Back to Journals » Infection and Drug Resistance » Volume 13
Acinetobacter baumannii Sequence Types Harboring Genes Encoding Aminoglycoside Modifying Enzymes and 16SrRNA Methylase; a Multicenter Study from Pakistan
Authors Khurshid M ![]() , Rasool MH, Ashfaq UA
, Rasool MH, Ashfaq UA ![]() , Aslam B, Waseem M
, Aslam B, Waseem M ![]() , Ali MA
, Ali MA ![]() , Almatroudi A
, Almatroudi A ![]() , Rasheed F
, Rasheed F ![]() , Saeed M, Guo Q, Wang M
, Saeed M, Guo Q, Wang M ![]()
Received 30 April 2020
Accepted for publication 29 July 2020
Published 18 August 2020 Volume 2020:13 Pages 2855—2862
DOI https://doi.org/10.2147/IDR.S260643
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sahil Khanna
Mohsin Khurshid,1,2 Muhammad Hidayat Rasool,2 Usman Ali Ashfaq,3 Bilal Aslam,2 Muhammad Waseem,2 Muhammad Akhtar Ali,4 Ahmad Almatroudi,5 Farhan Rasheed,6 Muhammad Saeed,2 Qinglan Guo,1 Minggui Wang1
1Institute of Antibiotics, Huashan Hospital, Fudan University, Shanghai 200040, People’s Republic of China; 2Department of Microbiology, Government College University, Faisalabad, Pakistan; 3Department of Bioinformatics & Biotechnology, Government College University, Faisalabad, Pakistan; 4School of Biological Sciences, University of the Punjab, Lahore, Pakistan; 5Department of Medical Laboratories, College of Applied Medical Sciences, Qassim University, Buraydah, Saudi Arabia; 6Allama Iqbal Medical College, Jinnah Hospital Lahore, Lahore, Pakistan
Correspondence: Mohsin Khurshid
Department of Microbiology, Government College University, Faisalabad, Pakistan
Tel +923334301513
Email [email protected]
Introduction: The aminoglycosides are widely used for the therapeutic management of infections caused by gram-negative bacteria, including the Acinetobacter baumannii strains. However, the resistance to the members of the aminoglycoside family, such as amikacin, gentamicin, and tobramycin, is increasingly being common among the clinical isolates.
Purpose: This study aimed to investigate the presence of 16SrRNA methylases and aminoglycoside modifying enzymes (AMEs) genes among aminoglycoside resistant A. baumannii isolates and to study the genetic diversity of the clinical population of A. baumannii in local hospitals.
Material and Methods: The 143 A. baumannii clinical strains were analyzed for antimicrobial susceptibility, genetic screening for enzymes conferring aminoglycosides resistance followed by the multilocus sequence typing.
Results: The 133/143 (93%) isolates were non-susceptible to at least one of the tested aminoglycosides, including amikacin, gentamicin, and tobramycin. The MIC distribution has shown that 87.486.7% strains were resistant to amikacin and gentamicin, respectively. The aphA 6, aadB, aacC 1, and aphA 1 were found in 74.1%, 59.4%, 16.1%, and 11.2% isolates, respectively, whereas the armA was found in 28% of the strains having a higher MIC value (MIC; ≥ 256μg/mL). The MLST data have shown that the ST589 and ST2 were the most common STs and corresponded to 51 (35.7%) and 38 (26.6%) isolates, respectively, and few of the isolates corresponding to these STs were found to harbor the armA gene with a variable genotypic profile for AMEs.
Discussion: The study has reported the incidence of various enzymes conferring aminoglycoside resistance among the A. baumannii clones for the first time from Pakistan. The findings suggest the possibility of transmission of aminoglycoside resistance determinants through the lateral gene transfer as well as clonal dissemination.
Keywords: aminoglycosides, armA, MLST, gentamicin, A. baumannii
Introduction
The remarkable ability of Acinetobacter baumannii strains to develop resistance to various groups of antimicrobial agents through a mutational event or lateral acquisition of foreign genes have made this pathogen successful in healthcare settings.1–4 The aminoglycosides along with beta-lactam drugs are used for the treatment of infections caused by Gram-negative multidrug-resistant (MDR) pathogens.5 The combination of beta-lactams and colistin is also recommended in some studies however as far as the clinical success is concerned, the combination therapy was not found to have any clear advantage over colistin monotherapy.6 Despite the associated side effects, aminoglycosides particularly amikacin, tobramycin, and gentamicin are frequently prescribed in the USA for the treatment of serious infections.7 However, many pathogenic bacteria have developed resistance to the aminoglycosides through modification of the antimicrobial drugs, alteration of the targets, ie, ribosomes, decreased permeability, and through efflux pumps.8–10
The aminoglycoside-modifying enzymes (AMEs) got substantial clinical impact because of their dissemination through transposons, plasmids, or integrons that are frequently known to harbor additional resistance determinants, including β-lactamases which helps to facilitate their dissemination. AMEs deactivate the aminoglycosides through acetylation, adenylation, and phosphorylation. Moreover, the AMEs possess a diverse resistance phenotype because of their variable spectrum of activity for particular aminoglycosides.7,11 The studies have reported that the phosphotransferase (APH), nucleotidyltransferase (ANT), and acetyltransferase (AAC) are mainly responsible for aminoglycoside resistance among the non-fermenters Gram-negative bacterial pathogens including the Acinetobacter baumannii and Pseudomonas aeruginosa. It is suggested that the prevalence of the genes conferring aminoglycoside resistance correlates with aminoglycoside usage and varies among different clinical settings and clinical isolates.12,13 The 16SrRNA methylases are increasingly being common among different gram-negative pathogens during the last decade which poses a serious threat to the clinical implications of aminoglycoside.14 The clinical isolates producing these 16SrRNA methylases were reported from various parts of the world and are associated with high-level resistance to aminoglycosides. Various 16SrRNA methylases have been identified such as armA, npmA, and rmtA–H.15
The study was designed to assess the occurrence of AMEs and 16SrRNA methylase among A. baumannii isolates obtained from multiple tertiary care centers of Pakistan for the first time. The multilocus sequence typing was performed to study the molecular epidemiology of aminoglycoside resistant A. baumannii clones in the local hospitals.
Materials and Methods
Bacterial Isolates
A total of 143 non-duplicate Acinetobacter baumannii isolates were collected from the five tertiary care medical centers of Lahore, Pakistan from January to July 2017. The isolates were cultured from clinical specimens of admitted patients including tracheal secretions (n=32), blood (n=26), sputum (n=22), urine (n=16), Pus (n=14), wound swab (n=10), CSF (n=7), Bronchial washings (n=6), endotracheal tube (n=5) catheter tip (n=3) and fluid (n=2). The clinical record of the patients from whom the specimens were recovered was reviewed retrospectively. All the isolates were originated from nosocomial infections as the strains were obtained after 48 hours of hospital admittance.
The clinical isolates were initially identified using API 20NE (bioMérieux, France) and were further verified using multiplex PCR to target the 425-bp fragment of the recA gene present in Acinetobacter spp. and the 208-bp fragment corresponding to the intergenic spacer region which is solely present in A. baumannii strains as described previously.2,4 Furthermore, the isolates were subjected to PCR to detect the intrinsic blaOXA-51-like genes as described previously.2,4 The strains were stored at –80°C (40% Glycerol stocks) using brain heart infusion (BHI) broth (Oxoid, UK) and were grown on Luria-Bertani Broth (Oxoid, UK).
Antimicrobial Susceptibility Testing
The disc diffusion assays for the following antimicrobial agents were performed: amikacin (AK) 30µg, gentamicin (CN) 10µg, tobramycin (TOB) 10µg, imipenem (IMP) 10µg, meropenem (MEM) 10µg, ceftazidime (CAZ) 30µg, cefotaxime (CTX) 30µg, cefepime (FEP) 30µg, ampicillin-sulbactam (SAM) 30µg, piperacillin-tazobactam (TZP) 110µg, doxycycline (DO) 30µg, ciprofloxacin (CIP) 5µg and trimethoprim-sulfamethoxazole (SXT) 25µg. The antimicrobial discs were obtained from Oxoid, UK. The minimal inhibitory concentrations (MICs) were determined by the agar dilution method except for colistin for which the broth micro-dilution method was used. The results were interpreted according to the Clinical and Laboratory Standards Institute (CLSI, 2018) guidelines. To interpret tigecycline susceptibility, criteria defined by food and drug authority (FDA, USA) were used, ie, isolates with MIC ≥ 8µg/mL were considered resistant.2,16 Escherichia coli (ATCC® 25,922 and ATCC® 35,218) and Pseudomonas aeruginosa ATCC® 27,853 were used as quality control strains.
Aminoglycoside Resistant Determinants
All the A. baumannii isolates were examined for the presence of AMEs (aphA1, aphA6, aacC1 aadA1, and aadB) and 16SrRNA methylase (armA, rmtA, rmtB, rmtC, rmtD, rmtE, and rmtF) using specific primers. The Positive products were purified using a DNA purification kit and were sent to Shanghai Jieli Biotechnology (Shanghai, China) for sequencing. The obtained sequences were analyzed using the BLAST (NCBI) tool.
MLST
The multilocus sequence typing (MLST) was performed for all (n=143) isolates according to the Pasteur scheme as described previously.3,17 Briefly, the fragments of seven different housekeeping genes including the cpn60, fusA, gltA, pyrG, recA, rplB, and rpoB were amplified and the sequences were analyzed by the PubMLST database https://pubmlst.org/bigsdb?db=pubmlst_abaumannii_seqdef.
Results
Overall 133/143 (93%) isolates were non-susceptible to at least one of the tested aminoglycosides including amikacin, gentamicin, and tobramycin (Disc diffusion method only). Among these 133 isolates, 99.2% isolates were resistant to the third and fourth generation cephalosporins including cefotaxime, ceftazidime, and cefepime as well as to the carbapenems, ie, imipenem and meropenem. The 129/133 (97%) isolates were non-susceptible to fluoroquinolones (ciprofloxacin) and 80.554.1% strains were resistant to trimethoprim-sulfamethoxazole and doxycycline, respectively. The MIC of amikacin and gentamicin showed that 125/143 (87.4%) and 124/143 (86.7%) strains were resistant to amikacin (MIC ≥64 µg/mL) and gentamicin (MIC ≥16µg/mL) as shown in Table 1. All the strain included in the study were found susceptible to tigecycline (MIC, ≤2µg/mL) and colistin (MIC, ≤2µg/mL) with MIC90 as 2 µg/mL and 1 µg/mL, respectively.
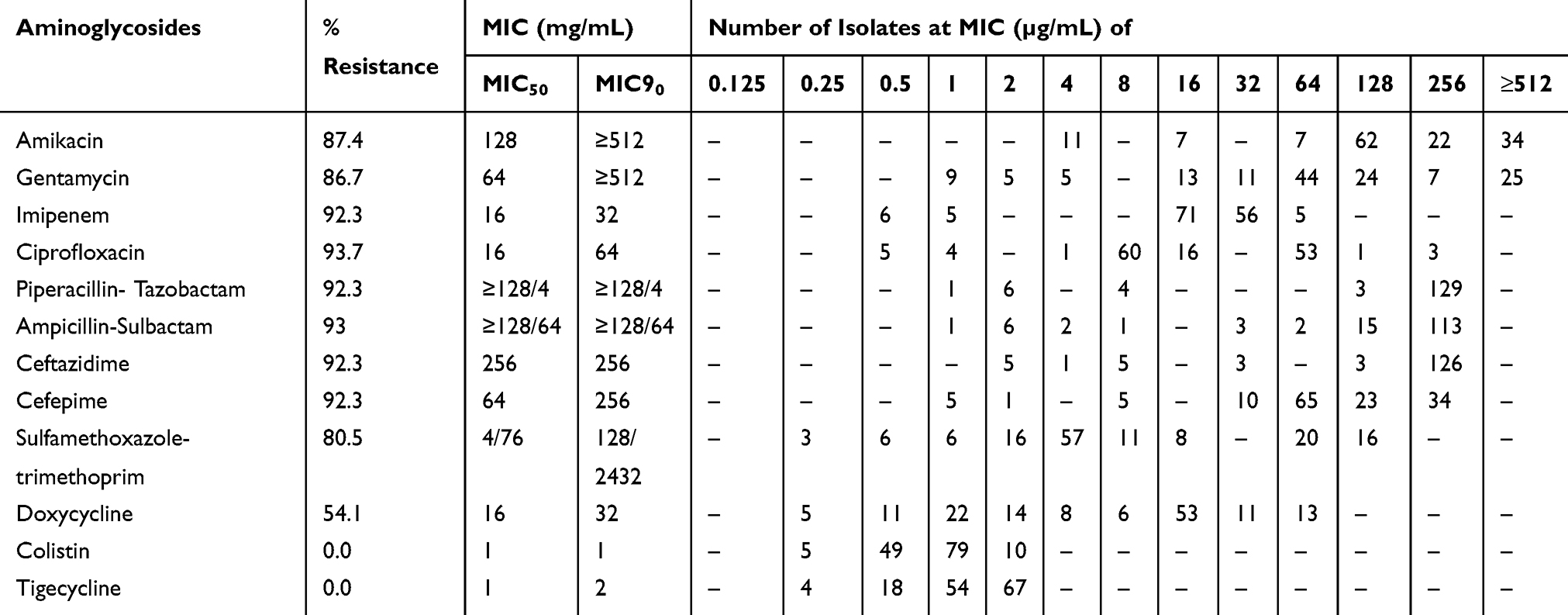 |
Table 1 Overall Distribution of MICs of Various Antimicrobial Agents Against A. Baumannii (n=143) Strains |
The isolates that were positive for the recA gene and 208-bp fragment of the intergenic spacer region were also positive for blaOXA-51 gene. All the isolates were screened for the genes encoding for aminoglycoside modifying enzyme (AMEs) and 16SrRNA methylase. Among all the investigated AMEs genes, the positive rates for aphA6, aadB, aacC1, and aphA1 were 74.1%, 59.4%, 16.1%, and 11.2%, respectively, whereas the aadA1 was not detected among the isolates. The armA was found in 28% of the strains (Table 2), whereas the rmtA-E genes were not found among any A. baumannii isolates.
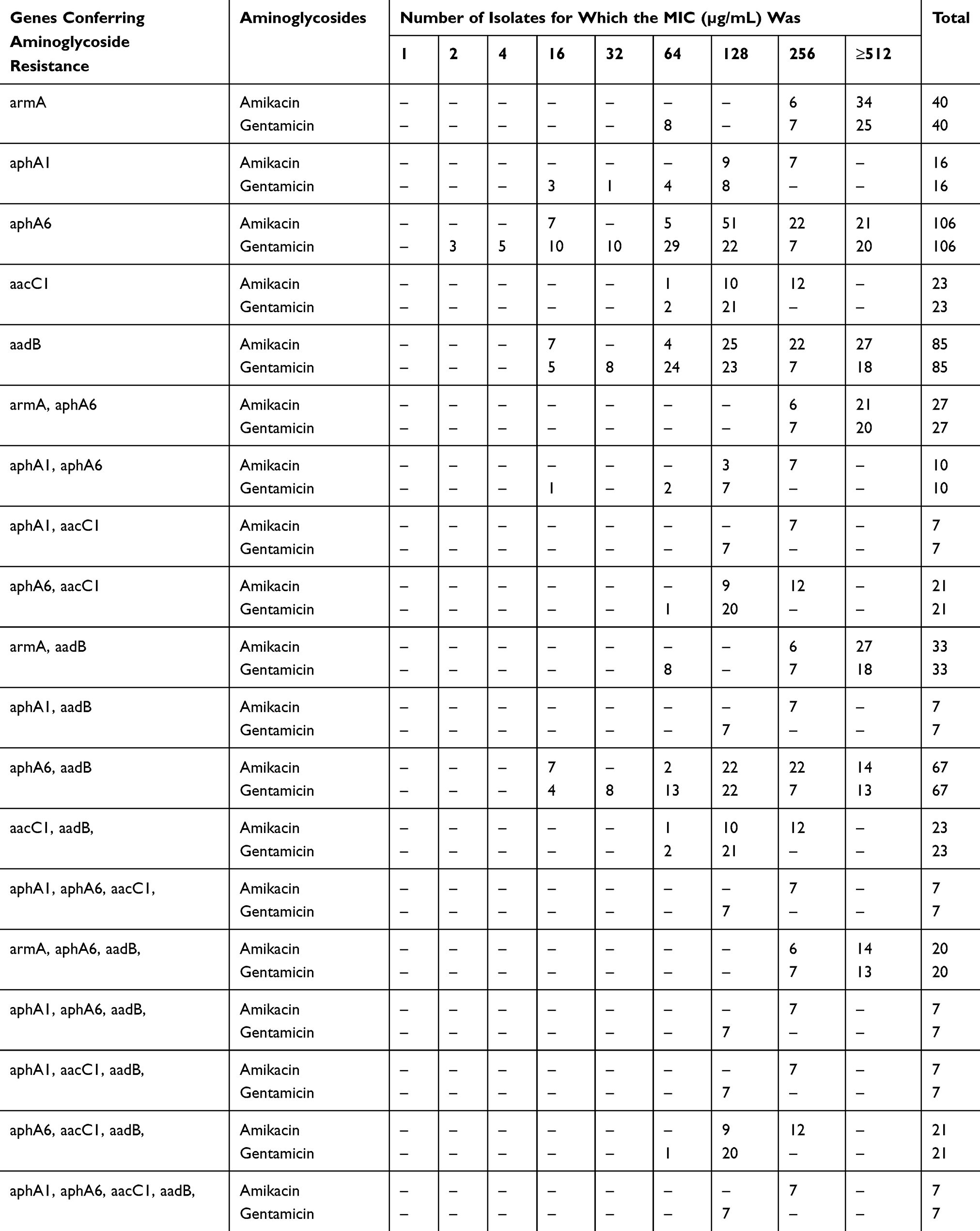 |
Table 2 Combinations of Aminoglycoside Resistant Determinants and Corresponding MICs for A. Baumannii Strains |
Among the sequence types (STs), ST589 and ST2 were the most common STs and corresponded to a total of 51 (35.7%) and 38 (26.6%) isolates, respectively. The 18 (12.6%) and 8 (5.6%) isolates belong to the ST642 and ST889, respectively. Among the isolates belonging to ST 589, armA was found in 18 out of 51 isolates while 40 isolates belonging to ST589 were positive for aphA6 gene.
The ST1209, New ST1, New ST2, New ST3, and New ST6 were sensitive to aminoglycosides and do not harbor any of the AMEs. The MLST types of A. baumannii isolates harboring the genes encoding for aminoglycoside resistance are summarized in Table 3.
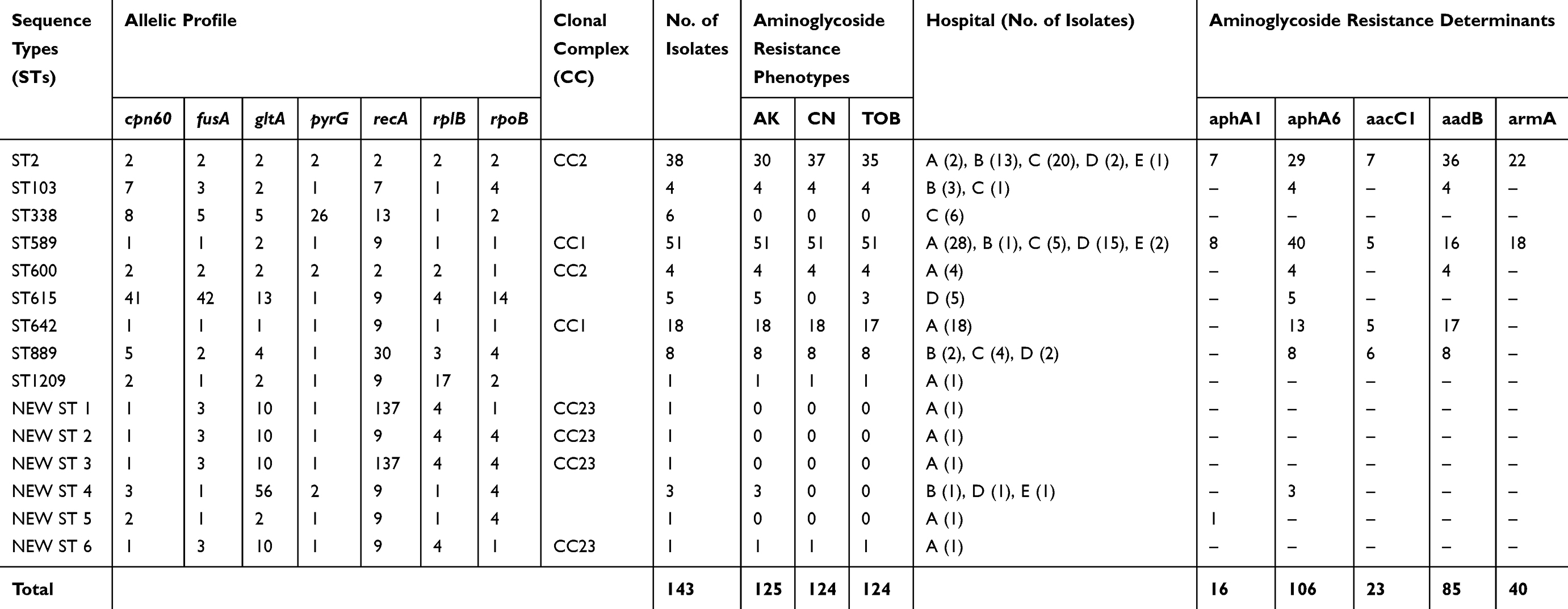 |
Table 3 Distribution of Aminoglycoside Resistance Determinants Among Different Sequence Types (STs) |
Discussion
The enzymes which confer resistance to aminoglycosides especially the 16SrRNA methylases possess a significant threat to public health. The treatment choices for the infections caused by the multidrug-resistant (MDR) or extensive drug resistance (XDR) bacteria have already been limited due to the increasing rate of resistance to carbapenems. In the present investigation, we have evaluated the presence of aminoglycoside resistance genes among A. baumannii isolates obtained from the tertiary care centers of Lahore, Pakistan for the very first time. The findings have established the fact that 93% of the isolates were non-susceptible to at least one of the three tested aminoglycosides (tobramycin, gentamicin, and amikacin).
The A. baumannii isolates were screened for the presence of the AME genes and 16SrRNA methylase. The AMEs, ie, aphA6, aadB, aacC1, and aphA1 were found in different combinations among the aminoglycoside resistant isolates, emphasizing the foremost role of AMEs in conferring aminoglycoside resistance among A. baumannii strains. The aphA6 [APH(3ʹ) VIa] was the commonest AME with an overall distribution rate of 74.1%, while its incidence was 79.7% among the isolates resistant to at least one of the tested aminoglycosides. The overall dominance of aphA6 among the A. baumannii isolates has been documented in neighboring countries as well as in many other countries.18,19 The presence of aphA6 in gentamicin and amikacin non-susceptible isolates in the present study is consistent with the previous studies which have shown that amikacin, gentamicin are the substrates for aphA6 enzymes.20
The aminoglycoside nucleotidyltransferase (ANT) gene, ie, aadB [ANT(2ʹ)-Ia] was also frequent with an overall incidence of 59.4%. The studies have widely reported the distribution of aadB as gene cassettes on class-I and class-II integrons, therefore, the transfer is easily mediated through the mobile genetic elements. These enzymes confer resistance to tobramycin, dibekacin, gentamicin, sisomicin, and kanamycin.21 The aminoglycoside acetyltransferase, ie, aacC1 [AAC(3)-Ia] was found in 23/133 (17.3%). The studies have reported that aacC1 confers resistance to gentamicin, tobramycin, and sisomicin although the frequency of this acetyltransferase varies among various studies.22,23
The study has shown that aminoglycoside resistant strains from the hospitals corresponded to diverse sequence types. The most common STs found in the study were the sequence type (ST) 589 belonging to clonal complex (CC) 1 and the sequence type (ST) 2 belonging to clonal complex (CC) 2 which comprised more than half of the isolates. However, the armA gene harboring isolates were belonging to ST589 and ST2, however, all the isolates corresponding to these two STs were not found to harbor the armA (16SrRNA methylase) gene. This shows heterogeneity among the STs as the isolates belonging to the same STs shown a different combination of genes. Although the studies suggest the spread of resistant bacterial strains harboring the resistant genes and plasmids through clonal dissemination, many studies proposed the transmission of the genes, as well as plasmids conferring the resistance phenotypes through the lateral gene transfer between the different strains of A. baumannii.24–27 The identification of same resistant determinants in the A. baumannii isolates with diverse STs and the isolates having the same genetic background or clones (MLST types) with the acquisition of dissimilar resistant genes in any individual clone suggests the possibility of horizontal transfer.27 This transmission happens through the dissemination of mobile elements or by clonal spread possibly vary due to the diverse conditions existing in different geographical areas.
A large number of studies have shown that CC2 (Pasteur scheme)/CC92 (Oxford Scheme) is prevalent throughout Asian countries as well as the widest worldwide distribution.28 The various STs belonging to CC2/CC92 (eg, ST2, and ST600) are single locus variants (SLV) to each other and have been reported from the Asian countries.17,29 It is suggested that A. baumannii clones belonging to CC2 have spread among the local hospitals and evolved, resulting in the development of different SLVs in the CC2.
The emergence of 16SrRNA methylase in addition to the multiple types of carbapenemases is evinced by the large numbers of recent studies describing the co-existence of these two mechanisms.30–32 Before the dissemination of 16SrRNA methylase, the resistance to various aminoglycosides involves the bacterial species to acquire and gather different AMEs. Therefore, the emergence of 16SrRNA methylases is attaining a similar significance for aminoglycoside resistance that different carbapenemases have achieved for beta-lactam resistance, ie, successfully halting the therapeutic efficacy of all agents from a class of antibiotics by the acquisition of a single enzyme for example blaNDM.
In conclusion, the study highlighted the heterogeneity of aminoglycoside resistant A. baumannii mediated by the armA and AMEs for the first time in the Pakistani hospitals. The disparities in the frequencies of different resistance determinants among the various clones may be associated with the differential use of antibiotics in different clinical settings. The present study describes the spreading of A. baumannii strains belonging to CC2 (ST2 and ST600) and CC1 (ST589 and ST642) in Pakistani hospitals. The dissemination of diverse STs of A. baumannii in different hospitals harboring AMEs and 16SrRNA methylases, ie, armA genes represents the establishment of this important nosocomial pathogen in the local hospitals and highlights the necessity of taking effective control measures.
Ethics Statement
The A. baumannii isolates in this study were part of the routine hospital laboratory procedure; therefore, the Institutional Review Board, Government College University Faisalabad, Pakistan exempted this research for written informed consent and approved the study vide letter number GCUF/IRB/4221 dated 29-04-2016.
Disclosure
The authors report no conflicts of interest in this work.
References
1. McGann P, Courvalin P, Snesrud E, et al. Amplification of aminoglycoside resistance gene aphA1 in Acinetobacter baumannii results in tobramycin therapy failure. MBio. 2014;5(2):e00915. doi:10.1128/mBio.00915-14
2. Khurshid M, Rasool MH, Ashfaq UA, Aslam B, Waseem M. Emergence of ISAba1 harboring carbapenem-resistant Acinetobacter baumannii isolates in Pakistan. Future Microbiol. 2017;12:1261–1269. doi:10.2217/fmb-2017-0080
3. Khurshid M, Rasool MH, Ashfaq UA, et al. Dissemination of blaOXA-23 harboring Carbapenem-resistant Acinetobacter baumannii clones in Pakistan. J Glob Antimicrob Resist. 2020;21:357–362. doi:10.1016/j.jgar.2020.01.001
4. Khurshid M, Rasool MH, Siddique MH, et al. Molecular mechanisms of antibiotic co-resistance among carbapenem resistant Acinetobacter baumannii. J Infect Dev Ctries. 2019;13(10):899–905. doi:10.3855/jidc.11410
5. Tamma PD, Cosgrove SE, Maragakis LL. Combination therapy for treatment of infections with gram-negative bacteria. Clin Microbiol Rev. 2012;25(3):450–470. doi:10.1128/CMR.05041-11
6. Nørgaard SM, Jensen CS, Aalestrup J, Vandenbroucke-Grauls C, de Boer MGJ, Pedersen AB. Choice of therapeutic interventions and outcomes for the treatment of infections caused by multidrug-resistant gram-negative pathogens: a systematic review. Antimicrob Resist Infect Control. 2019;8:170. doi:10.1186/s13756-019-0624-1
7. Krause KM, Serio AW, Kane TR, Connolly LE. Aminoglycosides: an overview. Cold Spring Harb Perspect Med. 2016;6:6. doi:10.1101/cshperspect.a027029
8. Sohail M, Rashid A, Aslam B, et al. Antimicrobial susceptibility of Acinetobacter clinical isolates and emerging antibiogram trends for nosocomial infection management. Rev Soc Bras Med Trop. 2016;49(3):300–304. doi:10.1590/0037-8682-0111-2016
9. Costello SE, Deshpande LM, Davis AP, Mendes RE, Castanheira M. Aminoglycoside-modifying enzyme and 16S ribosomal RNA methyltransferase genes among a global collection of Gram-negative isolates. J Glob Antimicrob Resist. 2019;16:278–285. doi:10.1016/j.jgar.2018.10.020
10. Aslam B, Wang W, Arshad MI, et al. Antibiotic resistance: a rundown of a global crisis. Infect Drug Resist. 2018;11:1645–1658. doi:10.2147/IDR.S173867
11. Vakulenko SB, Mobashery S. Versatility of aminoglycosides and prospects for their future. Clin Microbiol Rev. 2003;16(3):430–450. doi:10.1128/CMR.16.3.430-450.2003
12. Nemec A, Dolzani L, Brisse S, van den Broek P, Dijkshoorn L. Diversity of aminoglycoside-resistance genes and their association with class 1 integrons among strains of pan-European Acinetobacter baumannii clones. J Med Microbiol. 2004;53(Pt 12):1233–1240. doi:10.1099/jmm.0.45716-0
13. Taitt CR, Leski TA, Stockelman MG, et al. Antimicrobial resistance determinants in Acinetobacter baumannii isolates taken from military treatment facilities. Antimicrob Agents Chemother. 2014;58(2):767–781. doi:10.1128/AAC.01897-13
14. Fair RJ, Tor Y. Antibiotics and bacterial resistance in the 21st century. Perspect Medicin Chem. 2014;6:25–64. doi:10.4137/PMC.S14459
15. Ayad A, Drissi M, de Curraize C, et al. Occurrence of arma and rmtb aminoglycoside resistance 16S rRNA methylases in extended-spectrum β-Lactamases producing escherichia coli in algerian hospitals. Front Microbiol. 2016;7:1409. doi:10.3389/fmicb.2016.01409
16. Sader HS, Farrell DJ, Flamm RK, Jones RN. Variation in potency and spectrum of tigecycline activity against bacterial strains from U.S. medical centers since its approval for clinical use (2006 to 2012). Antimicrob Agents Chemother. 2014;58(4):2274–2280. doi:10.1128/AAC.02684-13
17. Al-Atrouni A, Hamze M, Jisr T, et al. Wide spread of OXA-23-producing carbapenem-resistant Acinetobacter baumannii belonging to clonal complex II in different hospitals in Lebanon. Int J Infect Dis. 2016;52:29–36. doi:10.1016/j.ijid.2016.09.017
18. Aghazadeh M, Rezaee MA, Nahaei MR, et al. Dissemination of aminoglycoside-modifying enzymes and 16S rRNA methylases among Acinetobacter baumannii and Pseudomonas aeruginosa isolates. Microb Drug Resist. 2013;19(4):282–288. doi:10.1089/mdr.2012.0223
19. Sheikhalizadeh V, Hasani A, Ahangarzadeh Rezaee M, et al. Comprehensive study to investigate the role of various aminoglycoside resistance mechanisms in clinical isolates of Acinetobacter baumannii. J Infect Chemother. 2017;23(2):74–79. doi:10.1016/j.jiac.2016.09.012
20. Davies J, Wright GD. Bacterial resistance to aminoglycoside antibiotics. Trends Microbiol. 1997;5(6):234–240. doi:10.1016/S0966-842X(97)01033-0
21. Ramirez MS, Tolmasky ME. Aminoglycoside modifying enzymes. Drug Resist Updat. 2010;13(6):151–171. doi:10.1016/j.drup.2010.08.003
22. Xiao-Min X, You-Fen F, Wei-Yun F, Zu-Huang M, Xing-Bei W. Antibiotic resistance determinants of a group of multidrug-resistant Acinetobacter baumannii in China. J Antibiot. 2014;67(6):439–444. doi:10.1038/ja.2014.18
23. Gholami M, Haghshenas M, Moshiri M, et al. Frequency of 16S rRNA methylase and aminoglycoside-modifying enzyme genes among clinical isolates of Acinetobacter baumannii in Iran. Iran J Pathol. 2017;12(4):329–338. doi:10.30699/ijp.2017.27989
24. Chen Y, Yang Y, Liu L, et al. High prevalence and clonal dissemination of OXA-72-producing Acinetobacter baumannii in a Chinese hospital: a cross sectional study. BMC Infect Dis. 2018;18(1):491. doi:10.1186/s12879-018-3359-3
25. Li Y, Guo Q, Wang P, et al. Clonal dissemination of extensively drug-resistant Acinetobacter baumannii producing an OXA-23 beta-lactamase at a teaching hospital in Shanghai, China. J Microbiol Immunol Infect. 2015;48(1):101–108. doi:10.1016/j.jmii.2014.04.005
26. Salto IP, Torres Tejerizo G, Wibberg D, Puhler A, Schluter A, Pistorio M. Comparative genomic analysis of Acinetobacter spp. plasmids originating from clinical settings and environmental habitats. Sci Rep. 2018;8(1):7783. doi:10.1038/s41598-018-26180-3
27. Valenzuela JK, Thomas L, Partridge SR, van der Reijden T, Dijkshoorn L, Iredell J. Horizontal gene transfer in a polyclonal outbreak of carbapenem-resistant Acinetobacter baumannii. J Clin Microbiol. 2007;45(2):453–460. doi:10.1128/JCM.01971-06
28. Saffari F, Monsen T, Karmostaji A, Azimabad FB, Widerström M. Significant spread of extensively drug-resistant Acinetobacter baumannii genotypes of clonal complex 92 among intensive care unit patients in a university hospital in southern Iran. J Med Microbiol. 2017;66(11):1656–1662. doi:10.1099/jmm.0.000619
29. Saharman YR, Karuniawati A, Sedono R, et al. Endemic carbapenem-nonsusceptible Acinetobacter baumannii-calcoaceticus complex in intensive care units of the national referral hospital in Jakarta, Indonesia. Antimicrob Resist Infect Control. 2018;7:5. doi:10.1186/s13756-017-0296-7
30. Karthikeyan K, Thirunarayan MA, Krishnan P. Coexistence of blaOXA-23 with blaNDM-1 and armA in clinical isolates of Acinetobacter baumannii from India. J Antimicrob Chemother. 2010;65(10):2253–2254. doi:10.1093/jac/dkq273
31. Nordmann P, Boulanger AE, Poirel L. NDM-4 metallo-beta-lactamase with increased carbapenemase activity from Escherichia coli. Antimicrob Agents Chemother. 2012;56(4):2184–2186. doi:10.1128/AAC.05961-11
32. Taj Z, Rasool MH, Almatroudi A, Saqalein M, Khurshid M. Extensively drug-resistant Acinetobacter baumannii belonging to international clone II from a pet cat with urinary tract infection; the first report from Pakistan. Pol J Microbiol. 2020;69(2):231–234. doi:10.33073/pjm-2020-017
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.