Back to Journals » International Journal of Nanomedicine » Volume 21
A Biocompatible Microdroplet Platform Incorporating MSC-Derived Exosomes for Bone Regeneration and Osteoporosis Treatment
Authors Sima J, Chen J, Liu X, Li D, Huang W, Wang Y, Ma Y, Zhang Y, Wang X
Received 23 April 2026
Accepted for publication 6 July 2026
Published 13 July 2026 Volume 2026:21 614120
DOI https://doi.org/10.2147/IJN.S614120
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Kamakhya Prakash Misra
Jincheng Sima,1,* Junrong Chen,1,* Xiliang Liu,2 Dinghong Li,1 Wenqing Huang,1 Yi Wang,1 Yaping Ma,1 Yi Zhang,1,2 Xin Wang1,3,4
1Department of Orthopaedic Surgery, Affiliated Hospital of Zunyi Medical University, Zunyi, Guizhou, People’s Republic of China; 2Department of Hygiene Toxicology, School of Public Health, Zunyi Medical University, Zunyi, Guizhou, People’s Republic of China; 3Zunyi Municipal Key Laboratory of Medicinal Biotechnology, Affiliated Hospital of Zunyi Medical University, Zunyi, Guizhou, People’s Republic of China; 4Institute of Medicinal Biotechnology, Affiliated Hospital of Zunyi Medical University, Zunyi, Guizhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yi Zhang, Department of Hygiene Toxicology, School of Public Health, Zunyi Medical University, Zunyi, Guizhou, People’s Republic of China, Tel +86 182 7566 2009, Fax +86 851 2860 9551, Email [email protected] Xin Wang, Department of Orthopaedic Surgery, Affiliated Hospital of Zunyi Medical University, Zunyi, Guizhou, People’s Republic of China, Tel +86 136 3928 8558, Fax +86 851 2860 8903, Email [email protected]
Introduction: Postmenopausal osteoporosis, characterized by progressive loss of bone mass and deterioration of skeletal microarchitecture, remains a major public health concern worldwide. Exosomes derived from mesenchymal stem cells (MSC-Exos) are nanosized extracellular vesicles enriched with bioactive cargo, have emerged as potent cell-free mediators of tissue regeneration and immunomodulation. However, their clinical translation is hindered by rapid clearance and insufficient retention at target sites. In this study, we developed a biocompatible microdroplet (MD)-based delivery platform incorporating MSC-derived exosomes (MD/Exos) and evaluated its therapeutic potential for osteoporosis treatment.
Methods: MSC-Exos were isolated from human bone marrow-derived mesenchymal stem cells (hBMSCs) and characterized by transmission electron microscopy and exosomal marker analysis. MD/Exos were fabricated and evaluated for cellular uptake and cytocompatibility. The effects of MD/Exos on hBMSC proliferation, osteogenic differentiation, and macrophage inflammatory responses were investigated in vitro. Therapeutic efficacy was further assessed in an ovariectomy-induced osteoporotic rat model through histological and cytokine analyses.
Results: MSC-Exos exhibited characteristic exosomal morphology and were efficiently internalized by both hBMSCs and RAW264.7 macrophages. The fabricated MD/Exos system demonstrated excellent cytocompatibility in vitro and favorable histocompatibility in vivo. In vitro, MD/Exos significantly enhanced osteogenic differentiation of hBMSCs and altered macrophage inflammatory cytokine profiles, suggesting a coordinated effect on osteogenesis and inflammatory regulation. In an ovariectomy-induced osteoporotic rat model, MD-Exo treatment effectively preserved trabecular bone microarchitecture, reduced marrow adiposity, and markedly inhibited osteoclast activity.
Conclusion: MD/Exos represent a promising exosome delivery strategy for osteoporosis treatment.
Keywords: bioactive microdroplets, MSC-Exos, biocompatibility, hBMSCs, osteogenesis, osteoporosis
Introduction
Postmenopausal osteoporosis is a skeletal disease characterized by increased bone mass loss and heightened fracture susceptibility, primarily attributed to diminished estrogen levels following menopause.1 With increasing life expectancy and the global aging population, osteoporosis represents a significant public health concern.2 Epidemiological data indicate that more than 14 million individuals in the United States and over 200 million people worldwide are affected by osteoporosis or osteoporotic fractures.3,4 Osteoporotic fractures, particularly of the hip and spine, are associated with chronic pain, long-term disability, diminished quality of life, thereby placing considerable burdens on both patients and healthcare systems.5 Currently, anti-resorptive agents remain the cornerstone of osteoporosis management.6 Among them, bisphosphonates are the most widely prescribed drugs and function by binding to the bone surface and inhibiting over-activation of osteoclasts and subsequent bone resorption, thereby reducing skeletal turnover.7 Denosumab is an FDA approved fully human monoclonal antibody for the treatment of osteoporosis.8,9 Denosumab functions by targeting and neutralizing receptor activator of nuclear factor kappa-B ligand (RANKL), the pivotal molecule involved in osteoclast differentiation, activation, and survival.9 Clinical evidence has demonstrated that denosumab significantly reduces fracture risk and improves bone mineral density with a favorable safety profile, offering an alternative strategy for postmenopausal osteoporosis management.9 However, despite their proven efficacy, long-term use of these agents has raised increasing concerns regarding their adverse effects, including atypical fractures, osteonecrosis of the jaw, and rebound-associated bone loss.10 Therefore, the development of novel and more effective therapeutic strategies for osteoporosis remains an urgent need.
Over recent decades, exosomes, nanosized extracellular vesicles typically ranging from 30 to 150 nm, have attracted increasing attention due to their fundamental role in mediating intercellular communication under both physiological and pathological conditions.11 Structurally, exosomes are enclosed by a lipid bilayer membrane that protects their cargo and enables stable transport in biological environments. Functionally, they serve as dynamic carriers of diverse bioactive molecules, including proteins, nucleic acids (such as DNA, mRNA, microRNA, and circular RNA), and lipids, thereby regulating multiple signaling pathways in recipient cells.12,13 Among the various cellular sources of exosomes, mesenchymal stem cells (MSCs) have attracted considerable attention due to their self-renewal capacity, immunomodulatory properties, and ability to differentiate into multiple cell types, including osteoblasts, chondrocytes, and adipocytes, etc.14 In recent years, increasing evidence has highlighted the therapeutic potential of exosomes derived from human bone marrow mesenchymal stem cells (hBMSCs) as a promising cell-free strategy for the treatment of a wide range of physiological and pathological disorders.15,16 Importantly, their nanoscale size facilitates efficient penetration across biological barriers, thereby enhancing their capacity for targeted delivery to specific tissues and cells.12 Moreover, MSC-derived exosomes (MSC-Exos) exhibit low immunogenicity and excellent biocompatibility, making them highly attractive candidates for safe and effective therapeutic applications in both preclinical and clinical conditions.12,17 Despite these advantages, exosome-based therapy still faces several critical translational barriers. One of the major limitations is their rapid clearance from systemic circulation, which significantly reduces bioavailability and therapeutic efficiency.18 In vivo tracking studies have demonstrated that systemically administered exosomes are rapidly cleared by organs of the mononuclear phagocyte system, resulting in short plasma half-life and poor tissue retention.19 Consequently, achieving therapeutic efficacy often requires repeated or high-dose administration, which may limit clinical feasibility. These challenges highlight the need for advanced delivery platforms designed to improve exosome protection and stability, thereby potentially enhancing their therapeutic efficacy at target sites.
Advanced nanocarrier systems have been widely developed to achieve spatiotemporally controlled drug delivery, particularly in cancer therapy applications.20 In recent years, microdroplets (MDs) have emerged as highly versatile and efficient carriers in the field of drug and gene delivery, owing to their unique physicochemical properties and favorable biocompatibility.21 Structurally, MDs share many similarities with nanodroplets and generally consist of a liquid perfluorocarbon core surrounded by a stabilizing shell composed of phospholipids, proteins, or other biocompatible surfactants.22 The incorporation of perfluorocarbon compounds within the core enhances droplet stability and responsiveness to acoustic stimulation, while the shell serves to prevent premature dissolution and facilitate the encapsulation of therapeutic cargo.22 The core-shell architecture of MDs confers enhanced physicochemical stability under physiological conditions, enabling efficient encapsulation and protection of therapeutic payloads from premature degradation or release prior to reaching the target site.23 Building upon our previous work demonstrating the therapeutic potential of a gene-silencing microdroplet platform in an ovariectomy-induced bone loss model,24 we sought to further enhance the regenerative capability of this delivery system by incorporating biologically active cargo. Although the previous platform successfully demonstrated the feasibility of MD-mediated therapeutic delivery, it lacked a regenerative component capable of directly promoting bone formation and tissue repair. Therefore, in the present study, we developed a novel microdroplet-exosome (MD/Exos) delivery platform by integrating hBMSCs-derived exosomes within microdroplet delivery system. We hypothesized that combining the biological activity of MSC-derived exosomes with the delivery capability of MDs would enhance osteogenic responses, improve bone remodeling, and attenuate bone loss in osteoporosis. Through the combined effects of exosome-mediated regulation of osteogenesis and bone remodeling, this platform represents a promising and versatile exosome delivery approach for osteoporosis treatment, offering a novel strategy to harness the therapeutic benefits of MSC-derived exosomes for the restoration of bone homeostasis and skeletal regeneration.
Materials and Methods
Cell Culture and Exosome Isolation
In this study, human bone marrow-derived mesenchymal stromal cells (hBMSCs; ATCC PCS-500-012, American Type Culture Collection, USA) were used for exosome isolation. The hBMSCs were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Life Technologies Pty Ltd., China) supplemented with 10% fetal bovine serum (FBS; Biological Industries, LTD, Beit Haemek, Israel), and 1% (v/v) penicillin/streptomycin (Solarbio, Beijing, China) under a 5% CO2 atmosphere at 37 °C. The RAW264.7 macrophages were obtained from the American Type Culture Collection (ATCC TIB-71, USA) and cultured in DMEM supplemented with 10% heat-inactivated FBS and 1% penicillin/streptomycin in a humidified incubator containing 5% CO2 at 37 °C. hBMSCs at passage 3 were grown in T75 flasks until they reached 90% confluence at 37 °C. Following three washes with phosphate-buffered saline (PBS), the cells were cultured with 10 mL DMEM supplemented with 10% exosome-depleted FBS at 37 °C for 24 h to collect exosome-containing conditioned medium (CM). To isolate exosomes, the CM was filtered through 0.22 μm filters to remove live cells and other large membranous structures. Subsequently, the filtered CM were centrifuged at 300×g at 4 °C for 10 min, followed by transfer to new tubes and centrifugation at 2,000×g at 4 °C for 20 min. Next, the CM was centrifuged in a 45Ti rotor (Beckman) at 10,000×g at 4 °C for 40 min. The supernatant was then spun at 100,000×g at 4 °C for 90 min to pellet the exosomes. The resulting exosome pellets were resuspended in PBS and stored at −80 °C.
Transmission Electron Microscopy (TEM)
Exosomes (5 μL) were deposited onto 200 mesh TEM copper grids coated with formvar carbon film and allowed to adhere for 5 minutes at room temperature. The grids were then stained with 1% uranyl acetate solution for a duration of 15–20 seconds. To remove any excess uranyl acetate, the grids were carefully rinsed with deionized water three times. Subsequently, samples were dried using Whatman filter paper. Images were captured using a JOEL transmission electron microscope (JEM-1400) at a voltage of 80 kV.
Western Blot
The expression of exosomal surface marker was evaluated by Western blot analysis. Briefly, the isolated exosome samples were lysed using RIPA buffer, and total proteins were separated using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Following electrophoresis, proteins were transferred onto PVDF membranes and blocked with Odyssey Blocking Buffer (LI-COR Biotechnology, Lincoln, NE, USA). Membranes were incubated with the following primary antibody: CD81 Antibody (B-11) (1:100, sc-166029, Santa Cruz). The following secondary antibody was used: IRDye 680RD goat anti-mouse IgG (H+L) (1:10,000; LI-COR Biotechnology, USA). The protein signal intensity was detected using Odyssey infrared Imaging System (LI-COR Biotechnology, USA).
Internalization of Exosomes Into Target Cells
Exosome labeling was performed using the PKH26 red fluorescent cell linker kit for general cell membrane labeling, following the protocol in our previous study (PKH26GL-1KT, Sigma, China).25 Briefly, purified exosomes were incubated with PKH26 dye and subsequently pelleted by ultracentrifugation using an ultracentrifuge (Optima, Beckman Coulter, USA) equipped with an SW41 Ti rotor at 100,000 × g for 70 min at 4 °C. The supernatant was carefully removed, and the exosome pellet was washed three times with PBS to remove unbound dye before final resuspension in PBS. As dye-only control, PBS without exosomes was processed in parallel following the same labeling and ultracentrifugation procedures to evaluate background fluorescence. For exosome uptake assays, PKH26-labeled exosomes were incubated with target cells at 37 °C for the indicated time periods. After incubation, cells were washed three times with PBS and examined using an IVIS imaging system (PerkinElmer/Xenogen, USA). For quantitative analysis, cells were washed twice with PBS, detached using 0.25% trypsin-EDTA, and analyzed by flow cytometry using a BD FACSCalibur system. Mean fluorescence intensity was calculated from the flow cytometry data to quantify exosome internalization. For confocal laser scanning microscopy, cells were washed three times with PBS and fixed with 4% paraformaldehyde. Following fixation, cells were permeabilized with 0.25% Triton X-100 and stained with Alexa Fluor 488-conjugated phalloidin for 1 h. Samples were then mounted using an antifade mounting medium containing DAPI (ProLong Gold Antifade Mountant, Thermo Fisher Scientific, China). Confocal images were acquired using a confocal laser scanning microscope (DM IRB, Leica Microsystems, Wetzlar, Germany). To evaluate whether MSC-Exos uptake induced cytotoxicity, hBMSCs and RAW264.7 macrophages were incubated with MSC-Exos for the indicated durations. After treatment, cells were stained with staining solution containing Calcein-AM, and images were acquired using confocal laser scanning microscopy (Leica, Wetzlar, Germany).
Fabrication and Characterization of MD/Exos
The MD-Exos was fabricated according to our previous study with slight modification. Briefly, exosomes (10 μg in terms of protein), 300 μL perfluorocarbon, and 4 mL of PBS, and 40 mg of bovine serum albumin (BSA, Sigma Aldrich, China) were used for MD/Exos fabrication. The resulting emulsion was ultracentrifuged (Beckman Coulter, Optima XPN-100, USA), and the MD/Exos were used for subsequent testing. To validate the successful internalization of exosomes into MDs, exosomes were fluorescently labeled using the PKH26 Red Fluorescent Cell Linker Kit, as described previously. A dye-only sample processed under identical conditions without exosomes was used as a negative control. The incorporation of PKH26-labeled exosomes into MDs (MD/Exos) was assessed by detecting fluorescence signals using an IVIS imaging system. To evaluate loading efficiency, CD81 expression was analyzed by Western blot in exosomes prior to loading and in residual exosomes collected from the supernatant after the loading process. The morphology of MD/Exos was further examined using confocal laser scanning microscopy and transmission electron microscopy (TEM). For TEM analysis, samples were fixed, air-dried, and visualized using a JEM-1400 JEOL TEM. To further validate the successful internalization of exosomes into MDs, hBMSCs were incubated with MD/Exos cells at 37 °C. Subsequently, the cells were washed three washes with PBS, followed by fixation using 4% paraformaldehyde. After rinsing, the samples were permeabilized with 0.25% Triton and stained with Alexa Fluor 488-labeled phalloidin for 1 h. Subsequently, the samples were mounted using an antifade mounting medium containing DAPI (P36935, Thermo Fisher Scientific, China). Images were acquired using a confocal laser scanning microscope (Leica DM IRB; Leica, Wetzlar, Germany). To validate exosomes internalization into target cells, the cellular uptake of MD/Exos into target cells was further evaluated using IVIS imaging system. To evaluate the release kinetics of MD-Exos, exosomes were collected from the supernatant of MD/Exos at indicated time points. The collected exosomes were then pelleted by centrifugation and incubated with macrophages to assess cellular uptake. Internalization was evaluated using IVIS imaging system, and fluorescence intensity was quantified to determine release kinetics and uptake efficiency. To further validate exosomal release, macrophages incubated with the released exosomes were fixed and imaged using confocal laser scanning microscopy (Leica DM IRB; Leica, Wetzlar, Germany).
Live and Dead Staining
Live/Dead staining assay was conducted using the Calcein/PI live/dead viability assay kit according to the manufacturer’s instructions (Beyotime, China). Briefly, the cells were stained with staining solution containing Calcein-AM and propidium iodide (PI) at room temperature. Calcein-AM marked viable cells with green fluorescence, while PI-stained nonviable cells red. After staining, cells were gently rinsed with PBS to remove excess dye. Confocal laser scanning microscopy (Leica, Wetzlar, Germany) was used to visualize stained cells at ×10 magnification.
Cell Morphology Analysis via Phalloidin/DAPI Staining
To assess cell morphology, phalloidin and DAPI staining were performed. Briefly, cells were fixed with 4% paraformaldehyde, permeabilized with 0.25% Triton X-100, and incubated with Alexa Fluor 594-labeled phalloidin for 1 hour. The nuclei were counterstained with antifade mounting medium with DAPI (Thermo Fisher Scientific, China). Imaging was performed using a confocal laser scanning microscope (Leica DM IRB, Wetzlar, Germany).
Internalization of MD/Exos Into Target Cells
The cellular uptake dynamics of MD/Exos into target cells was evaluated using confocal laser scanning microscopy. Briefly, cells were treated with FITC-labeled MD/Exos for 2 h, 6 h, 12 h, and 24 h. Following treatment, the cells were thoroughly rinsed with PBS to remove unbound MD/Exos. Subsequently, cells were fixed in 4% paraformaldehyde for 10 minutes at room temperature. To visualize the actin cytoskeleton, the fixed cells were stained with Alexa Fluor 594-labeled phalloidin followed by DAPI staining. The stained samples were subsequently examined using a confocal laser scanning microscope (Leica DM IRB; Leica, Wetzlar, Germany).
Scratch Wound Migration Assay
The migratory capacity of hBMSCs was evaluated using a scratch wound assay.26 After MD/Exos treatment, cells were stained with calcein-AM to visualize viable cells and imaged by fluorescence microscopy (Leica, Wetzlar, Germany).
Cell Protection Assay
To evaluate the cellular protective effect of MD/Exos, RAW264.7 macrophages were exposed to hydrogen peroxide (H2O2) at a final concentration of 400 µM. Cells were treated with or without MD/Exos, and cell viability was assessed using live/dead staining. The stained samples were subsequently examined using a confocal laser scanning microscope (Leica DM IRB; Leica, Wetzlar, Germany).
Intracellular Reactive Oxygen Species (ROS) Detection
Intracellular ROS levels were measured using the DCFDA/H2DCFDA Cellular ROS Assay Kit (ab113851, Abcam, USA). Briefly, following H2O2 exposure and MD/Exos treatment, cells were washed with PBS and incubated with DCFDA solution according to the manufacturer’s protocol. After staining, nuclei were counterstained with Hoechst 33258. The stained samples were subsequently examined using a confocal laser scanning microscope (Leica DM IRB; Leica, Wetzlar, Germany).
Assessment of Osteogenic Differentiation
To induce osteogenic differentiation, hBMSCs were cultured in osteogenic differentiation medium containing 10% FBS, 1% penicillin/streptomycin, 10 mM β-glycerophosphate (G9422, Sigma, China), 50 μM L-ascorbic acid 2-phosphate (49752, Sigma, China), and dexamethasone (100 nM, D4902, Sigma, China). ALP staining was conducted using the BCIP/NBT Alkaline Phosphatase Color Development Kit (Beyotime, Shanghai, China) following the manufacturer’s protocol. Imaging of stained cells were visualized using an inverted light microscope (Leica, Wetzlar, Germany). To assess cell viability during osteogenic differentiation, cells were subjected to live/dead staining as described above, and imaged via confocal laser scanning microscopy (Leica DM IRB; Leica, Wetzlar, Germany).
Immunofluorescence Staining of Osteogenic Markers
Immunofluorescence staining was performed to examine the expression of osteogenic marker: ALP. Briefly, cells were fixed with 4% paraformaldehyde, permeabilized with 0.25% Triton X-100, and blocked with 4% bovine serum albumin (BSA) for 1 hour at room temperature. Primary antibodies against ALP (1:100 dilution, Abcam) was used for overnight incubation at 4°C. After washing, cells were treated with a FITC-conjugated goat anti-rabbit IgG (H+L) secondary antibody. F-actin was visualized using Alexa Fluor 594-labeled phalloidin, and nuclei were counterstained with DAPI. Stained samples were mounted using antifade mounting medium with DAPI and visualized under a confocal laser scanning microscope (Leica DM IRB, Wetzlar, Germany).
Alizarin Red S Staining
Alizarin Red S staining was performed to assess mineralization. Briefly, cells were fixed with 4% paraformaldehyde, washed with deionized water, and stained with a 2% Alizarin Red S solution (pH 4.1) for 20 minutes at room temperature. Imaging was performed using an inverted light microscope (Leica, Wetzlar, Germany). To quantify mineral deposition, 10% cetylpyridinium chloride in sodium phosphate was used to dissolve the dye, and absorbance was measured spectrophotometrically at 570 nm.
Osteogenic-Related Gene Expression Analysis by Reverse Transcription-Quantitative PCR (RT-qPCR)
To analyze osteogenic gene expression, total RNA was extracted from hBMSCs using TRIzol reagent (Thermo Fisher Scientific, China) according to the manufacturer’s instructions. RNA concentration and purity were assessed using a NanoDrop spectrophotometer (NanoDrop Technologies). Reverse transcription was carried out with 500 ng of total RNA using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, China). Quantitative assessment of osteogenic-related gene expression was performed using using SYBR Green qPCR Master Mix on a real-time PCR system (QuantStudio, Thermo Fisher Scientific, USA). Relative gene expression levels were calculated using the comparative Ct (2−ΔΔCT) method.27
Human TGF Beta Array Analysis
Human TGF beta array analysis was performed following the protocols provided by the manufacturer (Ray Biotech, GA, USA). Briefly, cells were collected for array analysis on day 7. Protein concentration was determined using the Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, China). Membranes were placed in incubation tray wells and blocked with 2 mL of blocking buffer for 30 min at RT, followed by overnight incubation with equal amount of protein lysates at 4 °C. After washing, membranes were incubated overnight at 4 °C with the Biotinylated Antibody Cocktail, then treated with 2 mL of 1× HRP-Streptavidin solution for an additional overnight incubation. Protein spots were visualized using the Li‑Cor Odyssey M Imaging System and semi‑quantified with ImageJ software.
Macrophage Activation
Macrophage activation was conducted based on established protocols.28 Briefly, macrophages were seeded on tissue culture-treated coverslips and allowed to adhere overnight. The cells were then stimulated with Lipopolysaccharide (LPS, Escherichia coli O111:B4, Sigma, China), with or without MD/Exos treatment. Cells were washed with PBS, fixed in 4% paraformaldehyde, and subjected to fluorescence staining. F-actin was visualized using Alexa Fluor 594-labeled phalloidin, while nuclear staining was performed using DAPI. Samples were mounted with antifade mounting medium (Thermo Fisher Scientific, China). Images were acquired using a confocal laser scanning microscope (Leica DM IRB; Leica, Wetzlar, Germany). Cell spreading areas were semi-quantified using ImageJ software.
Cytokine Profiling Using Mouse Antibody Array
The cytokine profiles changes in the macrophages following MD/Exos treatment were examined using a Mouse Inflammation Array C1 (Ray Biotech, GA, USA), following the protocols provided by the manufacturer. Briefly, cells were harvested for cytokine antibody array analysis. Each array membrane was incubated with 2 mL of blocking buffer at room temperature for 30 minutes, followed by overnight incubation at 4°C with samples. After extensive washing with Wash Buffer I and II, membranes were incubated overnight at 4°C with a Biotinylated Antibody Cocktail. Subsequently, 2 mL of 1X HRP-Streptavidin solution was added to each well and incubated overnight at 4°C. After this incubation, the membranes underwent a final wash with wash buffer I and II to ensure thorough removal of any unbound streptavidin. Protein spots on the membranes were visualized using Li-Cor Odyssey M Imaging System. ImageJ software was utilized for semi-quantitative analysis of cytokine expression levels.
Animal Study
Female Sprague-Dawley (SD, 8 weeks old) rats underwent ovariectomy (OVX) or sham-operated (sham) were used for this study. All animal experiments were in accord with institutional animal use and care regulations approved by the Zunyi Medical University Committee (ZMU21-2203-009). For surgical procedures, the animals were anesthetized using 2% isoflurane with the eyes protected by applying the ophthalmic liquid gel. The anesthesia depth was continuously monitored throughout the procedure to minimize pain and distress. After post-OVX surgery recovery, the rats were divided into four groups: the first group consisted of sham-operated rats (sham) (n = 6); the second group comprised rats that underwent ovariectomy (OVX) (n = 6), the third group were OVX rats received MDs (n = 6), the fourth group were OVX rats received MD/Exos (n = 6). Intraperitoneal (I.P.) injection were performed twice a week (20 mg/kg in saline), on day 1 and day 4 throughout the whole experiment. The rats were euthanized at 10 weeks using carbon dioxide (CO2) inhalation in accordance with the recommendations of the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals. Following euthanasia, femoral samples were harvested for subsequent histological analysis.
Histological Assessment
To perform haematoxylin and eosin (H&E) staining, the bones were first carefully extracted and cleaned to remove any surrounding soft tissue. Following this, they were immersed in 10% neutral buffered formalin for fixation. Subsequent decalcification was conducted using a 10% ethylenediaminetetraacetic acid (EDTA, pH 7.4) solution. After the decalcification process, the samples were then embedded in paraffin wax and were cut into sections with a thickness of 5 µm using a microtome. These sections were then subjected to a sequential staining protocol using the Hematoxylin-Eosin (H&E) Staining Kit (Beyotime, Shanghai, China). Images were captured using an inverted light microscope (Leica, Wetzlar, Germany).
Tartrate-Resistant Acid Phosphatase (TRAP) Staining
Osteoclast activity was determined using tartrate-resistant acid phosphatase staining (TRAP staining) following protocols established in our previous study.24 Images were captured using an inverted light microscope (Leica, Wetzlar, Germany).
Immunohistochemical Staining
For immunohistochemistry analysis, the paraffin tissue sections were deparaffinized in 100% xylene followed by rehydration in a series of graded ethanol solutions. Antigen retrieval was performed using proteinase K (Thermo Fisher Scientific, China). After washing with PBS, the tissue sections were blocked with ReadyProbes 2.5% Normal Goat Serum (1X) (Thermo Fisher Scientific, China) and incubated overnight at 4°C with the following primary antibodies: Anti-Cathepsin K antibody (ab187647, Abcam, China), anti-MMP9 (ab76003, Abcam, China). Hematoxylin was used as counterstaining for cell nuclei. Images were captured using an inverted light microscope (Leica, Wetzlar, Germany).
Statistical Analysis
Data are presented as mean ± standard deviation (SD; n = 3). Statistical analyses were performed using GraphPad Prism version 7.02 (GraphPad Software, San Diego, CA, USA). For comparisons between two groups, an unpaired two-tailed Student’s t-test was used. For comparisons among three or more groups, one-way analysis of variance (ANOVA) followed by Tukey’s multiple-comparison test was performed. A p value < 0.05 was considered statistically significant.
Results
Purification and Characterization of MSC-Exos
As illustrated in Figure 1A, we first isolated exosomes from hBMSCs. Transmission electron microscopy (TEM) analysis confirmed the successful isolation of MSC-Exos, which exhibited the typical spherical or cup-shaped morphology characteristic of exosomes (Figure 1B). In our previous study, exosomes isolated using the same protocol were shown to express the classical exosomal marker CD63.25 In addition, nanoparticle tracking analysis was not newly performed as the size distribution of the isolated exosomes had been comprehensively characterized in our previous work.29 To characterize the exosomes obtained in the present study, Western blot analysis was performed to detect the exosomal marker CD81. As shown in Figure 1C, MSC-Exos exhibited strong CD81 expression, further confirming the successful isolation and characterization of exosomes from hBMSCs. Fluorescently labeled MSC-Exos were incubated with hBMSCs (Figure 1D and E) and RAW264.7 macrophages (Figure 1F and G). IVIS analysis revealed robust uptake of MSC-Exos by both cell types, with significantly higher fluorescence intensity compared to mock-treated controls. The uptake of MSC-Exos was further validated by flow cytometric analysis. A marked increase in fluorescence signal was detected in RAW264.7 cells following MSC-Exos treatment (Figure 1H and I), indicating effective exosome internalization. Similarly, hBMSCs exhibited significant uptake of MSC-Exos, as demonstrated by flow cytometric analysis and quantitative fluorescence measurements (Figure 1J and K).
 |
Figure 1 Isolation and characterization of MSC-derived exosomes (MSC-Exos). (A) Schematic diagram for the MSC-Exos isolation process. (B) Transmission electron microscopy (TEM) images showing the characteristic spherical or cup-shaped morphology of isolated MSC-Exos. (C) Western blot analysis confirmed the expression of the exosomal marker CD81 in the isolated exosomes. (D) Uptake of labeled MSC-Exos by hBMSCs visualized using the IVIS imaging system. (E) Quantification of fluorescence intensity. Significant difference: ****p=0.000016. (F) Uptake of labeled MSC-Exos by RAW264.7 macrophages visualized using the IVIS imaging system. (G) Quantification of fluorescence intensity. Significant difference: ****p=0.000003. (H) Flow cytometry analysis confirming MSC-Exos internalization in RAW264.7 cells. (I) Quantification of fluorescence signal in RAW264.7 cells treated with or without MSC-Exos. Significant difference: ****p=0.000003. (J) Internalization of MSC-Exos into hBMSCs. Cellular internalization of MSC-Exos by flow cytometric analysis. (K) Quantification of fluorescence signal in hBMSCs treated with or without MSC-Exos. Significant difference: ***p =0.0003. (L) Schematic diagram illustrating the in vitro internalization assay using hBMSCs. (M) Confocal laser scanning microscopy images of hBMSCs following mock treatment. Nuclei were counterstained with DAPI (blue), and F-actin filaments were stained with phalloidin (green). (N) Confocal microscopy images showing the internalization of PKH26-labeled MSC-Exos into hBMSCs at different time points. (O) Schematic diagram illustrating the in vitro internalization assay using RAW264.7 macrophages. (P) Confocal laser scanning microscopy images of RAW264.7 macrophages following mock treatment. (Q) Confocal laser scanning microscopy analysis demonstrated the internalization of PKH26-labeled MSC-Exos into RAW264.7 macrophages at different time points. Scale bar = 50 µm (low mag), Scale bar = 20 µm (high mag). |
To further investigate the uptake kinetics and internalization dynamics of MSC-Exos by target cells, we performed an in vitro uptake assay using two cell types: hBMSCs and RAW264.7 macrophages. Figure 1L shows the in vitro internalization assay in hBMSCs. As a control, hBMSCs were subjected to mock treatment, and their morphology was examined using confocal laser scanning microscopy. As shown in Figure 1M, mock-treated hBMSCs displayed normal cytoskeletal organization, with well-spread morphology and intact F-actin filaments. In addition, no red fluorescence was detected, confirming the absence of exosomal uptake under control conditions. To assess exosome internalization, hBMSCs were incubated with PKH26-labeled MSC-Exos, and uptake was examined at different time points using confocal microscopy (Figure 1N). A gradual increase in red fluorescence within the cytoplasm of hBMSCs was observed over time, indicating efficient and time-dependent uptake of exosomes. Notably, the MSC-Exos was localized predominantly in the perinuclear region. In addition, a live cell staining assay was conducted to assess the viability of hBMSCs following exposure to MSC-Exos. As shown in Supplementary Figure 1, the cells exhibited strong green fluorescence due to calcein-AM staining, indicative of viable cells with intact membranes. These results demonstrate that MSC-Exos treatment does not adversely affect hBMSCs viability. A similar approach was employed to assess MSC-Exos uptake in RAW264.7 macrophages (Figure 1O). Confocal microscopy of mock-treated RAW264.7 cells (Figure 1P) revealed typical macrophage morphology with intact nuclei, and no evidence of PKH26 signal, confirming the absence of exosomes in control conditions. Upon treatment with PKH26-labeled MSC-Exos, RAW264.7 macrophages exhibited robust internalization of exosomes, as evidenced by the progressive increase in red fluorescence within the cells at indicated time points (Figure 1Q). In addition, live staining results further confirmed strong green fluorescence in RAW264.7 macrophages following MSC-Exos uptake (Supplementary Figure 2). These findings demonstrate that MSC-Exos are efficiently internalized by both hBMSCs and macrophages. Collectively, these results confirm the successful isolation of MSC-Exos.
Synthesis and Characterization of MD/Exos
To enable visualization of exosomes during microdroplet fabrication and cellular uptake, MSC-Exos were fluorescently labeled with PKH26 prior to incorporation into microdroplets. A schematic diagram of the PKH26 labeling procedure and subsequent MD/Exos fabrication process is shown in Figure 2A. Fluorescence imaging demonstrated strong red fluorescence signals from PKH26-labeled exosomes, whereas the dye-only control without exosomes exhibited negligible fluorescence (Figure 2B), confirming successful and specific labeling of exosomes. Following MD/Exos fabrication, robust fluorescence signals were retained within the MD/Exos, indicating efficient internalization of exosomes into the MD structure (Figure 2B). The loading efficiency was then examined by Western blot analysis of CD81 expression. As shown in Figure 2C, CD81 was strongly detected in exosomes prior to loading, whereas a markedly reduced signal was observed in the residual exosomes remaining in the supernatant after loading. Quantitative analysis further confirmed a high loading efficiency (Figure 2C). In addition, representative confocal laser scanning microscopy (CLSM) images revealed distinct PKH26-positive signals localized within the MDs (Figure 2D). Transmission electron microscopy (TEM) further confirmed the spherical morphology of MD/Exos (Figure 2E). Previous study has shown that microbubble stability remains largely unaffected following exposure to temperatures of up to 50°C.23 To further evaluate the behavior of the MD/Exos system under physiological conditions, we examined its expansion characteristics using confocal microscopy. The results demonstrated a progressive increase in the size of MD/Exos over time while maintaining their spherical morphology, indicating gradual expansion and possible release of exosomes over the incubation period (Figure 2F). The ability of MD/Exos to deliver exosomes into target cells was next evaluated. CLSM images demonstrated pronounced intracellular red fluorescence in hBMSCs treated with MD/Exos, indicating effective cellular uptake of MSC-Exos delivered via the MD platform (Figure 2G). To further confirm the cellular uptake of MD/Exos, the internalization was examined in macrophages using the IVIS imaging system (Figure 2H). Strong fluorescence signals were detected in macrophages treated with MD/Exos, and quantitative analysis revealed a significantly higher fluorescence intensity compared with control groups (Figure 2J). Similarly, IVIS imaging of hBMSCs treated with MD/Exos showed pronounced fluorescence signals (Figure 2I and K). Finally, the release kinetics of MSC-Exos from MD/Exos were evaluated. Exosomes collected from the supernatant of MD/Exos were incubated with macrophages, and internalization was assessed using IVIS imaging. The results demonstrated sustained release and effective uptake of released exosomes by macrophages over time (Figure 2L and M). Confocal laser scanning microscopy further validated the internalization of released exosomes within macrophages (Figure 2N).
 |
Figure 2 Synthesis and characterization of MD/Exos. (A) Schematic diagram illustrating the PKH26-based fluorescent labeling of exosomes and the subsequent fabrication of MD/Exos. (B) Representative fluorescence images acquired using the IVIS imaging system showing PKH26-labeled exosomes. A dye-only sample processed under identical conditions in the absence of exosomes was included as a negative control. PKH26-labeled exosomes were further incorporated into microdroplets to generate MD/Exos (red), and IVIS imaging confirmed the successful internalization of MSC-Exos within MDs. (C) Western blot analysis of CD81 expression in exosomes before loading and in residual exosomes remaining in the supernatant after loading; loading efficiency is shown on the right. (D) Representative confocal laser scanning microscopy images demonstrating the morphology of MD/Exos. (E) TEM image showing the spherical morphology of MD/Exos. Scale bar = 500 nm. (F) Representative confocal laser scanning microscopy images demonstrating the expansion of MD/Exos. Scale bar = 500 µm. (G) Confocal microscopy images showing the cellular uptake of MSC-Exos delivered via MD/Exos (red) in hBMSCs. (H) Evaluation of MD/Exos (red) internalization in macrophages using the IVIS imaging system. (I) Assessment of MD/Exos internalization by hBMSCs using the IVIS imaging system. (J) Quantitative analysis of fluorescence intensity of MD/Exos uptake in macrophages. Significant difference: ****p=0.000033. (K) Quantification of fluorescence intensity in hBMSCs. Significant difference: ****p=0.000003. (L) Release kinetics of MSC-Exos from MD/Exos. Exosomes collected from the supernatant of MD/Exos were incubated with macrophages, and internalized exosomes were evaluated using IVIS imaging system. (M) Quantitative analysis of fluorescence intensity. (N) Representative confocal laser scanning microscopy images showing macrophage internalization of released exosomes. |
MD-Exo Shows Good Biocompatibility in vitro
The biocompatibility of MD-Exo was assessed by evaluating its effects on cell viability and morphology in hBMSCs and RAW264.7 macrophages (Figure 3A). Live/dead staining was first performed to determine the viability of hBMSCs following MD-Exo treatment. As shown in Figure 3B, both the MD and MD/Exos-treated groups exhibited strong green fluorescence with minimal red staining, indicating the absence of detectable cytotoxicity. To further investigate whether MD-Exo affected cellular morphology, hBMSCs were subjected to phalloidin and DAPI staining. As illustrated in Figure 3C, no significant morphological alterations were observed across the experimental groups, suggesting that MD-Exo does not negatively impact hBMSCs growth. We next examined the biocompatibility of MD-Exo in RAW264.7 macrophages (Figure 3D). Live/dead staining demonstrated comparable cell viability between the control and MD/Exos-treated groups, with no evidence of increased cell death following treatment (Figure 3E). In addition, phalloidin and DAPI staining revealed no obvious morphological alterations or changes in response to MD-Exo exposure (Figure 3F). Collectively, these results indicate that MD-Exo does not induce cytotoxic effects or morphological alterations on RAW264.7 macrophages or hBMSCs, demonstrating its excellent biocompatibility. In addition, the scratch wound assay revealed a significantly enhanced wound closure in the MD-Exo-treated group compared to untreated controls, indicating elevated migratory activity of hBMSCs (Supplementary Figure 3). Furthermore, live/dead staining results revealed a higher proportion of viable cells in the Exos- and MD/Exos-treated group compared to untreated controls, indicating their protective effect against H2O2-induced cell death (Supplementary Figure 4). Intracellular reactive oxygen species (ROS) levels were assessed by DCFDA fluorescence staining in hBMSCs (Supplementary Figure 5) and RAW264.7 macrophages (Supplementary Figure 6) following H2O2 exposure with or without MD/Exos treatment. H2O2 stimulation markedly increased ROS fluorescence intensity, indicating pronounced oxidative stress. A similarly elevated ROS signal was observed in the MDs-treated group, suggesting that MDs alone did not alleviate oxidative damage. In contrast, treatment with MD/Exos significantly reduced intracellular ROS levels, as evidenced by the markedly decreased DCFDA fluorescence compared with both the control and MDs-treated groups.
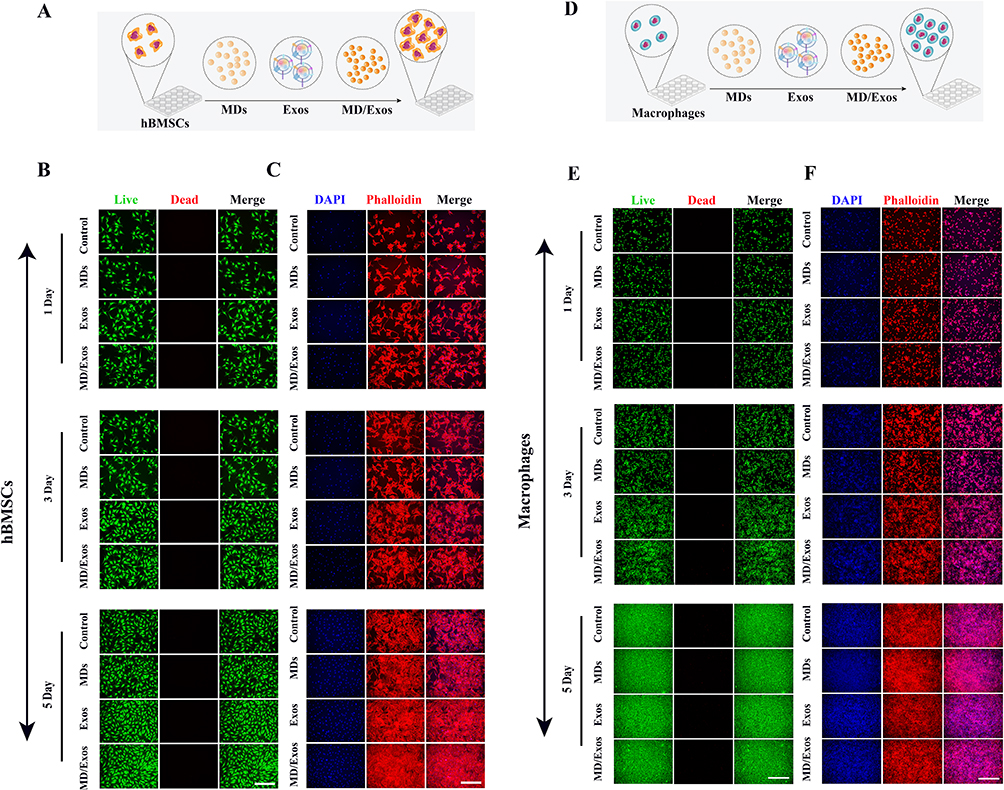 |
Figure 3 In vitro biocompatibility evaluation of MD-Exo in hBMSCs and RAW264.7 macrophages. (A) Schematic diagram for the in vitro study using hBMSCs. (B) Representative confocal laser scanning microscopy images of hBMSCs treated with or without MD-Exo following live/dead staining. Viable cells are stained green (calcein-AM), while dead cells are stained red (PI). Scale bar = 500 μm. (C) Representative confocal laser scanning microscopy images of hBMSCs treated with or without MD-Exo following DAPI/phalloidin staining. Cell nuclei and cytoskeleton are shown in blue and red, respectively. Scale bar = 500 μm. (D) Schematic diagram for the in vitro study using RAW264.7 macrophages. (E) Representative confocal laser scanning microscopy images of RAW264.7 macrophages treated with or without MD-Exo following live/dead staining. Viable cells are stained green (calcein-AM), while dead cells are stained red (PI). Scale bar = 500 μm. (F) Representative confocal laser scanning microscopy images of RAW264.7 macrophages treated with or without MD-Exo following DAPI/phalloidin staining. Scale bar = 500 μm. |
MD-Exo are Internalized by hBMSCs and RAW264.7 Macrophages in a Time-Dependent Manner
To investigate the internalization efficiency of MD-Exo in vitro, we also performed cellular uptake assays using hBMSCs and murine RAW264.7 macrophages. We first assessed MD-Exo internalization in hBMSCs (Figure 4A). As a negative control, cells were treated with vehicle only, and no green punctate fluorescence was observed under confocal microscopy, indicating the absence of MD-Exo internalization (Figure 4B). In contrast, when hBMSCs were incubated with FITC-labeled MD-Exo and imaged at various time points, we observed a time-dependent increase in cytoplasmic green fluorescence (Figure 4C), indicating efficient internalization of MD-Exo. These findings support the capacity of hBMSCs to internalize MD-Exo effectively. Next, we assessed the internalization of MD-Exo in RAW264.7 macrophages (Figure 4D). Control cells treated with vehicle alone retained their characteristic morphology, with well-defined nuclear staining and intact cytoskeletal structures, as shown in Figure 4E. Upon treatment with FITC-labeled MD-Exo, however, we observed a clear and progressive increase in intracellular green fluorescence over time (Figure 4F). The distribution pattern of MD-Exo was similar to that observed in hBMSCs. Collectively, these results demonstrate that MD-Exo are efficiently taken up by both hBMSCs and RAW264.7 macrophages in vitro.
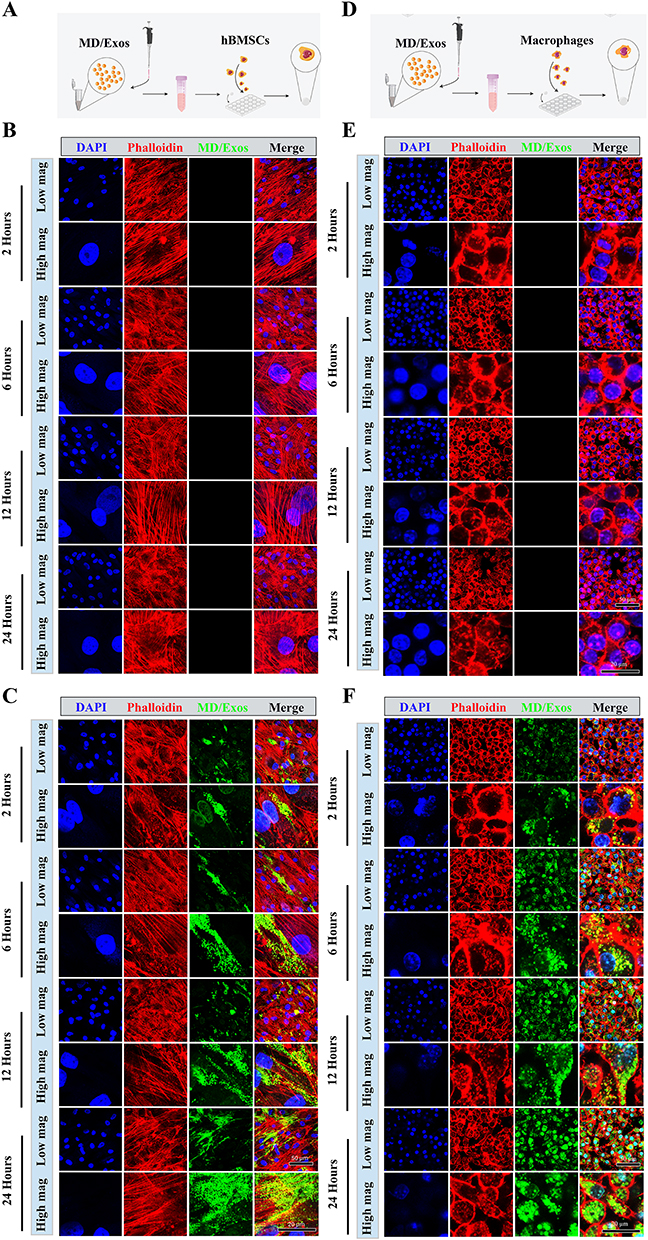 |
Figure 4 Efficient internalization of MD-Exo by hBMSCs and RAW264.7 macrophages in vitro. (A) Schematic diagram illustrating the experimental design for assessing MD-Exo internalization in hBMSCs. (B) Confocal laser scanning microscopy images of hBMSCs under mock treatment conditions. Nuclei were counterstained with DAPI (blue), and F-actin filaments were stained with phalloidin (red). (C) Confocal microscopy images showing the internalization of MD-Exo into hBMSCs. (D) Schematic diagram illustrating the internalization assay conducted in RAW264.7 macrophages. (E) Confocal microscopy images of RAW264.7 macrophages following mock treatment. (F) Confocal analysis revealing time-dependent uptake of MD-Exo by RAW264.7 macrophages. |
MD-Exo Treatment Promotes Osteogenic Differentiation and Mineralization in hBMSCs
We next investigate the impact of MD-Exo on the osteogenic differentiation of hBMSCs (Figure 5A). First, we evaluated the long-term viability of hBMSCs during osteogenic differentiation using live/dead staining. The live/dead assay at days 7 and 14 indicated that MD-Exo treatment did not compromise hBMSC viability or adherence compared to untreated controls (Supplementary Figure 7). Immunofluorescence staining for alkaline phosphatase (ALP), an early marker of osteogenic differentiation, revealed a marked increase in ALP expression in hBMSCs treated with MD-Exo compared to untreated controls (Figure 5B). Quantitative analysis of fluorescence intensity confirmed a significant increase in ALP levels in the MD-Exo group (Figure 5C). Next, we examined ALP expression using ALP staining in hBMSCs treated with or without MD-Exo. As shown in Figure 5D–G, the MD-Exo treatment revealed heightened ALP expression compared to the non-treated cells. This finding strongly implies the potentiated osteogenic differentiation potential following MD-Exo treatment. To assess late-stage osteogenesis and mineral deposition, Alizarin Red S staining was performed. As shown in Figure 5H, MD-Exo-treated hBMSCs showed enhanced mineralized matrix formation compared to untreated controls. Quantitative analysis of Alizarin Red S staining confirmed a marked increase in mineral deposition in hBMSCs following MD-Exo treatment (Figure 5I). By day 28, the difference in mineral deposition became even more pronounced, as evidenced by the robust Alizarin Red S staining in the MD-Exo group (Figure 5J). Quantification of Alizarin Red S staining further confirmed a sustained and significant enhancement in mineralization (Figure 5K). Finally, we validated the osteogenic phenotype of hBMSCs by examining the expression of osteogenesis-related marker genes. Real-time PCR analysis revealed significantly upregulated expression of key osteogenic markers following treatment with MD-Exo, including ALP (p= 0.0074), collagen type I (COLI) (p= 0.000008), osteocalcin (OCN) (p=0.000031), and osteopontin (OPN) (p=0.000039) (Figure 5L). These findings collectively indicate that MD-Exo enhances osteogenic differentiation and promotes mineralization in hBMSCs.
 |
Figure 5 MD-Exo treatment promotes osteogenic differentiation and mineralization in hBMSCs. (A) Schematic diagram for the in vitro study. (B) Representative immunofluorescence staining images showing alkaline phosphatase (ALP) expression in hBMSCs. Scale bar = 100 µm. (C) Quantification of ALP immunofluorescence intensity. Significant difference: ****p < 0.0001. (D) ALP staining images at day 7 showing elevated ALP activity in MD-Exo-treated cells. Scale bar = 500 µm. (E) Quantification of ALP staining areas, indicating a significant increase in ALP activity upon MD-Exo treatment. Significant difference: ****p < 0.0001. (F) ALP staining at day 14 further illustrates increased ALP activity in the MD-Exo group. (G) Quantification of ALP staining areas. Significant difference: ****p < 0.0001. (H) Alizarin Red S staining images demonstrating increased mineral deposition in MD-Exo-treated hBMSCs. Scale bar = 500 µm. (I) Quantitative analysis of Alizarin Red S staining reveals significantly higher mineralization in the MD-Exo group. Significant difference: ****p < 0.0001. (J) Representative Alizarin Red S staining images at day 28 further indicate enhanced mineral deposition. (K) Quantification of alizarin red S staining. Significant difference: ****p < 0.0001. (L) The expression of osteogenesis-related gene was evaluated using real-time PCR. GAPDH was used as a housekeeping gene. Significant difference: **p < 0.01, ***p < 0.001, ****p < 0.0001. (M) Representative membrane images from the Human TGF-β cytokine antibody array showing differential expression patterns between MDs-only and MD-Exos groups. Cell lysates were collected and analyzed using the Human TGF-β Array C2 platform. (N) Quantitative densitometric analysis of cytokine array. Significant difference: *p < 0.05, **p < 0.01. |
To elucidate the potential molecular mechanisms responsible for the enhanced osteogenic differentiation observed following MD/Exos treatment, cytokine profiling was performed using the Human TGF-β Array C2 platform under irradiated conditions (Figure 5M). Densitometric analysis identified significant upregulation of multiple osteogenic and TGF-β/BMP pathway-associated factors, including Activin A (p=0.1733), beta-IG-H3 (p= 0.0162), BMP4 (p=0.0011), BMP6 (p=0.0082), BMP9 (p=0.0238), BMP15 (p=0.0116), BMPRIA (p=0.0104), GDF15 (p=0.0026), SMAD4 (p=0.0133), TGFb1 (p=0.0017), and TGFb2 (p=0.0074) (Figure 5N). Collectively, these results indicate that MD/Exos may enhance osteogenic differentiation through activation of the TGF-β/BMP signaling network, thereby creating a pro-osteogenic microenvironment that supports osteoblast lineage commitment.
MD/Exos Modulate Macrophage Inflammatory Cytokine Responses in vitro
We next investigated whether MD/Exos could modulate macrophage inflammatory responses (Figure 6A). Morphological assessment by confocal laser scanning microscopy was conducted at day 3 following treatment. The results showed significant changes in cell spreading, with the formation of multinucleated giant cells observed by day 3 in both the control and MD-treated groups. In contrast, MD-Exo treatment resulted in a marked reduction in the formation of multinucleated giant cells, indicating its inhibitory effect on multinucleated giant cells formation (Figure 6B). Quantitative analysis of macrophage spreading area revealed a significant difference between MD/Exos-treated and untreated groups (Supplementary Figure 8). To comprehensively evaluate alterations in cytokine expression patterns following MD-Exo treatment, we further performed a cytokine array analysis. This approach allows for the simultaneous assessment of multiple cytokine expression, providing a comprehensive overview of cytokine profiles in response to MD-Exo treatment. (Figure 6C and D). The results indicated a significant downregulation of GCSF (p=0.0113), IL-4 (p=0.0107), IL-6 (p=0.0304), IL-12 p70 (p=0.0011), LIX (p=0.0130), MCP-1 (p=0.0036), M-CSF (p=0.0017), MIP-1 alpha (p=0.0050), MIP-1 gamma (p=0.0078), RANTES (p=0.0112), and TNF RI (p=0.0024) following MD-Exo treatment (Figure 6E). Collectively, these findings indicate that MD/Exos suppress excessive macrophage inflammatory activation and reshape cytokine secretion profiles.
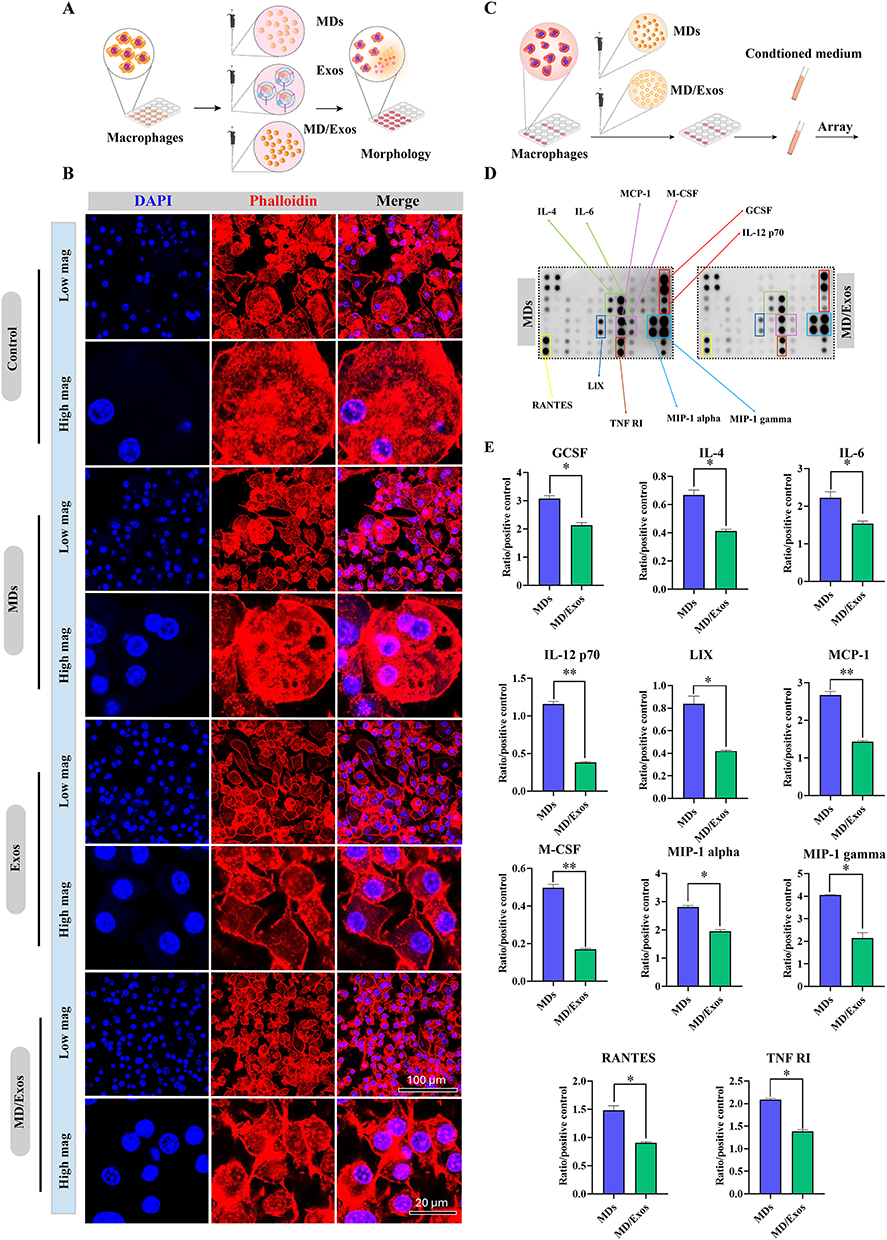 |
Figure 6 Regulation of macrophage inflammatory responses by MD/Exos in vitro. (A) Schematic illustration depicting the effects of MD/Exos on macrophage. (B) Confocal laser scanning microscopy images showing macrophage morphology at day 3 post-treatment. Actin filaments were labeled using phalloidin (red), and the cell nuclei were stained with DAPI (blue). (C) Schematic diagram for the in vitro cytokine antibody array study. (D) Cytokine antibody array analysis of macrophages treated with or without MD-Exo. (E) Quantification of cytokine array. The mean integrated pixel density of the pair of duplicate spots were quantified using imageJ software after subtraction of the background signal and normalized by the positive control. Significant difference: *p < 0.05, **p < 0.01. |
The Effect of MD-Exo in Preventing Osteoporosis in OVX Rats
The therapeutic efficacy of MD-Exo in preventing ovariectomy (OVX)-induced osteoporotic bone loss was evaluated using an established OVX rat model (Figure 7A). Histological examination of major organs by H&E staining revealed no obvious pathological abnormalities in the MD-Exo-treated group, indicating favorable in vivo biosafety and good biocompatibility of the treatment (Figure 7B). We next examined the structural alterations in animals with or without MD-Exo treatment. As shown in Figure 7C, OVX animals exhibited severe deterioration of trabecular bone microarchitecture, characterized by increased trabecular separation and a marked accumulation of adipocytes within the bone marrow cavity. Similar pathological features were observed in the OVX + MDs group, suggesting that MDs alone did not effectively prevent OVX-induced bone loss. In contrast, rats treated with MD-Exo exhibited marked preservation of trabecular structure, with reduced trabecular separation and fewer marrow adipocytes (Figure 7G), indicating significant attenuation of osteoporotic bone loss. To further investigate the effects of MD-Exo on osteoclast activity, TRAP staining was performed. OVX rats showed a substantial increase in TRAP-positive osteoclasts, whereas MD-Exo treatment markedly reduced the number of TRAP-positive cells (Figure 7D and H). Furthermore, we examined the expression levels of osteoclast-related markers cathepsin K (CTSK) and matrix metalloproteinase 9 (MMP9) via immunohistochemical staining. Our results indicated a substantial reduction in the expression of CTSK (Figure 7E and I) and MMP9 (Figure 7F and J) in response to MD-EXO treatment. Taken together, these results indicated the promising role of MD-Exo as a potential therapeutic intervention for osteoporosis, while exhibiting excellent in vivo safety.
 |
Figure 7 Therapeutic effects of MD-Exo on ovariectomy-induced osteoporotic bone loss in rats. (A) Schematic diagram illustrating the experimental design and treatment protocol for the in vivo study. (B) Representative H&E–stained sections of major organs, showing no obvious histopathological abnormalities, indicating good biosafety of MD-Exo treatment. Dotted boxes indicate the regions selected for higher-magnification imaging. Scale bar: 250 µm. (C) Representative H&E-stained femoral sections. Compared with the sham group, both the OVX and OVX + MDs groups exhibited pronounced trabecular bone deterioration, characterized by increased trabecular separation and a higher number of adipocytes in the bone marrow cavity. In contrast, MD-Exo treatment markedly preserved trabecular architecture and effectively attenuated OVX-induced bone loss. (D) Representative TRAP staining images. The MD-Exo-treated group showed a substantial reduction in the number of TRAP-positive osteoclasts compared with OVX controls, indicating suppressed osteoclast activity. (E) Immunohistochemical analysis of cathepsin K (CTSK) expression. (F) Immunohistochemical analysis of matrix metalloproteinase 9 (MMP9) expression. For panels C-F, scale bars = 500 µm (left panels), 250 µm (middle panels), and 100 µm (right panels). (G) Quantitative analysis of adipocyte area/total area. Significant difference: ****p < 0.0001. (H) Quantitative analysis of TRAP-positive osteoclasts. Significant difference: ****p=0.00001. (I) Quantification of CTSK-positive cells. Significant difference: ****p=0.00002. (J) Quantification of MMP9-positive cells. Significant difference: ***p=0.0007. |
Discussion
Osteoporosis remains a major public health concern, primarily driven by an imbalance between bone resorption and bone formation. Clinically, osteoporotic patients will experience an increased bone fragility and a heightened risk of fractures, particularly in the hip, spine, and wrist-posing a significant burden on both patients and healthcare systems worldwide. With the global population aging rapidly, the prevalence of osteoporosis is rising, thus highlighting an urgent need for more effective and targeted therapeutic interventions.30 In recent years, mesenchymal stem cell-derived exosomes have emerged as a promising acellular therapeutic strategy due to their intrinsic regenerative and immunomodulatory properties. MSC-Exos can modulate the bone microenvironment by targeting multiple stages of bone healing, thereby offering a natural yet potent approach to restoring skeletal homeostasis. However, the clinical translation of exosome-based therapies has been hindered by several intrinsic limitations, including labor-intensive and low-yield isolation protocols, poor structural stability, rapid clearance, and the lack of efficient tissue-specific targeting mechanisms.31
To overcome these limitations, the field of tissue regeneration has increasingly shifted from native exosome therapies toward engineered exosome platforms that combine biological, molecular, and biomaterial engineering approaches to enhance their therapeutic efficacy and functionality. Through genetic modification, cargo engineering, biomaterial integration, and microenvironmental preconditioning, engineered exosomes have demonstrated superior regenerative and immunomodulatory capabilities in preclinical bone regenerating models.32 For example, Li et al engineered bone-targeting exosomes enriched with METTL3 mRNA and demonstrated that these exosomes enhanced osteogenic differentiation while suppressing adipogenic differentiation of BMSCs through METTL3-mediated m6A modification of FOXO1. In ovariectomized mouse models of postmenopausal osteoporosis, the engineered exosomes effectively reduced bone loss and restored skeletal homeostasis.33 Furthermore, engineered exosome-membrane hybrid nanoparticles have been developed to target vascular dysfunction in osteoporosis.34 Tang et al also developed transcriptomics-guided exosome-encapsulated bifunctional nanosheets, which target PI3K/AKT pathway, enabling simultaneous regulation of inflammation, osteoclast activity, and osteogenesis to improve bone regeneration in osteoporosis.35 Similarly, Long et al demonstrated that Scutellaria baicalensis Georgi-preconditioned MSC-derived exosomes enhanced osteogenesis and alleviated osteoporosis through Semaphorin 3A (Sema3A)-mediated signaling,36 whereas Si et al engineered curcumin-loaded exosome-containing hydrogel microspheres that promoted bone regeneration via coordinated immunomodulation, osteogenesis, angiogenesis, and DNA damage repair.37 Xiang et al developed a bone-targeting exosome-loaded hydrogel that combined ROS-scavenging nanoparticles with 3D-cultured BMSC-derived exosomes, thereby improving exosome retention, relieving oxidative stress, activating Wnt/β-catenin signaling, and enhancing bone regeneration.38 In addition, our group previously developed a bioactive hybrid scaffold incorporating NFATc1-silencing siRNA-loaded ultrasound-responsive microdroplets and MSC-derived exosomes. This multifunctional platform simultaneously inhibited osteoclastogenesis and promoted osteogenic differentiation, thereby enhancing bone regeneration through the coordinated regulation of bone resorption and bone formation.39 Building upon these advances, we engineered a novel microdroplet-exosome (MD/Exos) platform that integrates MSC-derived exosomes with a dynamic microdroplet delivery system and investigated its therapeutic efficacy in both in vitro and in vivo models.
Previous studies indicated that MSC-Exos play a pivotal role in promoting osteogenesis in vitro and enhancing bone regeneration in vivo.40 Unlike MSCs, exosomes exhibit low immunogenicity and higher biocompatibility, making them attractive candidates for cell-free therapeutic strategies. In vitro studies have consistently demonstrated that treatment of naïve mesenchymal stem cells or pre-osteoblasts with MSC-derived exosomes enhances the expression of early and late osteogenic markers, including alkaline phosphatase (ALP), osteocalcin (OCN), runt-related transcription factor 2 (RUNX2), and collagen type I (COL1A1), alongside increased matrix mineralization.41–43 Molecularly, MSC-derived exosomes have been shown to initiate and sustain osteogenic differentiation by modulating key signaling pathways such as Wnt/β-catenin, PI3K/Akt, MAPK, TGF-β, and BMP/Smad.44–47 In addition, in vivo animal models and preclinical studies have revealed that exosome administration significantly accelerates bone defect healing, improves callus formation, and supports the regeneration of structurally and functionally mature bone tissue.48,49 Consistent with these findings, we also demonstrated that MSC-derived exosomes effectively promote osteogenic differentiation, as evidenced by increased expression of key osteogenic markers, enhanced alkaline phosphatase activity, and robust matrix mineralization in treated hBMSCs. Our results further support the notion that MSC-Exos act as potent mediators of bone regeneration by delivering pro-osteogenic signals that activate critical intracellular pathways involved in skeletal tissue development. In addition, our in vitro evaluation provides compelling evidence that MD/Exos significantly enhance the osteogenic potential of hBMSCs without compromising cell viability or structural integrity, highlighting their robust bioactivity. Notably, the pro-osteogenic effects observed with MD/Exos were comparable to those elicited by conventional MSC-derived exosomes, suggesting that MD-Exos possess a similarly potent capacity to stimulate osteogenic differentiation and bone matrix formation in hBMSCs. This finding highlights the therapeutic promise of MD/Exos as a viable and effective alternative to standard MSC-Exos in regenerative applications.
To gain insight into the molecular mechanisms underlying the enhanced osteogenic differentiation induced by MD/Exos, a Human TGF-β Array analysis was performed. The array revealed increased expression of several factors associated with osteogenesis and the TGF-β/BMP signaling network. Previous studies have identified BMP6 and BMP9 as key osteogenic regulators capable of promoting mesenchymal stem cell differentiation toward the osteoblastic lineage and enhancing bone formation both in vitro and in vivo.50 BMPRIA (BMP receptor type IA) is a transmembrane serine/threonine kinase receptor that initiates intracellular BMP signaling upon ligand binding.51 Upon binding of BMP ligands to BMPRIA, the receptor forms a receptor complex with type II BMP receptors, leading to phosphorylation and activation of downstream receptor-regulated SMAD proteins. The phosphorylated SMADs subsequently interact with SMAD4 to form a transcriptional complex that translocates to the nucleus, where it cooperates with other transcriptional regulators to modulate the expression of genes involved in osteogenic differentiation and bone formation.52 The increased expression of BMPRIA and SMAD4 further suggests an enhanced capacity for BMP signal transduction, as these molecules are critical mediators of canonical TGF-β/BMP signaling. Collectively, these findings suggest that MD/Exos promote osteogenic differentiation through the coordinated upregulation of multiple components of the TGF-β/BMP signaling network. Although the present study demonstrated the therapeutic efficacy of MD/Exos, the specific exosomal cargos responsible for these effects were not investigated. Future studies incorporating comprehensive miRNA and proteomic profiling, will be important to identify the key bioactive molecules and further enhance the therapeutic potential of this platform.
In the present study, we further investigated the effects of MD/Exos on inflammatory macrophages, as these cells are increasingly recognized as key regulators of the bone microenvironment and central coordinators of inflammation, bone remodeling, and tissue repair.53 At the tissue level, the osteoporotic bone microenvironment is characterized by a complex and dynamic interplay of chronic inflammation, cellular dysfunction, metabolic disturbances, and aberrant signaling pathways, all of which collectively impair the regenerative capacity of bone. Importantly, both senile and postmenopausal osteoporosis are associated with a persistent low-grade inflammatory state accompanied by dysregulated expression of multiple cytokines and inflammatory mediators,54 which further amplifies pathological signaling within the bone microenvironment and accelerates disease progression. At the cellular level, inflammatory macrophages drive bone loss by secreting a broad array of cytokines and inflammatory mediators that enhance osteoclast differentiation and activity while suppressing osteogenic differentiation, ultimately disrupting bone homeostasis in favor of excessive bone resorption.55 Therefore, a comprehensive assessment of how MD/Exos modulate inflammatory mediator expression and cytokine networks is crucial for understanding their capacity to remodel the bone microenvironment under chronic inflammatory conditions. We observed that untreated macrophages exhibited increased cellular spreading and the formation of multinucleated giant cells (MGCs), particularly by day 3 post-treatment. The cellular fusion of macrophages into MGCs is a hallmark of chronic inflammation and represents a defining feature of the granulomatous response to persistent infection, foreign bodies, and unresolved tissue injury.56 Interestingly, treatment with MD/Exos markedly reduced the formation of MGCs, suggesting that the platform may attenuate excessive macrophage activation and limit cellular responses associated with chronic inflammation.
Accumulating evidence suggests that MSC-Exos functions as key regulators of intercellular communication within injured tissues by delivering a diverse array of bioactive cargos. Through these molecular components, MSC-Exos are capable of modulating multiple signaling pathways associated with inflammation, tissue repair, angiogenesis, and bone remodeling.57 For example, previous studies have shown that MSC-Exos can influence macrophage-associated inflammatory responses, which are commonly reflected by changes in cytokine expression profiles. These changes often include reduced levels of pro-inflammatory mediators such as TNF-α, IL-1β, and IL-6, together with modulation of anti-inflammatory and tissue-repair-associated cytokines such as IL-10.58 Such cytokine remodeling has been observed across a range of disease models, including cardiovascular and pulmonary inflammatory conditions,59,60 where MSC-Exos attenuates excessive inflammatory signaling and contribute to tissue repair processes.61 Consistent with these observations, our cytokine antibody array analysis revealed that MD/Exos treatment altered the expression profiles of multiple inflammation-associated cytokines. These findings suggest that MD/Exos may influence the inflammatory microenvironment through coordinated regulation of cytokine networks rather than through isolated effects on individual signaling molecules. Given the central role of cytokines in mediating intercellular crosstalk among macrophages, osteoblasts, osteoclasts, and mesenchymal stem cells, alterations in the local cytokine milieu may have important implications for bone remodeling and tissue regeneration by profoundly influencing cellular recruitment, differentiation, survival, and functional activity, ultimately shaping the regenerative capacity of the bone tissue.54 Although cytokine antibody array analysis identified multiple inflammatory mediators altered by MD/Exos treatment, individual cytokines were not further validated in the present study. Future investigations will incorporate targeted ELISA and qPCR analyses, together with functional studies, to confirm the observed cytokine changes and further elucidate the molecular mechanisms underlying the inflammation-regulating effects of MD/Exos.
Bone homeostasis is tightly regulated by the dynamic and coordinated interactions among osteoclasts and osteoblasts, which collectively maintain the balance between bone resorption and bone formation.62 In alignment with these findings, our study sought to evaluate the in vivo efficacy of MD/Exos in mitigating estrogen-deficiency-induced bone loss using a well-established OVX rat model, which closely mimics the clinical pathophysiology of postmenopausal osteoporosis [55]. As expected, untreated OVX rats displayed classic osteoporotic features, including severe trabecular bone deterioration, increased spacing between trabeculae, and significant infiltration of adipocytes into the bone marrow. In contrast, MD/Exos treatment significantly preserved trabecular architecture and reduced adipocyte infiltration within the bone marrow compartment, resulting in histological features that more closely resembled those observed in healthy controls. These findings demonstrate that MD/Exos effectively attenuate estrogen deficiency-induced skeletal degeneration and support the therapeutic potential of this platform for osteoporosis treatment. Importantly, the in vivo observations complement our in vitro findings and collectively support a unified mechanism of action centered on the maintenance of skeletal homeostasis. The favorable therapeutic outcomes observed in the present study are corroborated by substantial preclinical evidence demonstrating the efficacy of stem cell-derived exosome-based therapies in restoring bone homeostasis and promoting skeletal regeneration.63–65 Mechanistically, these therapeutic effects are mediated through the delivery of diverse bioactive cargos capable of modulating key signaling pathways involved in bone remodeling. For instance, Wang et al demonstrated that MSC-derived exosomes enriched with miR-27a effectively suppressed osteoclast differentiation while simultaneously enhancing osteoblast activity, thereby preventing osteoporotic bone loss.66 Likewise, Li et al reported that exosomal miR-186 derived from human BMSCs promoted osteogenesis in ovariectomized rats through regulation of the Hippo signaling pathway, a critical pathway governing cellular proliferation and lineage commitment within bone tissue.67 Together, these studies highlight the ability of stem cell-derived exosomes to serve as biological nanocarriers capable of delivering bioactive cargos that regulate key pathways involved in osteogenesis, osteoclastogenesis, and bone microenvironment homeostasis. The favorable outcomes achieved with MD/Exos in the present study further extend this concept and demonstrate the promise of engineered exosome-based platforms for osteoporosis therapy. While additional mechanistic studies are warranted, the present work provides strong evidence that MD/Exos represents a promising exosome-based therapeutic strategy for the treatment of osteoporosis.
Limitations and Future Perspectives
However, there are many limitations for our study. Although the present study primarily focused on evaluating the therapeutic efficacy of MD/Exos in osteoporosis, future investigations will further explore the functional characteristics of the microdroplet delivery platform. In particular, we will systematically evaluate the effects of different ultrasound parameters, including acoustic intensity, frequency, exposure duration, and treatment intervals, on MD/Exos behavior, cargo release efficiency, and therapeutic outcomes. In vivo, although histological analyses demonstrated that MD/Exos effectively preserved bone microarchitecture following treatment, micro-computed tomography (micro-CT) analysis was not performed. Inclusion of micro-CT analysis in future studies will provide a more comprehensive and quantitative evaluation of bone structural changes. Second, the in vivo safety evaluation in the present study was limited to endpoint histological analysis. Comprehensive systemic safety assessments, including serum biochemical markers of liver and kidney function and long-term toxicity studies, were not conducted. Therefore, the systemic biosafety and long-term biocompatibility of the MD/Exos platform remain to be fully established. In addition, another notable limitation of our study is the absence of an exosome-only treatment group in the in vivo experiments. Although this comparison would have provided valuable insight into the therapeutic efficacy of integrating exosomes into the microdroplet delivery system, the exclusion of the exosome-only group was mainly due to the substantial technical and financial burden associated with producing sufficient quantities of high-purity exosomes required for repeated systemic administration over the 10-week experimental period. Future studies employing optimized exosome production and purification methods will be necessary to enable a direct comparative evaluation between free exosomes and the MD/Exos delivery system.
Conclusion
By marrying the biological activity of MSC-derived exosomes with the microdroplet-based delivery platform, this system provides an effective approach to enhance exosomal contents delivery and improve bone repair in osteoporosis. This platform not only advances the field of bone regeneration but also sets the stage for broader applications of microdroplet-based exosome delivery in regenerative medicine.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was financially supported by the National Natural Science Foundation of China (Grant No. 82560425, 82560664, 82460429), Guizhou Science and Technology Program Project (Grant No. Qiankehe Foundation Basic MS [2025] 370, Basic MS [2025] 410, Basic - ZK[2024] General 296, and - ZK[2023] General 502), Zunyi Science and Technology Program Project (Grant No. Zunshikehe HZ zi [2025] 385), Zunyi Municipal Key Laboratory of Medicinal Biotechnology (Grant No. ZSKH-SYS-[2025] 03), Zunyi Science and Technology Innovation Team for Nutrition and Health (Grant No. Zun KCTD [2025] 89), and Future Eminent Clinician Plan of Zunyi Medical University (Grant No.2022-02).
Disclosure
Jincheng Sima and Junrong Chen are co-first authors for this study. The authors declare that they have no competing interests in this work.
References
1. Munoz M, Robinson K, Shibli-Rahhal A. Bone health and osteoporosis prevention and treatment. Clin Obstet Gynecol. 2020;63(4):770–24. doi:10.1097/GRF.0000000000000572
2. Salari N, Darvishi N, Bartina Y, et al. Global prevalence of osteoporosis among the world older adults: a comprehensive systematic review and meta-analysis. J Orthop Surg Res. 2021;16(1):669. doi:10.1186/s13018-021-02821-8
3. Rinonapoli G, Ruggiero C, Meccariello L, Bisaccia M, Ceccarini P, Caraffa A. Osteoporosis in men: a review of an underestimated bone condition. Int J Mol Sci. 2021;22(4). doi:10.3390/ijms22042105
4. Adejuyigbe B, Kallini J, Chiou D, Kallini JR. Osteoporosis: molecular pathology, diagnostics, and therapeutics. Int J Mol Sci. 2023;24(19). doi:10.3390/ijms241914583
5. Porter JL, Varacallo MA. Osteoporosis(Archived). StatPearls; 2025.
6. Zhao X, Patil S, Xu F, Lin X, Qian A. Role of biomolecules in osteoclasts and their therapeutic potential for osteoporosis. Biomolecules. 2021;11(5). doi:10.3390/biom11050747
7. Ha P, Kwak JH, Zhang Y, et al. Bisphosphonate conjugation enhances the bone-specificity of NELL-1-based systemic therapy for spaceflight-induced bone loss in mice. NPJ Microgravity. 2023;9(1):75. doi:10.1038/s41526-023-00319-7
8. Kendler DL, Cosman F, Stad RK, Ferrari S. Denosumab in the treatment of osteoporosis: 10 years later: a narrative review. Adv Ther. 2022;39(1):58–74. doi:10.1007/s12325-021-01936-y
9. Miller PD. Denosumab: anti-RANKL antibody. Curr Osteoporos Rep. 2009;7(1):18–22. doi:10.1007/s11914-009-0004-5
10. Solling AS, Tsourdi E, Harslof T, Langdahl BL. Denosumab discontinuation. Curr Osteoporos Rep. 2023;21(1):95–103. doi:10.1007/s11914-022-00771-6
11. van Niel G, Carter DRF, Clayton A, Lambert DW, Raposo G, Vader P. Challenges and directions in studying cell-cell communication by extracellular vesicles. Nat Rev Mol Cell Biol. 2022;23(5):369–382. doi:10.1038/s41580-022-00460-3
12. Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol. 2021;16(7):748–759. doi:10.1038/s41565-021-00931-2
13. Di Bella MA. Overview and update on extracellular vesicles: considerations on exosomes and their application in modern medicine. Biology. 2022;11(6). doi:10.3390/biology11060804
14. Ullah I, Subbarao RB, Rho GJ. Human mesenchymal stem cells - current trends and future prospective. Biosci Rep. 2015;35(2). doi:10.1042/BSR20150025
15. Jiao YR, Chen KX, Tang X, et al. Exosomes derived from mesenchymal stem cells in diabetes and diabetic complications. Cell Death Dis. 2024;15(4):271. doi:10.1038/s41419-024-06659-w
16. Tan F, Li X, Wang Z, Li J, Shahzad K, Zheng J. Clinical applications of stem cell-derived exosomes. Signal Transduct Target Ther. 2024;9(1):17. doi:10.1038/s41392-023-01704-0
17. Kou M, Huang L, Yang J, et al. Mesenchymal stem cell-derived extracellular vesicles for immunomodulation and regeneration: a next generation therapeutic tool? Cell Death Dis. 2022;13(7):580. doi:10.1038/s41419-022-05034-x
18. Tzng E, Bayardo N, Yang PC. Current challenges surrounding exosome treatments. Extracell Vesicle. 2023;2:100023. doi:10.1016/j.vesic.2023.100023
19. Morishita M, Takahashi Y, Nishikawa M, et al. Quantitative analysis of tissue distribution of the B16BL6-derived exosomes using a streptavidin-lactadherin fusion protein and iodine-125-labeled biotin derivative after intravenous injection in mice. J Pharm Sci. 2015;104(2):705–713. doi:10.1002/jps.24251
20. Entzian K, Aigner A. Drug delivery by ultrasound-responsive nanocarriers for cancer treatment. Pharmaceutics. 2021;13(8). doi:10.3390/pharmaceutics13081135
21. Zhang Y, Cao J, Jian M, et al. Fabrication of Interleukin-4 encapsulated bioactive microdroplets for regulating inflammation and promoting osteogenesis. Int J Nanomed. 2023;18:2019–2035. doi:10.2147/IJN.S397359
22. Shakya G, Cattaneo M, Guerriero G, Prasanna A, Fiorini S, Supponen O. Ultrasound-responsive microbubbles and nanodroplets: a pathway to targeted drug delivery. Adv Drug Deliv Rev. 2024;206:115178. doi:10.1016/j.addr.2023.115178
23. Rovers TA, Sala G, van der Linden E, Meinders MB. Effect of temperature and pressure on the stability of protein microbubbles. ACS Appl Mater Interfaces. 2016;8(1):333–340. doi:10.1021/acsami.5b08527
24. Zhang Y, Dang Y, Huang M, Ma Y, Zhang D, Wang X. Development of bioactive and ultrasound-responsive microdroplets for preventing ovariectomy (OVX)-induced osteoporosis. J Mater Chem B. 2023;11(47):11344–11356. doi:10.1039/d3tb01726e
25. Zhang Y, Huo M, Wang Y, et al. A tailored bioactive 3D porous poly(lactic-acid)-exosome scaffold with osteo-immunomodulatory and osteogenic differentiation properties. J Biol Eng. 2022;16(1):22. doi:10.1186/s13036-022-00301-z
26. Cory G. Scratch-wound assay. Methods Mol Biol. 2011;769:25–30. doi:10.1007/978-1-61779-207-6_2
27. Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3(6):1101–1108. doi:10.1038/nprot.2008.73
28. Wei F, Neal CJ, Sakthivel TS, Kean T, Seal S, Coathup MJ. Multi-functional cerium oxide nanoparticles regulate inflammation and enhance osteogenesis. Mater Sci Eng C Mater Biol Appl. 2021;124:112041. doi:10.1016/j.msec.2021.112041
29. Wang X, Ao J, Lu H, et al. Osteoimmune modulation and guided osteogenesis promoted by barrier membranes incorporated with S-nitrosoglutathione (GSNO) and mesenchymal stem cell-derived exosomes. Int J Nanomed. 2020;15:3483–3496. doi:10.2147/IJN.S248741
30. Vaishya R, Iyengar KP, Jain VK, Vaish A. Demystifying the risk factors and preventive measures for osteoporosis. Indian J Orthop. 2023;57(Suppl 1):94–104. doi:10.1007/s43465-023-00998-0
31. Palakurthi SS, Shah B, Kapre S, et al. A comprehensive review of challenges and advances in exosome-based drug delivery systems. Nanoscale Adv. 2024;6(23):5803–5826. doi:10.1039/d4na00501e
32. Hu W, Wang W, Chen Z, Chen Y, Wang Z. Engineered exosomes and composite biomaterials for tissue regeneration. Theranostics. 2024;14(5):2099–2126. doi:10.7150/thno.93088
33. Li T, Zhao J, Yuan J, et al. Harnessing engineered exosomes as METTL3 carriers: enhancing osteogenesis and suppressing lipogenesis in bone marrow mesenchymal stem cells for postmenopausal osteoporosis treatment. Mater Today Bio. 2025;32:101648. doi:10.1016/j.mtbio.2025.101648
34. Wang F, Yang S, Zhu Y, et al. Mechanical energy-charged navigating pump repair natural vascular scaffolds to reverse osteoporosis. Mater Today Bio. 2025;35:102431. doi:10.1016/j.mtbio.2025.102431
35. Tang R, Liu G, Wang C, et al. Transcriptomics guided engineering of exosome-encapsulated bifunctional nanosheets targeting the immune-PI3K/Akt axis for osteoporosis therapy. Adv Sci. 2025;12(47):e11327. doi:10.1002/advs.202511327
36. Long C, Li X, Fan Y, et al. Effects and mechanisms of engineered exosomes pretreated with Scutellaria baicalensis Georgi on osteoporosis. PLoS One. 2025;20(10):e0333897. doi:10.1371/journal.pone.0333897
37. Si Y, Dong S, Li M, et al. Curcumin-encapsulated exosomes in bisphosphonate-modified hydrogel microspheres promote bone repair through macrophage polarization and DNA damage mitigation. Mater Today Bio. 2025;32:101874. doi:10.1016/j.mtbio.2025.101874
38. Xiang K, Hao M, Zhang Z, Zhang K, Sun H, Zhang L. Engineering 3D-BMSC exosome-based hydrogels that collaboratively regulate bone microenvironment and promote osteogenesis for enhanced cell-free bone regeneration. Mater Today Bio. 2025;32:101881. doi:10.1016/j.mtbio.2025.101881
39. Luo P, Zhang Y, Huang M, Luo G, Ma Y, Wang X. Microdroplets encapsulated with NFATc1-siRNA and exosomes-derived from MSCs onto 3D Porous PLA scaffold for regulating osteoclastogenesis and promoting osteogenesis. Int J Nanomed. 2024;19:3423–3440. doi:10.2147/IJN.S443413
40. Ren S, Lin Y, Liu W, Yang L, Zhao M. MSC-Exos: important active factor of bone regeneration. Front Bioeng Biotechnol. 2023;11:1136453. doi:10.3389/fbioe.2023.1136453
41. Zhang Y, Bai J, Xiao B, Li C. BMSC-derived exosomes promote osteoporosis alleviation via M2 macrophage polarization. Mol Med. 2024;30(1):220. doi:10.1186/s10020-024-00904-w
42. Al-Sharabi N, Mohamed-Ahmed S, Shanbhag S, et al. Osteogenic human MSC-derived extracellular vesicles regulate MSC activity and osteogenic differentiation and promote bone regeneration in a rat calvarial defect model. Stem Cell Res Ther. 2024;15(1):33. doi:10.1186/s13287-024-03639-x
43. Tang J, Wang X, Lin X, Wu C. Mesenchymal stem cell-derived extracellular vesicles: a regulator and carrier for targeting bone-related diseases. Cell Death Discov. 2024;10(1):212. doi:10.1038/s41420-024-01973-w
44. Wang B, Lyu FJ, Deng Z, et al. Therapeutic potential of stem cell-derived exosomes for bone tissue regeneration around prostheses. J Orthop Translat. 2025;52:85–96. doi:10.1016/j.jot.2025.03.007
45. Dou X, Wei X, Liu G, et al. Effect of porous tantalum on promoting the osteogenic differentiation of bone marrow mesenchymal stem cells in vitro through the MAPK/ERK signal pathway. J Orthop Translat. 2019;19:81–93. doi:10.1016/j.jot.2019.03.006
46. Torrecillas-Baena B, Pulido-Escribano V, Dorado G, Galvez-Moreno MA, Camacho-Cardenosa M, Casado-Diaz A. Clinical potential of mesenchymal stem cell-derived exosomes in bone regeneration. J Clin Med. 2023;12(13). doi:10.3390/jcm12134385
47. Li D, Li D, Wang Z, et al. Signaling pathways activated and regulated by stem cell-derived exosome therapy. Cell Biosci. 2024;14(1):105. doi:10.1186/s13578-024-01277-7
48. Lu J, Wang QY, Sheng JG. Exosomes in the repair of bone defects: next-generation therapeutic tools for the treatment of nonunion. Biomed Res Int. 2019;2019:1983131. doi:10.1155/2019/1983131
49. Tan SHS, Wong JRY, Sim SJY, et al. Mesenchymal stem cell exosomes in bone regenerative strategies-a systematic review of preclinical studies. Mater Today Bio. 2020;7:100067. doi:10.1016/j.mtbio.2020.100067
50. Beederman M, Lamplot JD, Nan G, et al. BMP signaling in mesenchymal stem cell differentiation and bone formation. J Biomed Sci Eng. 2013;6(8A):32–52. doi:10.4236/jbise.2013.68A1004
51. Sanchez-Duffhues G, Williams E, Goumans MJ, Heldin CH, Ten Dijke P. Bone morphogenetic protein receptors: structure, function and targeting by selective small molecule kinase inhibitors. Bone. 2020;138:115472. doi:10.1016/j.bone.2020.115472
52. Wu M, Wu S, Chen W, Li YP. The roles and regulatory mechanisms of TGF-beta and BMP signaling in bone and cartilage development, homeostasis and disease. Cell Res. 2024;34(2):101–123. doi:10.1038/s41422-023-00918-9
53. Weivoda MM, Bradley EW. Macrophages and Bone Remodeling. J Bone Miner Res. 2023;38(3):359–369. doi:10.1002/jbmr.4773
54. Lu X, Zhang A, Zhang C, Guo D, Li ZA, Xie D. Targeting the osteoporotic bone microenvironment: mechanistic insight and therapeutic biomaterials for accelerating bone regeneration. Bioact Mater. 2026;62:96–122. doi:10.1016/j.bioactmat.2026.02.024
55. Wang W, Liu H, Liu T, Yang H, He F. Insights into the role of macrophage polarization in the pathogenesis of osteoporosis. Oxid Med Cell Longev. 2022;2022:2485959. doi:10.1155/2022/2485959
56. Sissons JR, Peschon JJ, Schmitz F, Suen R, Gilchrist M, Aderem A. Cutting edge: microRNA regulation of macrophage fusion into multinucleated giant cells. J Immunol. 2012;189(1):23–27. doi:10.4049/jimmunol.1102477
57. Wu T, Liu Y, Wang S, Shi C. MSC-derived extracellular vesicles: roles and molecular mechanisms for tissue repair. Int J Nanomed. 2025;20:7953–7974. doi:10.2147/IJN.S525394
58. Chen S, Saeed A, Liu Q, et al. Macrophages in immunoregulation and therapeutics. Signal Transduct Target Ther. 2023;8(1):207. doi:10.1038/s41392-023-01452-1
59. Xu R, Zhang F, Chai R, et al. Exosomes derived from pro-inflammatory bone marrow-derived mesenchymal stem cells reduce inflammation and myocardial injury via mediating macrophage polarization. J Cell Mol Med. 2019;23(11):7617–7631. doi:10.1111/jcmm.14635
60. Sun X, Shan A, Wei Z, Xu B. Intravenous mesenchymal stem cell-derived exosomes ameliorate myocardial inflammation in the dilated cardiomyopathy. Biochem Biophys Res Commun. 2018;503(4):2611–2618. doi:10.1016/j.bbrc.2018.08.012
61. Wang J, Xia J, Huang R, et al. Mesenchymal stem cell-derived extracellular vesicles alter disease outcomes via endorsement of macrophage polarization. Stem Cell Res Ther. 2020;11(1):424. doi:10.1186/s13287-020-01937-8
62. Xia D, Qian Q, Wang S, Dong X, Liu Y. Alendronate functionalized bone-targeting pomolic acid liposomes restore bone homeostasis for osteoporosis treatment. Int J Nanomed. 2024;19:7983–7996. doi:10.2147/IJN.S462514
63. Hu Y, Zhang Y, Ni CY, et al. Human umbilical cord mesenchymal stromal cells-derived extracellular vesicles exert potent bone protective effects by CLEC11A-mediated regulation of bone metabolism. Theranostics. 2020;10(5):2293–2308. doi:10.7150/thno.39238
64. Gong L, Chen B, Zhang J, et al. Human ESC-sEVs alleviate age-related bone loss by rejuvenating senescent bone marrow-derived mesenchymal stem cells. J Extracell Vesicles. 2020;9(1):1800971. doi:10.1080/20013078.2020.1800971
65. Chen CY, Rao SS, Tan YJ, et al. Extracellular vesicles from human urine-derived stem cells prevent osteoporosis by transferring CTHRC1 and OPG. Bone Res. 2019;7:18. doi:10.1038/s41413-019-0056-9
66. Wang Y, Zhou X, Wang D. Mesenchymal stem cell-derived extracellular vesicles inhibit osteoporosis via microRNA-27a-induced inhibition of DKK2-mediated Wnt/beta-catenin pathway. Inflammation. 2022;45(2):780–799. doi:10.1007/s10753-021-01583-z
67. Li L, Zhou X, Zhang JT, et al. Exosomal miR-186 derived from BMSCs promote osteogenesis through hippo signaling pathway in postmenopausal osteoporosis. J Orthop Surg Res. 2021;16(1):23. doi:10.1186/s13018-020-02160-0
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Knockdown of HDAC9 Inhibits Osteogenic Differentiation of Human Bone Marrow Mesenchymal Stem Cells Partially by Suppressing the MAPK Signaling Pathway
Wang B, Gong S, Han L, Shao W, Li Z, Xu J, Lv X, Xiao B, Feng Y
Clinical Interventions in Aging 2022, 17:777-787
Published Date: 12 May 2022
Fabrication of Interleukin-4 Encapsulated Bioactive Microdroplets for Regulating Inflammation and Promoting Osteogenesis
Zhang Y, Cao J, Jian M, Zhou Z, Anwar N, Xiao L, Ma Y, Zhang D, Zhang J, Wang X
International Journal of Nanomedicine 2023, 18:2019-2035
Published Date: 19 April 2023
Nanotechnology in Orthopedic Care: Advances in Drug Delivery, Implants, and Biocompatibility Considerations
Xia L, Zhou C, Li Q, Liu L, Jiang C, Dai H, Zhang H, Zhao J, Liang W
International Journal of Nanomedicine 2025, 20:9251-9274
Published Date: 21 July 2025