Back to Journals » International Journal of Nanomedicine » Volume 21
Advances in Tetrahedral Framework Nucleic Acids (tFNAs) for Lung Injury Repair: Mechanisms, Therapeutic Applications, and Future Directions
Authors Song J, Chen X, Hou Q, Zhao P, Zu X ![]()
Received 24 March 2026
Accepted for publication 20 May 2026
Published 10 June 2026 Volume 2026:21 611318
DOI https://doi.org/10.2147/IJN.S611318
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Eng San Thian
Jipeng Song,1,2,* Xin Chen,2,* Quanjie Hou,1,2,* Pan Zhao,2 Xianpeng Zu1,2
1Department of Natural Medicinal Chemistry, School of Pharmacy, Naval Medical University, Shanghai, 200433, People’s Republic of China; 2School of Pharmacy, Shandong University of Traditional Chinese Medicine, Jinan, 250355, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xianpeng Zu, Department of Natural Medicinal Chemistry, School of Pharmacy, Naval Medical University, No. 325 Guohe Road, Yangpu District, Shanghai, People’s Republic of China, Email [email protected] Pan Zhao, School of Pharmacy, Shandong University of Traditional Chinese Medicine, No. 4655 Daxue Road, Changqing District, Jinan, People’s Republic of China, Email [email protected]
Abstract: Lung injury is a pathological condition caused by a range of direct or indirect factors that disrupt the alveolar-capillary barrier and damage the structure of the lung parenchyma, leading to impaired gas exchange and compromised respiratory function. Lung injury shows a high incidence worldwide and remains associated with high mortality, and safe, effective, and reliable therapeutic strategies for this condition are still lacking. Tetrahedral framework nucleic acids (tFNAs) represent a significant advancement in nucleic acid nanotechnology. As novel therapeutic agents and nanocarriers, tFNAs possess an intrinsic capacity to scavenge reactive oxygen species, thus mitigating oxidative stress. Additionally, their unique tetrahedral geometry facilitates endocytosis-mediated cellular uptake and confers them with excellent tissue permeability, enabling them to traverse the lung mucosal barrier. These properties collectively enhance cell–drug interactions. As a result, tFNAs show multidimensional therapeutic potential for the treatment of lung injuries and provide innovative strategies for managing complex pathological conditions. This review summarizes the recent advances in the application of tFNAs for lung injury repair, with a focus on their mechanisms of action, treatment strategies, and current challenges. The goal of this review is to promote breakthroughs in lung injury treatment. This infographic illustrates all aspects of tetrahedral framework nucleic acid (tFNA) in lung injury treatment. The upper left part, “Intelligent Design of tFNA”, covers responses to ROS, miRNA, pH, and enzymes, which trigger drug release. The upper right part, “Challenges Faced by tFNA”, lists objectives, biosafety, and high cost. The lower right part, “tFNA as Delivery Carrier”, highlights cantilever, sequence replacement, mosaic, and vertex. The lower left part, “tFNA as a Drug”, emphasizes anti-inflammatory effects on macrophages (M1 to M2), ROS neutralization, immune regulation of T cells, and tissue repair. At the bottom, the advantages of tFNA include biodegradability, programmability, cell uptake, and tissue permeability.An infographic on the design, challenges, advantages and applications of tFNAs in the treatment of lung injury.
Keywords: self-assembled DNA nanomaterials, tetrahedral framework nucleic acid, lung injury, pulmonary fibrosis, drug delivery, targeted therapy
Introduction
Lung injury has long been a serious public health concern worldwide. The common etiologies of lung injury include pneumonia, smoking, transfusion-related complications, and other inflammatory conditions. In recent years, with rising environmental pollution, the number of lung injury cases has increased substantially, impacting global incidence rates, patients’ quality of life, and overall public health safety.1–3 Lung injury, as a core pathological link in the onset and progression of respiratory diseases, is characterized by pathological changes, disruption of structural integrity, and gas exchange dysfunction in lung tissue. Based on the disease course and clinical features, lung injury can be classified into two major categories: acute lung injury (ALI) and chronic lung injury. ALI is a clinical syndrome characterized by diffuse alveolar damage, inflammatory exudation, and pulmonary edema. Pathologically, it manifests as extensive injury to the alveolar epithelium and vascular endothelium, exudation of protein-rich edematous fluid, and formation of hyaline membranes.4,5 Its most severe and critical form, acute respiratory distress syndrome (ARDS), is frequently encountered in clinical practice, characterized by rapid onset and high mortality rates. In an international observational study conducted in 2019, the morbidity and mortality rates among 23280 ARDS patients across 27 countries were found to be 17% and 3%, respectively, highlighting the complex and multifactorial pathogenesis of this condition.6 Furthermore, a separate study covering 50 countries and regions found that ARDS accounted for 10.4% of intensive care unit (ICU) admissions, with mortality rates reaching up to 40%.7 Unfortunately, research shows that long-term quality of life remains impaired among ARDS survivors.8,9 In contrast to ALI, chronic lung injury involves prolonged, progressive structural and functional damage to lung tissues and typically progresses slowly. Its prominent pathological changes include persistent inflammation, fibroblast activation, and excessive deposition of extracellular matrix, ultimately leading to irreversible damage and pathological changes.10 Chronic lung injury serves as a common pathological foundation for a range of chronic lung diseases, including pulmonary fibrosis, radiation-induced lung injury, lung cancer, and chronic obstructive pulmonary disease, all of which pose serious threats to human health and safety. However, conventional treatment approaches such as pharmacotherapy, surgical interventions, and physical therapies often exhibit limited efficacy, lead to significant side effects, and carry a high risk of recurrence. In this context, the rapid development of nanomaterial science has offered new opportunities to optimize therapeutic strategies for lung injury and to advance the development of targeted pharmacotherapies.
Owing to the emergence and rapid development of nanomaterials, revolutionary biomedical tools have now been developed. Deoxyribonucleic acid (DNA), composed of monomeric nucleotides, serves as the molecular basis for storing genetic information. With its characteristic double helix structure, DNA plays a critical role in regulating biological functions. In 1982, Professor Nadrian Seeman proposed the groundbreaking concept that DNA could be utilized as a structural material capable of self-assembling into geometrically defined nanoscale objects. This idea laid the foundation for the development of DNA nanostructures.11 Among the most promising of these structures are tetrahedral framework nucleic acids (tFNAs), first synthesized by Turberfield et al in the early 2000s using a one-pot annealing method.12 tFNAs are novel three-dimensional nanomaterials constructed via DNA nanotechnology. They consist of four equidistant, programmable single-stranded DNA molecules and self-assemble based on the Watson-Crick base pairing principle. Owing to their unique design, tunability, and biocompatibility, tFNAs have garnered significant attention and have emerged as a versatile platform for applications in diagnostics, therapeutics, drug delivery, and regenerative medicine. In recent years, the optimization of tFNAs for biomedical applications has become a major focus of research. Numerous studies have demonstrated the potential of tFNAs in biosensing,13,14 targeted drug delivery,15–17 tissue regeneration,18,19 and even as therapeutic agents for serious diseases.15,20,21 These broad application prospects have attracted considerable interest from researchers in the field of biomedicine.
With advances in science and technology and growing clinical demands, the field of medical materials is entering a new era of innovation. The emergence of novel medical materials is increasingly supporting precision medicine and personalized therapy.22 Compared to conventional materials, the new medical materials offer superior biocompatibility, flexibility, biodegradability, and multifunctional therapeutic potential. They not only help reduce healthcare costs but also maintain or enhance clinical efficacy, providing patients with safer and more effective diagnostic and treatment plans. tFNAs are a classic example of this new class of advanced medical materials. Building upon their biological properties and regulatory functions, recent studies have highlighted their use as therapeutic agents and drug carriers.
This review explores the potential clinical applications of tFNAs in the context of lung injury repair, analyzes the challenges associated with their clinical translation, and provides an outlook on future development directions. The goal of this review is to inform and guide ongoing development in the field of nanoscale framework nucleic acid structures to address key healthcare needs.
Pathological Features and Challenges for Treating Lung Injury
The lungs are highly complex organs. The endothelial cells of the lungs, as key components of the capillary-alveolar unit, form a semipermeable selective barrier essential for maintaining normal pulmonary function. This barrier regulates blood flow, restricts excessive fluid extravasation, and hinders the infiltration of inflammatory cells.23 An intact alveolar epithelial barrier maintains a relatively dry alveolar environment, free from inflammatory cell infiltration, thereby ensuring effective gas exchange.24 The vast surface area of these endothelial cells enables the lungs to efficiently oxygenate blood before it enters the systemic circulation. Therefore, the structural integrity of the alveolar epithelium and capillary endothelium is crucial for maintaining lung homeostasis.
ALI
ALI is a clinical syndrome characterized by acute diffuse alveolar damage. It presents with bilateral pulmonary infiltrates on chest imaging as well as hypoxemia, without left atrial hypertension. ALI is primarily triggered by direct lung insults, such as pneumonia and aspiration, or by systemic inflammatory events, including sepsis. ARDS represents the severe end-stage manifestation of ALI, marked by profound hypoxemia, extensive alveolar edema, and a higher mortality rate. Both ALI and ARDS share similar pathophysiological mechanisms, including increased pulmonary capillary permeability, inflammatory cell infiltration, and disruption of the alveolar-epithelial barrier. However, ARDS is associated with more severe respiratory impairments and multi-organ dysfunction. Overall, ALI and ARDS are serious multifactorial respiratory conditions associated with lung inflammation and can be highly fatal.
The pathogenesis of ALI/ARDS involves multiple factors, including pneumonia caused by environmental particles, bacterial or viral infections, sepsis, trauma, the aspiration of gastric contents, drowning, and transfusion-related infections.25–27 These triggers initiate an inflammatory response that is mediated by polymorphonuclear leukocytes (PMNs), macrophages, alveolar epithelial cells, and endothelial cells. Following direct or indirect injury to lung tissue, these cells become activated. Then, they begin secreting pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-1 (IL-1), and IL-8, along with inflammatory mediators such as elastase, collagenase, and various chemokines. This molecular cascade leads to further cellular damage and perpetuates a self-amplifying cycle of inflammation.28 In the presence of pro-inflammatory factors, endothelial cells express adhesion molecules such as P-selectin, E-selectin, and intercellular adhesion molecule-1, which interact with anti-receptors such as P-selectin glycoprotein ligand-1 and β-2 integrin on PMNs. These interactions allow endothelial cells to capture PMNs, which subsequently transmigrate through the endothelium to reach sites of inflammation.23 The activation and migration of PMNs provoke alveolar inflammation, damaging both the vascular endothelium and alveolar epithelium. This damage compromises the integrity of the alveolar-capillary barrier, increasing the permeability of vascular endothelial and epithelial barriers and allowing the leakage of protein-rich fluids—including plasma and inflammatory cells—into the alveolar space. Such leakage results in pulmonary edema, a reduced alveolar surface area, and impaired gas exchange.29–31 Therefore, the effective management of pulmonary edema is essential to maintain adequate gas exchange in patients with ALI/ARDS.
At present, the treatment strategies for ALI/ARDS include respiratory support therapy, cell transplantation, gene therapy, and drug treatment. The drugs commonly used in clinical practice to treat ALI include glucocorticoids, anti-oxidative stress drugs, non-steroidal anti-inflammatory drugs, and vasodilators, but these agents often cause significant adverse effects. The prolonged or high-dose administration of glucocorticoids can increase the risk of infection, hyperglycemia, osteoporosis, gastrointestinal ulcers, and other adverse outcomes.32 Moreover, the pathogenesis of ALI/ARDS involves a complex interplay of cellular processes and molecular pathways, which form intricate regulatory networks. This complexity hinders the identification of precise therapeutic targets.33 Consequently, there is a pressing need to develop more targeted pharmacological treatments with reduced side effects.
Chronic Lung Injury
Pulmonary Fibrosis
Pulmonary fibrosis is a fatal, chronic, and progressive fibrotic interstitial lung disease characterized by excessive extracellular matrix (ECM) deposition and the formation and thickening of alveolar wall scars. The primary pathological process of pulmonary fibrosis involves the chronic inflammation triggered by initial injury or exposure to internal or external pathogenic factors. This eventually leads to aberrant tissue repair and progressive functional impairment of culminating in respiratory failure and death. Currently, the imbalance between tissue damage and wound repair is considered the fundamental mechanism underlying fibrosis. This imbalance manifests in two major physiological processes: the dysregulated proliferation or apoptosis of fibroblasts and myofibroblasts, and the abnormal synthesis or degradation of ECM components.34 Myofibroblasts, a specialized group of fibroblasts characterized by the presence of stress fibers and adhesion plaques, play a central role in mediating wound healing and are key contributors to pulmonary fibrosis.35 Transforming growth factor-β1 (TGF-β1), a pro-fibrotic cytokine derived from macrophages, promotes the proliferation of fibroblasts and enhances their differentiation into myofibroblasts via the fibroblast-to-myofibroblast transition (FMT), and it also enhances ECM contraction by stimulating the formation of contractile stress fibers.15 Furthermore, TGF-β1 induces the epithelial–mesenchymal transition (EMT), wherein alveolar epithelial cells acquire mesenchymal characteristics, leading to increased ECM production and accumulation in the alveolar wall.36 This process enhances cellular migratory capacity and causes irreversible damage to the alveolar architecture. Under normal physiological conditions, fibroblasts migrate to sites of injury and differentiate into myofibroblasts under the regulation of TGF-β1. These myofibroblasts are responsible for producing ECM components such as collagen, proteoglycans, fibronectin, and other matrix molecules in response to inflammatory and chemotactic signals from immune cells and macrophages. In healthy tissues, ECM production is tightly regulated, and most myofibroblasts undergo apoptosis after wound healing. Meanwhile, a small proportion of these cells transform into quiescent fibroblasts to maintain tissue homeostasis.37,38 However, persistent inflammation promotes sustained myofibroblast differentiation and inhibits myofibroblast apoptosis. This leads to excessive ECM accumulation and abnormal deposition, ultimately causing alveolar wall scarring and fibrotic remodeling.31,34,39
Unfortunately, once pulmonary fibrosis becomes irreversible, the median survival time after diagnosis is only 3 to 5 years.40 Given that its mortality rate is higher than that of many cancers, pulmonary fibrosis is often referred to as a “tumor-like disease.” In 2014, pirfenidone (PFD) and nintedanib were approved for the treatment of clinical pulmonary fibrosis by the US Food and Drug Administration. Although these drugs improve lung function and enhance patients’ quality of life, they do not eradicate the disease. Moreover, both compounds are poorly soluble in water, undergo rapid degradation and excretion, and do not achieve sufficient concentrations at sites of fibrosis, resulting in low treatment efficacy. Additionally, they are associated with significant adverse effects, including dermatologic reactions (rash), gastrointestinal bleeding, vomiting, and severe diarrhea, which limit their widespread clinical use.41,42 Importantly, the management of pulmonary fibrosis continues to be hindered by the lack of curative therapies, as current treatments merely slow disease progression instead of inducing complete remission. Known molecular targets for pulmonary fibrosis include TGF-β1, receptor tyrosine kinases, connective tissue growth factor, and the Wnt/β-catenin signaling pathway. However, the pharmacological agents targeting these pathways are still being developed. Therefore, there is an urgent need to enhance the targeted delivery of therapeutic agents to lung tissue in order to improve drug efficacy and clinical outcomes.
Radiation-Induced Lung Injury (RILI)
Radiotherapy is a common strategy for the treatment of malignant tumors in the chest and abdomen, including gastric cancer, lymphomas, and breast tumors. However, the lung is a radiosensitive organ. Hence, radiotherapy inevitably causes some damage to lung tissues. Therefore, RILI limits the ability to increase radiation doses to tumor tissues and consequently reduces the overall effectiveness of radiotherapy.43 RILI includes both early radiation pneumonitis and late-stage pulmonary fibrosis, and its pathogenesis involves a complex interplay of multiple cell types and molecular mediators.44 Early radiation pneumonitis is caused by an exudative inflammatory response. Following the exposure of lung tissues to ionizing radiation, reactive oxygen species (ROS) are generated in large quantities. Alveolar epithelial cells, as an important component of the alveolar barrier, are the first to experience oxidative damage. This damage initiates a series of inflammatory reactions and PMN chemotaxis, leading to the altered secretion of inflammatory cytokines, chemokines, and growth factors such as IL-4, IL-13, interferon-γ (IFN-γ), prostaglandin E2 (PGE2), and TGF-β1.45,46 These chemotactic signals recruit PMNs and macrophages to the alveoli and interstitium of the lungs, triggering an acute inflammatory response that is accompanied by extensive fibrin exudation. In the later stages of RILI, abnormal tissue repair processes result in the excessive secretion and deposition of ECM, which disrupts the normal tissue architecture, reducing alveolar surface area and progressively impairing pulmonary function. Macrophage-derived TGF-β1 plays a crucial role in promoting the accumulation of fibroblasts and myofibroblasts. Notably, it drives fibrotic progression by facilitating alveolar EMT and FMT, ultimately causing the structural remodeling of lung tissues.47,48
Currently, the treatment of RILI is associated with several major challenges. First, the disease course exhibits spatiotemporal heterogeneity: the acute phase necessitates the rapid suppression of the inflammatory cascade, whereas the chronic phase requires the reversal of fibrosis. In addition, significant individual variations in radiosensitivity complicate the development of personalized treatment plans. As RILI is a complication of radiotherapy for thoracic and abdominal tumors, any therapeutic interventions need to avoid interference with ongoing tumor treatment. In addition, drug delivery is a major challenge because the physiological barriers within the lungs make it challenging to achieve therapeutic concentrations at the site of injury following systemic administration, while inhaled formulations show limitations such as uneven distribution and poor formulation stability.
Lung Cancer
Lung cancer is a malignant respiratory tract tumor that originates in the epithelium of the bronchial mucosa and has long remained a significant clinical challenge. Similar to other chronic diseases, lung cancer is a multifactorial condition that develops slowly under the influence of various risk factors. These include long-term exposure to cigarette smoke, environmental pollutants (eg, PM25), and specific chemical agents.25,49 One of the primary mechanisms through which these risk factors contribute to tumorigenesis is the targeted induction of DNA damage in epithelial cells, leading to mutations in oncogenes, such as RAS and ALK gene rearrangements and loss, as well as the inactivation of tumor suppressor genes such as TP53, RB, and P16. These genetic alterations result in uncontrolled cell proliferation, resistance to apoptosis, and disruption of cell cycle regulation.50 Growth factors—peptide molecules produced in large quantities by various types of cells—play pivotal roles in regulating cell proliferation, differentiation, metastasis, and apoptosis. The receptors for these growth factors are located on the cell surface and are critical for transmitting extracellular signals into the intracellular environment. Among these, epidermal growth factor (EGF) and its receptor (EGFR) act as key regulators in lung cancer development. The disruption of these molecules is a key contributor to the pathogenesis of lung cancer.51 The formation of blood vessels is also crucial for both normal and pathological tissue growth. This process is typically self-limiting, but in malignant tumors, abnormal angiogenesis occurs because the division rate and survival time of endothelial cells become several hundred-fold higher. Vascular endothelial growth factor (VEGF) is among the most important and strongest angiogenic mediators. In cancer, including lung cancer, the VEGF signaling pathway is persistently activated, shifting the balance in favor of pro-angiogenic over anti-angiogenic signals, thereby promoting tumor vascularization and growth. VEGF mRNA is abundantly expressed in tumor cells, including lung cancer cells. Tumor cells facilitate capillary proliferation by secreting VEGF, enabling the establishment of a tumor-specific microcirculation.52,53 Meanwhile, the resident immune cells within the alveolar microenvironment are polarized by tumor cells to produce immunosuppressive cytokines such as IL-10, creating an immunosuppressive microenvironment that further supports tumor development and progression.54
In the context of lung cancer drug therapy, three major clinical challenges have emerged: enhancing drug targeting, overcoming multidrug resistance, and minimizing adverse effects. These challenges collectively limit the efficacy of current treatments in inhibiting tumor growth, metastasis, and recurrence.55 Therefore, there is an urgent need to develop safe and precise drug delivery systems, formulate multi-targeted therapeutic strategies, and explore dynamic drug resistance monitoring technologies based on tumor evolution trajectories. The mechanism diagram illustrating lung injury for Figure 1.
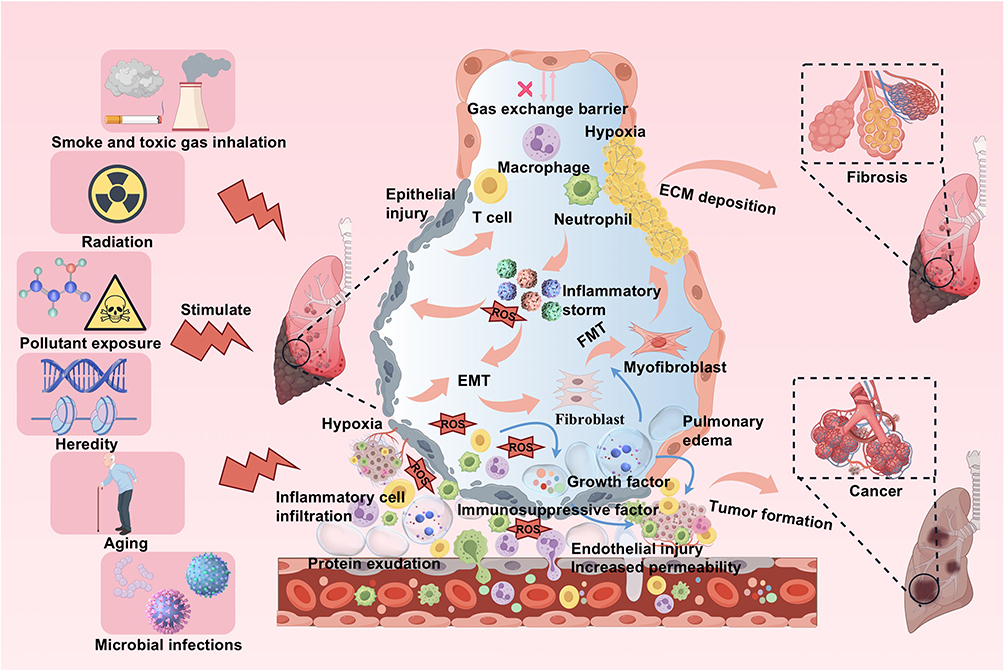 |
Figure 1 External stimuli impair normal cell behavior and lung parenchyma in the alveoli, destroy the alveolar-capillary barrier, and induce lung injury. Factors such as smoke, radiation, pollutants, genetics, and microorganisms can damage normal cell behavior in the alveoli, causing inflammatory storms and the release of growth factors, while also disrupting the alveolar capillary barrier, leading to increased vascular permeability, infiltration of inflammatory cells (macrophages, neutrophils, T cells), and infiltration of protein-rich fluids into the alveolar cavity. This results in pulmonary edema, ECM deposition, EMT, FMT, and immune dysfunction. It will eventually lead to hypoxia and may lead to serious consequences such as fibrosis and tumor formation. |
Emergence of Nanomaterials and the Characteristics of tFNAs
Emergence of Nanomaterials
Humankind’s understanding of matter at the nanoscale level can be traced back to the 4th century BC, when the ancient Greek philosopher Democritus proposed the atomic theory. During the Middle Ages, alchemists unintentionally produced gold sols by heating metal salt solutions. These colloidal systems, which exhibited unique optical properties, were used to color glass, although their underlying nature remained unknown at the time.
Nanomaterials are defined as materials that possess at least one dimension within the nanoscale range (1–100 nm) or are composed of nanoscale units in a three-dimensional configuration. Lying between micro and macroscale systems, nanomaterials encompass a variety of structures, including nanoparticles, nanosheets, and nanocapsules.56 Based on their intrinsic properties, nanomaterials can be classified into several categories, such as inorganic nanomaterials, organic nanomaterials, metallic nanomaterials, and biological nanomaterials. In 1857, Michael Faraday conducted the first systematic study on the synthesis and properties of gold nanoparticles.57 This observation marked the early identification of the size-dependent properties of nanoparticles. However, progress in the field remained limited for decades due to inadequate technologies. Nevertheless, a significant breakthrough was made after the invention of the scanning tunneling microscope, which enabled the direct visualization and manipulation of individual atoms for the first time. This technological advancement catalyzed the emergence of nanomaterials and the rapid development of associated technologies. Following this, numerous innovative design concepts and novel nanomaterials such as DNA nanostructures were introduced, unlocking new opportunities across a wide array of sectors, including healthcare, environmental protection, construction, and agriculture. In the field of medicine in particular, the integration of nanotechnology is transforming traditional models of disease prevention, diagnosis, and treatment.
Nanotechnology and biomaterials have since jointly opened novel avenues of research. DNA is a fundamental biomolecule with high efficiency and stability, having evolved over time, and has found applications for treating various clinical diseases. In 2004, Turberfield et al introduced tFNAs, the simplest polyhedral DNA nanostructures. Following this, in 2006, scientists developed a groundbreaking technology for DNA self-assembly known as DNA origami.58 The rise of DNA origami technology created new opportunities and strategies for the development of DNA nanomaterials.
DNA nanostructures, as a new class of nanomaterials developed in recent years, have undergone four stages of evolution.59 2D DNA tiles constituted the first iteration of these nanostructures, while the second generation included rigid DNA double crossover structures. This was followed by the third generation of 3D DNA nanostructures, and finally the fourth generation of arbitrarily shaped DNA nanostructures. These progressive improvements have significantly lowered the complexity of fabricating large-scale, multi-layered DNA structures with dynamic environmental response capabilities, expanding the applications of DNA nanomaterials.60,61 Compared to other polyhedral DNA structures, the synthesis of tetrahedral nanostructures is simpler and more efficient, and yields of up to 90% can be achieved (versus 76% for dodecahedra and only 69% larger buckyball-like structures). This is consistent with the general decrease in yield as the size of the target structure increases.62 Therefore, among the various types of DNA polyhedra, tFNAs remain the most studied and impactful.
Characteristics of tFNAs
One of the most promising applications of nanomaterials in biomedicine lies in drug delivery. However, with expanding research into nanotechnology, the challenges associated with liposomes, nanoparticles, and other carriers—such as excessive toxicity, non-degradability, insufficient loading efficiency, poor stability, and lack of specificity—have gradually become apparent.63,64 The accumulation of toxic nanoparticles and the difficulty of degrading these inorganic nanomaterials have necessitated the development of nanomaterials with high biocompatibility and degradability. In this context, tFNAs stand out as representative third-generation nanomaterials due to their unique structural and functional properties.
Notably, tFNAs possess features such as programmability, biocompatibility, degradability, and high stability.12,65,66 Their editability is reflected in the flexibility of their structural design, and researchers can precisely control the size of tFNAs by adjusting the sequence length of individual DNA strands. The editability of tFNAs also manifests in the presence of numerous modifiable sites, which can be controlled via chemical coupling or complementary base pairing to assemble functional groups at specific sites. This allows functions to be achieved after modification.66 Meanwhile, DNA tetrahedra can be intelligently controlled through clever designs to respond to various stimuli. Since DNA is a natural biomacromolecule, DNA materials exhibit good biocompatibility, facilitating their use in various biological applications. tFNAs can also be efficiently taken up by various cells, such as lung fibroblasts, Schwann cells, and chondrocytes, and promote significant cell proliferation.15,67,68 They also show limited cytotoxicity, which is another important aspect of biocompatibility, indicating their potential role in promoting cell growth. As DNA nanostructures, tFNAs can be gradually degraded in vivo by nucleases to produce nucleotide fragments without the long-term risk of accumulation.69 Their inherent stability derives from a hydrogen bonding network formed by complementary base pairing, enabling tight strand binding, which prevents their dissociation. The tetrahedral geometric shape exhibits good spatial symmetry, effectively distributing molecular stresses and further enhancing structural stability. This tetrahedral configuration contributes to the stability of tFNAs in complex environments, which is fundamental for biological applications. Compared to linear DNA, tFNAs show significantly higher resistance to enzymatic degradation in solutions containing 10% fetal bovine serum, largely due to the triple branching connections at their vertices and their unexposed 3′ ends, which collectively enhance structural integrity.70,71 In contrast to 24-hour DNA octahedra, tFNAs maintain their integrity within cells for at least 48 hours, likely due to their higher DNA density and shorter oligonucleotide components.72–74 Additionally, tFNAs as DNA nanomaterials have side lengths under 10 nm and can self-assemble quickly.12 Thus, tFNAs are providing considerable advantages as new carrier materials.
Biological Mechanisms Underlying the Role of tFNAs in Lung Injury Repair
Targeted Delivery and Synergistic Drug Treatment
Traditional drug delivery systems have several limitations, including inadequate bioavailability, suboptimal targeting, and a heightened risk of cytotoxicity. With the rapid advancement of nanomedicine, the integration of nanotechnology into precision drug delivery has become a major focus of contemporary biomedical research. This is particularly pertinent in the context of pulmonary disease treatment, wherein precise drug delivery, minimal adverse effects, and improved therapeutic efficacy are critical objectives. Harnessing the unique properties of DNA, researchers are developing efficient, safe, targeted, and multifunctional DNA nanostructures as delivery vehicles. The construction of intelligent therapeutic DNA nanomachines presents both substantial scientific interest and clinical promise. Extensive studies have demonstrated that tFNAs, designed based on the principles of Watson-Crick base pairing in single-stranded DNA, can achieve the precise loading and targeted delivery of functional nucleic acids, aptamers, or small-molecule drugs. This is accomplished through strategies such as vertex modification, mosaic modification, sequence replacement, and cantilever design, enhancing the therapeutic potential of tFNAs (Figure 2). Although tFNAs can serve as a structure for loading various therapeutic agents, the methods and rates of loading may vary. Vertex modification involves functionalizing the vertices of tFNAs with specific groups that may serve to stabilize the tFNA nanostructure, facilitate molecular recognition, or deliver biologically active compounds.75–77 Mosaic modification entails the conjugation of functional molecules or chemical groups to the double-helical regions of tFNAs. In some studies, for instance, embedding pirfenidone into the double-helical groove of tFNAs led to the formation of a stable complex and significantly enhanced therapeutic outcomes.15 In contrast, sequence replacement involves substituting one or more strands, or specific segments of strands, within the tFNA with custom-designed nucleic acid sequences to achieve functionalization. Through this strategy, combined with the precise design of each strand and a stable combination of DNA duplexes, three single strands of a tFNA were replaced with three independent miR-155 inhibitors, effectively restoring immune homeostasis in the lungs.78 Finally, cantilever modification refers to the attachment of functional groups or molecules to the side arms of tFNAs. This involves designing the base sequences such that the intersection of the 5′ and 3′ ends lies at the edge, center, or another non-terminal region of the tFNA. In previous studies, chemotherapeutic agents were successfully applied to the side arms of tFNAs for tumor treatment using this method.79 Additionally, specific DNA sequences within tFNAs themselves have demonstrated inherent therapeutic effects in cases of some diseases.15
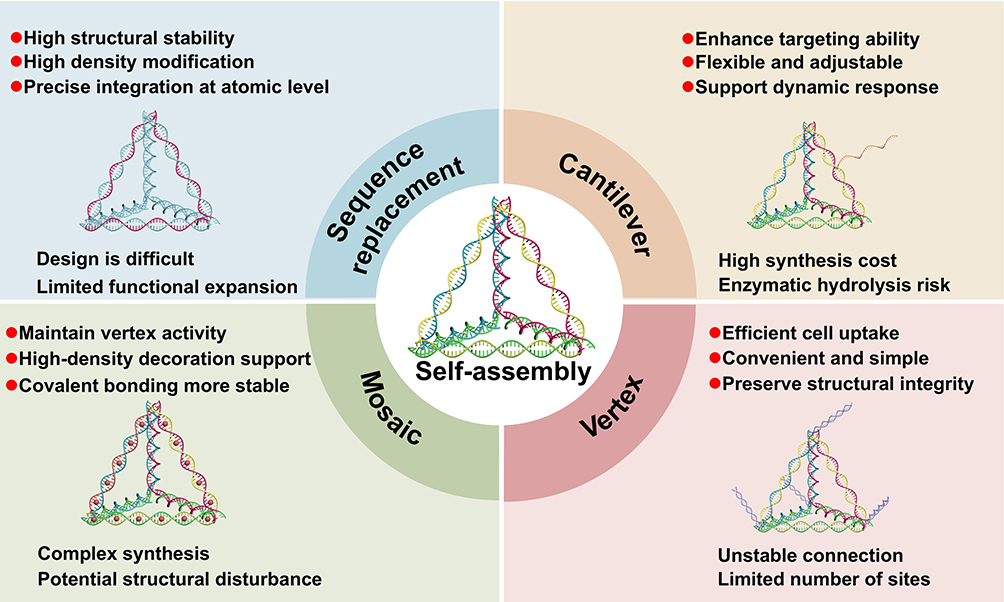 |
Figure 2 The advantages and disadvantages of multifunctional modification methods based on nano-platform tFNAs in the treatment of lung injury. |
One of the most significant advantages of the tFNA structure is its ability to enhance endocytosis and tissue penetration. Compared to most organic or inorganic nanoparticles, tFNAs are natural biomacromolecules with excellent biocompatibility and biodegradability and can penetrate mammalian cells without requiring lipid-based transfection. Single-stranded and double-stranded DNA molecules are typically impermeable to the plasma membrane due to their polyanionic properties. Moreover, exposed oligodeoxynucleotides are not readily internalized and are rapidly degraded by nucleases in the serum or the cytoplasm.80 However, the unique spatial topology of tFNAs gives them excellent transmembrane permeability, allowing them to act as effective transport vehicles. Upon contact with the cell membrane, the antenna-like extensions of tFNAs adhere to the membrane, reducing charge repulsion and redistributing membrane surface charges. They then efficiently traverse the cell membrane via the caveolin-mediated pathway, facilitating successful cellular uptake. When conjugated with additional aptamers for nuclear localization, tFNAs can also bypass lysosomal degradation.81,82 This caveolin-mediated internalization mechanism involves the formation of caveolae (membrane invaginations rich in caveolin proteins) that selectively capture and transport specific molecules into cells, enabling targeted delivery. The high intracellular uptake efficiency of tFNAs, along with their surface-exposed nucleotide sequences, enables the co-delivery of therapeutic cargos, including negatively charged nucleic acids, aptamers, and neutrally charged peptides that selectively recognize overexpressed markers at injury sites.72,83,84 The stability of DNA nanostructures is influenced by factors such as concentration, intracellular or extracellular nuclease activity, and innate immune responses. Notably, tFNAs have demonstrated structural stability in living cells for up to 48 hours, meeting the essential requirements for effective drug delivery systems.85 In co-delivery applications, tFNAs protect charged molecules from enzymatic degradation through steric hindrance and enhance cellular uptake by modulating surface charge, thereby reducing systemic toxicity while preserving therapeutic efficacy.86 These advances provide a solid theoretical and technical foundation for the development of lung injury treatments.
Anti-Inflammatory and Immunoregulatory Effects
Inflammation is typically triggered by host immune responses to physical or chemical stimuli or pathogenic microorganisms. In several diseases, uncontrolled inflammation is a key pathological process, and is mediated primarily through four mechanisms: ROS-induced oxidative stress, the JAK-STAT pathway, the nuclear factor κ light chain enhancer (NF-κB) pathway, and the mitogen-activated protein kinase (MAPK) pathway.87 Hence, tFNAs could modulate inflammatory signaling pathways via multiple targets. Zhang et al comprehensively analyzed the immunomodulatory effects of tFNAs on RAW264.7 cells and found that 250 nM tFNA could reduce the expression of nitric oxide, IL-1β, IL-6, and TNF-α in lipopolysaccharide (LPS)-stimulated RAW264.7 cells by inhibiting MAPK phosphorylation.88 Following this, the promising anti-inflammatory potential of tFNAs was extensively explored.15,89 Macrophages, as key effector cells responsible for secreting pro-inflammatory cytokines, phagocytosing pathogens, and clearing cellular debris, are central to the initiation and progression of inflammatory responses. These cells can polarize into either M1 or M2 phenotypes in response to microenvironmental signals, adopting either pro-inflammatory or anti-inflammatory roles, respectively.90 Inducible nitric oxide synthase (iNOS) and arginase serve as markers for the M1 and M2 phenotypes, respectively. M1 macrophages, characterized by iNOS expression, secrete pro-inflammatory cytokines that contribute to secondary inflammation and are thus referred to as pro-inflammatory macrophages. In contrast, M2 macrophages, marked by arginase expression, release factors that promote tissue repair and are known as repair macrophages. Owing to their excellent biocompatibility, tFNAs exhibit strong anti-inflammatory effects after cellular uptake. They can promote the polarization of M1 macrophages toward the M2 phenotype and suppress the production of inflammatory cytokines such as TNF-α and IL-6, thereby exerting potent anti-inflammatory effects.91
T cells are known contributors to autoimmune diseases, and they also play a crucial role in the development of systemic and local inflammation. Self-assembled tFNAs have been shown to enter and accumulate in the cytoplasm of primary human T cells, where they inhibit the excessive secretion of pro-inflammatory cytokines by activated T cells. Mechanistically, tFNAs regulate T cell-mediated adaptive immunity by inhibiting the phosphorylation of c-Jun N-terminal kinase (JNK) and extracellular signal-regulated kinase (ERK) under physiological conditions. In vitro studies further confirm that tFNAs possess significant immunomodulatory effects under pathological conditions, effectively suppressing pro-inflammatory cytokine secretion.89 Moreover, related research has demonstrated that tFNAs can regulate both T cell differentiation and macrophage polarization, highlighting their broad immunomodulatory potential (Figure 3).65,92
 |
Figure 3 The tFNAs promote lung injury repair by driving M2 transition, inhibiting T cells, scavenging ROS, and regulating cell cycles. tFNAs play an immunomodulatory role by promoting macrophage polarization to the reparative M2 phenotype, regulating T cell factors, inhibiting pro-inflammatory factors such as TNF-α and IFN-γ, and enhancing the anti-inflammatory factor IL-10. Together, these changes inhibit inflammation, promote tissue healing and vascular repair, and reduce scar formation. The green upward and downward arrows indicate upregulation and downregulation, respectively; the blue and red T-arrows indicate inhibition. |
Anti-Oxidative Stress
Oxidative stress, resulting from an imbalance between oxidative and antioxidant capacities within an organism, plays a critical role in the pathogenesis of numerous diseases. In inflammatory microenvironments, ROS levels are elevated, contributing to the release of proteases and other detrimental intermediates. This process directly damages intracellular proteins and lipids and thereby exacerbates oxidative stress. Studies have shown that the ROS-induced apoptosis of alveolar epithelial cells significantly impairs normal lung tissue repair and accelerates the progression of fibrosis.93 Notably, tFNAs have been identified as highly effective ROS scavengers, providing both antioxidant and anti-apoptotic benefits across a range of diseases. This ROS-eliminating capacity can be attributed to the intrinsic properties of DNA, which naturally possesses the ability to neutralize ROS.94 In contrast to single-stranded DNA, the unique tetrahedral geometry and efficient cellular uptake of tFNAs enhance their ROS-clearing potential.95 The interplay of ROS, inflammation, and apoptosis has been implicated in the pathogenesis of fibrosis. Research by Zhang et al demonstrated a significant reduction in ROS generation in rat type II alveolar epithelial cells (RLE-6TN) following treatment with tFNA. This indicated that tFNAs are promising DNA-based nanomedicine agents, capable of modulating pulmonary fibrosis progression through both anti-inflammatory effects and suppression of ROS production.96 Heme oxygenase-1 (HO-1), a cytoprotective factor regulated by nuclear factor erythroid 2-related factor 2 (NRf2), plays a pivotal role in alleviating cellular stress, controlling apoptosis, and managing inflammation. Notably, tFNAs have been shown to inhibit LPS-induced ROS generation and apoptotic cell death in macrophages by upregulating HO-1 mRNA expression.88 The potent anti-inflammatory and antioxidant properties exhibited by tFNAs upon cellular uptake present a novel approach for organ protection (Figure 3).
Promoting Tissue Repair and Regeneration
Interestingly, tFNAs also exert beneficial effects on cellular behavior, with their potential for tissue repair and regeneration attributed to multiple molecular mechanisms. These include the inhibition of apoptosis, stimulation of cell division, promotion of cell proliferation and migration, suppression of inflammatory cytokine expression, reduction of extracellular polysaccharide and biofilm formation, and regulation of ROS production.97 Collectively, these functions contribute to enhanced wound healing, minimize scar formation, and improve the regeneration of vascular and neural tissues. Studies have demonstrated that tFNAs promote cell proliferation by activating Wnt/β-catenin signaling.98 In particular, mouse L929 fibroblasts were found to exhibit the highest proliferation rate after 24 hours of exposure to 250 nM tFNA. This effect was likely associated with the upregulation of cyclin-dependent kinase-like protein, which regulates the S phase of the cell cycle. However, it is important to note that tFNAs did not significantly affect the cell cycle in bronchial epithelial cells, cancer cells, or macrophages (Figure 3).
Specific Therapeutic Applications of tFNAs in Lung Injury
Therapy for ALI
ALI and its severe form, ARDS, are rapidly progressive forms of pneumonia. In these conditions, the early inhibition of inflammation is necessary to prevent further disease progression. These pathological processes are closely related to the cytokine storm mediated by macrophages, which play a key role in the defense against pathogen invasion, clearance of cellular debris, and regulation of inflammatory responses in ALI.99 Upon activation, macrophages trigger the release of pro-inflammatory mediators within lung tissue, which in turn promote PMN infiltration and compromise the integrity of the endothelial barrier. This leads to increased pulmonary microvascular permeability, resulting in interstitial and alveolar edema, atelectasis, and hyaline membrane formation. This ultimately causes acute hypoxic respiratory dysfunction or respiratory failure. Recent studies have shown that tFNAs can regulate oxidative stress and macrophage-mediated inflammatory responses by inhibiting the phosphorylation of MAPK family proteins, including ERK1/2, JNK1/2/3, and p38.88
The widespread misuse of antibiotics has led to the emergence of drug-resistant pathogens.100 These pathogens resist the effect of drugs by forming biofilms and release hemolytic toxins into the host’s circulatory system while also damaging cellular membranes, significantly increasing membrane permeability. Furthermore, they impair macrophage activation, suppress host immune responses, and promote the recruitment and proliferation of myeloid-derived suppressor cells, thereby disrupting both innate and adaptive immunity. This immune evasion ultimately results in severe pathological damage and dysfunction in lung tissues.101,102 To overcome both the immunosuppressive effects and drug resistance mechanisms of these pathogens, researchers have developed biomimetic nano-delivery systems based on tFNAs, with simultaneously optimized antibacterial treatment and immune regulation.103 One such system was prepared by covalently loading the tFNA skeleton with tobramycin and coating it with a neutrophil–platelet-derived hybrid membrane to enable precise and targeted antibiotic delivery, toxin neutralization, as well as immune microenvironment modulation.103 The biomimetic nano-therapeutic significantly enhanced drug accumulation at the site of pulmonary infection through active targeting mechanisms facilitated by the biomimetic membrane. Simultaneously, the pattern recognition receptors on the membrane surface enabled the synchronous clearance of bacteria and their secreted toxins.
Inhibiting pathogenic microRNAs within the body is considered a key therapeutic strategy, as these regulatory molecules are involved in approximately 60% of intracellular protein expression processes. Meanwhile, given the sensitivity of DNA molecules to ROS, the use of exogenous DNA nanostructures to scavenge intracellular ROS has demonstrated functional efficacy. Yao et al developed a three-dimensional t-sponge carrier that integrates an antisense miR-155 aptamer and a ROS-scavenging module onto a tFNA platform, achieving dual-mode synergistic regulation through “base complementary trapping” and “redox-sensitive binding”.78 When delivered via nebulization to modulate the local immune microenvironment in ALI, the tFNAs facilitated the rapid penetration of microRNA inhibitors and enhanced pulmonary retention. This combination approach effectively reduced the intracellular levels of pathogenic miR-155 and ROS. The target protein of miR-155, suppressor of cytokine signaling 1 (SOCS-1), is a key regulatory factor that promotes the degradation of NF-κB. By adsorbing miR-155, which is highly expressed during inflammation, t-sponge reduces the inhibition of SOCS-1, thereby enabling the negative regulation of inflammation via the NF-κB pathway. This downregulation decreases the mRNA expression of LPS-induced ALI-related inflammatory factors (TNF-α, IL-6, IL-1β, and iNOS) and the chemokine CXCL1. Additionally, it promotes macrophage polarization toward the anti-inflammatory M2 phenotype, remodels the local immune microenvironment, and mitigates oxidative stress, thus alleviating lung injury.
Therapy for Pulmonary Fibrosis
The onset and progression of pulmonary fibrosis represent highly complex processes driven by epithelial cell injury due to inflammatory cytokines, oxidative stress, and other pathological factors. This process is further complicated by excessive EMT, FMT, and ECM accumulation, along with the activation of multiple associated signaling pathways. Zhang et al developed a pulmonary fibrosis cell model using alveolar epithelial cells derived from lung tissue and induced fibrosis using TGF-β1 stimulation to explore the therapeutic potential of tFNAs.96 Their findings indicated that tFNAs can downregulate the expression of critical fibrosis markers, including α-smooth muscle actin (α-SMA), collagen I (Col I), and fibronectin. This antifibrotic effect is achieved through the inhibition of ROS production and the downregulation of the Smad 2/Smad 3 signaling pathway, which plays a significant role in regulating EMT and ECM production. Collectively, these mechanisms contribute to the inhibition of fibrosis progression. Existing medications for fibrosis are primarily administered orally in the form of tablets or capsules. However, oral administration presents several limitations, including high dosage requirements, limited targeting capability, and no treatment precision. In contrast, tracheal aerosol delivery is a widely accepted and non-invasive route of administration. Nebulized inhalation can address the shortcomings of oral administration by reducing the dosage and frequency of administration. One study showed that the nebulized inhalation of drugs can increase the local concentration of PFD in the lungs and accelerate the rate of drug absorption.104,105 In a recent study, Xie et al delivered PFD via intratracheal nebulization using a Pt complex formed by loading PFD onto tFNAs.15 This strategy significantly improved the solubility and bioavailability of PFD, enabling targeted and quantitative delivery to the lungs. In vivo, the Pt complex enhanced immune regulation, exerted anti-inflammatory and antioxidant effects, and modulated the TGF-β1/Smad signaling pathway. These combined effects led to improved body weight recovery in pulmonary fibrosis mouse models, restored lung tissue architecture, and reduced mortality. In vitro studies further confirmed the antifibrotic efficacy of Pt, as evidenced by the downregulation of collagen I and α-SMA expression.15 Additionally, Pt inhibited excessive fibroblast activation and EMT in epithelial cells exposed to TGF-β1, while also suppressing the production of the pro-inflammatory factor NOD-like receptor thermal protein domain-associated protein 3 (NLRP3) and oxidative mediator NADPH oxidase-4 (NOX4).
In addition, Li et al found that the signaling pathway responsible for macrophage polarization, ie, the CCL2/CCR2 axis (including JAK/STAT, MAPK, and NF-κB signaling pathways), is activated in pulmonary fibrosis The activation of this axis promotes M1 macrophage-mediated myofibroblast activation and exacerbates alveolar inflammation. Given the central role of this signaling axis in macrophage migration, proliferation, and polarization, targeting the CCL2/CCR2 axis represents a promising therapeutic strategy.106 To this end, Li et al engineered a multi-adhesive terminal tFNA platform for the delivery of quadruple siCCR2 RNA in a mouse model of persistent pulmonary fibrosis mediated by pulmonary alveolar inflammation, induced by bleomycin and chest irradiation. By targeting the CCL2/CCR2 axis to deplete M1 macrophages and inhibit their recruitment to the lung parenchyma, the activation of fibroblasts was effectively suppressed, thereby mitigating fibrotic progression.105 Overall, these findings regarding tFNAs provide novel perspectives for the application of novel nucleic acid nanomaterials for the treatment of fibrotic diseases.
Therapy for Lung Cancer
In recent decades, targeted therapies and immunotherapies have significantly improved survival outcomes for patients with advanced lung cancer. However, the frequent emergence of drug resistance continues to compromise the long-term prognosis of these patients.107 Drug resistance refers to the capacity of tumor cells to activate various defense mechanisms that diminish or negate the cytotoxic and inhibitory effects of anticancer drugs. Paclitaxel (PTX), a widely used chemotherapeutic agent, promotes microtubule polymerization, disrupts cellular function, and induces apoptosis. However, its clinical efficacy is significantly limited by resistance.108 The major mechanisms underlying PTX resistance in tumors include decreased drug influx, enhanced drug efflux, activation of DNA repair pathways, and inhibition of apoptosis.109 o overcome these limitations, a PTX/tFNA drug delivery system was developed, in which PTX was loaded onto tFNAs.110 Studies have shown that tFNAs enhance the uptake of PTX by cancer cells and prevent drug efflux by exploiting caveolin-mediated endocytosis and exocytosis pathways. Moreover, the PTX/tFNA system downregulates the expression of multidrug resistance genes and P-glycoprotein, thereby limiting drug efflux, increasing cancer cell death, and reversing PTX resistance in lung cancer. In addition, the progression of lung cancer is heavily influenced by the tumor microenvironment (TME), a complex system of tumor cells, surrounding tissues, and immune cells that is characterized by hypoxia, immunosuppression, low pH, and inflammatory cell infiltration.111 Immune suppression is the primary factor hindering tumor elimination and has a direct impact on treatment efficacy. In a recent study, a novel inhalation strategy using nebulized tetrahedral DNA nanomachines was proposed. Subsequently, an inhalable pH-responsive tetrahedral DNA nanomachine was engineered for the treatment of metastatic lung cancer. This carrier incorporated immunomodulatory CpG oligonucleotides and antagonistic DNA aptamers that targeted programmed cell death ligand 1 (PD-L1).112 This carrier system was designed to release CpG and PD-L1 aptamers specifically in the acidic TME through the fine-tuning of their molar ratio, thereby blocking the PD-1/PD-L1 immune checkpoint axis. The CpG oligonucleotides were then internalized by antigen-presenting cells, triggering anti-tumor immune activation and cytokine secretion. Additionally, the use of the inhalation approach could bypass the lung mucosal barrier, significantly enhance drug accumulation within the tumor site, suppress tumor growth, and prolong survival in murine models. This study confirmed that genetically engineered tetrahedral DNA structures can effectively penetrate the lung mucus and tissue barriers to deliver immunotherapeutic agents directly to tumor tissues. Nonetheless, further optimization is required to achieve sustained therapeutic effects and improved clinical outcomes. A diagram of the mechanistic pathway of the tFNAs nano-platform for treating lung injury is shown in Figure 4.
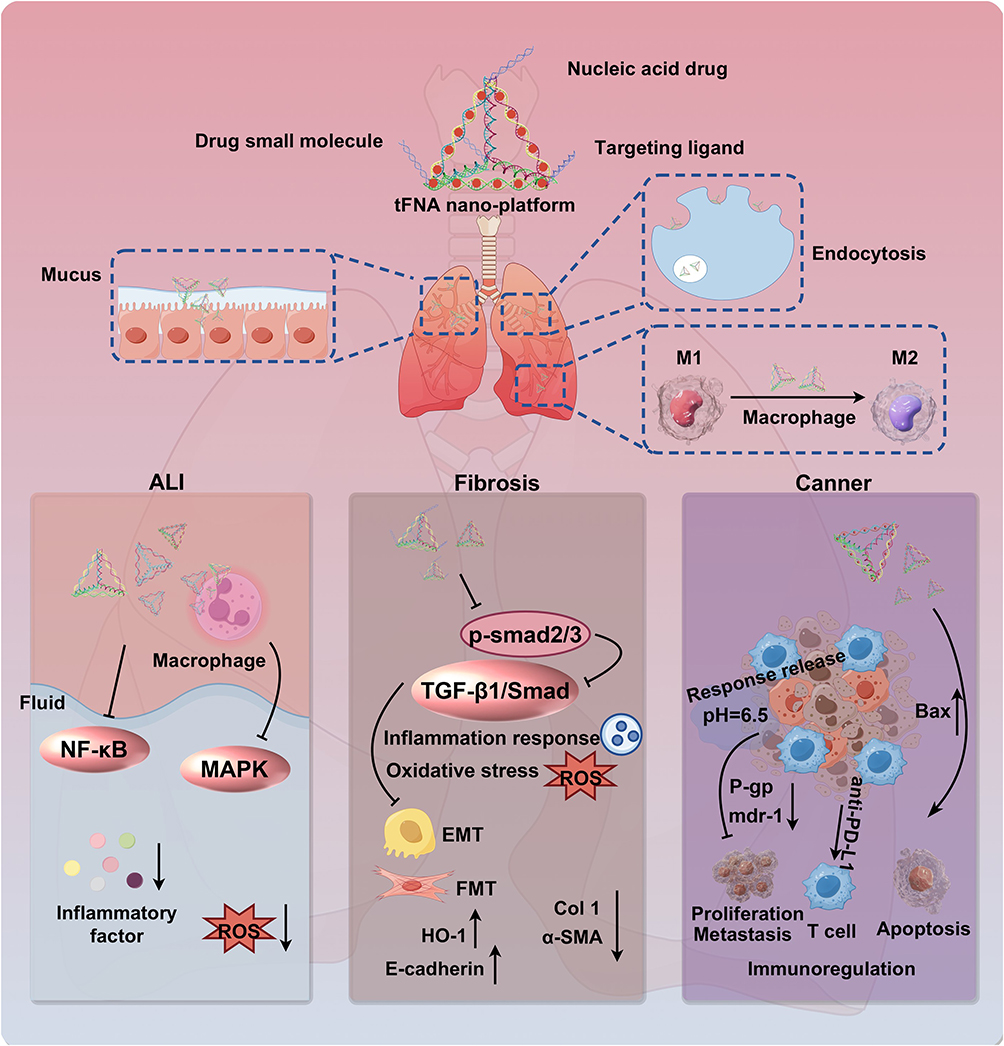 |
Figure 4 The tFNAs nano-platform penetrates the mucosal barrier for targeted, efficient drug delivery to treat lung injury. After the tFNAs-based nanoplatform penetrates the mucus barrier and is endocytosed by macrophages, it promotes macrophage polarization from the pro-inflammatory M1 to the reparative M2 phenotype. The platform simultaneously modulates key pathological pathways by inhibiting inflammation (NF-κB/MAPK), fibrosis (TGF-β1/Smad), ROS, and EMT to restore immune homeostasis and achieve integrated therapeutic efficacy. The black upward and downward arrows indicate upregulation and downregulation, respectively; the black T-arrows indicate inhibition. |
Beyond this, recent investigations have also revealed differences in the microbial composition of tumor tissues and normal tissues.113 Specific microorganisms have been found to preferentially accumulate at tumor sites, suggesting their potential utility as precision carriers for anticancer drugs. Building on this concept, a novel bacterial nanoplatform was developed to target tumor-associated biofilms and modulate the TME.114 This strategy involved modifying the anti-tumor aptamer AS1411 and the chemotherapeutic agent 5-fluorouracil, conjugating both to a single-stranded DNA-based tFNA, and using Streptococcus mutans as a biological carrier. This system enabled synergistic biofilm targeting and immune modulation, offering considerable potential for clinical translation. In the future, biomimetic nanocomposites based on bacterial carriers and tetrahedral DNA nanomedicines could be expanded to the treatment of a wide range of malignancies. Table 1 summarizes some preclinical studies on the use of tFNAs for treating lung injury, highlighting their design and therapeutic outcomes.
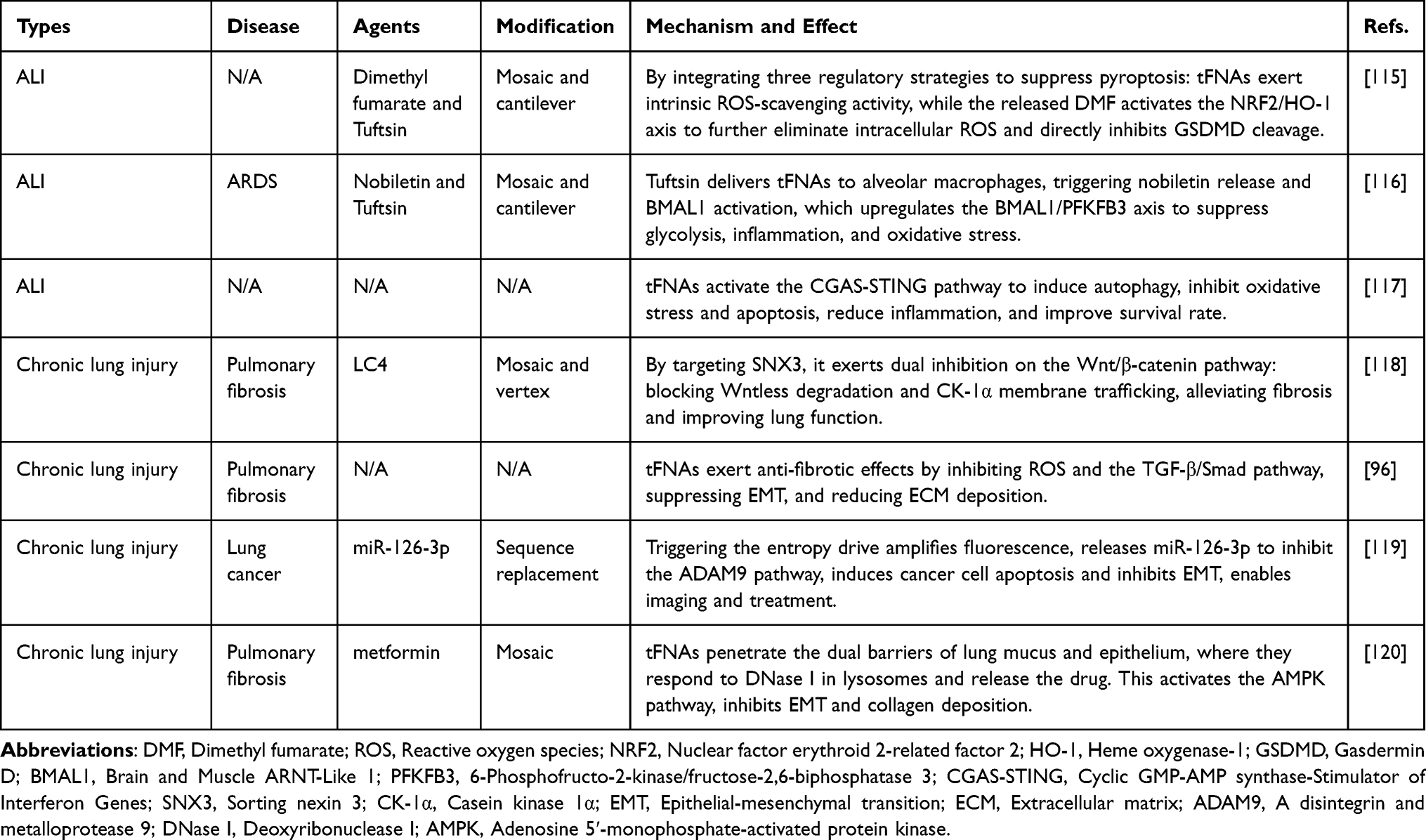 |
Table 1 Application and Mechanism of tFNAs for Treating Lung Injury |
Optimization and Clinical Challenges of tFNAs
With the rapid development of DNA nanotechnology and molecular biology, DNA nanocarriers and nucleic acid therapies have advanced considerably, demonstrating immense potential in the field of biomedicine. tFNAs, the simplest DNA polyhedral structures, have shown great biomedical potential. However, the clinical translation of these tFNAs is still associated with several challenges, and specific solutions are required to ensure their successful integration into medical practice.
Targeted Optimization
Due to their exceptional programmability, biocompatibility, and significant cellular uptake and tissue penetration capabilities, tFNAs have demonstrated significant potential as delivery platforms for various bioactive molecules. However, some limitations remain, including susceptibility to enzymatic degradation and insufficient ability to cross physiological barriers. Although tFNAs can increase the intracellular uptake of aptamers and small-molecule drugs, these agents are loaded onto the tFNA exoskeleton. The drug loading, biological stability, and off-target effects of these static structures under complex biological conditions thus remain unclear. For the delivery of small interfering RNA (siRNA) or microRNA, the “tail-like” structures at the apex can be modified using simple and convenient designs, but the sticky ends protruding from the exoskeleton are prone to degradation during transport.121 Additionally, off-target effects remain a major concern in many gene therapy applications.
The precise regulation of target genes using oligonucleotides has garnered increasing interest due to the remarkable therapeutic effects and enormous potential for improvement. However, achieving targeted delivery, enhancing functional performance, overcoming nuclease degradation, and ensuring successful lysosomal escape remain challenging.122 To overcome these obstacles, researchers have proposed an active strategy designed to optimize tFNA targeting, promote the stable encapsulation of DNA tetrahedral vectors, and enable controlled, stimulus-responsive release. Thus, a versatile platform tailored for diverse drug delivery requirements can be fabricated.123 In one study, siRNA was successfully encapsulated within dynamic lysosome-activated tFNA nanocapsules to achieve targeted, stimulus-responsive release.124 Thus, a versatile platform tailored for diverse drug delivery requirements can be fabricated. In one study, siRNA was successfully encapsulated within dynamic lysosome-activated tFNA nanocapsules to achieve targeted, stimulus-responsive release. This method involving complete encapsulation could prevent the premature dissociation of the drug–carrier complex at physiological temperatures, offer sufficient amounts of cargo securing force, protect siRNA against enzymes, and achieve pH-dependent release in target organelles. The targeting efficiency could be further improved through antibody-coupling strategies. For instance, doxorubicin (DOX), a potent chemotherapeutic agent, was embedded within a tFNA vector and coupled with the pro-apoptotic peptide D-(KLAKLAK)2 (KLA) to achieve mitochondrial targeting.125 In this system, DOX exerted cytotoxic effects against tumor cells, while KLA facilitated mitochondrial localization of DOX and activated intrinsic apoptotic pathways, enhancing anticancer efficacy. However, major transport barriers, including the blood–brain barrier and the plasma membrane, still limit the effective delivery of tFNAs. Although fluorescence imaging studies have confirmed that tFNAs enter cells primarily via caveolin-mediated endocytosis, this receptor-dependent pathway does not guarantee the efficient transport of DNA nanostructures across all barriers. Therefore, it remains critical to develop tFNA-based drug delivery systems capable of penetrating a range of different physiological barriers, increasing selective uptake by diseased tissues or cells, and reducing their uptake in healthy cells and tissues.126
Security Assessment
The safety and stability of tFNAs in vivo are critical to their biological applications and are key prerequisites for advancing their clinical translation. While mononucleotides are inherently biocompatible and biodegradable, their incorporation into DNA nanostructures may alter these properties. Although current evidence supports the good biocompatibility of tFNAs, the potential for long-term toxicity cannot be ignored. Previous studies have also demonstrated the safety of single-dose administration. However, comprehensive histopathological analysis after long-term, high-dose treatment, particularly in major organs such as the liver, kidneys, and spleen, is still lacking. Therefore, further investigation is still needed to fully assess the potential toxic effects of tFNAs. Recent research has provided important insights into the in vivo distribution, metabolism, and biosafety of tFNAs. In one study, tFNAs were labeled with the fluorescent dye Cy5 and their distribution in mice was tracked using in vivo imaging.127 Following the tail vein injection of Cy5-labeled tFNA, the fluorescence intensity increased in multiple organs in a time-dependent manner. Notably, strong signals were detected in the kidneys and brain during the early post-injection phase, confirming that tFNA accumulated rapidly in these tissues after entering the bloodstream. In addition, the fluorescence signal gradually shifted toward the bladder, suggesting that the tFNA was effectively processed and metabolized by the kidneys.128,129 Similarly, other studies have also demonstrated that tFNAs are excreted via the kidneys. This rapid clearance suggests that tFNAs can be effectively eliminated from the systemic circulation, thus reducing the risk of adverse reactions following long-term exposure. The findings suggest that the risk of tFNA accumulation in major organs is low, further supporting the safety of tFNAs. However, such clearance dynamics may also limit the sustained accumulation of tFNAs in target tissues, such as lung lesions, the available evidence is insufficient. In this context, a comprehensive analysis of the affected metabolic pathways and elimination mechanisms will be crucial to clarify the pharmacokinetic properties of tFNAs.
Furthermore, a detailed long-term evaluation of the immunomodulatory effects of tFNAs is also warranted. Despite being the simplest DNA nanostructures, tFNAs exhibit significantly greater structural complexity than conventional oligonucleotides. Their three-dimensional configuration and potential exogenous modifications can trigger immune responses as foreign entities, raising concerns about immunogenicity. Current immunological assessments largely rely on in vitro cellular models, while in vivo immune responses (such as antibody production and complement activation) remain inadequately explored. Recent studies suggest that tFNAs can partially activate the cGAS-STING pathway, a key inflammatory signaling cascade, and that the extent of activation correlates positively with the length of the DNA single strand.130 However, these findings have been validated only in vitro using a single cell type, and it remains unclear whether such activation translates into downstream inflammatory responses in vivo. In addition, factors such as the dosage, frequency, and route of tFNA administration must be carefully considered to optimize tFNA-based therapy while minimizing the risk of excessive immune activation. As such, a comprehensive evaluation of the immunostimulatory properties of tFNAs is essential before promoting their widespread clinical application.131
Clinical Translation Bottlenecks
Preclinical research on tFNAs has shown promising results in various disease models. However, it remains uncertain whether tFNAs can effectively transport therapeutic molecules that exceed their own size and molecular weight. Adding biomolecules on the vertices or arms of tFNAs may disrupt their inherent conformational flexibility, potentially compromising the stability, rigidity, and overall geometric structure of the assembled nanostructure.65 It is well established that the size, shape, and surface charge of DNA nanostructures influence their cellular uptake mechanisms, intracellular transport, and eventual destination. Further studies are needed to determine whether conjugation with complex biomolecular structures affects the internalization of tFNAs.132 Furthermore, current in vivo studies are primarily limited to murine models, and the therapeutic efficacy of tFNAs in humans remains to be further evaluated and verified. The transition from laboratory research to clinical application presents numerous challenges, the most important of which include production costs, scalability, consistency, and adherence to high manufacturing standards. For practical biomedical applications, it is essential to produce large quantities of high-purity functional DNA nanostructures. This places high demands on the reproducibility of the nanomaterial synthesis process and impacts systematic preclinical toxicity assessments and early-stage in vivo investigations. Although self-assembly technology has enabled the laboratory-scale fabrication of DNA nanostructures, issues such as low production yield, significant batch-to-batch variability, and high costs make scaling up challenging. These limitations create significant barriers in preventing the clinical translation of tFNAs. Addressing these problems is crucial to realize the full therapeutic potential of tFNAs for biomedical applications. Several convenient and cost-effective purification techniques for DNA nanostructures have been implemented at the laboratory level, including Bellot’s agarose gel electrophoresis-based separation method and Lin’s rate-zonal centrifugation technique for purifying DNA origami nanostructures via rate zone centrifugation. Additionally, methods such as PCR amplification and microbial fermentation have been explored for cost control and the mass production of DN.133,134 However, these approaches are yet to achieve reliable and consistent results in large-scale DNA production. To overcome these challenges, researchers have proposed the DELIVER framework, which provides a roadmap for clinical research on nanomedicine.135 This framework comprehensively covers all the steps from design to clinical application and aims to systematically guide the development, optimization, and adoption of nanomedicines and nucleic acid therapies in clinical practice.
Conclusion and Future Prospects
This article systematically reviews the research progress on tFNAs in lung injury repair. As a three-dimensional nanomaterial constructed using DNA nanotechnology, tFNAs can be used in clinical applications due to their unique tetrahedral structure, excellent biocompatibility, programmability, biodegradability, and high stability. Their unique ability to penetrate cell membranes and tissues without adjuvants makes them ideal candidates for drug delivery. They are small, can be distributed accurately, administered quickly and specifically, and have minimal impact on other organs. In specific application scenarios, tFNAs have been successfully used for anti-inflammatory immune regulation in ALI/ARDS, anti-fibrotic therapy in pulmonary fibrosis, and reversal of drug resistance and remodeling of the immune microenvironment in lung cancer, providing innovative strategies and a theoretical basis for the clinical treatment of lung injury. In summary, the biomedical applications of tFNAs are supported by two core aspects: (1) The unique physicochemical properties of DNA molecules, including sequence programmability, dynamic regulation of cell behavior, and efficient ROS scavenging, which collectively provide a versatile molecular toolbox for designing intelligent delivery systems; and (2) The spatial advantages conferred by their tetrahedral topology, which facilitate enhanced cellular uptake through geometric adaptability, improve deep tissue penetration, and enable modular assembly for functional integration in complex biological environments.
Although tFNAs have demonstrated notable efficacy in the treatment of lung injuries, their therapeutic potential extends well beyond this domain. With the rapid advancement of DNA nanotechnology and a growing understanding of lung injury mechanisms, future research should aim to explore and ensure optimization in several key areas: (1) Standardized manufacturing: The fabrication of tFNAs involves a complex self-assembly mechanism and stringent purity standards. Achieving stable, efficient, and large-scale production will necessitate breakthroughs in synthesis technologies and process innovations. (2) Engineering of intelligent carriers: tFNAs are programmable and can respond to the environment. By leveraging physiological stimuli such as pH, ROS, temperature, specific oligonucleotides, or adenosine triphosphate (ATP) levels, the spatiotemporally dynamic and controllable release of drug molecules can be achieved for lung injury treatment, and the accuracy of drug delivery and treatment specificity can be improved. (3) Integration of artificial intelligence (AI): Continued advancements in AI algorithms, combined with improvements in DNA origami techniques, offer the potential to tailor tFNA-based therapies to individual patients. AI can not only be used to analyze the characteristics of individual lesions (through imaging and biomarker data) but also to design optimized DNA sequences and generate standardized databases, thereby expanding the capabilities of tFNAs for targeted disease treatment. (4) In-depth mechanistic analysis: Using spatial transcriptomics and single-cell multi-omics, future studies should explore the dynamic transcriptional reprogramming of immune cells, epithelial cells, and interstitial cells in lung tissue before and after tFNAs treatment. This could help elucidate key pathways involved in immune reprogramming and the reversal of lung injury pathology. (5) Optimization of research models: Although DNA-based drug delivery systems are not yet in clinical use, the field is advancing rapidly. Establishing lung injury models that more closely resemble human pathology (eg, organoids and humanized mouse models) and evaluating the differential effects of tFNAs at various stages of disease could be instrumental in accelerating the clinical translation of tFNAs (Figure 5). If significant advancements are achieved in these areas, tFNAs could emerge as a novel paradigm for nanodrug therapy in cases of lung injury treatment and offer a scalable framework for addressing chronic inflammation, fibrosis, and malignancies in other organ systems.
 |
Figure 5 Optimization and exploration of a nano-drug-loading platform for lung injury based on tFNAs in the future. This future research strategy foresees an AI-driven approach to optimize lung injury models and tFNAs-based platforms. It integrates standardized tFNAs manufacturing with intelligent, environmentally responsive carrier engineering (eg, responsive to pH, ROS, ATP) to enable targeted and intelligent therapy. |
Abbreviations
ALI, Acute lung injury; ARDS, Acute respiratory distress syndrome; AMPK, Adenosine 5′-monophosphate-activated protein kinase; ADAM9, A disintegrin and metalloprotease 9; ATP, Adenine triphosphate; α-SMA, α-smooth muscle actin; AI, Artificial intelligence; BMAL1, Brain and Muscle ARNT-Like 1; Col 1, Collagen I; CGAS-STING, Cyclic GMP-AMP synthase-Stimulator of Interferon Genes; CK-1α, Casein kinase 1α; DNA, Deoxyribonucleic acid; DOX, Doxorubicin; DMF, Dimethyl fumarate; DNase I, Deoxyribonuclease I; ECM, Extracellular matrix; EMT, Epithelial-mesenchymal transition; ERK, Extracellular signal regulated kinases; FMT, Fibroblast to Myofibroblast transition; GSDMD, Gasdermin D; HO-1, Heme oxygenase-1; IL-1, Interleukin-1; IL-4, Interleukin-4; IL-8, Interleukin-8; IL-13, Interleukin-13; IFN-γ, Interferon-γ; iNOS, Inducible nitric oxide synthase; IL-1β, Interleukin-1β; IL-6, Interleukin-6; ICU, Intensive Care Unit; JNK, c-Jun N-terminal kinase; LPS, Lipopolysaccharide; MAPK, Mitogen-activated protein kinase; NF-κB, Nuclear factor κ light chain enhancer; NLRP3, NOD-like receptor thermal protein domain associated protein 3; NOX4, NADPH oxidase-4; NRF2, Nuclear factor erythroid 2-related factor 2; PMNs, Polymorphonuclear leukocytes; PFD, Pirfenidone; PGE2, Prostaglandin E2; PTX, Paclitaxel; PD-L1, Programmed cell death ligand 1; PFKFB3, 6-Phosphofructo-2-kinase/fructose-2,6-biphosphatase 3; RILI, Radiation-induced lung injury; ROS, Reactive oxygen species; siRNA, Small interfering RNA; SNX3, Sorting nexin 3; tFNA, Tetrahedral framework nucleic acid; TNF-α, Tumour necrosis factor-alpha; TGF-β1, Transforming growth factor-β1; TME, Tumor microenvironment; VEGF, Vascular endothelial growth factor.
Acknowledgment
We thank Figdraw for their assistance in creating the figures. We thank Home for Researchers editorial team (https://www.home-for-researchers.com) for language editing service.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Shanghai Municipal Health Commission Project (2024QN057), Military Pharmacy Key Discipline Specialty Group Construction Project (2024-HJZDXK-JS-12, 2024-HJZDXK-SZ-01), Sailing Talent Program of Naval Medical University.
Disclosure
The authors report there are no conflicts of interest in this work.
References
1. Zhang ZY, Zhu DW, Cui B, et al. Association between particulate matter air pollution and lung cancer. Thorax. 2020;75(1):85–24. doi:10.1136/thoraxjnl-2019-213722
2. Kallet RH, Zhuo HJ, Ho K, et al. Lung injury etiology and other factors influencing the relationship between dead-space fraction and mortality in ARDS. Respir Care. 2017;62(10):1241–1248. doi:10.4187/respcare.05589
3. Zhao C, Pu WY, Wazir J, et al. Long-term exposure to PM2.5 aggravates pulmonary fibrosis and acute lung injury by disrupting Nrf2-mediated antioxidant function. Environ Pollut. 2022;313:120017. doi:10.1016/j.envpol.2022.120017
4. Liu HQ, Dong JL, Xu CL, et al. Acute lung injury: pathogenesis and treatment. J Transl Med. 2025;23:926. doi:10.1186/s12967-025-06994-2
5. Liu YM, Meng JY, Liu J, et al. NLRP3 inflammasome in acute lung injury: cumulative evidence for traditional Chinese medicine and natural products. Chin Herb Med. 2025;17(4):703–719. doi:10.1016/j.chmed.2025.06.002
6. Khemani RG, Smith L, Lopez-Fernandez YM, et al. Paediatric acute respiratory distress syndrome incidence and epidemiology (PARDIE): an international, observational study. Lancet Respir Med. 2019;7(2):115–128. doi:10.1016/s2213-2600(18)30344-8
7. Bellani G, Laffey JG, Pham T, et al. Epidemiology, patterns of care, and mortality for patients with acute respiratory distress syndrome in intensive care units in 50 countries. JAMA. 2016;315(8):788–800. doi:10.1001/jama.2016.0291
8. Dowdy DW, Bienvenu OJ, Dinglas VD, et al. Are intensive care factors associated with depressive symptoms 6 months after acute lung injury? Crit Care Med. 2009;37(5):1702–1707. doi:10.1097/CCM.0b013e31819fea55
9. Bos LDJ, Ware LB. Acute respiratory distress syndrome: causes, pathophysiology, and phenotypes. Lancet. 2022;400(10358):1145–1156. doi:10.1016/s0140-6736(22)01485-4
10. Cha MJ, Solomon JJ, Lee JE, et al. Chronic lung injury after COVID-19 pneumonia: clinical, radiologic, and histopathologic perspectives. Radiology. 2024;310(1):e231643. doi:10.1148/radiol.231643
11. Seeman NC. Nucleic acid junctions and lattices. J Theor Biol. 1982;99(2):237–247. doi:10.1016/0022-5193(82)90002-9
12. Goodman RP, Schaap IA, Tardin CF, et al. Rapid chiral assembly of rigid DNA building blocks for molecular nanofabrication. Science. 2005;310(5754):1661–1665. doi:10.1126/science.1120367
13. Yu MC, Zhang YY, Zhang MX, et al. Enzymatically cyclic activated biosensor based on a Tetrahedral DNA framework for precise tumor in situ molecular imaging. ACS Sens. 2024;9(10):5302–5311. doi:10.1021/acssensors.4c01493
14. Zhu FL, Bian XJ, Zhang HC, et al. Controllable design of a nano-bio aptasensing interface based on tetrahedral framework nucleic acids in an integrated microfluidic platform. Biosens Bioelectron. 2021;176:112943. doi:10.1016/j.bios.2020.112943
15. Xie YT, Shi SR, Lv WT, et al. Tetrahedral framework nucleic acids delivery of pirfenidone for anti-inflammatory and antioxidative effects to treat idiopathic pulmonary fibrosis. ACS Nano. 2024;18(39):26704–26721. doi:10.1021/acsnano.4c06598
16. Li YF, Li JF, Xu S, et al. Tetrahedral framework nucleic acid-based delivery of astaxanthin suppresses chondrocyte pyroptosis and modulates oxidative stress for the treatment of osteoarthritis. Adv Healthcare Mater. 2024;13(28):e2401452. doi:10.1002/adhm.202401452
17. He JJ, Chen W, Chen XY, et al. Tetrahedral framework nucleic acid loaded with glabridin: a transdermal delivery system applicated to anti-hyperpigmentation. Cell Prolif. 2023;56(12):e13495. doi:10.1111/cpr.13495
18. Li PX, Fu LW, Liao ZY, et al. Chitosan hydrogel/3D-printed poly(ε-caprolactone) hybrid scaffold containing synovial mesenchymal stem cells for cartilage regeneration based on tetrahedral framework nucleic acid recruitment. Biomaterials. 2021;278:121131. doi:10.1016/j.biomaterials.2021.121131
19. Zou WQ, Lu J, Zhang LY, et al. Tetrahedral framework nucleic acids for improving wound healing. J Nanobiotechnol. 2024;22(1):113. doi:10.1186/s12951-024-02365-z
20. Tan L, Xie JZ, Liao CQ, et al. Tetrahedral framework nucleic acids inhibit Aβ-mediated ferroptosis and ameliorate cognitive and synaptic impairments in Alzheimer’s disease. J Nanobiotechnol. 2024;22(1):682. doi:10.1186/s12951-024-02963-x
21. Zhu JW, Yang YT, Ma WJ, et al. Antiepilepticus effects of tetrahedral framework nucleic acid via inhibition of gliosis-induced downregulation of glutamine synthetase and increased AMPAR internalization in the postsynaptic membrane. Nano Lett. 2022;22(6):2381–2390. doi:10.1021/acs.nanolett.2c00025
22. Gutierrez AM, Frazar EM, Klaus MVX, et al. Hydrogels and hydrogel nanocomposites: enhancing healthcare through human and environmental treatment. Adv Healthcare Mater. 2022;11(7):e2101820. doi:10.1002/adhm.202101820
23. Millar MW, Fazal F, Rahman A. Therapeutic targeting of NF-κB in acute lung injury: a double-edged sword. Cells. 2022;11(20):3317. doi:10.3390/cells11203317
24. Johnson ER, Matthay MA. Acute lung injury: epidemiology, pathogenesis, and treatment. J Aerosol Med Pulm Drug Deliv. 2010;23(4):243–252. doi:10.1089/jamp.2009.0775
25. Zhang H, Kang J, Guo WY, et al. An optimal medicinal and edible Chinese herbal formula attenuates particulate matter-induced lung injury through its anti-oxidative, anti-inflammatory and anti-apoptosis activities. Chin Herb Med. 2023;15(3):407–420. doi:10.1016/j.chmed.2022.08.003
26. Li N, Zou SS, Wang B, et al. Targeting immunometabolism against acute lung injury. Clin Immunol. 2023;249:109289. doi:10.1016/j.clim.2023.109289
27. Hu LY, Wang BL, Jiang Y, et al. Risk factors for transfusion-related acute lung injury. Respir Care. 2021;66(6):1029–1038. doi:10.4187/respcare.08829
28. He YQ, Zhou CC, Yu LY, et al. Natural product derived phytochemicals in managing acute lung injury by multiple mechanisms. Pharmacol Res. 2021;163:105224. doi:10.1016/j.phrs.2020.105224
29. Ranieri VM, Rubenfeld GD, Thompson BT, et al. Acute respiratory distress syndrome: the Berlin Definition. JAMA. 2012;307(23):2526–2533. doi:10.1001/jama.2012.5669
30. Matute-Bello G, Frevert CW, Martin TR. Animal models of acute lung injury. Am J Physiol Lung Cell Mol Physiol. 2008;295(3):L379–399. doi:10.1152/ajplung.00010.2008
31. Savin IA, Zenkova MA, Sen’kova AV. Pulmonary fibrosis as a result of acute lung inflammation: molecular mechanisms, relevant in vivo models, prognostic and therapeutic approaches. Int J Mol Sci. 2022;23(23):14959. doi:10.3390/ijms232314959
32. Mokra D, Mikolka P, Kosutova P, et al. Corticosteroids in acute lung injury: the dilemma continues. Int J Mol Sci. 2019;20(19):4765. doi:10.3390/ijms20194765
33. Heukels P, Moor CC, von der Thüsen JH, et al. Inflammation and immunity in IPF pathogenesis and treatment. Respir Med. 2019;147:79–91. doi:10.1016/j.rmed.2018.12.015
34. Upagupta C, Shimbori C, Alsilmi R, et al. Matrix abnormalities in pulmonary fibrosis. Eur Respir Rev. 2018;27(148):180033. doi:10.1183/16000617.0033-2018
35. Ju XT, Wang K, Wang CJ, et al. Regulation of myofibroblast dedifferentiation in pulmonary fibrosis. Respir Res. 2024;25(1):284. doi:10.1186/s12931-024-02898-9
36. Strippoli R, Moreno-Vicente R, Battistelli C, et al. Molecular mechanisms underlying peritoneal EMT and fibrosis. Stem Cells Int. 2016;2016:3543678. doi:10.1155/2016/3543678
37. Mayr CH, Sengupta A, Asgharpour S, et al. Sfrp1 inhibits lung fibroblast invasion during transition to injury-induced myofibroblasts. Eur Respir J. 2024;63(2):2301326. doi:10.1183/13993003.01326-2023
38. Hinz B. Formation and function of the myofibroblast during tissue repair. J Invest Dermatol. 2007;127(3):526–537. doi:10.1038/sj.jid.5700613
39. Liu F, Lagares D, Choi KM, et al. Mechanosignaling through YAP and TAZ drives fibroblast activation and fibrosis. Am J Physiol Lung Cell Mol Physiol. 2015;308(4):L344–357. doi:10.1152/ajplung.00300.2014
40. Richeldi L, Rubin AS, Avdeev S, et al. Idiopathic pulmonary fibrosis in BRIC countries: the cases of Brazil, Russia, India, and China. BMC Med. 2015;13:237. doi:10.1186/s12916-015-0495-0
41. Varone F, Sgalla G, Iovene B, et al. Nintedanib for the treatment of idiopathic pulmonary fibrosis. Expert Opin Pharmacother. 2018;19(2):167–175. doi:10.1080/14656566.2018.1425681
42. Meyer KC, Decker CA. Role of pirfenidone in the management of pulmonary fibrosis. Ther Clin Risk Manag. 2017;13:427–437. doi:10.2147/tcrm.S81141
43. Arroyo-Hernández M, Maldonado F, Lozano-Ruiz F, et al. Radiation-induced lung injury: current evidence. BMC Pulm Med. 2021;21(1):9. doi:10.1186/s12890-020-01376-4
44. Hanania AN, Mainwaring W, Ghebre YT, et al. Radiation-induced lung injury: assessment and management. Chest. 2019;156(1):150–162. doi:10.1016/j.chest.2019.03.033
45. Giuranno L, Ient J, De Ruysscher D, et al. Radiation-Induced Lung Injury (RILI). Front Oncol. 2019;9:877. doi:10.3389/fonc.2019.00877
46. Beach TA, Finkelstein JN, Chang PY. Epithelial Responses in Radiation-Induced Lung Injury (RILI) allow chronic inflammation and fibrogenesis. Radiat Res. 2023;199(5):439–451. doi:10.1667/rade-22-00130.1
47. Roy S, Salerno KE, Citrin DE. Biology of radiation-induced lung injury. Semin Radiat Oncol. 2021;31(2):155–161. doi:10.1016/j.semradonc.2020.11.006
48. Wang P, Yan ZY, Zhou PK, et al. The promising therapeutic approaches for radiation-induced pulmonary fibrosis: targeting radiation-induced mesenchymal transition of alveolar Type II epithelial cells. Int J Mol Sci. 2022;23(23):15014. doi:10.3390/ijms232315014
49. Freeman B, Mamallapalli J, Bian T, et al. Opportunities and challenges of kava in lung cancer prevention. Int J Mol Sci. 2023;24(11):9539. doi:10.3390/ijms24119539
50. Zochbauer-Muller S, Gazdar AF, Minna JD. Molecular pathogenesis of lung cancer. Annu Rev Physiol. 2002;64:681–708. doi:10.1146/annurev.physiol.64.081501.155828
51. Smolarz B, Łukasiewicz H, Samulak D, et al. Lung cancer-epidemiology, pathogenesis, treatment and molecular aspect (Review of Literature). Int J Mol Sci. 2025;26(5):2049. doi:10.3390/ijms26052049
52. Zhao YS, Guo SP, Deng J, et al. VEGF/VEGFR-targeted therapy and immunotherapy in non-small cell lung cancer: targeting the tumor microenvironment. Int J Biol Sci. 2022;18(9):3845–3858. doi:10.7150/ijbs.70958
53. Frezzetti D, Gallo M, Maiello MR, et al. VEGF as a potential target in lung cancer. Expert Opin Ther Targets. 2017;21(10):959–966. doi:10.1080/14728222.2017.1371137
54. Chen SY, Wang MN, Lu TQ, et al. JMJD6 in tumor-associated macrophage regulates macrophage polarization and cancer progression via STAT3/IL-10 axis. Oncogene. 2023;42(37):2737–2750. doi:10.1038/s41388-023-02781-9
55. Zhu XJ, Yu ZJ, Feng LB, et al. Chitosan-based nanoparticle co-delivery of docetaxel and curcumin ameliorates anti-tumor chemoimmunotherapy in lung cancer. Carbohydr Polym. 2021;268:118237. doi:10.1016/j.carbpol.2021.118237
56. Zhang YY, Ai LS, Gong YS, et al. Preparation and usage of nanomaterials in biomedicine. Biotechnol Bioeng. 2023;120(10):2777–2792. doi:10.1002/bit.28472
57. Kheirandish A, Sepehri Javan N, Mohammadzadeh H. Modified Drude model for small gold nanoparticles surface plasmon resonance based on the role of classical confinement. Sci Rep. 2020;10(1):6517. doi:10.1038/s41598-020-63066-9
58. Rothemund PW. Folding DNA to create nanoscale shapes and patterns. Nature. 2006;440(7082):297–302. doi:10.1038/nature04586
59. Chen JH, Seeman NC. Synthesis from DNA of a molecule with the connectivity of a cube. Nature. 1991;350(6319):631–633. doi:10.1038/350631a0
60. Zadeh JN, Steenberg CD, Bois JS, et al. NUPACK: analysis and design of nucleic acid systems. J Comput Chem. 2011;32(1):170–173. doi:10.1002/jcc.21596
61. Douglas SM, Marblestone AH, Teerapittayanon S, et al. Rapid prototyping of 3D DNA-origami shapes with caDNAno. Nucleic Acids Res. 2009;37(15):5001–5006. doi:10.1093/nar/gkp436
62. He Y, Ye T, Su M, et al. Hierarchical self-assembly of DNA into symmetric supramolecular polyhedra. Nature. 2008;452(7184):198–201. doi:10.1038/nature06597
63. Wang AZ, Langer R, Farokhzad OC. Nanoparticle delivery of cancer drugs. Annu Rev Med. 2012;63:185–198. doi:10.1146/annurev-med-040210-162544
64. Allen TM, Mumbengegwi DR, Charrois GJ. Anti-CD19-targeted liposomal doxorubicin improves the therapeutic efficacy in murine B-cell lymphoma and ameliorates the toxicity of liposomes with varying drug release rates. Clin Cancer Res. 2005;11(9):3567–3573. doi:10.1158/1078-0432.Ccr-04-2517
65. Zhang T, Tian TR, Zhou RH, et al. Design, fabrication and applications of tetrahedral DNA nanostructure-based multifunctional complexes in drug delivery and biomedical treatment. Nat Protoc. 2020;15(8):2728–2757. doi:10.1038/s41596-020-0355-z
66. Zhang XL, Liu NX, Zhou M, et al. The application of tetrahedral framework nucleic acids as a drug carrier in biomedicine fields. Curr Stem Cell Res Ther. 2021;16(1):48–56. doi:10.2174/1574888x15666200422103415
67. Yao YX, Wen YT, Li YJ, et al. Tetrahedral framework nucleic acids facilitate neurorestoration of facial nerves by activating the NGF/PI3K/AKT pathway. Nanoscale. 2021;13(37):15598–15610. doi:10.1039/d1nr04619e
68. Shao XR, Lin SY, Peng Q, et al. Tetrahedral DNA nanostructure: a potential promoter for cartilage tissue regeneration via regulating chondrocyte phenotype and proliferation. Small. 2017;13(12):1602770. doi:10.1002/smll.201602770
69. Vilcapoma J, Patel A, Chandrasekaran AR, et al. The role of size in biostability of DNA tetrahedra. Chem Commun. 2023;59(34):5083–5085. doi:10.1039/d3cc01123b
70. Keum JW, Bermudez H. Enhanced resistance of DNA nanostructures to enzymatic digestion. Chem Commun. 2009;(45):7036–7038. doi:10.1039/b917661f
71. Carter MLJ, Rusling DA, Gurr S, et al. Stability of the different arms of a DNA tetrahedron and its interaction with a minor groove ligand. Biophys Chem. 2020;256:106270. doi:10.1016/j.bpc.2019.106270
72. Walsh AS, Yin H, Erben CM, et al. DNA cage delivery to mammalian cells. ACS Nano. 2011;5(7):5427–5432. doi:10.1021/nn2005574
73. Vindigni G, Raniolo S, Ottaviani A, et al. Receptor-mediated entry of pristine octahedral DNA nanocages in mammalian cells. ACS Nano. 2016;10(6):5971–5979. doi:10.1021/acsnano.6b01402
74. Zagorovsky K, Chou LY, Chan WC. Controlling DNA-nanoparticle serum interactions. Proc Natl Acad Sci U S A. 2016;113(48):13600–13605. doi:10.1073/pnas.1610028113
75. Wang ZG, Xue QW, Tian WZ, et al. Quantitative detection of single DNA molecules on DNA tetrahedron decorated substrates. Chem Commun. 2012;48(77):9661–9663. doi:10.1039/c2cc35208g
76. Schlapak R, Danzberger J, Armitage D, et al. Nanoscale DNA tetrahedra improve biomolecular recognition on patterned surfaces. Small. 2012;8(1):89–97. doi:10.1002/smll.201101576
77. Zhao HP, Li MQ, Lu SS, et al. The enhancement of enzyme cascading via tetrahedral DNA framework modification. Analyst. 2023;148(4):906–911. doi:10.1039/d2an02097a
78. Yao L, Zhang GR, Wang Y, et al. Development of an inhalable DNA tetrahedron MicroRNA sponge. Adv Mater. 2025;37(4):e2414336. doi:10.1002/adma.202414336
79. Zhang J, Guo YY, Ding F, et al. A camptothecin-grafted DNA tetrahedron as a precise nanomedicine to inhibit tumor growth. Angew Chem Int Ed Engl. 2019;58(39):13794–13798. doi:10.1002/anie.201907380
80. Dykman L, Khlebtsov B, Khlebtsov N. Drug delivery using gold nanoparticles. Adv Drug Deliv Rev. 2025;216:115481. doi:10.1016/j.addr.2024.115481
81. Wang Y, Lu XH, Wu XH, et al. Chemically modified DNA nanostructures for drug delivery. Innovation. 2022;3(2):100217. doi:10.1016/j.xinn.2022.100217
82. Liang L, Li J, Li Q, et al. Single-particle tracking and modulation of cell entry pathways of a tetrahedral DNA nanostructure in live cells. Angew Chem Int Ed Engl. 2014;53(30):7745–7750. doi:10.1002/anie.201403236
83. Lim DY, Hwang BH. Aptamer-modified tetrahedral DNA nanostructure-immobilized liposome for specific gene delivery and potential cancer theragnostic. Biotechnol J. 2024;19(1):e2300156. doi:10.1002/biot.202300156
84. Lee H, Lytton-Jean AK, Chen Y, et al. Molecularly self-assembled nucleic acid nanoparticles for targeted in vivo siRNA delivery. Nat Nanotechnol. 2012;7(6):389–393. doi:10.1038/nnano.2012.73
85. Zhang YX, Ma WJ, Zhu Y, et al. Inhibiting methicillin-resistant staphylococcus aureus by tetrahedral DNA nanostructure-enabled antisense peptide nucleic acid delivery. Nano Lett. 2018;18(9):5652–5659. doi:10.1021/acs.nanolett.8b02166
86. Zhang T, Cui WT, Tian TR, et al. Progress in biomedical applications of tetrahedral framework nucleic acid-based functional systems. ACS Appl Mater Interfaces. 2020;12(42):47115–47126. doi:10.1021/acsami.0c13806
87. Han RF, Xiao Y, Bai QQ, et al. Self-therapeutic metal-based nanoparticles for treating inflammatory diseases. Acta Pharm Sin B. 2023;13(5):1847–1865. doi:10.1016/j.apsb.2022.07.009
88. Zhang Q, Lin SY, Shi SR, et al. Anti-inflammatory and antioxidative effects of tetrahedral DNA nanostructures via the modulation of macrophage responses. ACS Appl Mater Interfaces. 2018;10(4):3421–3430. doi:10.1021/acsami.7b17928
89. Zhou M, Tang YL, Lu YF, et al. Framework nucleic acid-based and neutrophil-based nanoplatform loading baicalin with targeted drug delivery for anti-inflammation treatment. ACS Nano. 2025;19(3):3455–3469. doi:10.1021/acsnano.4c12917
90. Su L, Zhang WY, Wu XL, et al. Glycocalyx-mimicking nanoparticles for stimulation and polarization of macrophages via specific interactions. Small. 2015;11(33):4191–4200. doi:10.1002/smll.201403838
91. Liu XY, Yu ZY, Wu Y, et al. The immune regulatory effects of tetrahedral framework nucleic acid on human T cells via the mitogen-activated protein kinase pathway. Cell Prolif. 2021;54(8):e13084. doi:10.1111/cpr.13084
92. Chen XY, Xu Z, Gao Y, et al. Framework nucleic acid-based selective cell catcher for endogenous stem cell recruitment. Adv Mater. 2024;36(50):e2406118. doi:10.1002/adma.202406118
93. Anathy V, Roberson E, Cunniff B, et al. Oxidative processing of latent Fas in the endoplasmic reticulum controls the strength of apoptosis. Mol Cell Biol. 2012;32(17):3464–3478. doi:10.1128/mcb.00125-12
94. Zhu YJ, Shi RJH, Lu WT, et al. Framework nucleic acids as promising reactive oxygen species scavengers for anti-inflammatory therapy. Nanoscale. 2024;16(15):7363–7377. doi:10.1039/d3nr05844a
95. Zhang T, Tian TR, Lin YF. Functionalizing framework nucleic-acid-based nanostructures for biomedical application. Adv Mater. 2022;34(46):e2107820. doi:10.1002/adma.202107820
96. Zhang TX, Gao Y, Xiao DX, et al. Nucleic acid based tetrahedral framework DNA nanostructures for fibrotic diseases therapy. Appl Mater Today. 2020;20:100725. doi:10.1016/j.apmt.2020.100725
97. Dou YK, Cui WT, Yang X, et al. Applications of tetrahedral DNA nanostructures in wound repair and tissue regeneration. Burns Trauma. 2022;10:tkac006. doi:10.1093/burnst/tkac006
98. Peng Q, Shao XR, Xie J, et al. Understanding the biomedical effects of the self-assembled tetrahedral DNA nanostructure on living cells. ACS Appl Mater Interfaces. 2016;8(20):12733–12739. doi:10.1021/acsami.6b03786
99. Malainou C, Abdin SM, Lachmann N, et al. Alveolar macrophages in tissue homeostasis, inflammation, and infection: evolving concepts of therapeutic targeting. J Clin Invest. 2023;133(19):e170501. doi:10.1172/jci170501
100. Flemming HC, Wingender J, Szewzyk U, et al. Biofilms: an emergent form of bacterial life. Nat Rev Microbiol. 2016;14(9):563–575. doi:10.1038/nrmicro.2016.94
101. Esteban-Fernández de Ávila B, Angsantikul P, Ramírez-Herrera DE, et al. Hybrid biomembrane-functionalized nanorobots for concurrent removal of pathogenic bacteria and toxins. Sci Robot. 2018;3(18):eaat0485. doi:10.1126/scirobotics.aat0485
102. Zhao KL, Li WJ, Li J, et al. TesG is a type I secretion effector of Pseudomonas aeruginosa that suppresses the host immune response during chronic infection. Nat Microbiol. 2019;4(3):459–469. doi:10.1038/s41564-018-0322-4
103. Zhang SH, Chen TY, Lu WT, et al. Hybrid cell membrane-engineered nanocarrier for triple-action strategy to address pseudomonas aeruginosa infection. Adv Sci. 2025;12(6):e2411261. doi:10.1002/advs.202411261
104. Weng CM, Li GH, Zhang DD, et al. Nanoscale porphyrin metal-organic frameworks deliver siRNA for alleviating early pulmonary fibrosis in acute lung injury. Front Bioeng Biotechnol. 2022;10:939312. doi:10.3389/fbioe.2022.939312
105. Tian XD, Bera H, Guo X, et al. Pulmonary delivery of reactive oxygen species/glutathione-responsive paclitaxel dimeric nanoparticles improved therapeutic indices against metastatic lung cancer. ACS Appl Mater Interfaces. 2021;13(48):56858–56872. doi:10.1021/acsami.1c16351
106. Li C, Feng XR, Li SH, et al. Tetrahedral DNA loaded siCCR2 restrains M1 macrophage polarization to ameliorate pulmonary fibrosis in chemoradiation-induced murine model. Mol Ther. 2024;32(3):766–782. doi:10.1016/j.ymthe.2024.01.022
107. Karasarides M, Cogdill AP, Robbins PB, et al. Hallmarks of Resistance to Immune-Checkpoint Inhibitors. Cancer Immunol Res. 2022;10(4):372–383. doi:10.1158/2326-6066.Cir-20-0586
108. Teow HM, Zhou ZY, Najlah M, et al. Delivery of paclitaxel across cellular barriers using a dendrimer-based nanocarrier. Int J Pharm. 2013;441(1–2):701–711. doi:10.1016/j.ijpharm.2012.10.024
109. Gottesman MM, Fojo T, Bates SE. Multidrug resistance in cancer: role of ATP-dependent transporters. Nat Rev Cancer. 2002;2(1):48–58. doi:10.1038/nrc706
110. Xie X, Shao X, Ma W, et al. Overcoming drug-resistant lung cancer by paclitaxel loaded tetrahedral DNA nanostructures. Nanoscale. 2018;10(12):5457–5465. doi:10.1039/c7nr09692e
111. Wu JY, Chen J, Feng YJ, et al. Tumor microenvironment as the “regulator” and “target” for gene therapy. J Gene Med. 2019;21(7):e3088. doi:10.1002/jgm.3088
112. Fan Q, Li ZH, Yin J, et al. Inhalable pH-responsive DNA tetrahedron nanoplatform for boosting anti-tumor immune responses against metastatic lung cancer. Biomaterials. 2023;301:122283. doi:10.1016/j.biomaterials.2023.122283
113. Riquelme E, Zhang Y, Zhang LL, et al. Tumor microbiome diversity and composition influence pancreatic cancer outcomes. Cell. 2019;178(4):795–806.e712. doi:10.1016/j.cell.2019.07.008
114. Zhan YX, Zhang YX, Mao CC, et al. Improved tumor infiltration and immunomodulation for tumor therapy: a pathway based on tetrahedral framework nucleic acids coupled bacterial nanocells. Nano Lett. 2023;23(1):353–362. doi:10.1021/acs.nanolett.2c03943
115. Zhang YL, Pan ML, Ma F, et al. Macrophage-targeted nanocarriers based on tetrahedral DNA nanostructure alleviate sepsis-induced acute lung injury by triple-pathway suppression of pyroptosis. ACS Appl Mater Interfaces. 2026;18(14):20128–20145. doi:10.1021/acsami.6c01459
116. Zhang YL, Li B, Fan ZJ, et al. Tetrahedral DNA nanostructure-based biomimetic nanovesicles attenuate sepsis-associated ARDS by suppressing glycolysis via the BMAL1/PFKFB3 axis. Adv Sci. 2026:e23782. doi:10.1002/advs.202523782
117. Wei CR, Liu YY, Pan BH, et al. TFNAs induce autophagy via activation of the nonclassical cGAS-STING signalling pathway to attenuate acute lung injury in mice. BMC Pulm Med. 2025;25(1):514. doi:10.1186/s12890-025-03970-w
118. Ma DH, Yu WJ, Zhou H, et al. Targeting sorting nexin 3 to treat pulmonary fibrosis by dual modulating Wnt/β-catenin signaling. Cell Death Dis. 2026;17(1):43. doi:10.1038/s41419-025-08248-x
119. Guo XL, Qiu XP, Hu XL, et al. APE1-Triggered Inhalable Microsphere (ATIM) for in situ non-small cell lung cancer theranostics. Adv Mater. 2026:e20216. doi:10.1002/adma.202520216
120. Jiang FL, Han Q, Deng H, et al. Inhalable tetrahedral DNA nanoplatforms for dual-barrier penetration and sustained deep-lung delivery of small-molecule drugs. Chem Eng J. 2026;528:172522. doi:10.1016/j.cej.2025.172522
121. Qin X, Xiao LR, Li N, et al. Tetrahedral framework nucleic acids-based delivery of microRNA-155 inhibits choroidal neovascularization by regulating the polarization of macrophages. Bioact Mater. 2022;14:134–144. doi:10.1016/j.bioactmat.2021.11.031
122. Kim B, Park JH, Sailor MJ. Rekindling RNAi therapy: materials design requirements for in vivo siRNA delivery. Adv Mater. 2019;31(49):e1903637. doi:10.1002/adma.201903637
123. Tian TR, Zhang T, Shi SR, et al. A dynamic DNA tetrahedron framework for active targeting. Nat Protoc. 2023;18(4):1028–1055. doi:10.1038/s41596-022-00791-7
124. Gao Y, Chen XY, Tian TR, et al. A lysosome-activated tetrahedral nanobox for encapsulated siRNA delivery. Adv Mater. 2022;34(46):e2201731. doi:10.1002/adma.202201731
125. Yan JQ, Chen J, Zhang N, et al. Mitochondria-targeted tetrahedral DNA nanostructures for doxorubicin delivery and enhancement of apoptosis. J Mater Chem B. 2020;8(3):492–503. doi:10.1039/c9tb02266j
126. Hu QQ, Li H, Wang LH, et al. DNA nanotechnology-enabled drug delivery systems. Chem Rev. 2019;119(10):6459–6506. doi:10.1021/acs.chemrev.7b00663
127. Shi SR, Li YJ, Zhang T, et al. Biological effect of differently sized tetrahedral framework nucleic acids: endocytosis, proliferation, migration, and biodistribution. ACS Appl Mater Interfaces. 2021;13(48):57067–57074. doi:10.1021/acsami.1c20657
128. Jiang DW, Rosenkrans ZT, Ni DL, et al. Nanomedicines for renal management: from imaging to treatment. Acc Chem Res. 2020;53(9):1869–1880. doi:10.1021/acs.accounts.0c00323
129. Jiang DW, Im HJ, Boleyn ME, et al. Efficient renal clearance of DNA tetrahedron nanoparticles enables quantitative evaluation of kidney function. Nano Res. 2019;12(3):637–642. doi:10.1007/s12274-019-2271-5
130. Zou KX, Zhang PF, Wang YQ, et al. Investigation and regulation of DNA nanostructures on activating cGAS-STING signaling. Small Methods. 2024:e2401041. doi:10.1002/smtd.202401041
131. Yu JW, Liu ZY, Jiang W, et al. De novo design of an RNA tile that self-assembles into a homo-octameric nanoprism. Nat Commun. 2015;6:5724. doi:10.1038/ncomms6724
132. Gratton SE, Ropp PA, Pohlhaus PD, et al. The effect of particle design on cellular internalization pathways. Proc Natl Acad Sci U S A. 2008;105(33):11613–11618. doi:10.1073/pnas.0801763105
133. Bellot G, McClintock MA, Lin CX, et al. Recovery of intact DNA nanostructures after agarose gel-based separation. Nat Methods. 2011;8(3):192–194. doi:10.1038/nmeth0311-192
134. Lin CX, Perrault SD, Kwak M, et al. Purification of DNA-origami nanostructures by rate-zonal centrifugation. Nucleic Acids Res. 2013;41(2):e40. doi:10.1093/nar/gks1070
135. Joyce P, Allen CJ, Alonso MJ, et al. A translational framework to DELIVER nanomedicines to the clinic. Nat Nanotechnol. 2024;19(11):1597–1611. doi:10.1038/s41565-024-01754-7
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
From Pioneering Discoveries to Innovative Therapies: A Journey Through the History and Advancements of Nanoparticles in Breast Cancer Treatment
Basingab FS, Alshahrani OA, Alansari IH, Almarghalani NA, Alshelali NH, Alsaiary AH, Alharbi N, Zaher KA
Breast Cancer: Targets and Therapy 2025, 17:27-51
Published Date: 21 January 2025
Progress in the Application of Novel Nanomaterials in Targeted Therapy for Liver Cancer

Wei X, Cao W, Wang S, Zhang Y, Gao Z, Wang S, Yao L, Zhang Z, Li X, Deng W, Xie Y, Li M
International Journal of Nanomedicine 2025, 20:2623-2643
Published Date: 3 March 2025
Targeting Multiple Pathophysiological Axes in COPD: Nanomaterial Advances
Zhang Q, Xu S, Yang C, Wang X, Liu T, Zhang X, Qu C, Wu J, Yang J, Xing X
International Journal of Nanomedicine 2025, 20:11989-12007
Published Date: 1 October 2025
Exosomes as Emerging Therapeutic Strategies in Primary Osteoporosis: A Narrative Review
Chen G, Li H, Chen R, Chen G
Drug Design, Development and Therapy 2025, 19:9099-9115
Published Date: 7 October 2025
Bridging the Gap: The Role of Advanced Formulation Strategies in the Clinical Translation of Nanoparticle-Based Drug Delivery Systems
Herdiana Y
International Journal of Nanomedicine 2025, 20:13039-13053
Published Date: 29 October 2025