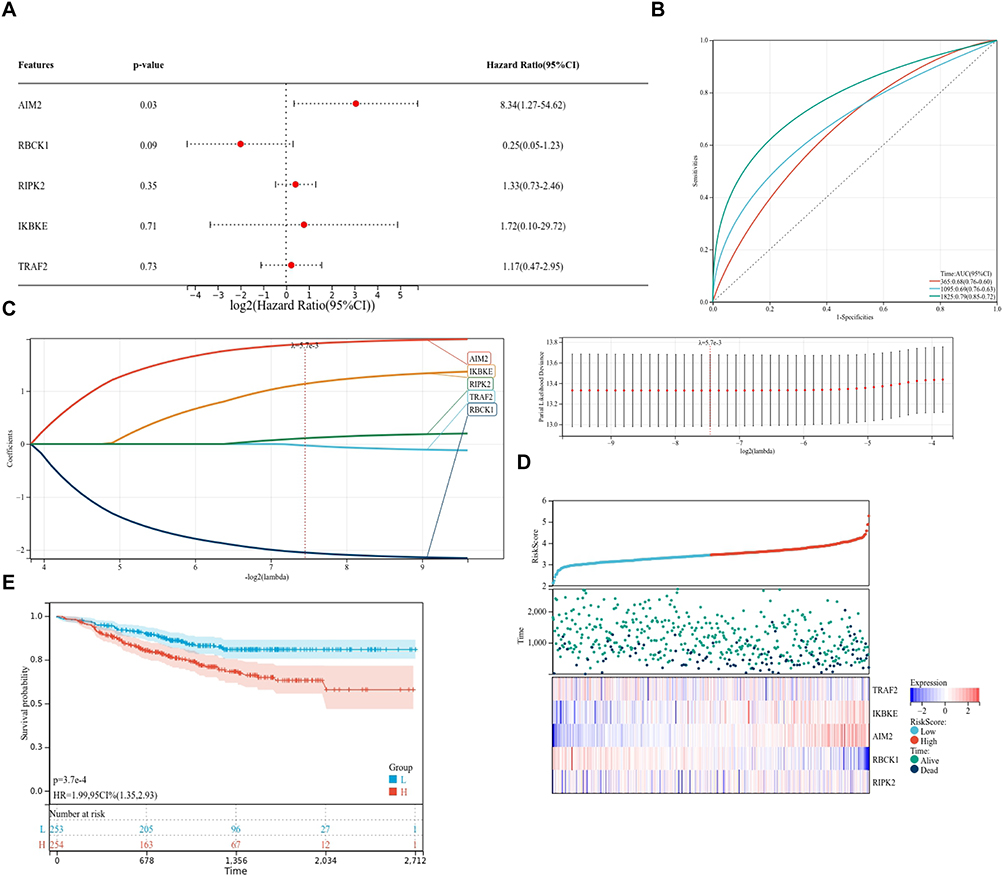 |
Figure 4 AIM2 has a strong association with TNBC. (A–D) AIM2 had the strongest positive association with risk factors in TNBC patients. (E) TNBC patients with high expression of AIM2 had lower survival curves. |
IRF3 is Associated with AIM2-Related Regulation of Radioresistant TNBC Cells
To investigate whether IRF3 is involved in AIM2-associated regulation of radioresistant TNBC cells, BT-549-RR and MDA-MB-231-RR cells were divided into five groups: sh-NC, sh-AIM2, OE-NC, OE-IRF3, and sh-AIM2+OE-IRF3. AIM2 mRNA expression was first evaluated by RT-qPCR. Compared with the sh-NC group, AIM2 mRNA expression was decreased in the sh-AIM2 group. Compared with the OE-NC group, AIM2 mRNA expression was increased in the OE-IRF3 group, whereas AIM2 mRNA expression in the sh-AIM2+OE-IRF3 group was lower than that in the OE-IRF3 group (Figure 5A).
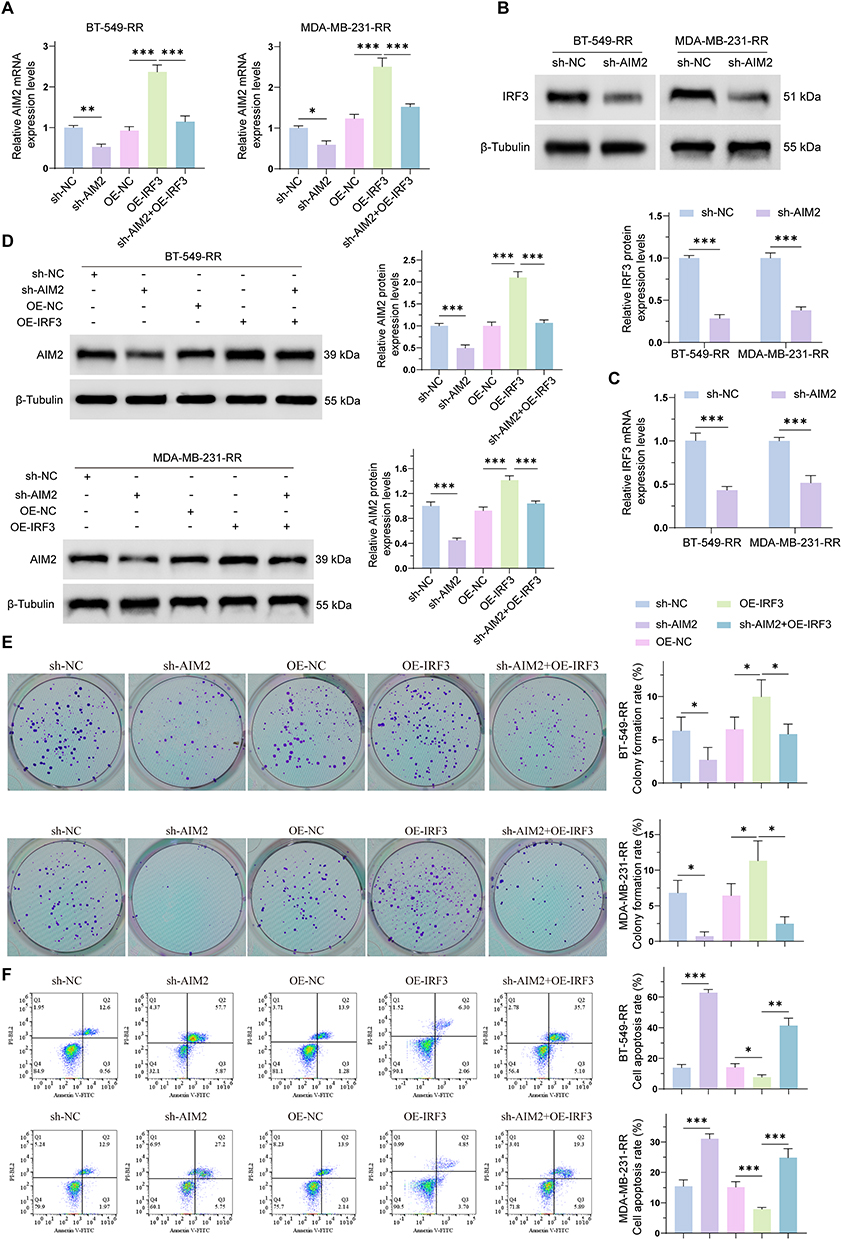 |
Figure 5 IRF3 is associated with AIM2-related regulation in radioresistant TNBC cells. (A) RT-qPCR analysis of AIM2 mRNA expression in BT-549-RR and MDA-MB-231-RR cells in the sh-NC, sh-AIM2, OE-NC, OE-IRF3, and sh-AIM2+OE-IRF3 groups. (B) Western blot analysis of IRF3 protein expression in BT-549-RR and MDA-MB-231-RR cells after AIM2 knockdown. (C) RT-qPCR analysis of IRF3 mRNA expression in BT-549-RR and MDA-MB-231-RR cells after AIM2 knockdown. (D) Western blot analysis of AIM2 protein expression in BT-549-RR and MDA-MB-231-RR cells in the sh-NC, sh-AIM2, OE-NC, OE-IRF3, and sh-AIM2+OE-IRF3 groups. (E) Colony formation assay of BT-549-RR and MDA-MB-231-RR cells in the indicated groups. (F) Flow cytometry analysis of apoptosis in BT-549-RR and MDA-MB-231-RR cells in the indicated groups. Data are presented as mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001. |
We next assessed whether AIM2 knockdown affected IRF3 expression. Western blot analysis showed that IRF3 protein expression was decreased in the sh-AIM2 group compared with the sh-NC group in both BT-549-RR and MDA-MB-231-RR cells (Figure 5B). Consistently, RT-qPCR analysis showed that IRF3 mRNA expression was also reduced after AIM2 knockdown (Figure 5C). In addition, Western blot analysis of AIM2 protein expression across the five groups showed that AIM2 was decreased in the sh-AIM2 group, increased in the OE-IRF3 group, and reduced in the sh-AIM2+OE-IRF3 group compared with the OE-IRF3 group (Figure 5D). These findings suggest a potential association between AIM2 and IRF3 expression in radioresistant TNBC cells.
Colony formation assays showed that AIM2 knockdown reduced the clonogenic ability of radioresistant TNBC cells compared with the sh-NC group. IRF3 overexpression increased colony formation compared with the OE-NC group, whereas the sh-AIM2+OE-IRF3 group showed reduced colony formation compared with the OE-IRF3 group (Figure 5E). Flow cytometry with Annexin V/PI staining further showed that AIM2 knockdown increased apoptosis, while IRF3 overexpression decreased apoptosis. The apoptotic rate in the sh-AIM2+OE-IRF3 group was higher than that in the OE-IRF3 group, indicating that AIM2 knockdown attenuated the anti-apoptotic effect associated with IRF3 overexpression (Figure 5F).
Together, these results suggest that IRF3 is associated with AIM2-related regulation of colony formation and apoptosis in radioresistant TNBC cells. However, further studies are needed to clarify whether this association is mediated directly through AIM2 lactylation and to define the downstream IRF3-related signaling pathways.
High-Lactate Conditions Weaken the Interaction Between AIM2 and IRF3
We further investigated whether high-lactate conditions affect AIM2 and IRF3 expression and their interaction in radioresistant TNBC cells. BT-549-RR and MDA-MB-231-RR cells were treated with 20 mM lactate (LA) for 24 h, with untreated cells used as controls.
RT-qPCR analysis showed that the mRNA expression levels of both AIM2 and IRF3 were reduced after LA treatment compared with the control group (Figure 6A). Western blot analysis further showed that AIM2 and IRF3 protein expression levels were also decreased under high-lactate conditions (Figure 6B). These results suggest that lactate treatment may downregulate AIM2 and IRF3 expression in radioresistant TNBC cells.
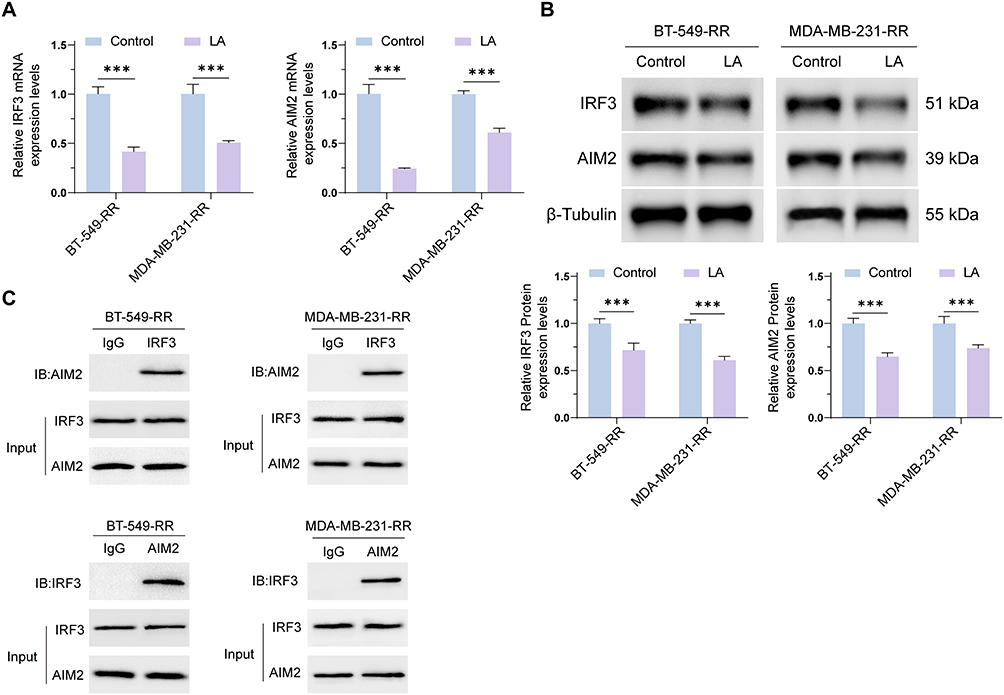 |
Figure 6 High-lactate conditions weaken the interaction between AIM2 and IRF3. (A) RT-qPCR analysis showed that AIM2 and IRF3 mRNA expression levels were decreased in BT-549-RR and MDA-MB-231-RR cells after treatment with 20 mM lactate (LA) for 24 h compared with the control group. (B) Western blot analysis showed that AIM2 and IRF3 protein expression levels were decreased in BT-549-RR and MDA-MB-231-RR cells after LA treatment compared with the control group. (C) Co-immunoprecipitation (Co-IP) analysis of the interaction between AIM2 and IRF3 in BT-549-RR and MDA-MB-231-RR cells under control and high-lactate conditions. Data are presented as mean ± SD. ***P < 0.001. |
Co-immunoprecipitation (Co-IP) analysis was then performed to evaluate the interaction between AIM2 and IRF3. AIM2 and IRF3 were not detected in the IgG control pull-down, whereas AIM2 or IRF3 antibody pull-down detected the corresponding interacting proteins, supporting a potential interaction between AIM2 and IRF3. Compared with the control group, LA treatment reduced the co-immunoprecipitation signal between AIM2 and IRF3, suggesting that high-lactate conditions may weaken the AIM2–IRF3 interaction (Figure 6C).
Taken together, these findings suggest that high-lactate conditions may reduce AIM2 and IRF3 expression and weaken their interaction in radioresistant TNBC cells.
Discussion
In this study, we investigated the potential role of AIM2 in TNBC radioresistance and explored whether a lactate-rich microenvironment affects AIM2-related regulation. Radioresistant BT-549-RR and MDA-MB-231-RR cells were established by repeated fractional irradiation using a cumulative dose of 60 Gy, and AIM2 expression was increased after radiation exposure. AIM2 knockdown reduced colony formation and increased apoptosis in radioresistant TNBC cells, suggesting that AIM2 may be involved in maintaining the survival capacity of TNBC cells under radiation stress. These findings are consistent with the broader understanding that tumor radioresistance is regulated by multiple mechanisms, including DNA damage repair, apoptosis resistance, oxidative stress adaptation, metabolic reprogramming, inflammatory signaling, and immune escape.19
AIM2 is a cytosolic DNA sensor and inflammasome-related molecule with context-dependent roles in cancer. In breast cancer, AIM2 has been reported to suppress cell proliferation and tumor growth, suggesting a potential tumor-suppressive function in certain breast cancer contexts.9 AIM2-related pathways have also been implicated in breast cancer cell death regulation, as dihydroartemisinin was reported to induce pyroptosis through the AIM2/caspase-3/DFNA5 axis.10 However, AIM2 may exert different effects depending on tumor type and treatment context. For example, AIM2 was reported to sense radiation-induced DNA damage and mediate inflammasome-associated cell death and tissue injury, supporting a connection between AIM2 signaling and radiation response.11 In oral squamous cell carcinoma, AIM2 was shown to promote irradiation resistance, migration, and PD-L1 expression through STAT1/NF-κB activation.12 Together with our findings, these studies suggest that AIM2 may have cancer- and context-specific functions in the regulation of radiotherapy response.
Metabolic reprogramming is increasingly recognized as an important contributor to therapy resistance and immune escape in TNBC. TNBC cells and the surrounding tumor microenvironment often exhibit altered glucose metabolism, hypoxia-related metabolic adaptation, and lactate accumulation, which can reshape tumor cell behavior and antitumor immunity.6 Lactate is no longer considered merely a metabolic waste product; rather, it can act as a signaling molecule and substrate for lactylation, a lactate-derived post-translational modification of lysine residues.15 Recent studies have highlighted lactylation as a regulatory mechanism linking tumor metabolism, gene expression, immune modulation, and therapy resistance.14 In the present study, high-lactate treatment decreased AIM2 expression but increased the lactylation level of AIM2 and promoted its nuclear translocation. These observations suggest that lactate-rich conditions may alter AIM2 function through post-translational modification, although the specific lactylation sites and their functional consequences remain to be further validated.
Importantly, the present study did not perform mass spectrometry-based identification of AIM2 lactylation sites or site-directed mutagenesis experiments. Therefore, our findings should be interpreted as evidence that AIM2 lactylation is increased under high-lactate conditions, rather than definitive proof that specific AIM2 lactylation sites directly drive TNBC radioresistance. Future studies should use mass spectrometry to identify AIM2 lactylation sites and perform site-directed mutagenesis to determine whether these residues are required for AIM2 localization, AIM2–IRF3 interaction, and radiation response. Such experiments will be necessary to clarify whether AIM2 lactylation is a causal regulator or a metabolic stress-associated modification in TNBC radioresistance.
IRF3 is a key transcription factor involved in innate immune signaling and type I interferon-related responses. Radiation-induced DNA damage can activate cytosolic DNA sensing pathways, including cGAS-STING signaling, which may lead to IRF3 activation and type I interferon production.20 In this study, bioinformatics analysis suggested a potential relationship between AIM2 and IRF3, and Co-IP experiments supported a possible interaction between AIM2 and IRF3 in radioresistant TNBC cells. Functionally, IRF3 overexpression partially reversed the effects of AIM2 knockdown on colony formation and apoptosis, suggesting that IRF3 may participate in AIM2-associated regulation of TNBC radioresistance. However, the downstream IRF3-related pathways were not fully explored in this study. In particular, interferon-related genes, inflammatory mediators, DNA damage repair factors, and apoptosis-related signaling molecules require further investigation to define the downstream network of the AIM2–IRF3 axis.
Our results also showed that lactate treatment weakened the interaction between AIM2 and IRF3. This finding suggests a possible link between lactate accumulation, AIM2 lactylation, and disruption of AIM2–IRF3 regulatory signaling. However, because the current evidence is based mainly on in vitro cell models, the causal relationship among lactate exposure, AIM2 lactylation, AIM2 nuclear translocation, AIM2–IRF3 binding, and radioresistance remains to be further established. Additional rescue experiments, AIM2 lactylation-deficient mutants, IRF3 downstream pathway analysis, and time-course studies will help determine whether lactate-induced AIM2 lactylation directly mediates the observed changes in radiosensitivity.
Bioinformatics analysis indicated that higher AIM2 expression was associated with poorer prognosis, suggesting a potential clinical relevance of AIM2 in breast cancer. However, this finding should be interpreted cautiously because it was derived from public datasets and was not validated using independent clinical cohorts or TNBC tissue samples in the present study. Further validation using clinical specimens, immunohistochemistry, paired radiosensitive and radioresistant tumor samples, and independent survival cohorts will be necessary to determine whether AIM2 expression or AIM2 lactylation has prognostic or predictive value in TNBC.
Several limitations should be acknowledged. First, this study was mainly based on established TNBC cell lines and in vitro radioresistant models, and the findings require validation in animal models and clinical tissue samples. Second, although AIM2 lactylation was increased under high-lactate conditions, the specific lactylation sites and functional domains of AIM2 were not identified. Third, the downstream signaling pathways of IRF3, including type I interferon signaling, inflammatory cytokine regulation, DNA damage repair, and apoptosis-related pathways, were not comprehensively investigated. Fourth, the clinical analysis was based on public transcriptomic datasets and lacked independent experimental validation. Therefore, the present findings should be regarded as preliminary evidence supporting a potential lactate–AIM2–IRF3 regulatory mechanism in TNBC radioresistance.
In summary, our study suggests that AIM2 may be involved in the regulation of TNBC radioresistance and that high-lactate conditions may influence AIM2 function by increasing AIM2 lactylation and altering its interaction with IRF3. These findings provide a potential link between metabolic remodeling and innate immune-related signaling in TNBC radiosensitivity regulation. However, further mechanistic, in vivo, and clinical studies are required before AIM2 lactylation can be considered a clinically actionable therapeutic target.
Conclusions
In this study, AIM2 expression was increased in TNBC cells following radiation exposure, and AIM2 knockdown reduced colony formation while promoting apoptosis in radioresistant TNBC cells. These findings suggest that AIM2 may be involved in maintaining the survival capacity of TNBC cells under radiation stress. Under high-lactate conditions that mimic the tumor microenvironment, AIM2 expression was decreased, whereas AIM2 lactylation and nuclear translocation were increased, suggesting that lactate accumulation may affect AIM2 function through post-translational modification. In addition, IRF3 appeared to participate in AIM2-associated regulation of TNBC radioresistance, while lactate exposure weakened the interaction between AIM2 and IRF3.
Overall, these findings provide preliminary evidence that lactate-induced AIM2 lactylation may be involved in the regulation of TNBC radiosensitivity, potentially through effects on AIM2 localization and AIM2–IRF3 interaction. However, the specific lactylation sites of AIM2, the downstream IRF3-related signaling pathways, and the in vivo and clinical relevance of this mechanism require further investigation. Therefore, AIM2 lactylation should be considered a potential direction for future mechanistic studies rather than an established clinically actionable therapeutic target at this stage.
Data Sharing Statement
The data generated in this study are available upon request from the corresponding author (Dr. Qibiao Wu).
Ethical Approval
This study involved only in vitro experiments using established human cell lines and analyses of publicly available, de-identified gene expression datasets obtained from the Gene Expression Omnibus (GEO) database. No human participants were recruited, no human specimens were collected from individuals, and no identifiable personal information was accessed. According to Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (National Health Commission of China, February 18, 2023), ethical review may be exempted for research that uses (1) legally obtained public data, (2) anonymized information data, and/or (4) established human cell lines, provided that such research does not involve sensitive personal information, human intervention, or activities prohibited under the regulation. The present study met these exemption criteria because it utilized publicly available anonymized GEO datasets and commercially available established human cell lines exclusively. Therefore, informed consent was not required. Ethical review and exemption confirmation were obtained from the local Institutional Review Board (IRB).
Acknowledgments
We would like to sincerely thank Yongxi Zhang, Shiyu Gu, Qi Su, Wenju Qin, Hongjie Wu, and Su Wang for their valuable assistance with data collection, experimental coordination, technical support, and helpful discussions during the course of this study. Their contributions greatly supported the successful completion of this work. We would like to express our sincere gratitude to the following individuals and institutions for their support and contributions to this work. First and foremost, we thank Science and Technology Development Fund, Macau SAR and the Chinese Medicine Guangdong Laboratory for the financial support that made this research possible. We are grateful to Professor Qibiao Wu for his valuable insights and guidance during the study. Their expertise greatly enhanced the quality of our research. We are thankful to Chinese Medicine Guangdong Laboratory for providing the necessary facilities and resources to conduct this research. Finally, we extend our deepest appreciation to our families for their unwavering support and patience throughout the course of this research.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by the Science and Technology Development Fund, Macau SAR (0048/2023/AFJ, 0164/2023/RIA3, 0121/2022/A3, 0127/2023/RIA2 and 0023/2024/ITP1).
Disclosure
The authors declare no competing interests in this work.
References
1. Leon-Ferre RA, Goetz MP. Advances in systemic therapies for triple negative breast cancer. BMJ. 2023;381:e71674. doi:10.1136/bmj-2022-071674
2. Lu L, Niu Z, Chao Z, Fu C, Chen K, Shi Y. Exploring the therapeutic potential of ADC combination for triple-negative breast cancer. Cellular and Molecular Life Sciences: CMLS. 2023;80(12):350. doi:10.1007/s00018-023-04946-x
3. Canha-Borges A, Nunes B, Quintas ST, et al. Emerging mechanisms of triple-negative breast cancer radioresistance: interplay between cancer cell mechanisms and the tumor immune microenvironment. Cancer Treat Rev. 2025;140:103026. doi:10.1016/j.ctrv.2025.103026
4. Wang Y, Liu Z, Lv Y, Long J, Lu Y, Huang P. Mechanisms of radioresistance and radiosensitization strategies for Triple Negative Breast Cancer. Transl Oncol. 2025;55:102351. doi:10.1016/j.tranon.2025.102351
5. Melia E, Parsons JL. DNA damage and repair dependencies of ionising radiation modalities. Bioscience Rep. 2023;43(10):BSR20222586. doi:10.1042/BSR20222586
6. Bao R, Qu H, Li B, Cheng K, Miao Y, Wang J. The role of metabolic reprogramming in immune escape of triple-negative breast cancer. Front Immunol. 2024;15:1424237. doi:10.3389/fimmu.2024.1424237
7. Sui L, Xi Y, Zheng S, Xiao Q, Liu Z. The Role of AIM2 in Cancer Development: inflammasomes and Beyond. J Cancer. 2025;16(1):157–16. doi:10.7150/jca.101473
8. Yu D, Zheng S, Sui L, Xi Y, He T, Liu Z. The role of AIM2 in inflammation and tumors. Front Immunol. 2024;15:1466440. doi:10.3389/fimmu.2024.1466440
9. Chen I, Ou-Yang F, Hung J, et al. AIM2 suppresses human breast cancer cell proliferation in vitro and mammary tumor growth in a mouse model. Mol Cancer Ther. 2006;5(1):1–7. doi:10.1158/1535-7163.MCT-05-0310
10. Li Y, Wang W, Li A, et al. Dihydroartemisinin induces pyroptosis by promoting the AIM2/caspase-3/DFNA5 axis in breast cancer cells. Chem-Biol Interact. 2021;340:109434. doi:10.1016/j.cbi.2021.109434
11. Hu B, Jin C, Li H, et al. The DNA-sensing AIM2 inflammasome controls radiation-induced cell death and tissue injury. Science. 2016;354(6313):765–768. doi:10.1126/science.aaf7532
12. Chiu H, Lee H, Lee H, et al. AIM2 promotes irradiation resistance, migration ability and PD-L1 expression through STAT1/NF-κB activation in oral squamous cell carcinoma. J Transl Med. 2024;22(1):13. doi:10.1186/s12967-023-04825-w
13. Chen L, Huang L, Gu Y, Cang W, Sun P, Xiang Y. Lactate-Lactylation Hands between Metabolic Reprogramming and Immunosuppression. International Journal of Molecular Sciences. 2022;23(19):11943. doi:10.3390/ijms231911943
14. Li H, Sun L, Gao P, Hu H. Lactylation in cancer: current understanding and challenges. Cancer Cell. 2024;42(11):1803–1807. doi:10.1016/j.ccell.2024.09.006
15. Zhang D, Tang Z, Huang H, et al. Metabolic regulation of gene expression by histone lactylation. Nature. 2019;574(7779):575–580. doi:10.1038/s41586-019-1678-1
16. Li J, Li Z, Zhang X, et al. Histone lactylation bridges metabolic reprogramming with chromatin-immune crosstalk in triple-negative breast cancer. Cancer Lett. 2026;639:218227. doi:10.1016/j.canlet.2025.218227
17. Zhang F, Manna S, Pop LM, Chen ZJ, Fu Y, Hannan R. Type I Interferon Response in Radiation-Induced Anti-Tumor Immunity. Semin Radiat Oncol. 2020;30(2):129–138. doi:10.1016/j.semradonc.2019.12.009
18. Wu Q, Leng X, Zhang Q, et al. IRF3 activates RB to authorize cGAS-STING-induced senescence and mitigate liver fibrosis. Sci Adv. 2024;10(9):eadj2102. doi:10.1126/sciadv.adj2102
19. Wu Y, Song Y, Wang R, Wang T. Molecular mechanisms of tumor resistance to radiotherapy. Mol Cancer. 2023;22(1):96. doi:10.1186/s12943-023-01801-2
20. Colangelo NW, Gerber NK, Vatner RE, Cooper BT. Harnessing the cGAS-STING pathway to potentiate radiation therapy: current approaches and future directions. Front Pharmacol. 2024;15:1383000. doi:10.3389/fphar.2024.1383000
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.