Back to Journals » International Journal of Nanomedicine » Volume 21
A Brain-Targeted Organophosphorus Hydrolase Fusion Protein Mediated by Angiopep-2 Neutralizes Central Organophosphates and Alleviates Neurotoxicity
Authors Xie Y, Zhai Y ![]() , Shang J, Ma M, Wang D, Xu S, Ran Y, Li Z, Gao X, Gao J
, Shang J, Ma M, Wang D, Xu S, Ran Y, Li Z, Gao X, Gao J ![]()
Received 19 February 2026
Accepted for publication 19 May 2026
Published 20 June 2026 Volume 2026:21 604162
DOI https://doi.org/10.2147/IJN.S604162
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Kamakhya Prakash Misra
Yanwei Xie,1,2 Yanan Zhai,1,2 Jiaxuan Shang,1 Ming Ma,1 Dan Wang,1 Shengye Xu,1 Yuhua Ran,1,2 Zhiping Li,1,2 Xiang Gao,1,2 Jing Gao1,2
1State Key Laboratory of National Security Specially Needed Medicines, Beijing, 100039, People’s Republic of China; 2Academy of Military Medical Sciences, Beijing, 100850, People’s Republic of China
Correspondence: Xiang Gao, Email [email protected] Jing Gao, Email [email protected]
Purpose: Organophosphates (OPs) exert neurotoxicity by inhibiting acetylcholinesterase (AChE) activity, leading to acetylcholine accumulation, excessive nervous system stimulation, and even fatal outcomes. Current clinical countermeasures focus on AChE reactivation and symptom antagonism but lack the ability to efficiently clear OPs in vivo, especially in the brain, failing to mitigate central neurotoxicity and resulting in persistent neuronal damage and permanent disability in survivors. Organophosphorus hydrolase (OPH) has robust OP clearance capacity but is hindered by poor blood-brain barrier (BBB) penetration. Thus, the purpose of this study is to address the BBB penetration challenge of OPH and develop an effective strategy for efficient clearance of brain-invading OPs to improve the prognosis of patients with OP-induced central neurotoxicity.
Patients and Methods: A fusion protein ANG-OPHDS5 was constructed by conjugating OPH mutant OPHDS5 with Angiopep-2 (ANG), a brain-targeting ligand that mediates transcytosis across the BBB via specific binding to low-density lipoprotein receptor-related protein 1 (LRP1). In vivo imaging and pharmacodynamic studies were performed to evaluate the brain-targeting ability, OP clearance efficiency, and neurotoxicity alleviation effect of ANG-OPHDS5.
Results: In vivo imaging and pharmacodynamic studies confirmed that the constructed fusion protein ANG-OPHDS5 efficiently achieved targeted delivery to the brain, effectively cleared OPs residing in the central nervous system (CNS), and significantly alleviated neurotoxicity induced by OPs.
Conclusion: Our strategy of centrally targeted delivery of OPH-based bioscavengers (ANG-OPHDS5) overcomes the limitations of current OP poisoning treatments, which lack efficient brain OP clearance capacity. This strategy holds great potential for improving the prognosis of patients with OP-induced central neurotoxicity.
Plain Language Summary: 1. Organophosphate poses a risk of brain damage, which can be effectively countered via cerebral delivery of organophosphorus hydrolase.
2. A brain-targeting recombinant fusion protein termed ANG-OPHDS5 was successfully constructed for efficient brain delivery.
3. ANG-OPHDS5 demonstrated potent antagonistic ability against lethal ethyl paraoxon toxicity and significantly alleviated accompanying central nervous system injury. The infographic consists of four sections. The first section, ’Fusion protein expression’ shows a gene construct with ligand, linker and protein components. It illustrates the process in E. coli BL21(DE3) leading to the production of ANG-OPHDS5 fusion protein. The second section, ’Poisoning treatment,’ depicts organophosphate exposure in mice, followed by intravenous administration of OPHDS5, leading to recovery. The third section, ’Central rescue,’ illustrates a synaptic gap with acetylcholine (ACh) and acetylcholinesterase (AChE) activity, highlighting the stopping of harmful effects. The fourth section, ’Blood-brain barrier delivery,’ shows a blood vessel with the delivery of substances across the blood-brain barrier into brain tissue, focusing on the synaptic gap area.Infographic on fusion protein expression, poisoning treatment, central rescue and blood-brain barrier delivery.
Keywords: organophosphorus hydrolase, angiopep-2, fusion protein, brain targeted delivery, organophosphate poisoning treatment
Introduction
Organophosphates (OPs), widely used as pesticides, exert their toxicity primarily by irreversibly inhibiting acetylcholinesterase (AChE). AChE localized at neuromuscular junctions, within the central nervous system (CNS), and on red blood cell membranes, plays a pivotal role in the degradation of acetylcholine. Inhibition of AChE leads to the accumulation of acetylcholine in synaptic clefts, causing persistent overstimulation of cholinergic nerves. The resultant typical clinical symptoms include salivation, muscle tremors, and respiratory distress, which may progress to coma or death in severe cases.1
OP exposure results in approximately 3 million poisoning cases and up to 300,000 deaths each year, imposing a significant global public health burden.2 Beyond their acute neurotoxic effects, OPs have also been linked to chronic neurological impairment, cognitive dysfunction, psychiatric manifestations, and multi-organ toxicity.3 The current standard treatment for OP poisoning involves atropine to block M-type cholinergic receptors and oxime reactivators (such as pralidoxime and obidoxime) to restore AChE activity.4 However, these drugs poorly penetrate the blood-brain barrier (BBB) and do not directly hydrolyze OP molecules. Consequently, OPs persist within the central nervous system, contributing to lasting neurological sequelae.5
To deliver such therapeutic agents effectively into the CNS, various drug delivery systems have been explored. Nanocarrier systems such as liposomes, polymeric micelles, and exosomes are also being actively explored for CNS delivery of therapeutic macromolecules. Intranasal administration can bypass the BBB via the olfactory neural pathway to deliver agents directly into brain tissue. However, these approaches still face challenges related to poor availability or high production costs.6–8 Receptor-mediated transcytosis represents a particularly promising approach.9,10 For instance, studies have demonstrated that fusion proteins constructed by linking paraoxonase 1 (PON1) to monoclonal antibodies targeting the insulin receptor on the BBB can facilitate the effective transport of the enzyme across the barrier while preserving hydrolytic activity, suggesting the potential of this strategy for protecting the CNS.11,12
Targeting ligands facilitate central delivery by mediating the transport of fusion proteins across the BBB through binding to specific receptors. Among these, Angiopep-2 binds to the low-density lipoprotein receptor-related protein 1 (LRP1), which is highly expressed on the surface of brain capillary endothelial cells (BCECs).13 The receptor-mediated transport mechanism is one of the most thoroughly studied pathways.14 Its advantage lies in its well-validated penetration efficiency, having been successfully applied to deliver various drugs (such as chemotherapeutic agents and opioid analgesics) for treating conditions like glioma and pain, demonstrating broad translational potential.15
Herein, we first report a robust strategy to thoroughly clear residual OP in the CNS post poisoning. This strategy involves the construction of a novel recombinant fusion protein via a standard E. coli system (Figure 1a). OPHDS5, a high-efficiency organophosphorus hydrolase variant developed in our laboratory based on the previously reported OPH‑S5 scaffold,16 was selected for its superior soluble expression profile and subsequently fused with Angiopep-2 via genetic engineering techniques. Leveraging the specific interaction between angiopep-2 and LRP1 on cerebrovascular endothelial cells, this fusion protein ANG-OPHDS5 can facilitate the transport of OPHDS5 across the BBB (Figure 1b), thereby enabling the simultaneous clearance of OP toxins in both peripheral and central regions. This study aims to address the critical unmet need of CNS detoxification after OP poisoning and provide a promising translational strategy for the clinical management of OP-induced neurological damage.
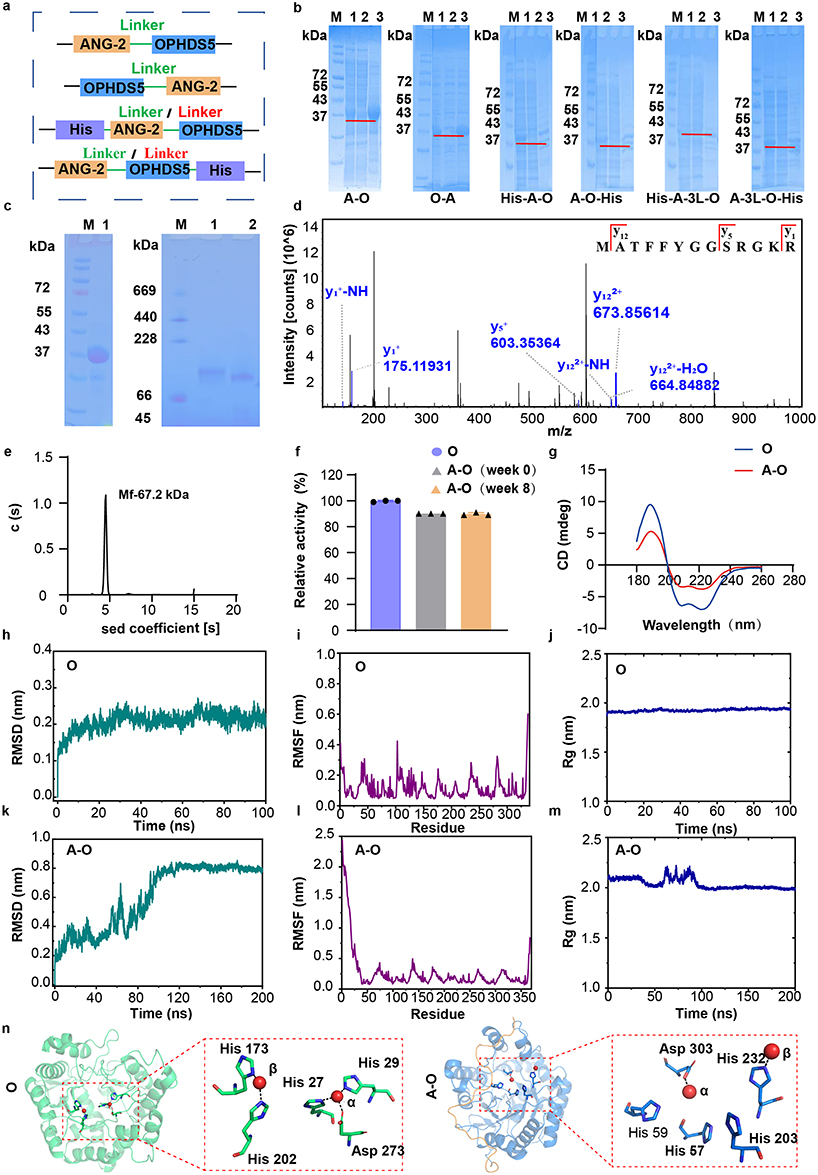 |
Figure 1 Design, screening and characterization of brain‑targeting fusion proteins (O represents OPHDS5, A-O represents ANG-OPHDS5). (a) Linear schematic of the recombinant fusion proteins; linker sequences are shown in green (SGGRGGA) and red (GGGGSGGGGSGGGGS). (b) SDS‑PAGE analysis of expression of the recombinant fusion protein. M, marker; 1, total bacterial lysate; 2, supernatant; 3, pellet. (c) Oligomeric state and molecular weight analysis of ANG‑OPHDS5 and OPHDS5 by SDS‑PAGE (left) and Native‑PAGE (right). M, marker; 1, ANG-OPHDS5; 2, OPHDS5. (d) Secondary mass spectrum (MS2) of the ANG sequence obtained via HPLC-MS/MS. (e) Molecular mass determination of ANG‑OPHDS5 using analytical ultracentrifugation. (f) Comparison of enzymatic activity between ANG‑OPHDS5 and OPHDS5, and stability of ANG‑OPHDS5 at 4 °C (n = 3). (g) Circular dichroism spectra of ANG‑OPHDS5 and OPHDS5. (h) Backbone root‑mean‑square deviation (RMSD, reflects overall structural stability) of OPHDS5 over time. (i) Per‑residue root‑mean‑square fluctuation (RMSF, reflects local residue flexibility) of OPHDS5. (j and m) Radius of gyration. (j) Radius of gyration (Rg, reflects protein structural compactness) of OPHDS5 over the simulation trajectory. (k) Backbone root‑mean‑square deviation (RMSD, reflects overall structural stability) of ANG‑OPHDS5 over time. (l) Per-residue root-mean-square fluctuation (RMSF, reflects local residue flexibility) of ANG-OPHDS5. (m) Radius of gyration (Rg, reflects protein structural compactness) of ANG‑OPHDS5 over the simulation trajectory. (n) Overall stable structures of the two proteins, with a detailed view of the interaction between Zn2⁺ and its coordinating residues. |
Materials and Methods
Study Design
This study aimed to develop a CNS-targeting biological scavenger for the emergency treatment of organophosphorus pesticide poisoning. To simultaneously target peripheral and central nervous systems, the recombinant fusion protein ANG-OPHDS5 was constructed by fusing the angiopep-2 peptide (ANG-2) to OPHDS5. The ANG-2 moiety facilitates drug delivery to the brain, while the OPHDS5 moiety hydrolyzes OPs into non- or low-toxicity products. The targeting specificity and mechanism of ANG-OPHDS5 were validated through in vivo imaging, immunofluorescence co-localization, and cellular uptake experiments. ANG-OPHDS5 was expressed in E. coli and purified via sequential steps of protamine sulfate precipitation, ammonium sulfate precipitation, affinity chromatography, anion-exchange chromatography, and gel filtration chromatography. Molecular dynamics simulations were employed to model the spatial structure of ANG-OPHDS5 and to disclose the rationale behind the observed enzymatic activity change. Pharmacodynamic efficacy was evaluated in a Sprague Dawley rat model of ethyl paraoxon poisoning. All animal experiments were conducted in accordance with the ARRIVE guidelines and approved by the Institutional Animal Care and Use Committee of the Academy of Military Medical Sciences (Approval No. DWZX-IACUC-2026-513).
Construction and Purification of CNS-Targeting Fusion Proteins
Six distinct expression plasmids were designed and constructed by cloning the coding sequences of ANG-2, linker and OPHDS5 into the pET-28a (+) vector. The recombinant plasmids were transformed into E. coli BL21 (DE3). Positive transformants were cultured in LB medium containing the appropriate antibiotic until reaching the exponential phase, followed by 0.2 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) induction at 25°C for 24 h. Bacterial pellets were collected, lysed by sonication, and centrifuged to separate the soluble and insoluble fractions. The soluble protein was purified through a multi-step process. Initial recovery used protamine sulfate and ammonium sulfate precipitation, followed by sequential affinity (Cytiva, HiTrapTM CaptoTM Phenyl ImpRes), anion-exchange (Cytiva, HiTrapTM Q HP), and gel filtration (Cytiva, Superdex 200 Increase 10/300GL) chromatography. Protein expression and purification were verified by SDS-PAGE and native PAGE, while concentration was quantified using the BCA assay kit (Thermo Scientific, Cat. No. 23227).
Structural Characterization of ANG-OPHDS5
Analytical ultracentrifugation technique (AUC) was utilized to determine the molecular weight and oligomeric state of ANG-OPHDS5 under native conditions. The sedimentation velocity experiments were performed at 20 °C using a Beckman Coulter Optima AUC instrument (40000 rpm), following the standard protocol.17 Samples were prepared in 50 mM HEPES buffer (pH 8.5) and loaded into 12 mm Epon centerpieces. During the experiment, absorbance scans were collected at 280 nm at 180 second intervals. Data were analyzed using SEDFIT software (version 16p36) with the continuous c(S) distribution model described by Schuck et al.18
The secondary structure and conformational stability of the proteins were assessed using circular dichroic (CD) spectrometer (Bio-Logic Science Instruments, MOS-450) equipped with a 150 W xenon lamp, with data acquired at a wavelength resolution of 0.1 nm. Samples were prepared in distilled water, at a final concentration of 0.2 mg/mL. The CD spectra were acquired from 180 to 280 nm, and the Spectra Manager software was used to analyze the data. The CONTIN algorithm was employed to evaluate the relative content of secondary structure.
Amino acid sequences of ANG-OPHDS5 were further determined by HPLC-MS/MS analysis. Target protein bands were excised from SDS-PAGE gels and purified through multi-steps, including destaining, reduction and digestion. The samples were desalted using C18 desalting columns (Qinglian Bai’ao Biotechnology Co., Ltd., QL-C18-C-10), and the final eluate was collected and lyophilized. The lyophilized powder was reconstituted in 0.1% formic acid for HPLC-MS/MS analysis (Q Exactive HF X mass spectrometer, protein analytical column, Cat. No. QL HPLC 100*15). Mass spectrometry analysis was performed in data-dependent acquisition mode. Detailed instrument parameters are provided in the Supplementary Methods.
Enzymatic Activity Assay
The enzyme activity was analyzed for the ability to hydrolyze ethyl paraoxon to p-nitrophenol (pNP) as described previously. A standard curve (R2 > 0.99) was established using 6 mM pNP, with absorbance measured at 405 nm. Reactions (total volume 1.005 mL) consisted of 5 μL ethyl paraoxon (10 μg/mL), 900 μL HEPES buffer (pH 8.0), and 100 μL enzyme solution (500 ng/mL). After incubation at 37 °C for 10 minutes, the reaction was terminated by adding 1 mL of 10% trichloroacetic acid, followed by 1 mL of 10% sodium carbonate for color development. Hydrolytic activity was quantified by measuring the absorbance of pNP at 405 nm using a microplate reader, with three replicate wells measured for each sample. The enzyme activity (U/mg) was calculated using the following formula:
Activity = [pNP]/(t × [E]), where [pNP] is the concentration of produced pNP (μM), t is the reaction time (10 minutes), and [E] is the final enzyme concentration in the reaction mixture (mg/mL).
Molecular Dynamics (MD) Simulations
The structural models for OPHDS5 and the fusion protein ANG-OPHDS5 were generated using AlphaFold 3.0 based on their respective amino acid sequences. Catalytic Zn2⁺ ions were then placed into the predicted active sites, resulting in the initial structures for simulation. Molecular dynamics (MD) simulations were performed using the GROMACS software. Each protein system was solvated in a cubic box of TIP3P water molecules. Sodium and chloride ions were then added at random positions to neutralize the system.
The CHARMM36 force field was employed for the simulations. Each system first underwent energy minimization using the steepest descent algorithm. This was followed by a 500 ps simulation in the canonical (NVT) and a 1000 ps equilibration in the isothermal-isobaric (NPT) ensemble. Finally, production MD simulations were conducted for 100 ns to 200 ns per system. Resulting trajectories were analyzed using tools within the GROMACS package. Structural visualization and further analysis were performed with Visual Molecular Dynamics, Python-based Molecular Viewer, and ChimeraX.
Immunofluorescence Staining for Brain Tissues
C57BL/6N mice (∼6 weeks old; male, Beijing SPF Biotechnology Co., Ltd.) were anesthetized via inhalation of isoflurane (induction concentration: 4% isoflurane; maintenance concentration: 1.5–2.0% isoflurane, delivered in oxygen) before all sampling procedures. Mice received an intravenous injection via the tail vein of either CY5.5-labeled ANG-OPHDS5 or OPHDS5 at a dose equivalent to 168 µg/kg of the OPHDS5 protein component. At 50 minutes post-injection, deeply anesthetized mice were transcardially perfused with PBS followed by 4% paraformaldehyde (PFA), and the brains were harvested and post-fixed in 4% PFA overnight at 4 °C. Complete perfusion fixation resulted in irreversible animal death, and no additional euthanasia procedures were required. All animal experimental procedures were performed in accordance with the American Veterinary Medical Association (AVMA) guidelines for laboratory animal care. After fixation, tissues were dehydrated, cleared, and embedded in paraffin according to standard histological protocols. Coronal sections (5 µm thickness) were cut using a microtome, mounted on slides, and deparaffinized. After rehydration through a graded ethanol series to PBS, antigen retrieval was performed using citrate buffer (pH 6.0). Sections were blocked with 10% normal goat serum containing 0.3% Triton X-100 in PBS for 1 h at room temperature, and then incubated overnight at 4 °C with an anti-CD31 primary antibody (1:200, Abcam, Cat. No. ab28364). After washing, sections were incubated with a corresponding secondary antibody (donkey anti-rat IgG H&L conjugated to Alexa Fluor 488, 1:200, Invitrogen, Cat. No. A-21206) for 60 minutes at room temperature. Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (Servicebio, Cat. No. G1012). Finally, sections were washed, and coverslipped with an anti-fade mounting medium. Fluorescence images were captured using a fluorescence microscope (Leica DMi8). To quantify the association of the injected constructs with the vasculature, images were analyzed for co-localization of the CY5.5 signal (from ANG-OPHDS5/OPHDS5) with CD31-positive vascular structures.
In vivo Imaging
Cy5.5-labeled ANG-OPHDS5 and OPHDS5 were administered via tail vein injection at a dose equivalent to 168 μg of the OPHDS5 protein component per kilogram of C57BL/6N mice body weight. The control group received an equal volume of normal saline. All mice were adequately anesthetized by inhalation of isoflurane (4% for induction, 1.5–2.0% for maintenance in oxygen) during in vivo imaging. At designated time points post-injection (15, 25, 50, and 100 min), the biodistribution was monitored using an in vivo imaging system (Guangzhou Guangyi Instrument, LASER 600). Fluorescence intensity was quantified, and its temporal profile was analyzed. After completion of all in vivo imaging experiments, all mice were humanely euthanized by deepening isoflurane inhalation (maintenance concentration increased to 5% in oxygen) until respiratory and cardiac arrest, in strict accordance with the American Veterinary Medical Association (AVMA) guidelines for laboratory animal euthanasia.
Cellular Uptake and Cytotoxicity Assay
Stably passaged mouse brain microvascular endothelial cells (bEnd.3) were purchased from Procell Life Science & Technology Co., Ltd. (Wuhan, China), a qualified commercial cell repository. Cells were seeded into 6-well plates at a density of 1×105 to 5×105 cells per well and cultured for 12 h to allow complete adhesion. The cells were then incubated with Cy5.5-labeled ANG-OPHDS5 or OPHDS5 at final protein concentrations of 5, 10, 20, 30 and 40 μg per well. After 6 h incubation, the cells were washed with PBS and detached using trypsin. Treated cells were washed again and analyzed by flow cytometer (Becton, Dickinson and Company, FACSLyric) to determine the cellular uptake rates of the two proteins. Additionally, to assess the time-dependent uptake, bEnd.3 cells were incubated with ANG-OPHDS5 at a fixed concentration of 20 μg per well for 15, 30, 60, 120, and 240 minutes. The cells were then collected and analyzed by flow cytometry (Becton, Dickinson and Company, FACSLyric) to measure the uptake rate at each time point.
For the cytotoxicity assay, bEnd.3 cells were incubated with varying amounts of ANG-OPHDS5 (0, 0.2, 0.4, 0.8, 1.0 mg/mL) for 1 h. Cell viability was assessed using the CCK-8 assay.
Analysis of Serum and Brain Biochemical Markers
Thirty adult male Sprague-Dawley (SD) rats were randomly divided into 5 groups (n=6 per group) and treated as follows: (1) Saline Control: received subcutaneous injection of normal saline. (2–3) Low-Dose Intoxication: injected subcutaneously with ethyl paraoxon (0.658 mg/kg, 2×LD50), followed one minute later by intravenous administration of either ANG-OPHDS5 or OPHDS5 (equimolar doses containing 120 µg/kg OPHDS5). These groups were designated as the ANG-OPHDS5 (2×LD50) group and the OPHDS5 (2×LD50) group. (4–5) High-Dose Intoxication: injected subcutaneously with ethyl paraoxon (8×LD50), followed one minute later by intravenous administration of either ANG-OPHDS5 or OPHDS5 (equimolar doses containing 120 µg/kg OPHDS5). These groups were designated as the ANG-OPHDS5 (8×LD50) group and the OPHDS5 (8×LD50) group.
Serum samples were collected at two time points: one day prior to treatment initiation and 24 h after the final administration. Prior to blood collection, all rats were adequately anesthetized by inhalation of isoflurane (4% for induction, 1.5–2.0% for maintenance in oxygen) to minimize pain and distress. Approximately 0.5 mL of whole blood was drawn from the orbital venous plexus into a dry EP tube, allowed to clot at room temperature for 2 h, and centrifuged at 2000 × g for 10 minutes at 4 °C. The serum supernatant was analyzed for AChE activity, superoxide dismutase (SOD) activity, and reduced glutathione (GSH) content using commercial assay kits (Solarbio; AChE: Cat. No. BC2025, SOD: Cat. No. BC5165, GSH: Cat. No. BC1175) according to the manufacturer’s instructions.
Seven days after intoxication, all rats were deeply anesthetized via inhalation of isoflurane (4% for induction, 1.5–2.0% for maintenance in oxygen) before cardiac perfusion. The rats were subjected to cardiac perfusion with ice-cold PBS, and the whole brain tissue was removed. Complete perfusion resulted in irreversible animal death, and no additional euthanasia procedures were required. Each brain was weighed and homogenized on ice in pre-cooled medium at a 1:10 (w/v) ratio. The homogenate was centrifuged as per kit specifications, and the supernatant was collected. AChE activity, SOD activity, and GSH content in the brain homogenate were measured using the respective commercial kits, following the protocols precisely. All animal experimental procedures, including anesthesia, blood collection, and tissue harvesting, were performed in strict accordance with the American Veterinary Medical Association (AVMA) guidelines for the humane care and use of laboratory animals.
Apoptosis Analysis of Brain Tissue
Twenty-four hours after the last administration, all rats in each group were deeply anesthetized via inhalation of isoflurane (4% for induction, 1.5–2.0% for maintenance in oxygen) prior to transcranial perfusion. Rats were transcranially perfused with 200 mL of pre-cooled normal saline followed by 200 mL of 4% paraformaldehyde (PFA). Complete perfusion fixation resulted in irreversible animal death, and no additional euthanasia procedures were required. The whole brains were removed, post-fixed in 4% PFA for 24 h, and processed for paraffin embedding following a standard dehydration, clearing, and embedding procedure. Paraffin-embedded tissues were coronally sectioned at 5 μm thickness.
For TUNEL staining, sections were dewaxed in xylene, rehydrated through a graded ethanol series, and subjected to antigen retrieval by incubation with 20 μg/mL proteinase K (Servicebio, Cat. No. G1205) at 37°C for 20 minutes. After three washes with PBS, 50 μL of TUNEL reaction mixture was applied to each section, followed by incubation at 37 °C in the dark for 60 minutes. Sections were washed again with PBS and mounted with anti-fade mounting medium containing DAPI. Images were captured using a fluorescence microscope. Apoptotic cells were identified by green fluorescence, while nuclei were counterstained blue with DAPI. All animal experimental procedures, including anesthesia and tissue harvesting, were performed in strict accordance with the American Veterinary Medical Association (AVMA) guidelines for the humane care and use of laboratory animals.
Behavioral Testing
Behavioral tests were conducted in a sound-attenuated room under consistent lighting. Adult Sprague-Dawley rats were acclimatized in a holding area adjacent to the testing room one day before testing. On the first test day, each rat was allowed to freely explore the empty arena for 10 minutes to reduce novelty stress. All animal procedures during behavioral testing were performed in strict accordance with the American Veterinary Medical Association (AVMA) guidelines for the humane care and use of laboratory animals.
For the open field test, a rat was gently positioned facing the wall at the center of the arena, and locomotor activity was video-recorded for 10 minutes. The arena was thoroughly cleaned with 75% ethanol between trials to eliminate odor and excretory cues from the previous animal. The ratio of distance traveled in the center zone to the total distance traveled was calculated as an indicator of anxiety-like behavior.
For the novel object recognition (NOR) test, rats explored two identical objects (cylinders) during the first trial (T1), and exploration of each object was recorded for 10 minutes. After a 2 h interval, one familiar object was replaced with a novel object (cone) for the second trial (T2), and exploration was recorded for another 10 minutes. Recognition memory was assessed using two indices: the discrimination index (DI) and the recognition index (RI). The DI was calculated as (Tnew - Tfam)/(Tnew + Tfam) and the RI was calculated as Tnew/(Tnew + Tfam), where Tnew and Tfam represent exploration time of the novel and familiar object, respectively. A DI > 0 and an RI > 0.5 were considered indicative of intact recognition memory. After completion of all behavioral tests, all rats were humanely euthanized by deepening isoflurane inhalation (maintenance concentration increased to 5% in oxygen) until respiratory and cardiac arrest, in compliance with AVMA guidelines for laboratory animal euthanasia.
Biocompatibility Assessment of ANG-OPHDS5
Adult Sprague-Dawley rats were randomly divided into two groups (n = 3): saline control group (daily subcutaneous injection of normal saline) and treatment group (daily subcutaneous injection of ANG-OPHDS5, 131 μg/kg, equivalent to 120 μg/kg OPHDS5 protein). Both groups were treated for three consecutive days. Twenty-four hours after the final dose, all rats were adequately anesthetized by inhalation of isoflurane (4% for induction, 1.5–2.0% for maintenance in oxygen) prior to blood collection. Blood was collected from the orbital venous plexus. One aliquot anticoagulated with EDTA-K2 was used for routine hematological analysis (white blood cell, red blood cell, platelet). The remaining blood was clotted and centrifuged to obtain serum for biochemical profiling of liver function (alanine aminotransferase, aspartate aminotransferase), kidney function (creatinine, blood urea nitrogen), and metabolic indices (glucose, total protein).
Following blood collection, the rats were kept under deep isoflurane anesthesia and subjected to transcardial perfusion with ice-cold normal saline. Complete perfusion resulted in irreversible animal death. The heart, liver, spleen, lungs, and kidneys were harvested, fixed in 4% PFA for 24 h, and processed for paraffin embedding. Sections (5 μm) were stained with hematoxylin and eosin (H&E) and examined under a light microscope for morphological assessment. All animal experimental procedures, including anesthesia, blood collection, transcardial perfusion and tissue harvesting, were performed in strict accordance with the American Veterinary Medical Association (AVMA) guidelines for laboratory animal care and use.
Statistical Analysis
Statistical analysis was performed via two-tailed paired Student’s t test or one-way/two-way ANOVA with Dunnett’s/Sidak’s multiple comparisons test. All data were analyzed by GraphPad Prism 9.5.0 and presented as means ±standard deviation (SD) with significance defined as P< 0.05 with 95% confidence intervals.
Results
Design and Screening of Brain-Targeting Fusion Proteins
We employed genetic engineering to design coding sequences for six distinct fusion brain-targeting proteins (Table S1). These sequences were ligated into expression vectors to generate the corresponding plasmids (Figure S1). The core design of these recombinant proteins involves fusing OPHDS5 with Angiopep-2 via specific amino acid linkers. The linkers used are flexible, hydrophilic, and unstructured sequences rich in glycine (G) and serine (S), SGGRGGA and GGGGSGGGGSGGGGS.19,20 Following DNA sequencing and small-scale expression in Escherichia coli, six recombinant fusion proteins were successfully produced (Figure 1a). Among them, ANG-OPHDS5, OPHDS5-ANG, His-ANG-OPHDS5 and ANG-OPHDS5-His employed SGGRGGA as linker, while His-ANG-3L-OPHDS5 and ANG-3L-OPHDS5-His used GGGGSGGGGSGGGGS as linker. Preliminary screening based on the soluble protein expression identified ANG-OPHDS5 as a promising candidate. All recombinant fusion proteins were confirmed by SDS-PAGE to match their theoretical molecular weights (Figure 1b).
Preparation and Characterization of ANG-OPHDS5
Native polyacrylamide gel electrophoresis (Native‑PAGE) revealed that ANG‑OPHDS5 migrated at a higher apparent molecular weight than unmodified OPHDS5, with an increase of approximately 7 kDa consistent with theoretical predictions (Figure 1c). Qualitative proteomic analysis by HPLC‑MS/MS confirmed the presence of the ANG sequence at the N‑terminus (Figure S2). Database matching and theoretical fragment‑ion analysis of the MS/MS spectra demonstrated high consistency between the observed peptide masses and the expected sequences, verifying successful expression of the recombinant fusion protein (Figure 1d).
Both wild‑type OPH and its engineered variant OPHDS5 predominantly exist as dimers in solution. Analytical ultracentrifugation (AUC) analysis showed that ANG‑OPHDS5 also adopts a similar dimeric state (Figure 1e). The observed lower-than-theoretical molecular weight phenomenon may be attributed to the N-terminal ANG fusion, which likely induces a more non-spherical protein conformation, thereby hindering its movement through the centrifugal matrix and altering its sedimentation properties. This inference aligns with the general principles governing protein dynamics and interactions in solution.21
The fusion enzyme exhibited favorable stability in solution at 4 °C, following a slightly lower initial activity compared to OPHDS5, with no significant further loss over 8 weeks(Figure 1f). Circular dichroism spectroscopy indicated that this activity change was accompanied by a conformational shift from α‑helix to β‑sheet (Figures 1g and S3).
To clarify the structural mechanism underlying the altered enzymatic activity, molecular dynamics simulations were performed in this study. The root-mean-square deviation (RMSD) results of OPHDS5 (Figure 1h) revealed that its overall structure remained stable in the early simulation stage. Root-mean-square fluctuation (RMSF) analysis (Figure 1i) indicated low conformational flexibility of amino acid residues in OPHDS5. Radius of gyration (Rg) measurement (Figure 1j) further confirmed high structural compactness of OPHDS5. In contrast, the RMSD profile of ANG-OPHDS5 (Figure 1k) exhibited distinct structural rearrangement at the initial stage. RMSF analysis (Figure 1L) identified prominent conformational flexibility in the N-terminal fused ANG segment, suggesting that this region is intrinsically disordered. Meanwhile, ANG-OPHDS5 presented markedly higher Rg values and fluctuations (Figure 1m), which was attributed to the flexible N-terminal extension. Moreover, the Rg variation trends were highly consistent with RMSD changes within the first 100 ns. Collectively, molecular dynamics simulations verified that ANG-OPHDS5 possessed reduced structural stability and enhanced molecular dynamics, and the disordered and flexible N-terminal region dominated its unique structural characteristics. Taken together, these structural analyses suggest a mechanistic model: The flexible ANG sequence likely interacts with the protein core, inducing conformational changes. Critically, this includes a shift in the side chains of the histidine residues that coordinate the catalytic zinc ion (Figure 1n). This displacement disrupts the zinc coordination geometry, compromises the structural integrity of the enzyme’s active site, and thereby accounts for the observed alteration in enzymatic activity.
Brain Targeting of ANG-OPHDS5
In order to verify the brain-targeting capability of the recombinant fusion protein ANG-OPHDS5, we labeled ANG-OPHDS5 and OPHDS5 with Cy5.5 dye for in vivo and in vitro evaluations (Figure 2a). Immunohistochemical assessment of the hippocampal and medullary oblongata regions collected 50 minutes after administering Cy5.5-ANG-OPHDS5 to C57BL/6N mice revealed that low-density lipoprotein receptor 1 was associated with the cerebral vasculature (delineated by CD31), while Cy5.5-ANG-OPHDS5 exhibited overlap with the vascular system, indicating the strong potential of ANG-OPHDS5 for targeted delivery to brain regions (Figure 2b). Concurrently, this provides a potential avenue for the emergency treatment of OP poisoning, as existing research indicates that early mortality from such poisoning is due to inhibition of the medullary respiratory center.22 Subsequently, we evaluated the distribution of ANG-OPHDS5 and OPHDS5 within the brain and assessed their in vivo blood-brain barrier penetration capability. Quantitative analysis of the time‑course fluorescence images (Figure 2c and d) revealed that at 25 min post‑injection, ANG‑OPHDS5 produced significantly higher brain fluorescence intensity than OPHDS5, with mean values of 8.70 ± 0.37 × 109 (n = 3) for ANG‑OPHDS5 and 7.63 ± 0.49 × 109 (n = 3) for OPHDS5 (p = 0.039). At other time points (15, 50, and 100 min), ANG‑OPHDS5 showed consistently higher mean values than OPHDS5, but the differences were not statistically significant (all p > 0.05). Fluorescence distribution in other parts of the body was similar between the two groups. Flow cytometry analysis indicated that the uptake rate of Cy5.5-labeled ANG-OPHDS5 by bEnd.3 cells was significantly higher than that of OPHDS5 (Figure 2e), reaching its peak at 2 h (Figure 2f). These results demonstrate that ANG-OPHDS5 possesses favorable central nervous system targeting properties.
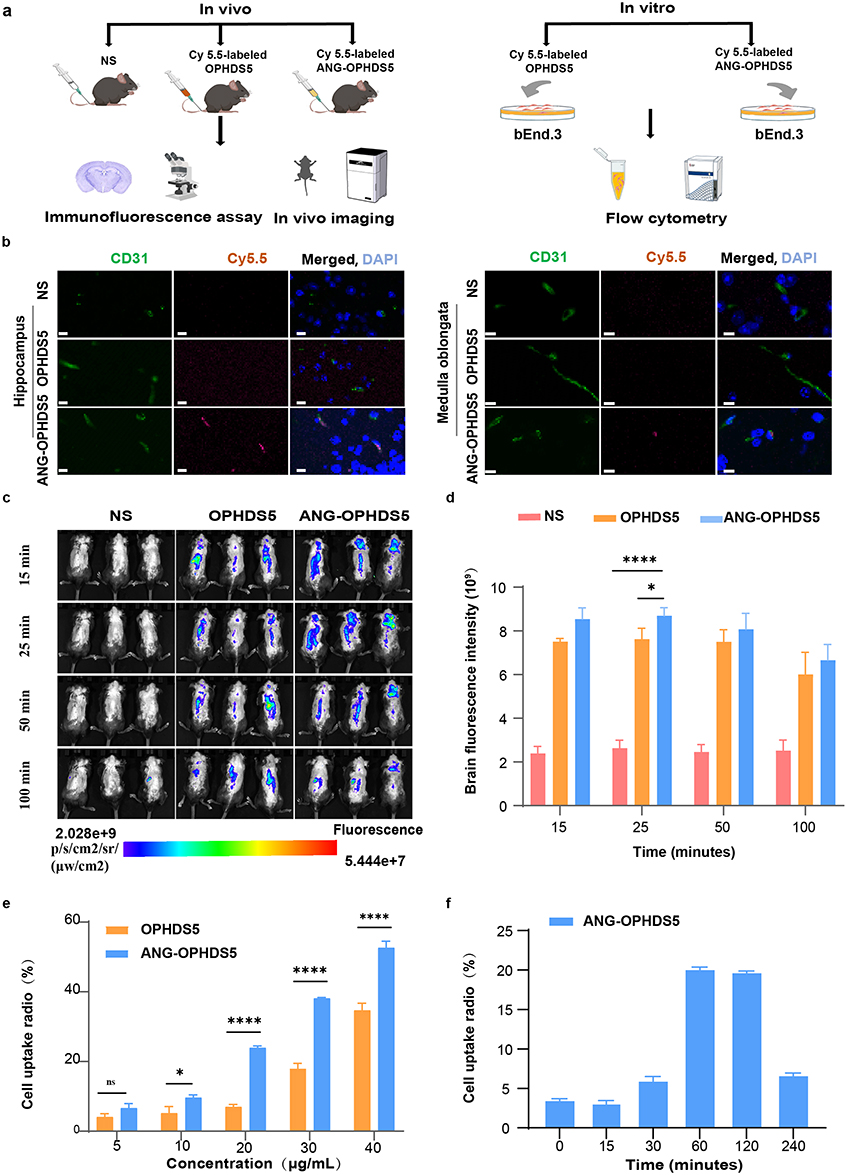 |
Figure 2 Evaluation of the brain‑targeting potential of ANG‑OPHDS5 in vivo and in vitro (NS, normal saline). (a) Schematic of the experimental workflow for evaluating the central nervous system targeting efficacy of Cy5.5‑labeled OPHDS5 and ANG‑OPHDS5 (Cy5.5, a fluorescent labeling reagent). (b) Representative image from a single experiment qualitatively showing co‑localization of ANG‑OPHDS5 (Cy5.5, red) with blood vessels (CD31, a vascular endothelial cell marker, green) in the brain at 50 minutes post‑systemic delivery in 6‑week‑old male C57BL/6N mice. Scale bar, 10 μm. (c) In vivo fluorescence images comparing the distribution of Cy5.5‑labeled ANG‑OPHDS5 and OPHDS5 in mice at 15, 25, 50, and 100 minutes after intravenous injection. (d) Quantification of brain‑region fluorescence signals from in vivo imaging at 15, 25, 50, and 100 minutes (n =3). (e) Flow cytometry analysis of the concentration‑dependent cellular uptake of Cy5.5‑labeled ANG‑OPHDS5 and OPHDS5 in bEnd.3 cells (n = 3). (f) Time‑dependent cellular uptake of Cy5.5‑labeled ANG‑OPHDS5 (20 μg/mL) in bEnd.3 cells analysed by flow cytometry (n=3). Data were analysed by one‑way/two‑way ANOVA with Dunnett’s/Sidak’s multiple‑comparisons test. All data are presented as mean ± S.D. ns, no statistical difference, *P < 0.05, ****P < 0.0001. |
Dual Benefits of ANG-OPHDS5: Antagonizing Lethal Ethyl Paraoxon Toxicity and Mitigating Central Nervous System Injury
We established an acute OP poisoning model by subcutaneous injection of ethyl paraoxon into the posterior neck of rats, simulating a clinical emergency rescue scenario after poisoning (Figure 3a). The median lethal dose (LD50) of ethyl paraoxon on SD rats was measured at 0.329 mg/kg by our lab.1 The experiment employed absolute lethal doses (2×LD50) and high lethal doses (8×LD50, 20×LD50) for exposure to comparatively evaluate the detoxification efficacy of ANG-OPHDS5 and OPHDS5. When the exposure dose of paraoxon increased from 2×LD50 to 20×LD50, the lethal time of intoxicated rats was rapidly reduced from approximately 11 minutes to 3 minutes, with obvious toxic symptoms appearing as early as 1 minute post-exposure. One minute after administering an absolutely lethal dose of ethyl paraoxon via subcutaneous injection, we immediately delivered either ANG-OPHDS5 or OPHDS5 solution (equimolar doses containing 120 µg/kg OPHDS5) through tail vein injection. We then continuously monitored the survival and toxic symptoms of the rats for 1 h and assigned standardized poisoning severity scores based on Table S2. The above treatment was repeated over the subsequent two days, and the body temperature and body weight of the rats were monitored throughout the period. No mortality happened in any group of rats. Symptom scores (Figure 3b) revealed that at equivalent exposure doses, the ANG-OPHDS5 group had lower scores than the OPHDS5 group, indicating milder toxic injury symptoms. Particularly at the high lethal dose of 8×LD50, animals in the OPHDS5 treatment group commonly exhibited symptoms of central nervous system impairment such as bradykinesia, whereas the ANG-OPHDS5 group did not show signs of neural injury. These results indicate that ANG-OPHDS5 administration is effective not only for peripheral tissues but also exhibits a good therapeutic effect on the central nervous system. All experimental groups of rats exhibited a dynamic body weight pattern post-exposure: a sharp initial increase (within 24 hours), followed by a slight decline (Day 2), and then another significant increase (Days 3–4) (Figure 3c). The early weight fluctuation may be related to the acute systemic inflammatory response and metabolic stress induced by poisoning.23 It is noteworthy that the amplitude of body weight change was generally greater in the OPHDS5 treatment group than in the ANG-OPHDS5 group throughout the observation period. The sustained weight gain in the later stage might be associated with trace residual organophosphorus compounds promoting the conversion of brown adipose tissue to white adipose tissue.24
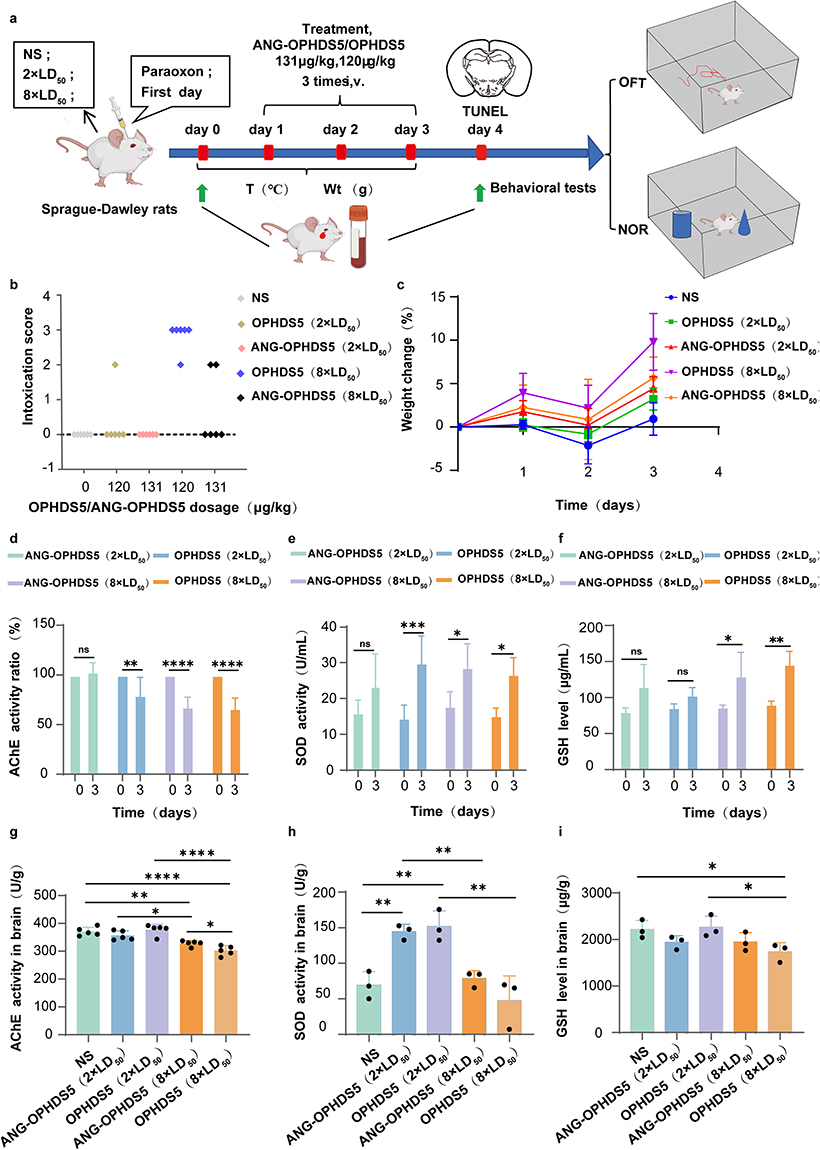 |
Figure 3 Acute rescue of subcutaneously intoxicated SD rats by ANG‑OPHDS5 and OPHDS5 (NS, normal saline). (a) Sprague‑Dawley rats received daily tail‑vein injections of ANG‑OPHDS5, OPHDS5, or NS for three consecutive days after ethyl paraoxon intoxication, followed by Open Field Test (OFT), novel‑object‑recognition (NOR) testing and tissue collection. (b) Intoxication score of rats following first aid treatment with ANG-OPHDS5 or OPHDS5 after intoxication with 2 × LD50 or 8 × LD50 of ethyl paraoxon, observation within 1 h (n = 6 biologically independent animals). (c) Body‑weight changes in rats intoxicated with lethal‑dose ethyl paraoxon and treated with ANG‑OPHDS5 or OPHDS5 (n = 6 biologically independent animals). (d) Post‑/pre‑treatment ratio of serum acetylcholinesterase (AChE) activity in rats treated with ANG‑OPHDS5 or OPHDS5 (n = 6 biologically independent animals). (e) Post‑/pre‑treatment ratio of serum superoxide dismutase (SOD) activity in rats treated with ANG‑OPHDS5 or OPHDS5 following three consecutive days of treatment (n = 6 biologically independent animals). (f) Post‑/pre‑treatment ratio of serum reduced glutathione (GSH) level in rats treated with ANG‑OPHDS5 or OPHDS5 for three consecutive days (n = 4 biologically independent animals). (g) Brain acetylcholinesterase (AChE) activity on day 7 post‑intoxication in rats treated with ANG‑OPHDS5 or OPHDS5 (n = 6 biologically independent animals). (h) Brain superoxide dismutase (SOD) activity on day 7 post‑intoxication in rats treated with ANG‑OPHDS5 or OPHDS5 (n = 3 biologically independent animals). (i) Brain reduced glutathione (GSH) level on day 7 post‑intoxication in rats treated with ANG‑OPHDS5 or OPHDS5 (n = 3 biologically independent animals). Statistical significance was determined using a paired two‑tailed t‑test (d–f) or one‑way ANOVA with Dunnett’s multiple comparisons test. Data are presented as mean ± S.D. ns, no statistical difference, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. |
Post-treatment blood biochemical analysis revealed that at the 2×LD50 dose, ANG-OPHDS5 treatment effectively restored AChE activity, whereas in all other treatment groups, enzyme activity remained at 50%–75% of baseline levels, within the range indicative of mild intoxication (Figure 3d). All groups showed a significant increase in SOD activity (Figure 3e), suggesting activation of the body’s antioxidant system, accompanied by a concurrent rise in GSH levels (Figure 3f), indicating that the body had entered a compensatory stress stage after the 3-day treatment. Next, we indirectly assessed the clearance of centrally present free ethyl paraoxon by measuring AChE activity in brain tissue 7 days post-exposure. The results showed that compared to normal saline control group, there was no significant difference in brain AChE activity in the 2×LD50 exposure group, while a decrease was observed in the high-dose (8×LD50) exposure groups. Furthermore, compared to the ANG-OPHDS5 group, the OPHDS5 treatment group exhibited a more pronounced reduction in AChE activity (Figure 3g), demonstrating the protective effect of ANG-OPHDS5 on central AChE. Brain SOD levels in the 8×LD50 exposure group were lower than in the 2×LD50 group (Figure 3h), indicating consumption of SOD due to the intense oxidative stress response that occurred in the brain post-exposure. The difference in brain GSH levels between the 8×LD50 groups reflects the advantage of ANG-OPHDS5 in mitigating local oxidative stress in the brain (Figure 3i).
TUNEL staining was performed on brain tissue to assess neuronal damage following rescue therapy. The results demonstrated that after three treatment cycles, rats treated with ANG‑OPHDS5 exhibited a lower level of neuronal apoptosis in the hippocampus and surrounding regions compared to those in the OPHDS5 group, under both 2× LD50 and 8× LD50 exposure doses (Figure 4a). When the exposure dose was increased to 20× LD50, this protective difference became more pronounced, with the ANG‑OPHDS5 group showing a significantly further reduction in hippocampal neuronal apoptosis (Figure 4b and c).
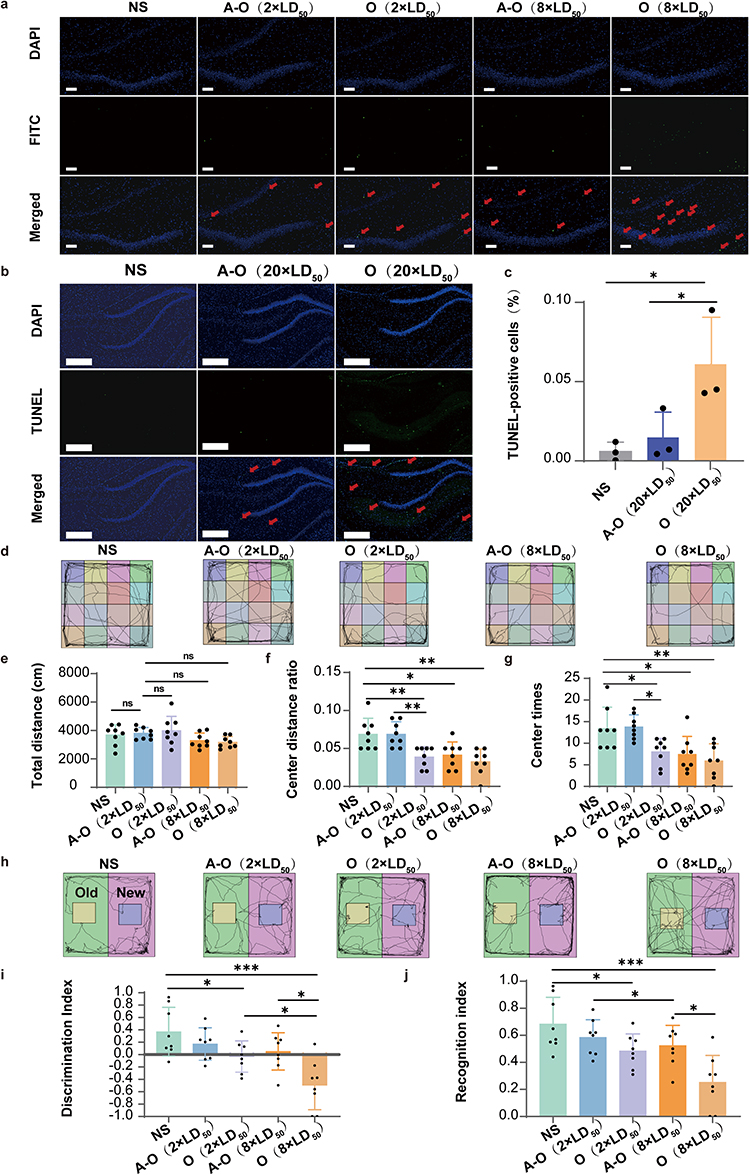 |
Figure 4 Neuroprotective properties of ANG-OPHDS5 (O represents OPHDS5, A-O represents ANG-OPHDS5, NS, normal saline). (a) Representative TUNEL staining images of the hippocampal region from rats intoxicated with 2×LD50 or 8×LD50 ethyl paraoxon and treated with the indicated proteins. Red arrows indicate TUNEL-positive cells. Scale bar, 100 μm. (b) Representative TUNEL staining images of the hippocampal region from rats intoxicated with 20×LD50 and treatment. Red arrows indicate TUNEL-positive cells. (c) Quantitative analysis of neuronal apoptosis levels in the hippocampal region following intoxication at 20×LD50 and treatment (n = 3 biologically independent animals). Scale bar, 200 μm. (d) Representative locomotion traces from the open-field test (OFT). (e) Total movement distance of experimental rats in each group. (n = 8 biologically independent animals). (f) Ratio of distance travelled in the centre zone to total distance travelled in the OFT (n = 8 biologically independent animals). (g) Number of entries into the centre zone in the OFT (n = 8 biologically independent animals). (h) Representative locomotion traces from the novel-object-recognition (NOR) test. (i) Discrimination index in the NOR test (n = 8 biologically independent animals). (j) Recognition index in the NOR test (n = 8 biologically independent animals). Data were analysed by one-way ANOVA with Dunnett’s multiple-comparisons test. All data are presented as mean ± S.D. ns, no statistical difference, *P < 0.05, **P < 0.01, ***P < 0.001. |
To compare the central protective effects of ANG-OPHDS5 and OPHDS5, we employed behavioral assays relevant to central neuronal damage. In the open field test, heterogeneous locomotor trajectory profiles were observed among all experimental groups. Rats in the normal saline and ANG-OPHDS5 protein-treated groups displayed a distinct predilection for central zone crossing compared with the OPHDS5-treated group (Figure 4d). For total distance traveled, a declining trend was observed in both 2×LD50 and 8×LD50 poisoned groups, with no statistically significant intergroup variation (Figure 4e), suggesting that basal locomotor capacity was not significantly modulated by poisoning dosage or protein rescue regimen, which could be explained by the pronounced peripheral rescue effects of ANG-OPHDS5 and OPHDS5.
Despite the lack of significant divergence in overall locomotor output, quantitative assessment of central zone exploration uncovered clear distinct OP dose-dependent disparities between the two protein-treated groups. At the 2×LD50 exposure dose, ANG-OPHDS5-treated rats exhibited significantly increased central zone entry frequency and prolonged central residence relative to the OPHDS5 group, corresponding to reduced anxiety-like behaviors and augmented exploratory drive. Nevertheless, this beneficial distinction was abrogated when the poisoning dose was elevated to 8×LD50 (Figure 4f and g), which may be attributed to the overwhelming organophosphorus burden within the central nervous system. These results collectively indicate that ANG-OPHDS5 affords superior alleviation of anxiety-like behaviors following rescue from 2×LD50 acute poisoning, conferring a favorable restorative effect on rat emotional state.
The novel object recognition (NOR) test revealed distinct spatial patterns in representative exploratory trajectories across experimental groups (Figure 4h). Rats in the normal saline group exhibited dense trajectory coverage around the novel object (purple area), demonstrating a clear exploratory preference for the novel stimulus and confirming intact recognition memory. In the 2×LD50 exposure cohorts, the ANG-OPHDS5 protein-rescued group displayed more frequent exploration of the novel object compared to OPHDS5 protein-rescued group. At the 8×LD50 exposure level, trajectories of OPHDS5 protein-rescued group were largely confined to the familiar object (yellow area) and arena periphery, with negligible exploration of the novel object, whereas ANG-OPHDS5 protein-rescued group retained partial exploratory behavior toward the novel stimulus.
Quantitative analysis of the discrimination index (DI) showed that the NS group exhibited a significantly positive DI, confirming the ability to distinguish between novel and familiar objects. Following 2×LD50 exposure, the DI of ANG-OPHDS5 was significantly higher than that of OPHDS5 group and remained above zero, indicating partial preservation of recognition memory. At 8×LD50 exposure, the DI of OPHDS5 group was significantly negative, reflecting a complete loss of novel object recognition, while ANG-OPHDS5 group maintained a DI near zero, suggesting residual recognition memory function (Figure 4i). Consistent with DI results, the recognition index was highest in the NS group, indicating a strong preference for novel object exploration. In the 2×LD50 exposure groups, ANG-OPHDS5 group exhibited a significantly higher recognition index than the OPHDS5 group. At 8×LD50 exposure, both groups showed a marked reduction in recognition index, but ANG-OPHDS5 group still maintained a significantly higher index than OPHDS5 group (Figure 4j).
Safety Assessment of ANG-OPHDS5
To assess the in vitro safety profile of ANG‑OPHDS5, its cytotoxicity was evaluated against mouse brain capillary endothelial cells (bEnd.3) by co‑incubation for 1 h (Figure 5a). The CCK‑8 assay showed no significant difference in cell viability compared to the control group, indicating no apparent cytotoxicity (Figure 5b). After 7 days of tail vein administration of a therapeutic dose of ANG‑OPHDS5 to rats, major organs and blood samples were collected for analysis (Figure 5c). The results showed no statistically significant differences in hematological and blood biochemical parameters compared to the saline control group (Figure 5d–e). Furthermore, histopathological examination of the major organs revealed no abnormal alterations or signs of toxicity (Figure 5f). Collectively, these findings demonstrate that ANG‑OPHDS5 exhibits promising safety under the tested experimental conditions.
 |
Figure 5 Safety assessment of ANG‑OPHDS5 (NS, normal saline). (a) Experimental workflow of the ANG‑OPHDS5 cytotoxicity assay in bEnd.3 cells. (b) Cytotoxicity of ANG‑OPHDS5 after 24h treatment at various concentrations (n = 6 biologically independent samples). (c) Schematic of the experimental timeline for repeated administration and sample collection. (d) Analysis of key hematological parameters in rats following ANG‑OPHDS5 administration. (e) Analysis of key serum biochemical parameters in rats following ANG‑OPHDS5 administration. (n = 3 biologically independent animals). (f) Representative haematoxylin and eosin (H&E) stained images of major organs (n = 3 biologically independent animals). Scale bar, 20 μm. Data were analysed by one‑way ANOVA with Dunnett’s multiple‑comparisons test. All data are presented as mean ± S.D. ns, no statistical difference. |
Discussion
Studies indicate that following poisoning from either a single acute exposure or repeated subclinical exposure to organophosphates, patients often experience delayed neuronal damage and neurobehavioral abnormalities even after effective emergency treatment.25 This is primarily because conventional antidotes fail to effectively cross the BBB and thus cannot clear the persistently present toxic molecules within the CNS. Current brain-targeted therapeutics for organophosphate poisoning are predominantly AChE reactivators.26,27 However, these drugs do not fundamentally address the persistence of organophosphates in the CNS, thereby allowing for the continued progression of neuronal damage.28 Consequently, the development of etiology-targeted, brain-focused therapeutic agents holds significant clinical importance and research value. Unlike conventional AChE reactivators and single symptomatic antidotes, the newly constructed ANG-OPHDS5 possesses precise blood-brain barrier penetration capability mediated by LRP1 receptor binding. It fundamentally eliminates residual organophosphate toxins in the CNS via enzymatic hydrolysis, rather than merely relieving cholinergic symptoms. On this basis, ANG-OPHDS5 further alleviates OP-triggered oxidative stress and secondary neuronal apoptosis, achieving dual effects of acute detoxification and long-term neuroprotection. This etiology-driven therapeutic strategy provides a distinct and superior alternative to existing reported antidote compounds.
This study constructed a novel fusion protein with CNS targeting capability, which can be expressed using E. coli BL21. The fusion protein leverages the high affinity of ANG for the LRP1 receptor on brain capillary endothelial cells to deliver the broad-spectrum organophosphorus hydrolase OPHDS5 to the CNS. A linker composed of flexible glycine and rigid serine residues was incorporated, which maintained the functionality of both protein domains while enhancing the soluble expression of the recombinant fusion protein.29 In an acute organophosphorus compound poisoning model, subcutaneous injection of a highly lethal dose (8×LD50) of ethyl paraoxon into the nape of the neck resulted in no mortality among rats treated with either ANG-OPHDS5 or OPHDS5. Compared to the OPHDS5 treatment group, the ANG-OPHDS5 group exhibited only mild peripheral intoxication symptoms. Furthermore, the recovery of acetylcholinesterase activity in brain tissue 7 days post-intoxication was significantly more pronounced in the ANG-OPHDS5 group, demonstrating its superior advantage in CNS-targeted therapy.
Studies have shown that exposure to OPs can lead to increased generation of reactive oxygen species and reactive nitrogen species, which induces lipid peroxidation, protein oxidation, and DNA damage, ultimately triggering cell apoptosis and neuroinflammation.30,31 In this experiment, at the 8×LD50 dose, the GSH content in the brain tissue of the OPHDS5 group was lower than that of the control group, whereas no significant difference was observed between the ANG-OPHDS5 group and the control group. The maintenance of normal GSH levels in the brain tissue of the ANG-OPHDS5 group indicates less depletion of antioxidant reserves, demonstrating the advantage of ANG-OPHDS5 in mitigating oxidative stress in brain tissue. Results from TUNEL staining suggest that ANG-OPHDS5 can more effectively protect hippocampal and surrounding neurons, particularly under high intoxication doses. HE and TUNEL results indicated that ANG-OPHDS5 more effectively protected hippocampal and surrounding neurons, particularly at high intoxication doses. Based on these indirect markers (AChE activity, oxidative stress, TUNEL, and behavior), we infer that ANG-OPHDS5 likely reduces the interaction of organophosphorus compounds with neural cells and may facilitate the clearance of CNS‑resident OPs. This significantly lowers neuronal oxidative stress, ultimately leading to the improved neural function seen in behavioral tests. This result signifies important progress in the development of next-generation antidotes against nerve agents and highly toxic pesticides, offering the potential to overcome the limitations of certain oxime drugs.
We have also noted that various bioactive compounds have been investigated for their neuroprotective effects against neurological diseases, often by targeting neuroinflammation and oxidative stress, two interconnected pathological processes that are also central to OP‑induced CNS injury. For instance, Astragaloside IV, a saponin from Astragalus membranaceus, attenuates neuronal damage in cerebral ischemia, Parkinson’s disease, and Alzheimer’s disease through anti‑inflammatory and anti‑apoptotic mechanisms. Unlike organophosphorus hydrolase (OPH), which directly hydrolyzes OP molecules, Astragaloside IV exerts its effects by modulating endogenous signalling pathways rather than directly detoxifying the toxicant.32 Similarly, chinonin, a natural xanthone from Anemarrhena asphodeloides, counteracts neuroinflammation and protects against cerebral ischemia and neuropathic pain by suppressing oxidative stress and microglial activation.33 The therapeutic value of suppressing neuroinflammation extends beyond natural products. For example, Apelin‑13 has been shown to reduce cognitive decline in Alzheimer’s disease by modulating microglial polarization and lowering pro‑inflammatory cytokines.34 Moreover, the critical role of inflammation in CNS injury is further underscored by autoimmune encephalitis, a prototypical immune‑mediated neurotoxic process that directly targets brain tissue.35 Collectively, these studies highlight that mitigating neuroinflammation and oxidative stress is a promising strategy across various neurological disorders, including OP poisoning.
A key signalling cascade central to neuroinflammation in many neurological disorders is the TLR4/NF‑κB pathway. Activation of TLR4 triggers NF‑κB nuclear translocation, promoting transcription of pro‑inflammatory mediators that exacerbate neuronal damage.36 This pathway is also implicated in OP‑induced neurotoxicity, where it links OP exposure to downstream oxidative stress and neuronal injury. Given the pivotal role of this cascade, therapeutic strategies that mitigate its pathological consequences, particularly oxidative stress and neuronal loss, hold great promise. Our results demonstrate that ANG‑OPHDS5 hydrolyzes OPs directly, thereby counteracting brain oxidative stress and reducing neuronal injury following OP poisoning, highlighting its neuroprotective potential.
It should be noted that in the in vivo imaging experiment, a statistically significant difference in brain fluorescence between ANG‑OPHDS5 and OPHDS5 was observed only at 25 min post‑injection, but not at 15, 50, or 100 min. This may be attributed to several factors, including the relatively low administered protein dose (~168 μg/kg), which limited the dynamic range; the fact that whole‑brain fluorescence includes signal from the blood pool, thereby contributing to the fluorescence detected in the OPHDS5 group; and the small sample size (n = 3 per group), which reduced statistical power. Nevertheless, the significantly higher cellular uptake of ANG‑OPHDS5 (Figure 2e and f) and its specific vascular co‑localization (Figure 2b) collectively support its brain‑targeting capability, with the advantage being most evident at the early time point.
Equally importantly, limitations related to the in vitro cellular uptake assay of ANG-OPHDS5 in bEnd.3 cells should be acknowledged. As a typical cell-penetrating peptide, ANG is widely recognized to facilitate transmembrane delivery through endocytic pathways; however, the detailed internalization process of ANG-OPHDS5 in bEnd.3 cells, as well as the specific membrane receptors governing this uptake (e.g., LRP1), have not been fully clarified in the present work. This is primarily because the core focus of this manuscript is to characterize the brain-targeted distribution, detoxification capacity, and neuroprotective effects of the recombinant ANG-OPHDS5, rather than to systematically explore its cellular uptake mechanism. In subsequent research, we will perform Western blotting and other molecular biological methods to identify and validate the expression and functional roles of specific receptors such as LRP1, so as to systematically elucidate the underlying molecular mechanism of ANG-OPHDS5 internalization.
Furthermore, given the well‑established role of the TLR4/NF‑κB signalling cascade in OP-triggered neuroinflammation and neuronal damage, future studies should further explore whether the neuroprotective efficacy of brain-targeted ANG‑OPHDS5 is mediated by the inhibition of this inflammatory pathway. Elucidating the regulatory relationship between ANG‑OPHDS5 and the TLR4/NF‑κB axis will deepen our mechanistic understanding of how direct intracerebral OP detoxification mediated by ANG‑OPHDS5 alleviates neuroinflammation and oxidative stress.
Admittedly, the receptor-mediated transport pathway is also subject to a binding site barrier effect. Excessive binding to endothelial cells may result in substantial drug retention at the BBB itself rather than complete brain delivery;37,38 optimizing binding affinity is under consideration. Simultaneously, investigating specific alterations in receptor expression on the blood-brain barrier following OPs intoxication will be a focus of future work. Beyond the current evaluation using ethyl paraoxon, the broad‑spectrum detoxification efficacy of ANG‑OPHDS5 should be further validated in poisoning models induced by a wider range of organophosphorus pesticides (e.g., malathion, dimethoate, dichlorvos) and nerve agents (e.g., sarin, soman, VX).39 In addition, combination strategies with existing antidotes such as atropine and oxime reactivators can be explored to assess potential synergistic effects, thereby providing more therapeutic options for complex clinical poisoning scenarios.40 Meanwhile, systematic pharmacokinetic studies are needed to elucidate its distribution, metabolism, and elimination profiles in both peripheral and central compartments, with particular emphasis on quantitative evaluation of its blood‑brain barrier penetration efficiency and retention time within brain tissue.41
Conclusions
In the present study, with a focus on acute or chronic poisoning caused by organophosphorus pesticides in the environment, the engineered ANG-OPHDS5 fusion protein not only achieved potent peripheral rescue efficacy but also protected central organophosphorus target enzymes, reduced apoptosis in hippocampal neurons, and improved behavioral outcomes. This represents a novel biodetoxification strategy integrating targeting capability, catalytic function, and scalable production, which holds significant translational medical value.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Ma M, Zhai Y, Jin Q, et al. N-terminal PEGylation enhances organophosphorus hydrolase catalysis for a promising fast and long-acting prophylactic candidate. J Hazard Mater. 2025;488:137336. doi:10.1016/j.jhazmat.2025.137336
2. Alozi M, Rawas-Qalaji M. Treating organophosphates poisoning: management challenges and potential solutions. Crit Rev Toxicol. 2020;50(9):764–19. doi:10.1080/10408444.2020.1837069
3. Massey N, Vasanthi SS, Holtkamp C, et al. Mitigating organophosphate nerve agent, soman (GD), induced long-term neurotoxicity: saracatinib, a Src Tyrosine Kinase inhibitor, as a potential countermeasure. J Neuroinflammation. 2025;22(1):199. doi:10.1186/s12974-025-03520-5
4. Eddleston M. Novel clinical toxicology and pharmacology of organophosphorus insecticide self-poisoning. Annu Rev Pharmacol Toxicol. 2019;59:341–360. doi:10.1146/annurev-pharmtox-010818-021842
5. Qiao L, Du X, Wang H, Wang Z, Gao S, Zhao CQ. Research progress on the strategies for crossing the blood-brain barrier. Mol Pharm. 2024;21(10):4786–4803. doi:10.1021/acs.molpharmaceut.4c00447
6. Butola M, Nainwal N. Non-Invasive techniques of nose to brain delivery using nanoparticulate carriers: hopes and hurdles. AAPS Pharm Sci Tech. 2024;25(8):256. doi:10.1208/s12249-024-02946-z
7. Furtado D, Björnmalm M, Ayton S, Bush AI, Kempe K, Caruso F. Overcoming the blood-brain barrier: the role of nanomaterials in treating neurological diseases. Adv Mater. 2018;30(46):e1801362. doi:10.1002/adma.201801362
8. Khan AR, Liu M, Khan MW, Zhai G. Progress in brain targeting drug delivery system by nasal route. J Control Release. 2017;268:364–389. doi:10.1016/j.jconrel.2017.09.001
9. Lin J, Yu Z, Gao X. Advanced noninvasive strategies for the brain delivery of therapeutic proteins and peptides. ACS Nano. 2024;18(34):22752–22779. doi:10.1021/acsnano.4c06851
10. Wang C, Wang S, Xue Y, et al. Intravenous administration of blood-brain barrier-crossing conjugates facilitate biomacromolecule transport into central nervous system. Nat Biotechnol. 2025;43(11):1783–1789. doi:10.1038/s41587-024-02487-7
11. Pardridge WM. Blood-brain barrier delivery for lysosomal storage disorders with IgG-lysosomal enzyme fusion proteins. Adv Drug Deliv Rev. 2022;184:114234. doi:10.1016/j.addr.2022.114234
12. Sonoda H, Morimoto H, Yoden E, et al. A blood-brain-barrier-penetrating anti-human transferrin receptor antibody fusion protein for neuronopathic mucopolysaccharidosis II. Mol Ther. 2018;26(5):1366–1374. doi:10.1016/j.ymthe.2018.02.032
13. Oller-Salvia B, Sánchez-Navarro M, Giralt E, Teixidó M. Blood-brain barrier shuttle peptides: an emerging paradigm for brain delivery. Chem Soc Rev. 2016;45(17):4690–4707. doi:10.1039/C6CS00076B
14. Ahmad N, Shaikh MF, Ansari MT, Arulsamy A. Nanoformulations in epilepsy therapy - a systematic review of emerging strategies in status epilepticus. Daru. 2025;33(2):31. doi:10.1007/s40199-025-00575-w
15. Habib S, Singh M. Angiopep-2-Modified nanoparticles for brain-directed delivery of therapeutics: a review. Polymers. 2022;14(4):712. doi:10.3390/polym14040712
16. Roodveldt C, Tawfik DS. Directed evolution of phosphotriesterase from Pseudomonas diminuta for heterologous expression in Escherichia coli results in stabilization of the metal-free state. Protein Eng Des Sel. 2005;18(1):51–58. doi:10.1093/protein/gzi005
17. Zhao H, Brautigam CA, Ghirlando R, Schuck P. Overview of current methods in sedimentation velocity and sedimentation equilibrium analytical ultracentrifugation. Curr Protoc Protein Sci. 2013;71. Chapter 20:Unit20.12. doi:10.1002/0471140864.ps2012s71
18. Schuck P, Perugini MA, Gonzales NR, Howlett GJ, Schubert D. Size-distribution analysis of proteins by analytical ultracentrifugation: strategies and application to model systems. Biophys J. 2002;82(2):1096–1111. doi:10.1016/S0006-3495(02)75469-6
19. Chen X, Zaro JL, Shen WC. Fusion protein linkers: property, design and functionality. Adv Drug Deliv Rev. 2013;65(10):1357–1369. doi:10.1016/j.addr.2012.09.039
20. Shamriz S, Ofoghi H, Moazami N. Effect of linker length and residues on the structure and stability of a fusion protein with malaria vaccine application. Comput Biol Med. 2016;76:24–29. doi:10.1016/j.compbiomed.2016.06.015
21. Anufriev I, Hoeppener S, Nischang I. PEG-Lipids: quantitative study of unimers and aggregates thereof by the methods of molecular hydrodynamics. Anal Chem. 2023;95(28):10795–10802. doi:10.1021/acs.analchem.3c01999
22. Carey JL, Dunn C, Gaspari RJ. Central respiratory failure during acute organophosphate poisoning. Respir Physiol Neurobiol. 2013;189(2):403–410. doi:10.1016/j.resp.2013.07.022
23. Hotchkiss RS, Karl IE. The pathophysiology and treatment of sepsis. N Engl J Med. 2003;348(2):138–150. doi:10.1056/NEJMra021333
24. Wang B, Tsakiridis EE, Zhang S, et al. The pesticide chlorpyrifos promotes obesity by inhibiting diet-induced thermogenesis in brown adipose tissue. Nat Commun. 2021;12(1):5163. doi:10.1038/s41467-021-25384-y
25. Zou S, Wang Q, He Q, et al. Brain-targeted nanoreactors prevent the development of organophosphate-induced delayed neurological damage. J Nanobiotechnology. 2023;21(1):256. doi:10.1186/s12951-023-02039-2
26. Fan N, Li Q, Liu Y, Ma B, Li M, Yin D. Preparation of an HI-6-loaded brain-targeted liposomes based on the nasal delivery route and the evaluation of its reactivation of central toxic acetylcholinesterase. Eur J Pharm Sci. 2023;184:106406. doi:10.1016/j.ejps.2023.106406
27. Zhang Y, He J, Shen L, et al. Brain-targeted delivery of obidoxime, using aptamer-modified liposomes, for detoxification of organophosphorus compounds. J Control Release. 2021;329:1117–1128. doi:10.1016/j.jconrel.2020.10.039
28. Terry AV. Functional consequences of repeated organophosphate exposure: potential non-cholinergic mechanisms. Pharmacol Ther. 2012;134(3):355–365. doi:10.1016/j.pharmthera.2012.03.001
29. Van Rosmalen M, Krom M, Merkx M. Tuning the flexibility of glycine-serine linkers to allow rational design of multidomain proteins. Biochemistry. 2017;56(50):6565–6574. doi:10.1021/acs.biochem.7b00902
30. Meyer C, Thippeswamy T. Organophosphate chemical nerve agents, oxidative stress, and NADPH oxidase inhibitors: an overview. Int J Mol Sci. 2025;26(19):9313. doi:10.3390/ijms26199313
31. Pearson JN, Patel M. The role of oxidative stress in organophosphate and nerve agent toxicity. Ann N Y Acad Sci. 2016;1378(1):17–24. doi:10.1111/nyas.13115
32. Hu E, Li Z, Li T, et al. A novel microbial and hepatic biotransformation-integrated network pharmacology strategy explores the therapeutic mechanisms of bioactive herbal products in neurological diseases: the effects of Astragaloside IV on intracerebral hemorrhage as an example. Chin Med. 2023;18(1):40. doi:10.1186/s13020-023-00745-5
33. Hui Z, Lai-Fa W, Xue-Qin W, Ling D, Bin-Sheng H, Li JM. Mechanisms and therapeutic potential of chinonin in nervous system diseases. J Asian Nat Prod Res. 2024;26(12):1405–1420. doi:10.1080/10286020.2024.2371040
34. Luo H, Xiang Y, Qu X, et al. Apelin-13 suppresses neuroinflammation against cognitive deficit in a streptozotocin-induced rat model of Alzheimer’s disease through activation of BDNF-TrkB signaling pathway. Front Pharmacol. 2019;10:395. doi:10.3389/fphar.2019.00395
35. Huang L, Tan J, Lin P, et al. Autoimmune encephalitis followed by hemophagocytic lymph histiocytosis: a case report. Front Immunol. 2024;15:1383255. doi:10.3389/fimmu.2024.1383255
36. Zhou C, Yu Z, Chen T, et al. Tanshinone IIA attenuates cerebral-ischemia-reperfusion-induced neuroinflammation by inhibiting the TLR4/NF-κB signaling cascade: a study integrating network pharmacology, bioinformatics, and experimental validation. Phytomedicine. 2025;149:157548. doi:10.1016/j.phymed.2025.157548
37. Petersen I, Morrison JI, Petrovic A, et al. A shorter linker in the bispecific antibody RmAb158-scFv8D3 improves TfR-mediated blood-brain barrier transcytosis in vitro. Sci Rep. 2024;14(1):30613. doi:10.1038/s41598-024-83627-6
38. Rosa A, Metzendorf NG, Efverström J, et al. Lowering the affinity of single-chain monovalent BBB shuttle scFc-scFv8D3 prolongs its half-life and increases brain concentration. Neurotherapeutics. 2025;22(1):e00492. doi:10.1016/j.neurot.2024.e00492
39. Lee N, Yun H, Lee C, et al. Engineered recombinant PON1-OPH fusion hybrids: potentially effective catalytic bioscavengers against organophosphorus nerve agent analogs. J Microbiol Biotechnol. 2021;31(1):144–153. doi:10.4014/jmb.2006.06044
40. Wilhelm CM, Snider TH, Babin MC, Jett DA, Platoff GE, Yeung DT. A comprehensive evaluation of the efficacy of leading oxime therapies in Guinea pigs exposed to organophosphorus chemical warfare agents or pesticides. Toxicol Appl Pharmacol. 2014;281(3):254–265. doi:10.1016/j.taap.2014.10.009
41. Israel LL, Braubach O, Galstyan A, et al. A combination of tri-leucine and angiopep-2 drives a polyanionic polymalic acid nanodrug platform across the blood-brain barrier. ACS Nano. 2019;13(2):1253–1271. doi:10.1021/acsnano.8b06437
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.