Back to Journals » Journal of Inflammation Research » Volume 18
Aging-Associated CD55⁺ Fibroblasts Promote Chronic Inflammation and ECM Dysregulation in Sarcopenia
Authors Xie X, Zhao J, Wu T, Yang Y, Zong Y, Li L, Gao Y, Jiang L ![]() , Cao Y
, Cao Y ![]() , Shen L
, Shen L
Received 25 May 2025
Accepted for publication 7 November 2025
Published 28 November 2025 Volume 2025:18 Pages 16729—16746
DOI https://doi.org/10.2147/JIR.S542613
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Xianfei Xie,1,2,* Jiawen Zhao,3,* Tong Wu,3,* Yi Yang,3,* Yuan Zong,3 Lei Li,4 Yiming Gao,3 Liting Jiang,3 Yuelong Cao,5 Linhui Shen6
1Shuguang Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai, People’s Republic of China; 2Department of Orthopedics, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, People’s Republic of China; 3Department of Stomatology, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine; College of Stomatology, Shanghai Jiao Tong University, Shanghai, People’s Republic of China; 4Department of Pathology, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, People’s Republic of China; 5Characteristic Diagnosis and Treatment Technology Research Institution, Shanghai Institute of Traditional Chinese Medicine, Shanghai, People’s Republic of China; 6Department of Geriatrics, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yuelong Cao, Characteristic Diagnosis and Treatment Technology Research Institution, Shanghai Institute of Traditional Chinese Medicine, Shanghai, People’s Republic of China, Email [email protected] Linhui Shen, Department of Geriatrics, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, People’s Republic of China, Email [email protected]
Background: Sarcopenia, defined as the progressive loss of muscle mass and function with aging, is a major contributor to frailty and disability in the elderly. Fibroblasts, traditionally considered passive extracellular matrix (ECM) producers, are increasingly recognized for their active roles in inflammation, fibrosis, and immune modulation. However, the heterogeneity and specific functions of fibroblast subtypes in sarcopenic muscle remain poorly understood.
Methods: We employed an integrative multi-omics and multi-modal approach combining single-nucleus RNA sequencing (snRNA-seq), untargeted metabolomics, and histological analyses to systematically characterize fibroblast populations in human skeletal muscle.
Results: Several distinct fibroblast subtypes were identified, among which a CD55⁺ fibroblast subpopulation (FB_4) emerged as a major contributor to the pro-inflammatory and fibrotic microenvironment of aged and sarcopenic muscle. FB_4 demonstrated activation of NF-κB signaling, glycolytic metabolism, and oxidative stress pathways. Histological validation confirmed increased expression of inflammatory and fibrotic markers such as TGF-β 1 and TLR4. Moreover, FB_4 activity correlated with senescence-associated secretory phenotype (SASP) expression and enhanced immune cell infiltration.
Conclusion: Our findings reveal CD55⁺ FB_4 as a critical driver of chronic inflammation and ECM remodeling in aging muscle. Targeting fibroblast-mediated inflammation and fibrosis may represent a promising therapeutic strategy for sarcopenia. The integration of transcriptomic, metabolic, and histological data provides new insights into fibroblast heterogeneity and offers a framework for future interventions in age-related muscle degeneration.
Keywords: senescence-associated secretory phenotype, extracellular matrix remodeling, skeletal muscle aging, fibroblast heterogeneity
Introduction
Sarcopenia is a geriatric syndrome characterized by accelerated skeletal muscle atrophy and functional decline, imposing growing biomedical and socioeconomic burdens on aging populations.1,2 Epidemiological studies report a wide prevalence range of 1% to 33% among community-dwelling elderly individuals.3 This degenerative condition not only diminishes muscular strength and aerobic capacity but also increases the risk of severe outcomes, such as falls, fractures, and mortality. The absence of effective therapeutic strategies contributes to significant psychological distress and economic strain on both affected individuals and healthcare systems, with current treatments providing only limited symptomatic relief.4,5 Although sarcopenia is strongly linked to aging, its underlying mechanisms are not yet fully understood. Emerging evidence suggests that non-myogenic cells within the muscle niche, including fibroblasts and endothelial cells, may actively sculpt the tissue microenvironment through paracrine signaling. However, the functional heterogeneity of these cells and their mechanistic contributions to sarcopenia progression remain elusive, hindering the development of targeted interventions.
Fibroblasts, once regarded merely as passive producers of the extracellular matrix (ECM), are now recognized as dynamic regulators of tissue homeostasis across various organs.6 In fibrotic diseases affecting the heart, liver, and lungs, specialized fibroblast subpopulations drive disease progression through cytokine secretion, metabolic reprogramming, and immune modulation.7 However, in skeletal muscle—a tissue that uniquely depends on mechanical and metabolic adaptability—the diversity and disease-specific roles of fibroblast subsets remain poorly understood.8 Traditionally, fibroblasts are known for their role in ECM production and remodeling, which provides structural support to tissues. Yet, during aging and disease, fibroblasts undergo activation, particularly transitioning into myofibroblasts, which contribute to pathological ECM remodeling, fibrosis, and the amplification of inflammation.9 The specific mechanisms underlying these processes remain to be fully elucidated.
Emerging evidence highlights chronic inflammation as a key regulator in the progression of sarcopenia, offering essential insights into the pathology of the condition. The accumulation of senescent cells in aging muscle creates a self-sustaining inflammatory niche through the release of senescence-associated secretory phenotype (SASP) factors. For instance, elevated IL-6 levels activate the ubiquitin-proteasome proteolytic pathway, fostering a pro-inflammatory environment.10,11 Similarly, TNF-α activates nuclear factor kappa B (NF-κB), driving a pro-inflammatory response, apoptosis, and muscle degradation.12,13 Increased interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α) levels contribute to a chronic inflammatory milieu that is closely associated with sarcopenia. Additionally, age-related lipid accumulation in skeletal muscle exacerbates mitochondrial oxidative stress, generating reactive oxygen species (ROS) that further promote cytokine hypersecretion and immune cell infiltration into muscle tissue, accelerating the decline in muscle function.14 While these findings emphasize the central role of inflammation in sarcopenia pathogenesis, the cellular entities orchestrating this inflammatory landscape remain poorly understood.14,15
In this study, we leverage both human muscle samples and a rat model of sarcopenia to comprehensively investigate age-related changes in skeletal muscle at the cellular and molecular levels. Employing a multi-omics and multi-modal data integration approach, we combined snRNA-seq, metabolomics, and histological analyses to identify key fibroblast subpopulations, including FB_4, and examine their contribution to the pro-inflammatory and fibrotic microenvironment in aged and sarcopenic muscle. By linking fibroblast activation with inflammation and metabolic reprogramming, our integrated approach provides valuable insights into novel therapeutic targets for mitigating the pathological processes driving sarcopenia.
Materials and Methods
Human Tissue Collecting
In accordance with the Declaration of Helsinki, all human tissue protocols were approved by the Ethics Committee of Ruijin Hospital, Shanghai Jiao Tong University School of Medicine (No. 2023–222). Informed consent was obtained from all participants. Vastus lateralis muscle samples were collected from young individuals (≤35 years), aged healthy individuals (≥60 years), and aged individuals with sarcopenia (≥60 years). Patients with sarcopenia were diagnosed according to the EWGSOP2 criteria.1,2 Patient characteristics and descriptive analyses are summarized in Table 1.
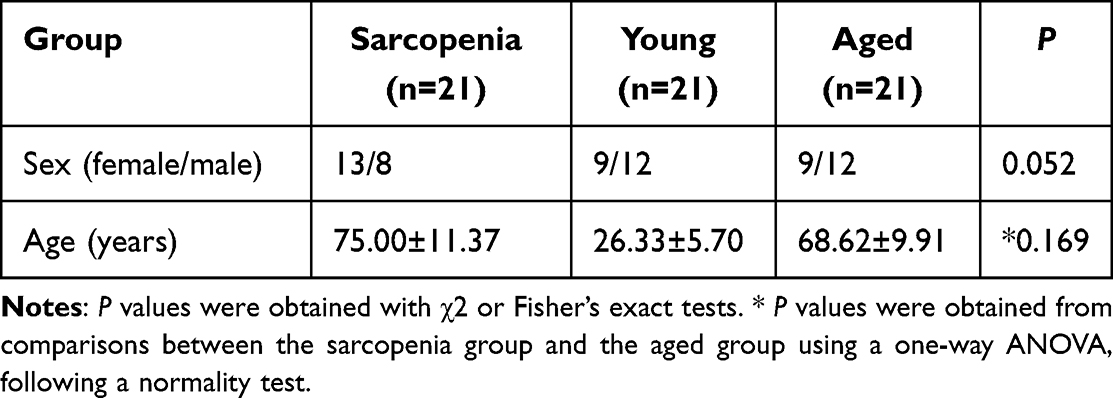 |
Table 1 Patient Characteristics and Descriptive Analysis |
Histological and Immunostaining
Fresh vastus lateralis muscle samples were frozen in liquid nitrogen-cooled isopentane and embedded in tragacanth gum. A portion of the fresh muscle was fixed in 4% neutral formaldehyde for 24 hours, dehydrated through a graded ethanol series, and embedded in paraffin. Sections were cut into 5-μm slices and stained with hematoxylin-eosin.
For immunohistochemical (IHC) staining, paraffin-embedded sections were dewaxed and rehydrated through graded ethanol incubations, followed by antigen retrieval and blocking. For frozen skeletal muscle sections, the dewaxing steps were omitted. Sections were incubated with primary antibodies against BCL6 (ZCL223, Leica, Germany), p21 (ab109520, Abcam, UK), CD44 (ZM-0051, ZSGB-BIO, China), TGF-β1 (GB11179-100; Servicebio, China), and TLR4 (GB12186-100, Servicebio, China) at 25 °C for 20 minutes. After washing with PBS, detection was carried out using the BOND Polymer Refine Detection Kit (DS9800, Leica Biosystems), which employs a polymer-based HRP secondary antibody system and does not rely on the ABC (Avidin–Biotin Complex) method, thereby avoiding issues with endogenous biotin. Sections were visualized with 3,3′-diaminobenzidine (DAB, K5007, Dako, Denmark) and counterstained with hematoxylin. Representative images were captured using the Leica Bond RX automated staining platform (Leica Biosystems).
For immunofluorescence (IF) staining, sections were deparaffinized and rehydrated before incubation with dihydroethidium (DHE, D7008, SIGMA, USA) at 37°C for 30 minutes. Nuclei were counterstained with DAPI. The antibodies used included α-SMA (1:500, GB12044; Servicebio, China) and collagen I (GB11022-3-100, Servicebio, China). Imaging was conducted using an upright fluorescence microscope (Nikon Eclipse C1, Tokyo).
Multi-Omics and Multi-Modal Data Integration Analysis
Integration of Human Datasets
Human data integration involved the use of two publicly available datasets. The single-nuclei cohort dataset GSE167186, retrieved from the public GEO database, comprised 19 young subjects, 29 aged subjects, and 24 patients with sarcopenia. Low-quality nuclei were excluded based on the following criteria: nFeature_RNA > 500 and nFeature_RNA < 500 and nCount_RNA < 10,000. This filtering process resulted in 101,862 single cells for further analysis.
Bulk RNA-seq datasets from sarcopenia cohorts were also obtained from the Gene Expression Omnibus (GEO) databases, including GSE16718616 and GSE111016.17 Data standardization was performed using the LIMMA package in R to generate a standardized matrix file. Raw data were normalized using FPKM, and normalized gene expression intensities were log2-transformed for subsequent analysis.
Single-Nucleus Transcriptomic Analysis and Cell Annotation
Normalization and clustering of cells were performed using the Seurat R package, with cluster visualization achieved via the uniform approximation and projection method (UMAP).18 Cluster annotation was carried out using the R package “SingleR”,19 with clusters representing the same cell type merged based on the expression of known marker genes. Gene set variation analysis (GSVA) was performed with the genes from Sn-RNA sequencing using the R Bioconductor package GSVA.20 Heatmaps to illustrate gene expression differences were generated using the R package “pheatmap”.
Intercellular Communication and Immune Infiltration Analysis
To assess cell–cell communication, particularly involving fibroblasts, the CellChat package21 was applied to GSE167186 to identify ligand–receptor interactions and secreted signaling pathways. Immune cell infiltration was quantified using ssGSEA and CIBERSORT,22,23 with scores compared between high- and low-SASP or FB_4 expression groups via the Wilcoxon test. Correlation and visualization were performed using Pearson coefficients, the Mantel test, and multiple R packages including “ggpubr”, “ESTIMATE”, “ggcor”, and “ComplexHeatmap”.
Gene Co-Expression Network and Functional Enrichment Analyses
The RNA-seq data from GSE167186 and GSE111016 were used for analysis. All samples across the three datasets were derived from the vastus lateralis muscle. WGCNA was performed using the R package WGCNA, including correlation, network analysis, and hierarchical clustering.24 Additionally, Proteomaps analysis was conducted on the sarcopenia group modules.
Functional enrichment analysis, including Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) pathway analysis, was conducted on differentially expressed genes using the clusterProfiler package for R25–27 and Metascape.28 Correlations were computed using Spearman’s rank correlation and visualized using the “ggplot2” R package, while scatter plots were generated using the “ggpubr” R package v0.4.0 with Pearson’s correlation. Protein-protein interaction (PPI) networks were assessed using the STRING database29 to explore pathways related to selected genes.
Animal Experiments
Animal Model of Sarcopenia
Naturally aged Sprague Dawley rats (>24 months old) were used as the animal model for sarcopenia.30 A total of 18 aged rats and 18 young rats (3 months old) were housed under standardized conditions at Ruijin Hospital, Shanghai Jiao Tong University School of Medicine. All animals used in this study were female. Vastus lateralis muscles were harvested for further analysis. All animal experiments and procedures were approved by the Beijing Vital River Laboratory Animal Technology Co., Ltd. (P2020100) and were conducted in accordance with the Guidelines for Ethical Review of Laboratory Animal Welfare (China, GB/T 35892–2018).
RNA Extractions From Rats’ Muscle Biopsies for Bulk Analysis
Rats’ muscle biopsies were homogenized in liquid nitrogen using a mortar and pestle. RNA was extracted from the resulting powdered samples with the RNeasy Fibrous Tissue Mini Kit (Qiagen) and processed using the QIAcube automatic processor (Qiagen). The integrity and concentration of the RNA were evaluated using the TapeStation 4200 (Agilent Technologies), with a cutoff for acceptable integrity set at an RNA integrity number (RIN) greater than 7. Batch-tag-seq libraries were generated from the RNA and sequenced on the HiSeq 4000 at the DNA Technologies Core, University of California, Davis.
Reads were aligned to the human genome using the STAR aligner, with GRCh38 as the reference genome. Counts were generated using the featureCounts function in the subread software. Genes with a total count of less than 10 were excluded from the analysis. Principal Component Analysis (PCA) of differentially expressed genes was performed using the DESeq2 package in R, with thresholds of absolute log Fold Change (logFC) > 1.5 and a False Discovery Rate (FDR) < 5%. Genes with a positive fold change were considered upregulated in the comparison group “B” when referring to “A vs B”.
Untargeted Metabolomics Analysis of Rats
For untargeted metabolomics, vastus lateralis muscle tissue from both sarcopenic and young rats was placed in a centrifuge tube. After metabolite extraction and quality control, the samples were analyzed using a SCIEX UPLC-Triple TOF 5600 system, equipped with an ACQUITY HSS T3 column (100 mm × 2.1 mm, 1.8 μm) for UPLC-MS/MS analysis. The conditions were set as follows: source temperature at 550°C, curtain gas (CUR) at 30 psi, and Ion Source Gas1 and Gas2 at 50 psi. Data were acquired using the Information Dependent Acquisition (IDA) mode. After filtering and normalizing, the data were subjected to further analysis, including PCA. Additionally, KEGG enrichment analysis was performed on the differential metabolites between the young and sarcopenic groups.
Statistical Analysis
Data were analyzed using GraphPad Prism software and SPSS software, with statistical significance determined by Student’s t-test. Each data was tested for normal distribution using the Shapiro–Wilks test. Results are presented as mean ± standard deviation (mean ± SD), with a significance level set at p<0.05.
Results
Identification of Age-Related Cellular Diversity in Skeletal Muscle
Single-nucleus RNA sequencing (snRNA-seq) was performed on muscle samples from six young and eleven aged donors (GSE167186) to identify age-associated cellular changes. Unsupervised clustering of the transcriptomic data led to the identification of 22 distinct cell clusters, which were annotated using canonical marker gene expression (Figure 1A–D). The two predominant cell populations identified were muscle fiber subtypes (eg, type I_1, I_2, II_1, II_2, II_3) and fibroblasts (FBs), which were further subdivided into LUM⁺ FAP and FBN1⁺ FAP subpopulations. Comparative analysis revealed three key findings: (1) LUM⁺ FAP cells were the dominant fibroblast subtype across all cohorts; (2) endothelial cell abundance was slightly increased in aged muscle; and (3) muscle fiber subtype distribution remained consistent between age groups (Figure 1E). Functional enrichment analysis indicated that FBs were primarily involved in ECM organization, while endothelial cells participated in angiogenesis and cell-cell adhesion pathways (Figure 1F and Supplementary Figure 1).
 |
Figure 1 (A) A brief flowchart of the single-nuclei sequencing process from skeletal muscle tissue in GSE167186. (B) tSNE plot of cells from young and aged muscle tissue, colored according to different cell clusters. (C and D) Feature plot (C) and violin plot (D) of specific markers used for muscle cell-type identification. Circle size represents the percentage of marker gene expression in cell types, while color shading indicates the average expression. (E) Bar plots showing the proportion of different cell types in young and aged groups. (F) Heatmap depicting gene expression in cell types. High to low expression is represented from red to grey. Enriched gene pathways for certain cell types are shown to the right of the heatmap. |
Fibroblasts Drive Pro-Inflammatory Gene Expression in Aged Muscle
A gene-level analysis was performed to further investigate the inflammatory environment in muscle aging. By intersecting highly variable genes from the 22 cell subpopulations with inflammation-related genes, 137 overlapping genes were identified (Figure 2A). KEGG and PPI analyses revealed enrichment in key immune and inflammatory pathways, including PI3K-Akt, JAK-STAT, and TNF signaling, as well as Th1/Th2 cell differentiation and T cell receptor signaling (Figure 2B and C). Further examination of senescence-associated secretory phenotype (SASP) genes identified fibroblasts, endothelial cells, and pericytes as major contributors to the inflammatory response, with FBs being the predominant source of cytokines and chemokines (Figure 2D and E). FB showed upregulation of IGFBP7, IL6ST, CXCL12, and EGFR, reinforcing its role in inflammation and immune dysregulation during muscle aging (Figure 2F and G).
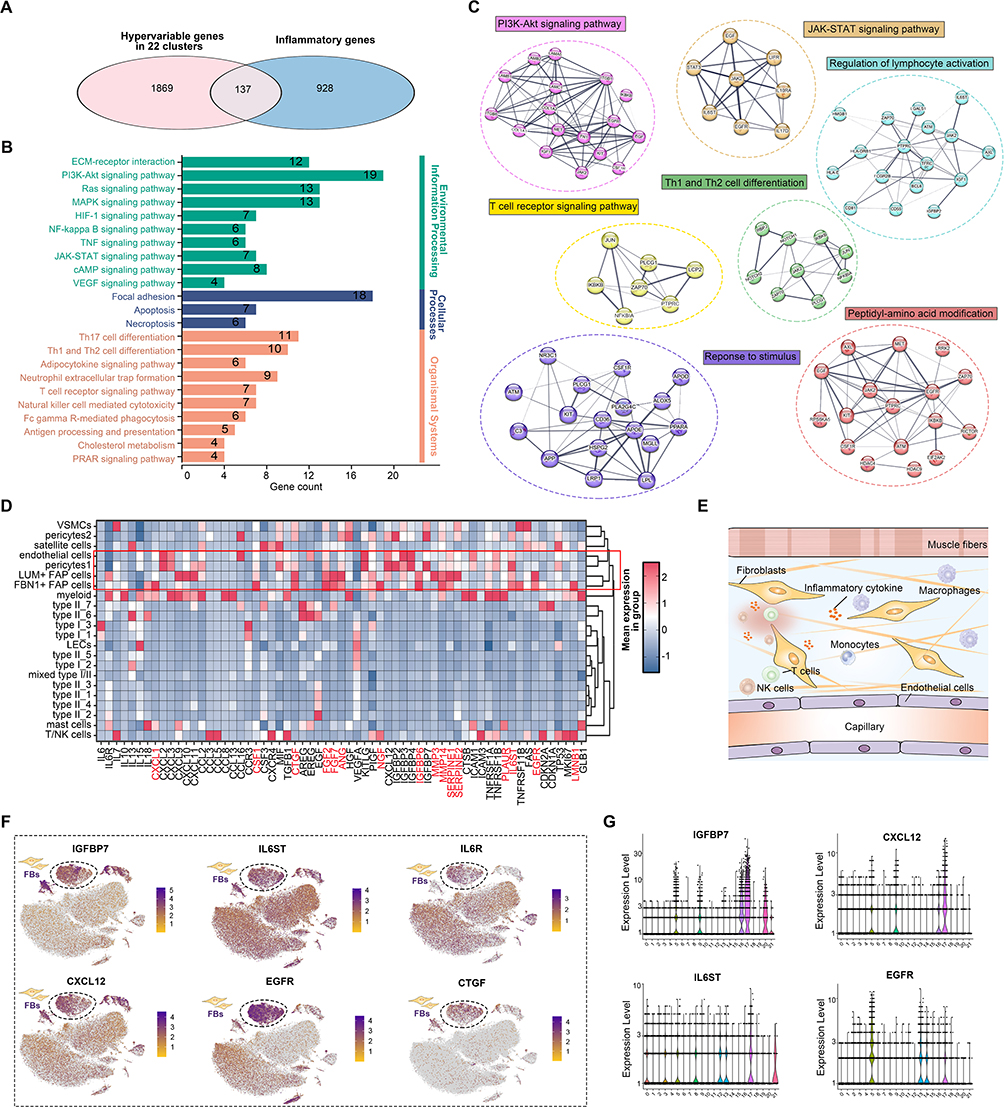 |
Figure 2 (A) Venn diagram displaying the overlap between hypervariable genes and inflammatory genes. (B) KEGG enrichment analysis of 137 overlapping genes. (C) PPI network of 137 overlapping genes. Circle nodes are color-coded by protein function, with line thickness representing the strength of data support. (D) Expression of SASP genes (rows) that are differentially expressed in cell types (columns). The heatmap shows the mean expression in each cell type. (E) Schematic depiction of fibroblasts within the inflammatory microenvironment. (F) Feature plots showing the high-expressed genes in fibroblasts. (G) Violin plots illustrating the high-expressed genes in fibroblasts. |
Fibroblasts Dominate Intercellular Signaling in Aged Muscle
Various cell types in skeletal muscle—including fibroblasts (FBs), satellite cells, and macrophages—were initially identified (Figure 3A). To analyze cell-cell communication within the muscle microenvironment, the CellChat tool was applied to model interactions between cell types. It was observed that FB-related communication lines were significantly thicker compared to those of other cell types, suggesting that FBs serve as dominant mediators of intercellular signaling in both young and aged groups (Figure 3B). This observation was reinforced by differential communication analysis between elderly and young cohorts (Figure 3C), in which the purple lines representing FB interactions—particularly from LUM⁺ FAP cells—exhibited the greatest interaction intensity in the aged group. Ligand-receptor pair analysis revealed increased communication probability between FBs and muscle fibers, particularly through LAMA2–DAG1 and HSPG2–DAG1 signaling (Figure 3D). Histological analysis confirmed muscle atrophy and enhanced fibrosis in the sarcopenic group, as indicated by an increased number of muscle fibers and higher expression levels of collagen I and α-SMA (Figure 3E and F).
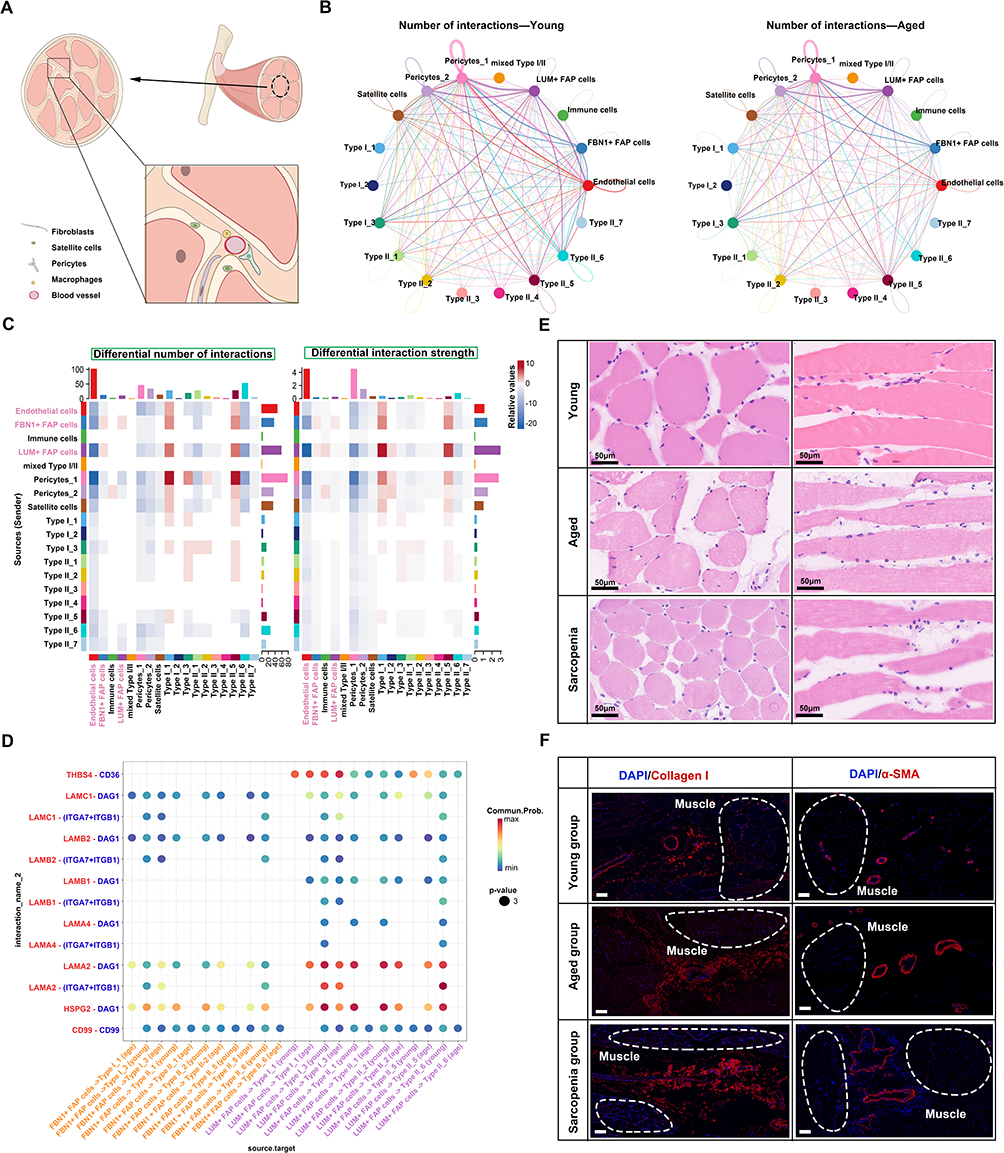 |
Figure 3 (A) Schematic view of cells related to skeletal muscle. (B) Circle plots visualizing the differential number of interactions in young and aged groups. (C) Heatmap showing the interaction numbers and strength of cell-cell communication in global samples. (D) Bubble heatmap displaying the strength of cell interactions for various ligand-receptor pairs. (E) HE staining of young, aged, and sarcopenia groups. Bar = 50 μm. (F) Immunofluorescence staining of DAPI, COL I, and α-SMA in the young, aged, and sarcopenia groups. Bar = 100 μm. |
Identification of Pro-Inflammatory FB_4 Subtype in Aged Muscle Fibroblasts
Fibroblasts (FBs) are key mediators of myogenesis and fibrosis via ECM production and remodeling, a process modulated by inflammatory cytokines. To dissect the heterogeneity and functions of FBs and endothelial cells (ECs), Cell-type annotation was refined through the use of canonical markers and correlation analysis, identifying ten distinct stromal and vascular subpopulations (Figure 4A–C). Hierarchical clustering revealed distinct transcriptional profiles among FBs, endothelial cells, and pericytes, with FB_4 clustering closely with FB_3 and FB_1, and a strong positive correlation with EC_3 (Figure 4D). Quantitative analysis indicated that FB subtypes were more abundant than endothelial cells, with the aged group contributing a higher proportion of FBs (Figure 4E). Functional enrichment analysis revealed that FB_4, which specifically expresses CD55, upregulates pathways related to arachidonic acid metabolism, suggesting an increase in the production of pro-inflammatory mediators such as prostaglandin E2 (PGE2). Furthermore, dysregulated autophagy and upregulated NF-κB signaling reinforce FB_4’s role in inflammation during muscle aging (Figure 4F–H). Additionally, the specific expression of EREG, IGFBP4, and MK167 in FB_4 (Figure 4I) reinforces its distinct role in age-related tissue remodeling.
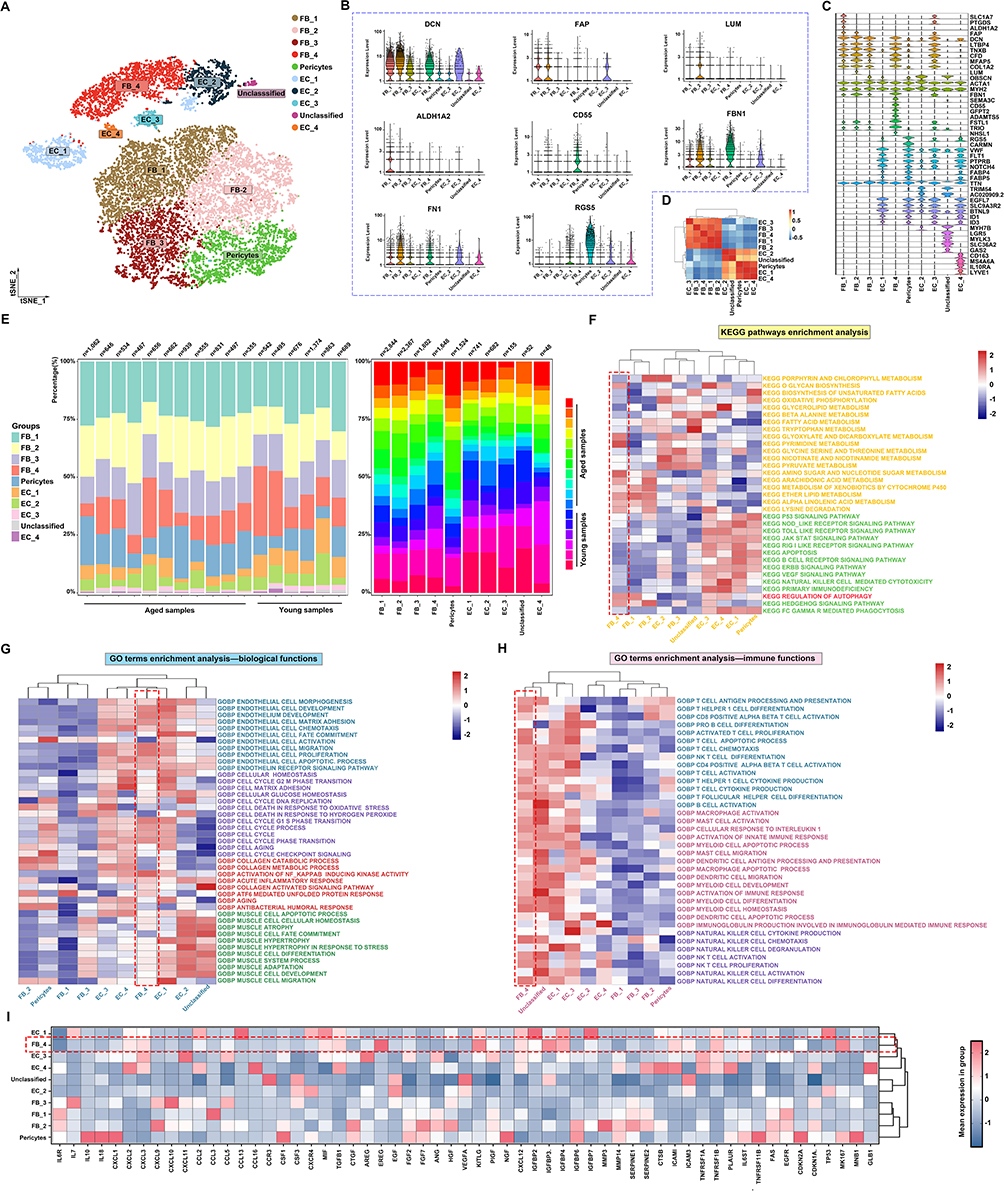 |
Figure 4 (A) tSNE plot of 10 clusters from fibroblasts and endothelial cells. (B and C) Violin plots of specific markers used for the identification of fibroblasts and endothelial cells. (D) Correlation analysis across different cell clusters. (E) Left bar plots show the proportion of different cell types in young and aged groups, while right bar plots display the distribution of young and aged samples across different cell types. (F–H) KEGG pathway enrichment analysis and GO term enrichment analysis. The red dashed box indicates the FB-4 subpopulation, the primary focus of this study. (I) Mean expression of inflammatory and aging-related genes across cell clusters. The red dashed box indicates the FB-4 subpopulation, the primary focus of this study. |
Enhanced FB_4-Mediated Communication in Aged Muscle Despite Overall Decline
Building on our identification of FB_4 as a key pro-inflammatory fibroblast subtype, intercellular communication in aging muscle was mapped using CellChat on snRNA-seq data. In comparison to the young group, the number of global interactions and interaction strength reduced sharply in aged group (Figure 5A). Then, we performed the cell chat in the groups and observed that the lines associated with fibroblasts are partly increased while the number of interactions globally decreased with advancing age (Figure 5B and C). To further investigate the changes in fibroblast subtypes in the aged group compared to the young group, we conducted cell communication analysis differential expressed genes between young and aged groups. The results demonstrated that the FB_4 received significantly increased signaling, indicating enhanced interactions in the aged group (Figure 5D and E). Specifically, collagen ligands (COL6A3, COL6A2, COL6A1) and their receptor CD44 exhibited increased interaction in aged muscle, with CD44 showing age-dependent upregulation (Figure 5F). Given the role of these collagen–CD44 interactions in maintaining muscle fiber integrity, this enhancement suggests a compensatory or pathogenic remodeling response. Pathway analysis revealed that FB_4 predominantly receives cues via the GAS and CXCL signaling axes (Figure 5G and H). Together, these findings position FB_4 as a central hub that dynamically reshapes specific signaling networks to drive the aberrant cell–cell communication and tissue remodeling characteristic of muscle aging.
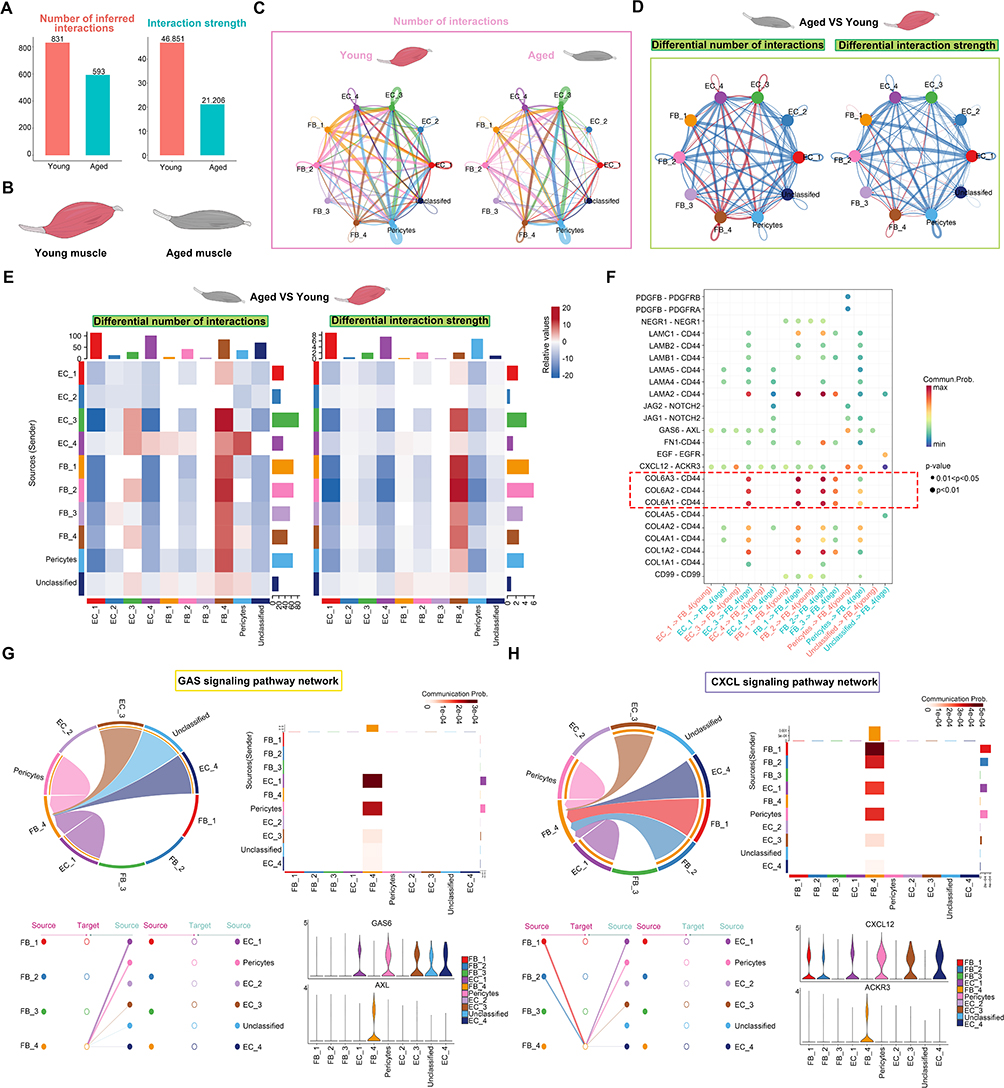 |
Figure 5 (A) Bar charts displaying the numbers and strength of interactions in young and aged groups. (B) Schematic representation of young and aged skeletal muscle. (C) Circle plots visualizing the differential number of interactions in young and aged samples. Node colors represent different cell types, with node size proportional to the number of interactions. The thickness of the strings represents the strength of communication. (D) Circle plots illustrating the differential number of interactions and interaction strength between Aged and Young groups. Node colors represent different cell types, with node size proportional to the number of interactions, and string thickness indicating communication strength. Brown indicates strengthened connections, while blue indicates weakened connections. (E) Heatmap depicting the interaction numbers and strength of cell-cell communication in global samples. Darker brown indicates a higher number and stronger intensity of connections, whereas darker blue represents fewer connections and weaker intensity. (F) Bubble heatmap displaying the strength of cell interactions for various ligand-receptor pairs. The size of the dots reflects the p-value, while the color of the dots represents communication probabilities. Empty spaces indicate a communication probability of zero. The red dashed box highlights ligand–receptor pairs with markedly stronger interaction strengths. (G and H) The chord plot, heatmap, hierarchical chart, and violin plot for GAS signaling pathway communication and CXCL signaling pathway. In the chord diagram, the width of each link indicates the strength of communication between cell clusters, while the width of each sector reflects the overall strength of communications to or from that cell group. |
FB_4 and SASP Genes Correlate with Inflammatory Pathways in Sarcopenic Muscle
To validate our findings, WGCNA was applied to RNA-seq data from young, aged, and sarcopenic muscle samples, identifying five trait-associated modules (p < 0.05; Figure 6A). The positively correlated green module was enriched in ECM–receptor interaction, TGFβ, PI3K–Akt, p53/senescence, FoxO, and autophagy pathways, consistent with pro-fibrotic and inflammatory remodeling (Figure 6B and C). In contrast, the pink module, negatively correlated with sarcopenia, was enriched in mitochondrial and metabolic processes, highlighting mitochondrial dysfunction in sarcopenia (Figure 6D and E). Proteomap analysis confirmed these findings (Figure 6F). Focusing on the green module’s PPI network further underscored inflammatory signaling, with hubs in PI3K–Akt, B cell activation, and lymphocyte proliferation pathways (Figure 6G). Sarcopenic muscle showed higher enrichment of FB_4, SASP factors, and PI3K–Akt-related genes compared to both young and aged controls (Figure 6H), further supporting FB_4’s role in driving inflammation and tissue remodeling (Figure 6I).
 |
Figure 6 (A) Gene dendrogram and co-expression modules between aged, sarcopenia, and young groups. The middle chart represents the correlation analysis between modules, while the bottom chart displays the module-trait relationship. (B and C) GO enrichment analysis for the pink and green modules. (D and E) KEGG analysis of the positive and negative correlation modules. (F) Proteomaps for the negative and positive correlation modules in the sarcopenia group. Each protein, encoded by a known gene ID, is depicted as a polygon, with functionally related proteins grouped together. (G) PPI network for the positive correlation modules, with genes color-coded by their functions. (H) Expression of genes related to FBs, SASP, and PI3K-AKT signaling pathway in the three groups. (I) Mantel test correlation between FB_4 and SASP-related genes. The heatmap illustrates the Pearson correlation between SASP-related factors. Line color represents p-values from the Mantel test, and line thickness indicates the magnitude of the Mantel correlation coefficient. Solid lines indicate positive correlations, while dashed lines represent negative correlations. |
Elevated P21, CD44, and BCL6 in Sarcopenic Muscle Connective Tissue
As summarized in Table 1, the three participant groups were well defined, with comparable sex distribution and expected differences in age, ensuring the validity of subsequent histological analyses. In these cohorts, P21, a key driver of cellular senescence in skeletal muscle, was markedly upregulated in the connective tissue of sarcopenic samples versus both aged and young controls, as shown by immunohistochemistry (Figure 7A). Similarly, CD44 immunoreactivity was significantly elevated in aged and sarcopenic muscles (Figure 7B), underscoring the importance of CD44-mediated signaling in fibroblast interactions. BCL6, linked to glycolytic muscle fibers and inflammatory regulation via PI3K–Akt signaling, was also enriched in sarcopenic tissue (Figure 7C), highlighting the interplay between chronic inflammation and senescence in the sarcopenic niche. To corroborate these histological findings at the gene-expression level, we scored bulk RNA-seq samples for the FB_4 signature using ssGSEA. Samples with high FB_4 scores showed significant upregulation of CD44 and several SASP-associated genes (Figure 7D), mirroring the immunohistochemical results and reinforcing FB_4’s role as a source of pro-inflammatory, senescence-associated signals in muscle aging. Consistent with this, markers of an inflammatory microenvironment, including TLR4 and TGFB1, were significantly elevated in aged and sarcopenic muscle compared with young controls. Correlation analyses further indicated that SASP-related genes, such as TGFB1, were positively associated with increased immune infiltration, while fibroblast marker FBN1 correlated strongly with monocytes and dendritic cells. These findings collectively highlight that elevated SASP expression in sarcopenic muscle is accompanied by enhanced immune activation and fibroblast–immune cell interactions (Supplementary Figure 2).
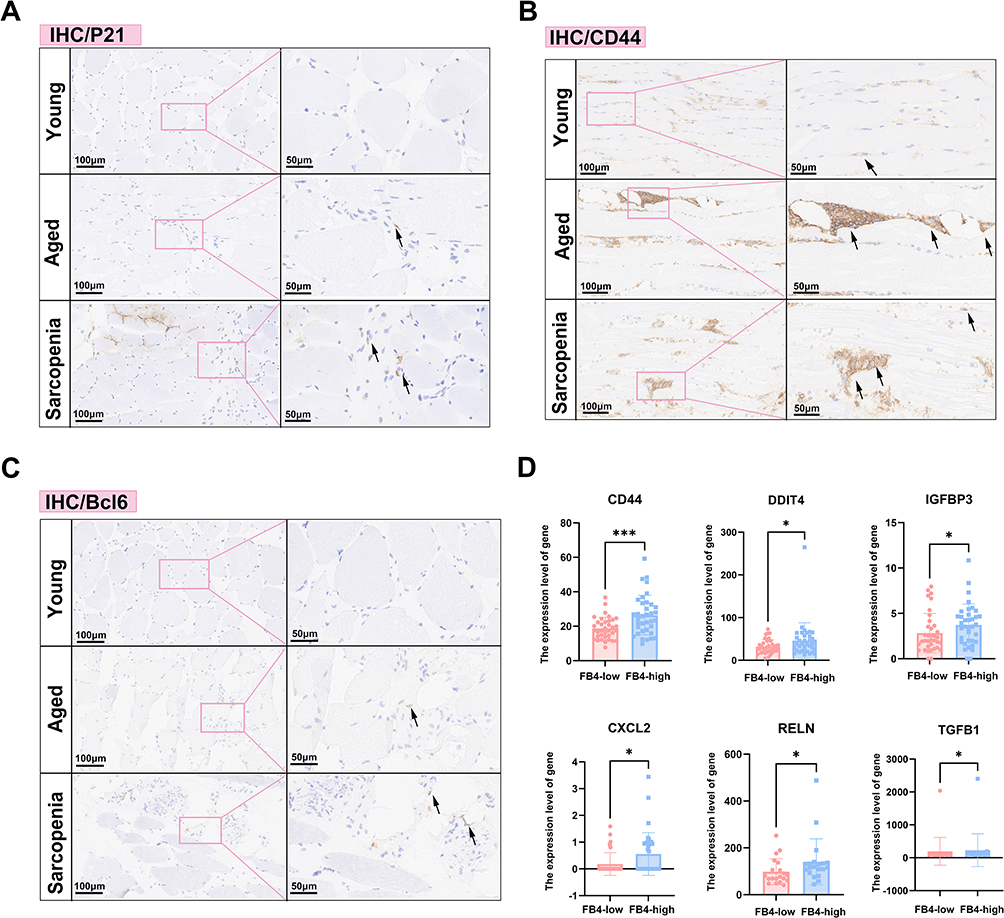 |
Figure 7 (A–C) Immunohistochemical staining for P21 (A), CD44 (B), and Bcl6 (C) in aged, young, and sarcopenia groups of clinical cohorts (black arrows indicate positive cells). (D) Summary graphs showing the expression levels of classical inflammatory genes in the FB4-low and FB4-high groups.*P < 0.05; ***P < 0.001. |
Glycolysis and Lysosomal Pathways Link to Inflammation in Sarcopenic Muscle
To further explore the metabolic changes in sarcopenic muscle, we performed untargeted metabolomics, bulk RNA-seq, and histological analyses on 12 sarcopenic and 12 young samples (Figure 8A). Histological analysis revealed smaller muscle fibers and increased fiber number in the sarcopenic group (Figure 8B). Metabolomics analysis identified distinct metabolic differences between sarcopenic and young muscle, with phospholipids being the major altered metabolites (Figure 8C). Differential abundance score analysis revealed enrichment in pathways related to glycolysis, gluconeogenesis, alanine, aspartate, and glutamate metabolism, as well as lysosomal function (Figure 8D). Metabolic stress is a key driver of aging and can trigger the senescence-associated secretory phenotype (SASP), with recent studies highlighting enhanced glycolysis in senescent cells. Correlation analysis showed that metabolites in glycolysis and FoxO signaling pathways were positively correlated with inflammation-related genes, while oxidative phosphorylation and AMPK signaling showed negative correlations (Figure 8E). Further analysis revealed that enzymes involved in glycolysis and lysosomal function were positively correlated with inflammatory genes, while ADP in oxidative phosphorylation showed a negative correlation (Figure 8F). These results suggest that metabolic enzymes play a key role in the pathophysiology of sarcopenia.
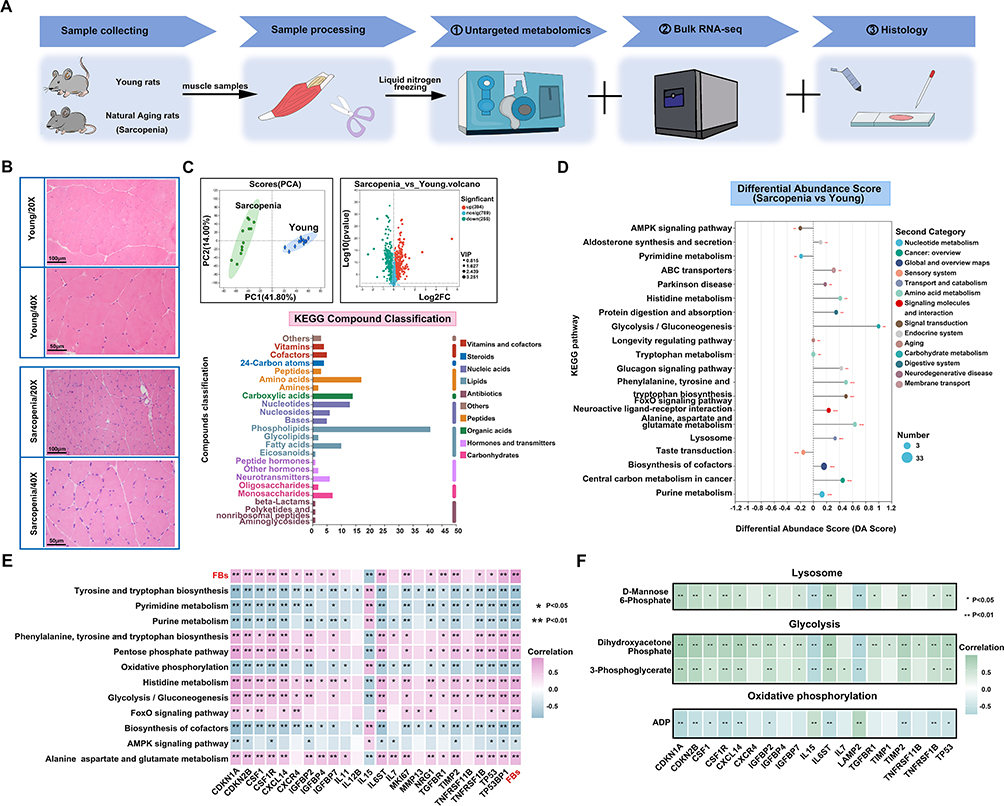 |
Figure 8 (A) Flowchart of rat tissue processing and analysis. (B) Hematoxylin-eosin staining of skeletal muscle in rats. (C) Untargeted metabolomics PCA analysis and volcano plot of sarcopenia and young rats. KEGG compound classification analysis of differentially expressed metabolites between aged and young samples. (D) Differential abundance score plot of enriched metabolic pathways. The size of the circles represents the number of differential metabolites. Positive DA-score (differential abundance score) indicates upregulation, while negative DA-score indicates downregulation. (E) Correlations between FBs and metabolic pathways, as well as SASP-related genes. (F) Correlations between enzymes in metabolic pathways and SASP-related genes. *P < 0.05; **P < 0.01; ***P < 0.001. |
Discussion
Chronic inflammation and cellular senescence are hallmarks of aging and age-related pathologies, manifesting as elevated inflammatory cytokines and accumulation of senescent cells.31,32 Senescent cells, primarily affecting FBs, endothelial cells, and macrophages, secrete pro-inflammatory mediators and amplify inflammatory signaling, thereby impairing muscle regenerative capacity.33–35 This study provides a comprehensive analysis of the cellular and molecular changes in skeletal muscle associated with aging and sarcopenia, focusing on the role of fibroblasts (FBs) in driving inflammation, fibrosis, and tissue remodeling. We identified key fibroblast subtypes, particularly FB_4, that contribute significantly to the pro-inflammatory and pro-fibrotic microenvironment characteristic of aging muscle. Our findings highlight the dynamic interplay between fibroblasts, immune cells, and the ECM, emphasizing the role of FBs in modulating muscle aging through inflammation- and metabolism-associated pathways.
The inflammatory microenvironment in aged skeletal muscle is primarily driven by pro-inflammatory cytokines (eg, IL-6, TNF-α, TGF-β) secreted by senescent cells, such as senescent fibroblasts or myocytes. These cytokines activate both immune cells and fibroblasts, leading to chronic inflammation, ECM remodeling, and fibrosis, while concurrently suppressing the regenerative capacity of muscle stem cells (satellite cells).36 Compounded by age-related immune dysfunction, mitochondrial impairment, and heightened oxidative stress, these factors collectively establish a self-perpetuating inflammatory cascade that exacerbates the progression of sarcopenia.37 One of the central findings of this study is the identification of distinct FB subtypes in aging muscle. Using snRNA-seq, we identified 22 cell populations, with LUM⁺ FAP and FBN1⁺ FAP fibroblasts as the dominant fibroblast subtypes in both young and aged muscle tissues. The relative abundance of these fibroblasts was found to increase with age, particularly in aged and sarcopenic muscle, indicating their central role in age-related changes in muscle structure. Furthermore, FBs exhibited characteristic upregulation of inflammatory mediators, including IGFBP7, IL6ST, and CXCL12, underscoring its central role in sustaining the pro-inflammatory milieu. FB_4 demonstrated robust activation of immune-related pathways such as NF-κB signaling, T cell activation, and macrophage chemotaxis, which demonstrated their role in amplifying inflammatory processes.
Fibroblasts (FBs) typically remain quiescent but secrete inflammatory cytokines in response to injury, which can drive their transformation into myofibroblasts. Myofibroblasts, also known as activated fibroblasts, play a key role as effector cells in various fibrotic diseases, including cardiac and hepatic fibrosis.38,39 These myofibroblasts are characterized by the overexpression of α-SMA and contribute to collagen deposition (mainly collagen type I), ECM remodeling, and cytokine secretion.38,40 The increased expression of collagen I and α-SMA observed in sarcopenic muscle supports the activation of fibroblasts and the subsequent increase in myofibroblast populations in sarcopenia. Fibroblasts are traditionally recognized for producing and remodeling the ECM to support muscle structure. However, our data show they also act as central hubs for intercellular signaling. We observed increased interactions between fibroblasts and muscle fibers through ligand–receptor pairs like LAMA2–DAG1 and HSPG2–DAG1 in aged muscle. Importantly, the positive correlation between FBs and pro-inflammatory immune cell populations, including M2c macrophages, and DCs, underscores the role of FBs in driving immune cell infiltration into the muscle tissue.
In addition to these immune-modulatory effects, the enrichment of CD55⁺ fibroblasts observed in our study highlights a potential link between fibroblasts and the complement system in sarcopenia. CD55, also known as decay-accelerating factor (DAF), is a membrane-bound complement regulatory protein that inhibits the formation of C3 and C5 convertases, thereby restraining excessive complement activation.41 While this mechanism normally protects tissues from complement-mediated damage, aberrant upregulation of CD55 in fibroblasts may reduce complement-driven clearance of senescent or damaged cells. In aging muscle, where CD55⁺ FB4 are enriched and simultaneously exhibit SASP-associated signaling, this dual role may paradoxically sustain chronic inflammation and ECM remodeling. Thus, the interplay between fibroblast activation and complement regulation represents an underappreciated axis in sarcopenia pathophysiology, and future studies should explore whether targeting complement–fibroblast interactions could provide therapeutic benefit.
SASP factors critically drive sarcopenia by sustaining chronic inflammation, disrupting metabolic homeostasis, and inhibiting muscle regeneration, and targeting SASP has emerged as a promising therapeutic avenue.42 Our study revealed an increase in senescence-associated secretory factors within muscle tissue from patients with sarcopenia, predominantly localized in the muscle interstitium. Senescent cells are characterized by permanent cell cycle arrest and the secretion of the SASP.43 Induced by various inflammatory signals, such as NF-κB, the SASP consists of a wide range of pro-inflammatory and immune-modulatory cytokines, including IL-6 and IL-8.43,44 Produced by senescent cells, the SASP recruits immune cells, clears senescent cells, and exacerbates the inflammatory response.43 In this study, samples were divided into two groups based on SASP expression levels to further explore the immune microenvironment in sarcopenia. The SASP-high group predominantly included patients with sarcopenia, with an increased presence of immune cells. The strong enrichment of SASP genes in the FB_4 fibroblast subset—and the tight correlation of TGFB1 expression with immune infiltration in SASP-high samples—underscores FB_4’s central role in orchestrating the pro-inflammatory microenvironment that drives muscle aging. Given that fibroblasts are the primary source of these inflammatory mediators, they represent attractive therapeutic targets for alleviating SASP-driven pathology in sarcopenia.
Recent studies have increasingly explored methods for integrating multimodal data, employing novel computational approaches to combine various data types.45 In our study, we integrated transcriptomic, metabolomic, and histological data to gain a comprehensive understanding of sarcopenia. After annotating cell clusters with specific markers like FAP and DCN, we identified FB_4 as a key subpopulation involved in ECM remodeling. Upregulation of FBN1 and enhanced collagen-CD44 interactions in FB_4 support its role in ECM remodeling in sarcopenia. Metabolomic analysis revealed FB_4’s involvement in inflammation-related pathways, including arachidonic acid metabolism and NF-κB signaling, and its role in immune activation. Histological analysis revealed increased abundance of TGF-β1 and TLR4 in the muscle interstitium of the aged and sarcopenic groups. TGF-β1, known to be upregulated in myocardial diseases, stimulates FB conversion into myofibroblasts by regulating their phenotype and function.46 TGF-β1 plays a critical role in FB activation and transdifferentiation, processes potentially mediated by Smad-related signaling pathways.47,48 Furthermore, TLR4, activated by damage-associated molecular patterns (DAMPs), is upregulated in muscle tissue, contributing to the fibrotic response.49,50 Accordingly, a pronounced fibrotic response was observed in both the aged and sarcopenic groups. Moreover, deep learning applied to digital histological scans may help identify subtle tissue features, enhancing early diagnosis.45 Future research should focus on developing integrated systems combining clinical, histological, and transcriptomic data.
In addition to inflammation, this study identifies metabolic changes in aging muscle that contribute to sarcopenia. Leveraging untargeted metabolomics—a powerful approach for global profiling of low-molecular-weight metabolites51—we identified pronounced glycolytic activation in sarcopenic rats, consistent with emerging evidence of age-dependent metabolic reprogramming marked by heightened glycolysis and mitochondrial dysfunction. Notably, lysosomal pathway activity exhibited a positive correlation with the senescence-associated secretory phenotype (SASP), whereas oxidative phosphorylation displayed an inverse relationship. This dichotomy aligns with recent reports implicating lysosomal cholesterol accumulation (driven by upregulated ABCA1 transporters) in SASP amplification during aging.52 Furthermore, senescence-associated oxidative stress amplifies reactive oxygen species (ROS) production, aggravating mitochondrial damage and suppressing oxidative phosphorylation.53 This indicates that metabolic stress is not merely a consequence but may actively contribute to the inflammatory environment by supplying energy or intermediates for SASP production. This link between metabolism and inflammation suggests that modulating metabolic pathways, such as reducing glycolysis, could help mitigate inflammation and slow muscle degeneration.
Our findings identify FB_4 as a potential therapeutic target to mitigate the inflammatory and fibrotic changes in aging muscle and sarcopenia. Targeting the inflammatory signaling pathways driven by FB_4 offers a novel strategy to alleviate chronic inflammation in aged muscle and potentially restore muscle function. For instance, inhibition of key pathways such as collagen–CD44 interactions or PI3K-Akt signaling could mitigate these inflammatory effects. Additionally, metabolic interventions aimed at restoring mitochondrial function or inhibiting glycolysis may provide further therapeutic benefits. Nevertheless, further functional studies will be required to directly establish causal links between CD55⁺ fibroblasts, complement signaling, and sarcopenia-associated tissue remodeling, which we plan to explore in subsequent research.
Conclusion
Our study provides a comprehensive overview of the cellular and molecular alterations in aging and sarcopenic muscle, identifying fibroblasts as central regulators of inflammation, fibrosis, and extracellular matrix (ECM) remodeling. Furthermore, we pinpointed FB4 as a key driver of these processes in muscle aging and sarcopenia. Through the integration of transcriptomic, metabolomic, and histological data, we demonstrated FB4’s critical role in ECM remodeling and immune activation, thereby sustaining a pro-inflammatory microenvironment. These findings suggest that targeting FB4-driven inflammatory pathways may offer novel therapeutic strategies to alleviate chronic inflammation and muscle degeneration. In addition, our study highlights the importance of metabolic reprogramming—including enhanced glycolysis and impaired mitochondrial function—providing complementary opportunities for metabolic interventions. However, further research is necessary to elucidate the precise molecular mechanisms through which FB_4 mediates inflammation, metabolism, and ECM remodeling, as well as its interactions with immune cells and muscle fibers, to fully harness its therapeutic potential.
Ethics Approval and Consent to Participate
In accordance with the Declaration of Helsinki, the studies involving human participants were reviewed and approved by the Ethics Committee of Ruijin Hospital, Shanghai Jiao Tong University School of Medicine. Informed consent was obtained from the patients.
Acknowledgments
We gratefully acknowledge the National Natural Science Foundation of China for their financial support and thank the contributors of the GEO database (http://www.ncbi.nlm.nih.gov/geo/) for making their datasets publicly available.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the National Natural Science Foundation of China (grant number: 82274553). The funder has no role in the conceptualization, design, data collection, analysis, decision to publish, or preparation of the manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Cruz-Jentoft AJ, Sayer AA. Sarcopenia. Lancet. 2019;393(10191):2636–2646. doi:10.1016/S0140-6736(19)31138-9
2. Cruz-Jentoft AJ, Bahat G, Bauer J, et al. Sarcopenia: revised European consensus on definition and diagnosis. Age Ageing. 2019;48(4):601. doi:10.1093/ageing/afz046
3. Cruz-Jentoft AJ, Landi F, Schneider SM, et al. Prevalence of and interventions for sarcopenia in ageing adults: a systematic review. Report of the International Sarcopenia Initiative (EWGSOP and IWGS). Age Ageing. 2014;43(6):748–759. doi:10.1093/ageing/afu115
4. Fielding RA, Vellas B, Evans WJ, et al. Sarcopenia: an undiagnosed condition in older adults. Current consensus definition: prevalence, etiology, and consequences. International working group on sarcopenia. J Am Med Dir Assoc. 2011;12(4):249–256. doi:10.1016/j.jamda.2011.01.003
5. Sarcopenia definition, diagnosis and treatment: consensus is growing - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/36273495/.
6. DeLeon-Pennell KY, Barker TH, Lindsey ML. Fibroblasts: the arbiters of extracellular matrix remodeling. Matrix Biol. 2020;91–92:1–7. doi:10.1016/j.matbio.2020.05.006
7. Liu X, Dai K, Zhang X, et al. Multiple fibroblast subtypes contribute to matrix deposition in pulmonary fibrosis. Am J Respir Cell Mol Biol. 2023;69(1):45–56. doi:10.1165/rcmb.2022-0292OC
8. Chapman MA, Meza R, Lieber RL. Skeletal muscle fibroblasts in health and disease. Differentiation. 2016;92(3):108–115. doi:10.1016/j.diff.2016.05.007
9. Younesi FS, Miller AE, Barker TH, Rossi FMV, Hinz B. Fibroblast and myofibroblast activation in normal tissue repair and fibrosis. Nat Rev Mol Cell Biol. 2024;25(8):617–638. doi:10.1038/s41580-024-00716-0
10. Jg R, Jd S, Gs L. Cellular and molecular mechanisms underlying age-related skeletal muscle wasting and weakness. Biogerontology. 2008;9(4). doi:10.1007/s10522-008-9131-0
11. Larsson L, Degens H, Li M, et al. Sarcopenia: aging-related loss of muscle mass and function. Physiol Rev. 2019;99(1):427–511. doi:10.1152/physrev.00061.2017
12. Kandarian SC, Jackman RW. Intracellular signaling during skeletal muscle atrophy. Muscle Nerve. 2006;33(2):155–165. doi:10.1002/mus.20442
13. Sousa-Victor P, García-Prat L, Muñoz-Cánoves P. Control of satellite cell function in muscle regeneration and its disruption in ageing. Nat Rev Mol Cell Biol. 2022;23(3):204–226. doi:10.1038/s41580-021-00421-2
14. Li CW, Yu K, Shyh-Chang N, et al. Pathogenesis of sarcopenia and the relationship with fat mass: descriptive review. J Cachexia Sarcopenia Muscle. 2022;13(2):781–794. doi:10.1002/jcsm.12901
15. Jimenez-Gutierrez GE, Martínez-Gómez LE, Martínez-Armenta C, Pineda C, Martínez-Nava GA, Lopez-Reyes A. Molecular mechanisms of inflammation in sarcopenia: diagnosis and therapeutic update. Cells. 2022;11(15):2359. doi:10.3390/cells11152359
16. Perez K, Ciotlos S, McGirr J, et al. Single nuclei profiling identifies cell specific markers of skeletal muscle aging, frailty, and senescence. Aging. 2022;14(23):9393–9422. doi:10.18632/aging.204435
17. Migliavacca E, Tay SKH, Patel HP, et al. Mitochondrial oxidative capacity and NAD+ biosynthesis are reduced in human sarcopenia across ethnicities. Nat Commun. 2019;10(1):5808. doi:10.1038/s41467-019-13694-1
18. Becht E, McInnes L, Healy J, et al. Dimensionality reduction for visualizing single-cell data using UMAP. Nat Biotechnol. 2018. doi:10.1038/nbt.4314
19. Aran D, Looney AP, Liu L, et al. Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage. Nat Immunol. 2019;20(2):163–172. doi:10.1038/s41590-018-0276-y
20. Hänzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinformatics. 2013;14:7. doi:10.1186/1471-2105-14-7
21. Jin S, Guerrero-Juarez CF, Zhang L, et al. Inference and analysis of cell-cell communication using CellChat. Nat Commun. 2021;12(1):1088. doi:10.1038/s41467-021-21246-9
22. Barbie DA, Tamayo P, Boehm JS, et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature. 2009;462(7269):108–112. doi:10.1038/nature08460
23. Newman AM, Liu CL, Green MR, et al. Robust enumeration of cell subsets from tissue expression profiles. Nat Methods. 2015;12(5):453–457. doi:10.1038/nmeth.3337
24. Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics. 2008;9:559. doi:10.1186/1471-2105-9-559
25. Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57. doi:10.1038/nprot.2008.211
26. Yu G, Wang LG, Han Y, He QY. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS. 2012;16(5):284–287. doi:10.1089/omi.2011.0118
27. ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/19237447/.
28. Zhou Y, Zhou B, Pache L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019;10(1):1523. doi:10.1038/s41467-019-09234-6
29. Szklarczyk D, Franceschini A, Wyder S, et al. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015;43(Database issue):D447–452. doi:10.1093/nar/gku1003
30. Christian CJ, Benian GM. Animal models of sarcopenia. Aging Cell. 2020;19(10):e13223. doi:10.1111/acel.13223
31. Inflammaging: a new immune-metabolic viewpoint for age-related diseases - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/30046148/.
32. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: an expanding universe. Cell. 2023;186(2):243–278. doi:10.1016/j.cell.2022.11.001
33. Senescence atlas reveals an aged-like inflamed niche that blunts muscle regeneration - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/36544018/.
34. Xu P, Wang M, Song WM, et al. The landscape of human tissue and cell type specific expression and co-regulation of senescence genes. Mol Neurodegener. 2022;17(1):5. doi:10.1186/s13024-021-00507-7
35. Zhu Y, Tchkonia T, Pirtskhalava T, et al. The Achilles’ heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. 2015;14(4):644–658. doi:10.1111/acel.12344
36. Nelke C, Dziewas R, Minnerup J, Meuth SG, Ruck T. Skeletal muscle as potential central link between sarcopenia and immune senescence. EBioMedicine. 2019;49:381–388. doi:10.1016/j.ebiom.2019.10.034
37. Chen M, Wang Y, Deng S, Lian Z, Yu K. Skeletal muscle oxidative stress and inflammation in aging: focus on antioxidant and anti-inflammatory therapy. Front Cell Dev Biol. 2022;10:964130. doi:10.3389/fcell.2022.964130
38. Cardiac fibrosis: myofibroblast-mediated pathological regulation and drug delivery strategies - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/33831476/.
39. Yang AT, Kim YO, Yan XZ, et al. Fibroblast activation protein activates macrophages and promotes parenchymal liver inflammation and fibrosis. Cell Mol Gastroenterol Hepatol. 2023;15(4):841–867. doi:10.1016/j.jcmgh.2022.12.005
40. Plikus MV, Wang X, Sinha S, et al. Fibroblasts: origins, definitions, and functions in health and disease. Cell. 2021;184(15):3852–3872. doi:10.1016/j.cell.2021.06.024
41. Álvarez-Lorenzo S, Montalvo-Castro RE, Jiménez-López J, et al. Membrane-bound complement regulatory proteins in breast cancer: are they best therapeutic targets? In: Breast Cancer Updates. IntechOpen; 2023. doi:10.5772/intechopen.109945
42. He Y, Xie W, Li H, Jin H, Zhang Y, Li Y. Cellular senescence in sarcopenia: possible mechanisms and therapeutic potential. Front Cell Dev Biol. 2021;9:793088. doi:10.3389/fcell.2021.793088
43. Birch J, Gil J. Senescence and the SASP: many therapeutic avenues. Genes Dev. 2020;34(23–24):1565–1576. doi:10.1101/gad.343129.120
44. Coppé JP, Patil CK, Rodier F, et al. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008;6(12):2853–2868. doi:10.1371/journal.pbio.0060301
45. Huang KB, Gui CP, Xu YZ, et al. A multi-classifier system integrated by clinico-histology-genomic analysis for predicting recurrence of papillary renal cell carcinoma. Nat Commun. 2024;15(1):6215. doi:10.1038/s41467-024-50369-y
46. Frangogiannis NG. Transforming growth factor-β in myocardial disease. Nat Rev Cardiol. 2022;19(7):435–455. doi:10.1038/s41569-021-00646-w
47. Leask A, Abraham DJ. TGF-beta signaling and the fibrotic response. FASEB J. 2004;18(7):816–827. doi:10.1096/fj.03-1273rev
48. Dobaczewski M, Bujak M, Li N, et al. Smad3 signaling critically regulates fibroblast phenotype and function in healing myocardial infarction. Circ Res. 2010;107(3):418–428. doi:10.1161/CIRCRESAHA.109.216101
49. Prabhu SD, Frangogiannis NG. The biological basis for cardiac repair after myocardial infarction: from inflammation to fibrosis. Circ Res. 2016;119(1):91–112. doi:10.1161/CIRCRESAHA.116.303577
50. TLR4-dependent fibroblast activation drives persistent organ fibrosis in skin and lung - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/29997297/.
51. Schrimpe-Rutledge AC, Codreanu SG, Sherrod SD, McLean JA. Untargeted metabolomics strategies-challenges and emerging directions. J Am Soc Mass Spectrom. 2016;27(12):1897–1905. doi:10.1007/s13361-016-1469-y
52. Roh K, Noh J, Kim Y, et al. Lysosomal control of senescence and inflammation through cholesterol partitioning. Nat Metab. 2023;5(3):398–413. doi:10.1038/s42255-023-00747-5
53. Amorim JA, Coppotelli G, Rolo AP, Palmeira CM, Ross JM, Sinclair DA. Mitochondrial and metabolic dysfunction in ageing and age-related diseases. Nat Rev Endocrinol. 2022;18(4):243–258. doi:10.1038/s41574-021-00626-7
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.