Back to Journals » Cancer Management and Research » Volume 12
4E-BP1Thr46 Phosphorylation Association with Poor Prognosis in Quantitative Phosphoproteomics of Portal Vein Tumor Thrombus Revealed that 4E-BP1Thr46 Phosphorylation is Associated with Poor Prognosis in HCC
Authors Lin X ![]() , Huang Y
, Huang Y ![]() , Sun Y, Tan X
, Sun Y, Tan X ![]() , Ouyang J, Zhao B
, Ouyang J, Zhao B ![]() , Wang Y
, Wang Y ![]() , Xing X
, Xing X ![]() , Liu J
, Liu J ![]()
Received 12 September 2019
Accepted for publication 30 November 2019
Published 7 January 2020 Volume 2020:12 Pages 103—115
DOI https://doi.org/10.2147/CMAR.S230849
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ahmet Emre Eşkazan
Xincong Lin,1,2,* Yao Huang,1,2,* Ying Sun,2 Xionghong Tan,2 Jiahe Ouyang,2 Bixing Zhao,2 Yingchao Wang,2 Xiaohua Xing,1,2 Jingfeng Liu1,2
1The First Affiliated Hospital of Fujian Medical University, Fuzhou 350007, People’s Republic of China; 2The United Innovation of Mengchao Hepatobiliary Technology Key Laboratory of Fujian Province, Mengchao Hepatobiliary Hospital of Fujian Medical University, Fuzhou 350025, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaohua Xing
The First Affiliated Hospital of Fujian Medical University, Fuzhou 350007, People’s Republic of China
Tel/Fax +86591-83705927
Email [email protected]
Jingfeng Liu
The First Affiliated Hospital of Fujian Medical University, Jiaotong Road 88, Fuzhou 350007, Fujian Province, People’s Republic of China
Tel/Fax +86591-83705927
Email [email protected]
Purpose: Early formation of portal vein tumor thrombosis (PVTT) is a key characteristic of hepatocellular carcinoma (HCC) metastasis, but to date, the aetiology of PVTT in HCC metastasis is largely unknown. We aim to find highly sensitive and specific biomarkers for the prediction of HCC prognosis.
Patients and methods: We used isobaric tags for relative and absolute quantitation (iTRAQ) based quantitative phosphoproteomics approach to investigate the molecular signatures of the HCC with PVTT in primary HCC tissues, surrounding non-cancerous tissues and PVTT tissues. The different proteome profiles in three groups were investigated and might reveal different underlying molecular mechanisms.
Results: In total, we identified 1745 phosphoproteins with 2724 phosphopeptides and 4594 phosphorylation sites in three groups. Among these phosphoproteins, 80 phosphoproteins were dysregulated in PVTT/Pan group, 51 phosphoproteins were dysregulated in HCC/Pan group, and 10 phosphoproteins were dysregulated in PVTT/HCC group. Furthermore, the phosphorylation of 4E-BP1 was elevated in HCC tissues and PVTT tissues in comparison with surrounding non-cancerous tissues, and the elevated fold change of phosphorylation level was higher than that in expression level of 4E-BP1. The further IHC analysis in acohort of 20 HCC tissues showed that the phosphorylation of 4E-BP1 on Thr46 might be closely related to HCC prognosis.
Conclusion: The high phosphorylation level of 4E-BP1Thr46 might serve as a biomarker for the diagnosis of early recurrence and metastasis of HCC.
Keywords: hepatocellular carcinoma, portal vein tumor thrombus, quantitative phosphoproteomics, 4E-BP1Thr46, prognosis
Introduction
Hepatocellular carcinoma (HCC) is the third leading cause of cancer-related death worldwide, and is responsible for a largest number of deaths (782 000 deaths, 8.2% of the total in 2018) according to the 2018 investigation of International Agency for Research on Cancer (IARC).1 Although surgical excision has made great progress, the prognosis of HCC patients remains poor. Approximately 70% of HCC patients suffer from metastasis and recurrence within 5 years after surgery in the residual liver after curative resection.2–7 But the mechanism of metastasis and recurrence of HCC was still unclear, which was the main cause for short-term survival and high fatality of HCC. Therefore, it is urgent to screen highly sensitive and specific biomarkers for the prediction of HCC prognosis and to in-depth investigate the metastasis/recurrence mechanisms.
Portal vein tumor thrombosis (PVTT) was an important prognostic factor and predictor for HCC metastasis and recurrence, which was a special type of HCC metastasis and caused by the invasion of HCC cells into the portal vein.8 Early formation of PVTT was a key characteristic of HCC metastasis, but the aetiology of PVTT in HCC metastasis was largely unknown, so far. The clear explanation of PVTT formation under HCC metastasis was necessary to investigate the metastasis mechanisms. Up to now, there have been only two reports on the PVTT associated proteomics. One was performed on 20 pairs of HCC tissues with PVTT and without PVTT, and PDIA6 and ApoA-I were identified to be closely related to vascular invasion feature of HCC through two-dimensional polyacrylamide gel electrophoresis combined with matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS).9 The other also utilized two-dimensional electrophoresis and tandem MS to compare and identify differentially expressed proteins between a type of distinct PVTT (dPVTT) and parenchyma tumor (PT) tissues, and C-kit was identified to be useful as a potential marker to distinguish dPVTT from PT.10 Phosphorylation is one of the major types of PTMs in eukaryotic species. Phosphorylation or dephosphorylation usually activates many different signal pathways, which are always associated with various diseases, including cancers.11,12 The development of proteomics has promoted us to detect the dynamic change of the whole phosphorylation in cancer samples easily.13–15 Based on various phosphopeptides enrichment technologies, aberrant kinase signaling pathways and phosphorylation sites (p-sites) can be identified by quantitative phosphoproteomics in HCC samples, which make us better understanding of the mechanisms of HCC progression. But the phosphoproteomics of HCC with PVTT has not been reported, which plays a critical role in HCC metastasis.
In this study, we aimed to screen highly sensitive and specific biomarkers for the prediction of HCC prognosis and investigate the molecular signatures of the HCC with PVTT, by quantitatively comparing the overall phosphoproteome of the primary HCC tissues, surrounding non-cancerous tissues and PVTT tissues using iTRAQ coupling with liquid chromatography-tandem mass spectrometry (LC-MS/MS). The results would help us better understanding of the mechanisms of PVTT, which was essential to HCC metastasis and recurrence.
Materials and Methods
Sample Collection
Tissue samples, including the cancerous, surrounding non-cancerous and portal vein tumor thrombus (PVTT) tissues were obtained from 6 primary HCC patients with portal vein tumor thrombus, respectively. All patients have undergone radical surgery at Mengchao Hepatobiliary Hospital of Fujian Medical University from January 2014 to February 2016. The absence of intrahepatic recurrence/metastasis in the residual liver was monitored by ultrasonography (US), computer tomography (CT) scan and angiography. All of the patients followed the Enrollment Eligibility Criteria: (1) the patient was diagnosed with HCC with portal vein tumor thrombus by post-operative pathological examinations; (2) Pre-operative serum HBs Ag (Hepatitis B surface antigen) positive, but HBs Ab (Hepatitis B surface antibody) negative and HCV (Hepatitis C virus) negative; (3) subject to the standard radical resection[7]: no distal metastasis was revealed in both pre and intra-operative examinations; no lesion was found in the rest of the liver during intra-operative ultrasonic scan; no cancer cell was found in the incisal margin at the post-operative pathological examinations; no recurrent/metastatic lesion was found at the ultrasonic and CT scan during the return visit after 2 months of surgery; (4) the elevated pre-operative serum AFP should decline to normal level after 2 months of post-operation; (5) the patient did not undergo any other intervention or therapies before surgery; and (6) The patients were followed every 3–6 months for the next 5 years after the surgery.
The project was approved for using human samples by the Institution Review Board of Mengchao Hepatobiliary Hospital of Fujian Medical University. The written informed consent was obtained from each participant before the operation. The use of clinical specimens is completely compliance to the “Declaration of Helsinki”.
Protein Preparation and iTRAQ Labeling
The tissues from patients were divided into 3 groups: cancerous tissues from HCC patients (HCC group, n = 6); surrounding non-cancerous tissues from HCC patients (Pan group, n = 6); and portal vein tumor thrombus tissues from HCC patients (PVTT group, n = 6). The protein preparation and iTRAQ labeling followed our previous reported procedures with modification.16 For each patient, proteins from 3 groups were, respectively, extracted with lysis buffer (9M Urea, 10 mM Tris-HCl (pH 8.0), 75 mM NaCl, 10 mM IAA, 1 mM NaF,1 mM β-glycerophosphate, 1 mM Sodium orthovanadate (Na3VO4), 1 mM Sodium pyrophosphate, 1 mM sodium dihydrogen phosphate, 1 mM PMSF, 1% phosphatase inhibitor cocktail 2 (Sigma, St. Louis, MO USA), 1% phosphatase inhibitor cocktail 3 (Sigma)), and 1 tablet of EDTA-free protease inhibitor cocktail (Roche, Basel, Switzerland) for every 10 mL lysis buffer. Then, the samples were followed by tissue homogenization on ice and precipitated by ice-cold acetone. After centrifugation at 17,000 g for 10 min at 4°C, the supernatant was collected and the protein concentration of the supernatant was determined by BCA assay (TransGen Biotech, Beijing, China) following the manufacturer’s protocol. After that, 100 μg proteins of each group was reduced by 10 mM DTT at 55°C for 30 min and alkylated by 50 mM IAA in the darkness at room temperature for 30 min. And the proteins were digested with trypsin via the FASP protocol with spin ultrafiltration units of molecular weight cut off 10,000 Dalton. Finally, the peptide mixture was further labeled using the iTRAQ reagent kit (AB SCIEX, USA) as follows: 3 groups (HCC group, Pan group and PVTT group) from one patient were labeled with 113, 114 and 115 isobaric tag, respectively; and the peptides from biological repetitions of above 3 groups from another one patient were labeled with 116, 117 and 118, respectively. In total, the iTRAQ 8-plex labeling was independently repeated 3 times, defining as A, B and C. And the mixed samples of 18 groups with equal amount were labeled with 119 in every 8-plex labeling, as an internal standard to balance each 8-plex labeling. The labeled peptides were mixed with equal amount in every 8-plex labeling and desalted with the Sep-Pak Vac C18 cartridges and then dried in a vacuum centrifuge for further usage.
Phosphopeptides Enrichment
Magnetic Titanium Dioxide Phosphopeptide Enrichment Kit (Thermo Fisher Scientific, Bremen, Germany) was used to enrich the phosphopeptides. Briefly, TiO2 magnetic beads were firstly washed by binding buffer three times. Then, the peptide mixtures were incubated with magnetic beads in loading buffer (80% ACN, 2% FA). After that, the magnetic beads were washed in turn by binding buffer and washing buffer. Finally, the phosphopeptides were eluted by elution buffer and dried in a vacuum centrifuge for further usage.
LC-MS/MS Analysis
The LC-MS/MS analysis follows our previously reported protocol with certain modification.17 Briefly, the phosphopeptides were separated by an EASY-nLC1000 system (Thermo Fisher Scientific, Bremen, Germany) and analyzed by a quadrupole-Orbitrap mass spectrometer (Q-Exactive Plus) (Thermo Fisher Scientific, Bremen, Germany) equipped with an online nano-electrospray ion source. Phosphopeptides were loaded onto the trap column (Thermo Scientific Acclaim PepMap C18, 100 μm × 2 cm) with a flow of 10 μL/min, and subsequently separated on the analytical column (Acclaim PepMap C18, 75 μm × 150 cm) with a gradient of 0% to 7% solvent B for 5 min, 7% to 28% solvent B for 105 min, 28% to 80% solvent B for 5 min followed by isocratic conditions at 80% solvent B for 5 min with a flow rate of 300 nL/min at 40°C.
The Q-Exactive Plus mass spectrometer was operated in the data-dependent mode to switch automatically between MS and MS/MS acquisition. The electrospray voltage of 2.1 kV at the inlet of the mass spectrometer was used. Survey full-scan MS spectra (m/z 350–1200) were acquired with a mass resolution of 70 K, followed by 15 sequential high energy collisional dissociation (HCD) MS/MS scans with a resolution of 17.5 K. In all cases, one microscan was recorded using dynamic exclusion of 30 s.
Data Analysis
The data analysis procedure was similar to our previous reports with certain adaption.18 Briefly, MS data acquiring were processed using Proteome Discoverer (Thermo Fisher Scientific, Bremen, Germany; version 2.1) against the uniprot human_database (released at 2014-04-10, 20,264 entries). The enzyme specificity of trypsin was set and maximally up to two missed cleavages were allowed. In the search, precursor mass tolerance was 10 parts per million (ppm) and fragment mass tolerance was 0.02 Da. The research also includes variable modifications of oxidation of methionine, phospho modification on S, T, Y (only for phosphoproteomics) and iTRAQ 8-plex labeling of tyrosine and fixed modification of carbamidomethylation of cysteine, iTRAQ modification of peptide N-terminus and lysine residues, were specified as variable modifications. For peptide and protein identification, the cutoff false discovery rates (FDR) were both set at 0.01.
Bioinformatics Analysis
The significant dysregulation was phosphoproteins or phosphopeptides with fold change >1.2 or <0.83 (log2 ratio lower than −0.26 or higher than 0.26) and p value <0.05. The Gene Ontology (GO) annotation and characteristic pathway analysis using DAVID Bioinformatics Resources 6.7 (https://david.ncifcrf.gov/). And the online tool Motif-X was utilized to predict the potential model of amino acid sequences in particular positions of ubiquitinated-15-mers (seven amino acids upstream of the ubiquitination site and seven downstream). And the statistical analysis in HCC patients was paired T-test for comparison of cancerous and their adjacent tissues, while unpaired T-test for different cancerous tissues. The p<0.05 was considered as statistically significant.
Immunoblot and Immunohistochemistry
Immunoblotting was performed as previously described.16 Briefly, HCC tissues were extracted as described above, separated by SDS-PAGE and transferred onto the NC membranes (Millipore, Bradford, MA). Then, the membranes were blocked using TBST buffer containing 5% BSA for 2 h at room temperature, followed by incubation with the primary antibody (anti-4E-BP1, 1/5000 dilution19 and anti-p-4E-BP1, 1/2000 dilution20) overnight at 4°C. Afterwards, the membranes were incubated with by corresponding secondary antibody for 1 hr at room temperature; after extensive washing with TBST buffer, the blots were revealed using enhanced chemoluminescence reagents and visualized by autoradiography.
Immunohistochemistry was performed on HCC histological section as previously described.21 Briefly, the slides were incubated in 1% hydrogen peroxide for 30 min to block endogenous peroxidase activity, and then, respectively, incubated with the primary antibody (anti-4E-BP1, 1/200 dilution and anti-p-4E-BP1, 1/200 dilution) for 30 min. Afterwards, the slides were washed, treated with the envision FLEX/HRP system for 20 min and revealed with the envision FLEX-DAB chromogen (Dako) and with Mayer’s Hematoxylin (Lille’s Modification) Histological Staining Regents (Dako) for 3 mins and distilled by water for 5 mins. The results were independently assessed by two pathologists double-blindly. All tissues were manually scored as 0 (negative), 1 (weak), 2 (strong) or 3 (very strong). Each case was considered to below expression if the final score was 0–1, while high expression if the final score was 2–3.
Results
Analysis of Phosphoproteome Profile of HCC with PVTT
The schematic overview of the experimental workflow is given in Figure 1, and the details of the optimized protocol were described in the Materials and Methods section. The proteomics dataset was used to normalize the change of phosphoproteins abundance against the changes in total protein levels, avoiding fluctuations due to variations in protein amount. In detail, the phosphorylation level was normalized against the standard proteomics using the ratio of phosphorylation level and protein level if these proteins were quantified in both phosphoproteomics and proteomics.
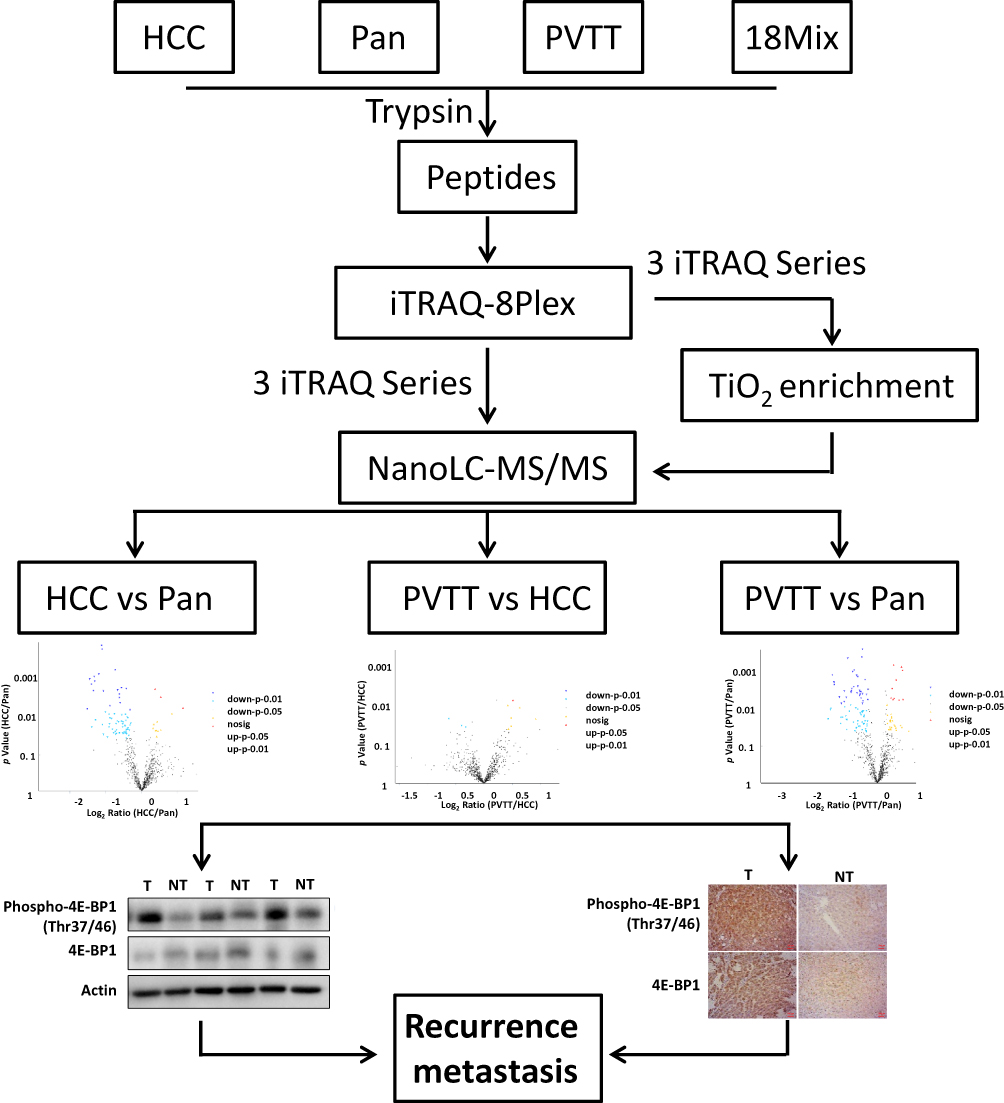 |
Figure 1 Experimental workflow. Total extracts of HCC tissues, surrounding non-cancerous tissues and their PVTT tissues were trypsin digested and directly labeled with iTRAQ-8plex. Phosphopeptides were then enriched using Magnetic Titanium Dioxide Phosphopeptide Enrichment Kit. The phosphoproteome was then analyzed through LC-MS/MS, and the phosphoprotein expression profile alternations between every two groups were performed. Finally, the candidates were verified in HCC patients by Western-Blot and Immunohistochemistry (IHC). |
In this study, all identified phosphopeptides fell into a precursor mass window of ±10 ppm with a mean mass error of only 0.50 ppm (Figure 2A), which suggested the accuracy of the phosphoproteome results. In total, we identified 1745 phosphoproteins from the HCC tissues, pan-cancerous tissue and their PVTT tissues. And the distribution of molecular weight (MW) and isoelectric point (pI) of the phosphoproteins were consistent with the previous studies, which suggesting that the overall phosphoproteome datasets of the HCC tissues had no strong bias (Figure 2B and C). Among these phosphoproteins, there were 2724 phosphopeptides with 4594 phosphorylation sites, and the phosphorylated serine (p-Ser) occupied a large proportion of about 89% of the identified p-sites, while the phosphorylated threonine (p-Thr) and tyrosine (p-Tyr) were relatively less, which demonstrated that the phosphorylation event mainly occurred in serine (Figure 2D). Furthermore, approximately 64% of phosphorylated peptides owned a single phosphorylation site, and the average number of phosphorylation sites in a peptide was 1.69, which indicated that the results of this study had no biases (Figure 2E). In addition, the number of the overlapped phosphoproteins among 3 iTRAQ 8-plex labeling replicates was 546, which were focused on for further analysis (Figure 2F, Table S1).
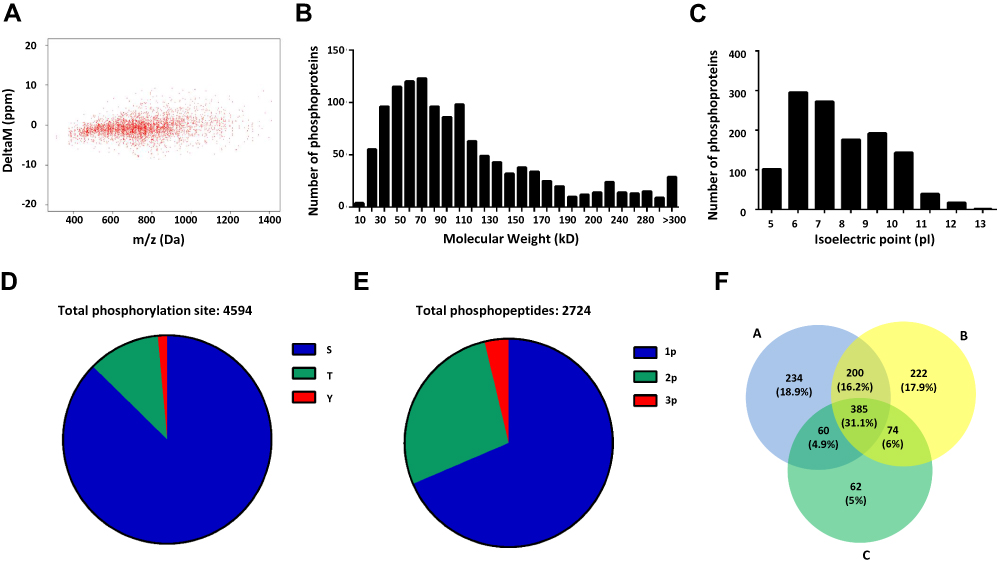 |
Figure 2 Quantitative phosphoproteome profiling of HCC with PVTT. (A) The precursor mass window of the identified phosphopeptides. (B) The distribution of molecular weight of the identified phosphoproteins. (C) The distribution of isoelectric point of the identified phosphoproteins. (D) Distribution of phosphorylation serine (S), phosphorylation threonine (T) and phosphorylation tyrosine (Y) sites. (E) Distribution of phosphopeptides depending on their number of p-sites. (F) Venn diagrams showed the overlaps of phosphoproteins identification in 3 repeated experiments. |
Dysregulation of Phosphoproteome Associated with PVTT
Among the 546 phosphoproteins overlapped in the 3 iTRAQ 8-plex labeling replicates, we set a threshold of mean fold change >1.2 or <0.83 (log2 ratio lower than −0.26 or higher than 0.26) with a p value less than 0.05 (paired T-test) to identify phosphoproteins significantly altered in HCC. According to this stringent criterion, 80 phosphoproteins were dysregulated in PVTT group comparing with Pan group (24 up- and 56 down-regulated), 51 phosphoproteins were dysregulated in HCC group comparing with Pan group (11 up- and 40 down-regulated), and 10 phosphoproteins were dysregulated in PVTT group comparing with HCC group (2 up- and 8 down-regulated) (Table 1, Table S2, Table S3, Table S4 and Figure 3A). These results revealed that the important phosphorylation events were associated with the initiation, development and metastasis of HCC.
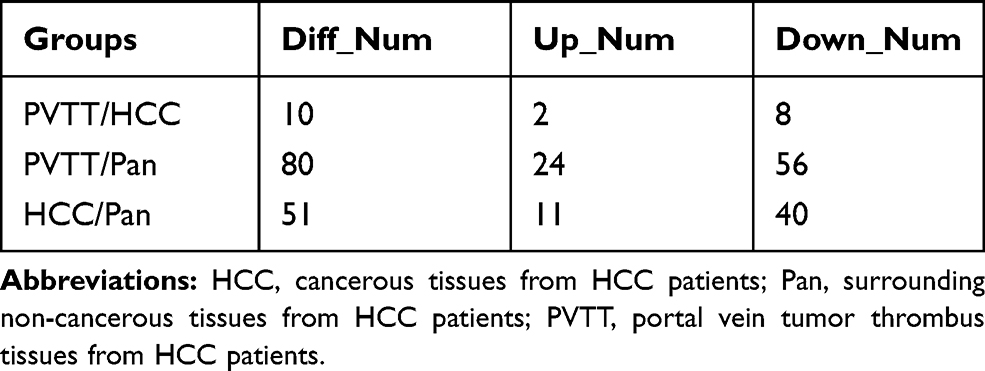 |
Table 1 The Number of the Dysregulated Phosphoproteins in Every Two Groups |
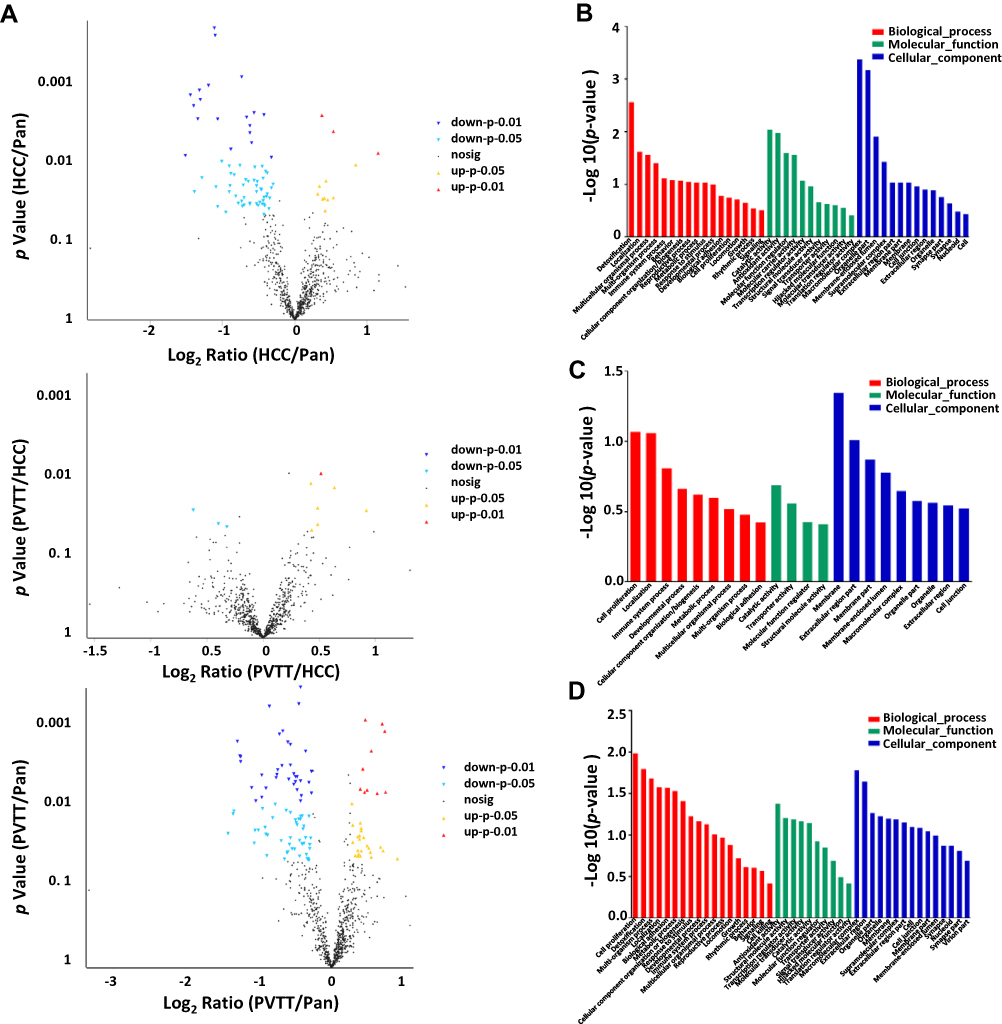 |
Figure 3 The alteration analysis and the involved biological process analysis associated with the dysregulated phosphorylation. (A) Volcano plot represented the phosphoprotein abundance changes in the HCC/Pan group, PVTT/HCC group and PVTT/Pan group, respectively. (B–D) The main biological processes associated with the dysregulated phosphorylation in the HCC/Pan group, PVTT/HCC group and PVTT/Pan group, respectively. |
To further reveal the importance of such phosphorylation events in HCC, we analyzed such phosphoprotein involved biological processes, molecular functions and cellular component by GO analysis. As shown in Figure 3B, the top 10 biological processes were enriched in HCC/Pan group according to the phosphoproteins showing up- or down-regulated phosphorylation. The liver is an important detoxifying organ, so it was reasonable that detoxification was top enriched in HCC.22 Multicellular organismal process, Multi-organism process, Cellular component organization/biogenesis and Metabolic process belong to material metabolism, which played an important role in the tumorigenesis and development of HCC. And the dysregulation of immune system process was associated with the initiation of HCC, which all might be the cause of hepatocarcinogenesis. And the molecular functions and cellular component of these dysregulated phosphoproteins were extensive and reasonable, which suggested the function of these depends on their molecular type and localization in the cell or organism. All of these revealed that the dysregulated phosphoproteins in HCC/Pan were associated with the occurrence and development of HCC.
In PVTT/HCC, the top 10 biological processes were enriched according to the phosphoproteins showing up- or down-regulated phosphorylation. Immune system process, cell proliferation and biological adhesion were actively associated with tumor invasion or metastasis, which might cause HCC invasion or metastasis.23–26 And the dysregulation of material metabolism was essential to HCC invasion or metastasis. And the molecular function and the cellular component of these phosphoproteins ensured the smooth completion of a series of biological processes related to invasion or metastasis (Figure 3C). The above results illustrated that the dysregulated phosphoproteins in PVTT/HCC were associated with the invasion and metastasis of HCC.
As a matter of course, the biological processes enriched in PVTT/Pan included not only the biological processes related to HCC occurrence and development, but also the biological processes related to HCC invasion and metastasis, which was consistent with previous reports and theoretical basis. Similarly, the molecular function and the cellular component of these phosphoproteins mainly enriched in Extracellular region, Membrane and Cell junction were also contributed to the occurrence and development, as well as invasion and metastasis of HCC (Figure 3D). These results suggested that the phosphoproteins involved in PVTT/Pan not only related to HCC occurrence and development, but also related to HCC invasion and metastasis.
The Key Signaling Pathway Associated to PVTT
To further reveal the possible phosphorylation events of HCC with PVTT, KEGG annotation was applied to analyze the signaling pathways that dysregulated phosphoproteins were involved in the analyzed samples. The analysis of KEGG annotations demonstrated that the top two enriched pathway of dysregulated phosphoproteins in HCC/Pan was PI3K-Akt signaling pathway and AMPK signaling pathway (Figure 4A). It has been reported that the PI3K/AKT signaling pathway was activated in HCC,27,28 which played an essential role in the development of HCC by enhancing tumor cell proliferation, differentiation and apoptosis.29,30 AMPK, a AMP‐activated serine/threonine protein kinase (AMPK), was well known for its controlling energy metabolism, which played an important role in the tumorigenesis and development of HCC. And the activation of AMPK may oppose HCC progression, decreased AMPK activity was associated with poor prognosis and aggressive clinicopathologic features.31,32 In this study, the overall decreased phosphorylation of AMPK signaling pathway might regulate the occurrence and development of HCC.
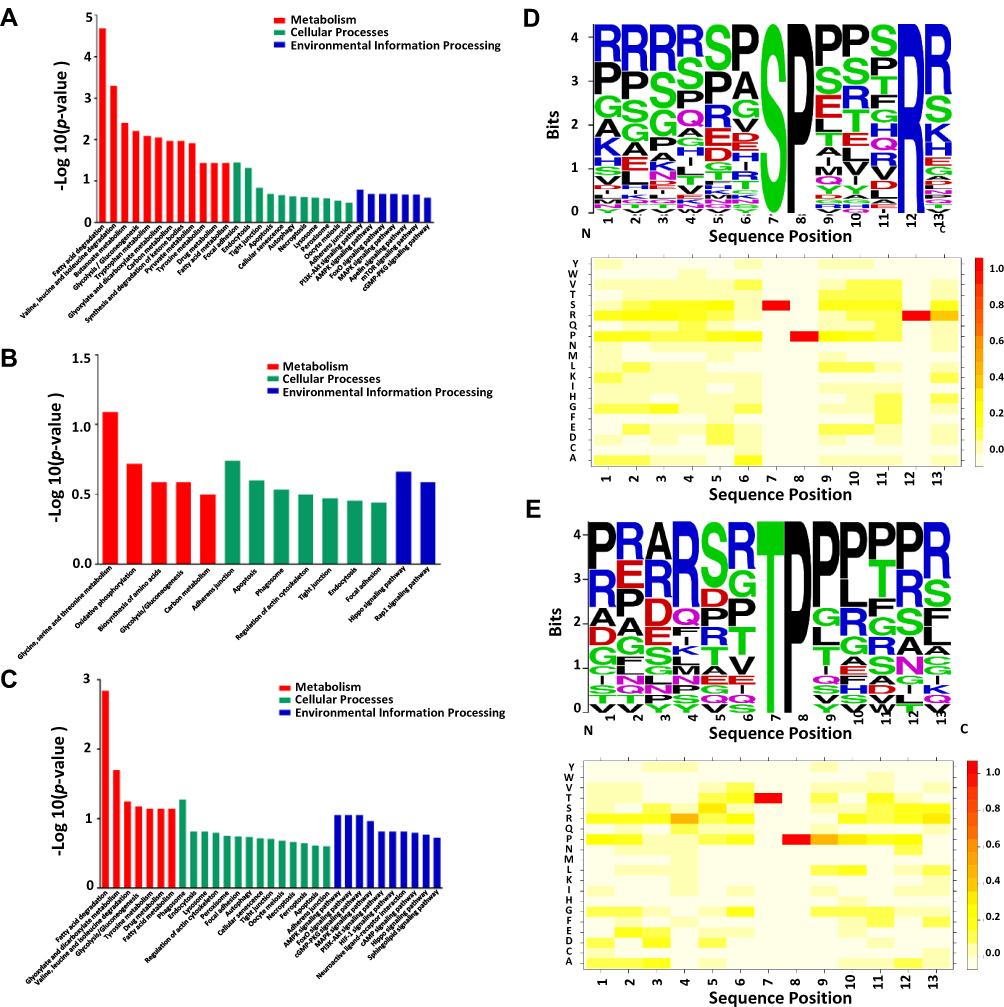 |
Figure 4 The key signaling pathways and motifs involved in the dysregulated phosphoproteins. (A–C) The key signaling pathways enriched by KEGG annotation in the HCC/Pan group, PVTT/HCC group and PVTT/Pan group, respectively. (D, E) The sequence motif analysis of p-sites in the phosphoproteome consisting of 14 residues surrounding the targeted site by Motif-X; and the heatmap for the distribution of amino acids flanking p-sites in the phosphoproteome in the HCC/Pan group and PVTT/Pan group, respectively. |
In PVTT/HCC, the enriched signaling pathway focused on Hippo signaling pathway and Rap1 signaling pathway (Figure 4B). Hippo signaling pathway plays an essential role in organ size control and tumor suppression,33 and it has been reported that misregulation of Hippo signaling pathway was involved in different types of cancers, including HCC.34 The down-regulation of the protein phosphorylation in HCC tissues suggested that the phosphorylation events might affect HCC invasion and metastasis through Hippo signaling pathway. And Rap1 signaling pathway was known for its important roles in the regulation of tumor cell migration, invasion, and metastasis.35,36 Rap1 signaling contributes to cell migration, invasion and even HCC metastasis through regulating cell adhesion, integrins and cadherins,37 modulating expression of matrix metalloproteinases (MMPs),38 controlling Rho GTPase-mediated regulation of cytoskeletal dynamics39,40 and also controlling cell proliferation.41
In PVTT/Pan, AMPK signaling pathway, FoxO signaling pathway, cGMP-PKG signaling pathway, MAPK signaling pathway, PI3K-Akt signaling pathway were mainly enriched (Figure 4C). Based on the above analysis, we knew that AMPK signaling pathway and PI3K-Akt signaling pathway both played an important role in the occurrence and development of HCC. The forkhead box O (FoxO) transcription factors were the most conserved targets of AKT, and it has been known for its important roles in metabolism regulation, cell cycle and response to stress factors.42 And cGMP-PKG signaling pathway, the inhibition of cyclic guanosine monophosphate phosphodiesterase (cGMPPDE), could increase intracellular cGMP levels to activate cGMP dependent protein kinase (PKG) and lead to the suppression of tumor cell growth and apoptosis.43,44 In addition, it has been reported that MAPK signaling pathway played an essential role in the development and aggressive behavior of tumor by enhancing tumor cell proliferation, differentiation, apoptosis and cell cycle.45,46 All of these signaling pathways demonstrated that the phosphoproteins involved in PVTT/Pan could help explain the occurrence, development and even invasion and metastasis of HCC.
The Structure Features of Phosphorylation Sites (p-Sites) in HCC
To better research the phosphorylation events in HCC with PVTT, we performed the Motif-x to analysis the structure features of phosphorylation sites (p-sites). In total, 6 serine motifs and 1 threonine motifs were significantly enriched in the HCC phosphoproteome, and the top representative motifs for phosphorylation serine (p-Ser) and phosphorylation threonine (p-Thr) are presented in Figure 4D and E. Among these, cyclin-dependent kinase (CDK) kinases, which phosphorylate many p53 sites, have strong preference for the SP motif.47 The motif [S]-D-X-E and [S]-X-D were acidic motifs, and might be recognized by casein kinase II (CK2).48,49 And motif R-X-X-[S] was a phosphorylation site consensus sequence specifically recognized by Ca2+/calmodulin-dependent protein kinases (CaMKs).50 HCC was characterized by high levels of CDKs activity, and CDK kinases play a role in regulating the cell cycle.51,52 CK2 is a ubiquitously expressed serine/threonine-kinase, which catalyzes phosphorylation of proteins involved in cell cycle regulation, cellular survival and differentiation.53 CaMKII is a ubiquitous mediator of Ca2+-linked signaling that phosphorylates a wide range of substrates to coordinate and regulate Ca22+-mediated alterations in cellular function.54 These results indicated that these kinases might play crucial roles in HCC, and the kinase-inhibitors play a role in anti-tumor activity and patient survival.
Moreover, to further analyze the relative abundances of the specific amino acids surrounding the modified site, HemI was used to visualize the heatmap for the items of PSP (7, 7) with a length of 15 amino acid residues around the p-site (seven upstream and seven downstream residues) (Figure 4D and E). It was obvious that the P at the +1 position was the amino acid with the highest frequency, and the R at the −3 position was also significantly enriched. And all of these heatmaps were well consistent with the motif analyses above.
Verification of Phosphopeptide Abundance
In this study, the phosphorylation of 4E-BP1 was elevated in HCC tissues and PVTT tissues comparing with surrounding non-cancerous tissues. It has been reported that the phosphorylation of 4E-BP1 could promote tumor angiogenesis and growth, and many phosphorylation sites on the 4E-BP1 protein have higher phosphorylation levels in cancer cells than that in adjacent tissues, which was well consistent with our experimental data. But how Thr46 phosphorylation of 4E-BP1 regulates its function was not clear. In our phosphoproteomic results, the phosphorylation of 4E-BP1 is upregulated 2.2-fold and 1.63-fold in the HCC group and PVTT group compared with Pan group. Therefore, significant upregulation of phosphorylated 4E-BP1 might increase the recurrence and metastasis during the progression of HCC pathology. The relative intensities of the reporter ion of 8-plex iTRAQ reagent of these two phosphopeptides were checked in the MS/MS spectra. As shown in Figure 5A, the expression levels of 4E-BP1Thr46 were significantly increased in the HCC group and PVTT group compared with Pan group (namely, 116>115>114 and 119>118>117). Therefore, they might be potential interesting biomarkers to predict HCC or PVTT. Here, we further verified the expression profile of these phosphorylated proteins on the tissues of HCC patients.
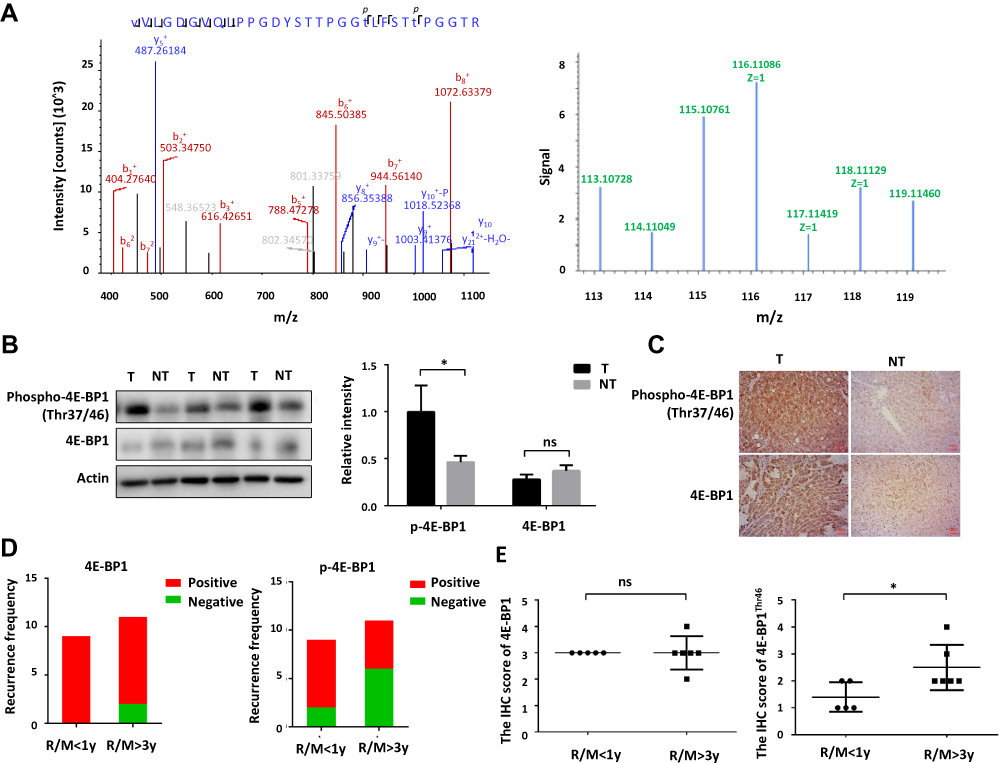 |
Figure 5 Validation of the differentially expressed phosphoproteins and phosphorylation site. (A) The relative intensity of reporter ion of the 8-plex iTRAQ reagent regarding 4E-BP1Thr46 in MS/MS spectra. (B) The level of 4E-BP1Thr37/Thr46 validated by Western-Blot are up-regulated in the HCC tissues comparing with surrounding non-cancerous tissues. (C) The level of 4E-BP1Thr37/Thr46 validated by IHC is up-regulated in the HCC tissues comparing with surrounding non-cancerous tissues. (D) The recurrence frequency of 4E-BP1Thr37/Thr46 was more positive in patients with early recurrence (recurrence within 1 year) than patients with late recurrence (recurrence after 3 years). (E) The IHC score of 4E-BP1Thr37/Thr46 was higher in patients with late recurrence (recurrence after 3 years) than patients with early recurrence (recurrence within 1 year). *p < 0.05. |
To validate the LC-MS/MS results, the protein expression level and phosphorylation level of 4E-BP1 was quantitated by Western-blot analysis using HCC patients’ tissues. As Figure 5B shown, phosphorylation level of 4E-BP1 was elevated in HCC tissue compared with the surrounding non-cancerous tissues, but the change of the protein expression level of 4E-BP1 was no statistical significance. And the further IHC analysis in a cohort of 20 HCC tissues also confirmed this conclusion, suggesting that the phosphorylation of 4E-BP1 on Thr46 might be closely related to HCC recurrence and metastasis (Figure 5C). When the IHC results of these 20 patients were carefully analyzed, we found that the phosphorylation of 4E-BP1 on Thr46 was more positive in patients with early recurrence (recurrence within 1 year) than patients with late recurrence (recurrence after 3 years) (Figure 5D). And even, the expression of 4E-BP1Thr46 was significantly higher in patients with late recurrence (recurrence after 3 years) than that in early recurrence (recurrence within 1 year) (Figure 5E). Therefore, the high phosphorylation level of 4E-BP1 on Thr46 might serve as a biomarker for the diagnosis of early recurrence of HCC.
Discussion
In order to study the phosphorylation events of PVTT in HCC, we compared the phosphorylation levels of HCC tissues, surrounding non-cancerous tissues and their PVTT tissues through a large-scale comparative phosphoproteomics approach. This was the first time for the large-scale exploration that focuses on the protein phosphorylation of PVTT in HCC.
Eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1) is a member of the translation inhibitor family, and also a substrate of the target of rapamycin signaling pathway. The translation initiation of eukaryotic cells is divided into 5ʹ hat-dependent and non-dependent and 5ʹ hats (IRES-mediated). Studies have shown that about 30% of 4E-BP1 was located in the nucleus, and its function was to restrict eIF4E in the nucleus and prevent it from entering the cytoplasm to participate in the formation of translation complex, while the function of 70% of 4E-BP1 located in the cytoplasm was still unknown.55 A recent study showed that 4E-BP1 in the cytoplasm may be involved in eukaryotic cell 5ʹ hat dependence, and the transcriptional orientation does not depend on the transformation process with the 5ʹ hat translation mode, thereby promoting tumor angiogenesis and growth.56
Since 4E-BP1 is a translational repressor, it should have a low expression level in cancer tissues, and the results in various cancer tissues and surrounding non-cancerous tissues showed that the expression of 4E-BP1 is elevated in cancer tissues.57 Phosphorylation of 4E-BP1 detaches it from the eukaryotic translation initiation factor 4E, and then promote the initiation of protein translation. It is a ubiquitous phenomenon that many phosphorylation sites on the 4E-BP1 protein have higher phosphorylation levels in cancer tissues than in adjacent tissues, which was well consistent with our experimental data.58,59 It has been reported that rapamycin-insensitive phosphorylation event at Thr46 is sufficient to prevent eIF4E: 4E-BP1 binding.19 And it also reported that 4E-BP1 phosphorylation by FRAP/mTOR on Thr-37 and Thr-46 was a priming event for subsequent phosphorylation of the carboxy-terminal serum-sensitive sites.20
Due to the important role of protein phosphorylation in cancer pathogenesis, phosphorylation site changes in tissues of different cancer types have been extensively and deeply explored. Inhibitors targeting these potential target sites have also been studied as anticancer drugs, and some indeed provide valuable methods for cancer treatment. Although there has been a lot of research in this work, however, the role of phosphorylation of imbalance in HCC mechanism needs more in-depth excavation, and new treatments will largely depend on the interpretation and study of these protein phosphorylation status change. The omics data provided in this paper provide substantial data for further understanding of protein phosphorylation and HCC development mechanisms. Of course, due to the limitations of high-throughput screening based on mass spectrometry, the sample size used in this study is still small, so more detailed data analysis and validation is still needed.
Conclusions
In this study, we have applied the iTRAQ-based quantitative phosphoproteomic approach to investigate the molecular signatures of the HCC with PVTT by quantitatively comparing the overall phosphoproteome of the primary HCC tissues, surrounding non-cancerous tissues and PVTT tissues. The results showed that different protein phosphorylation profiles in primary HCC tissues, surrounding non-cancerous tissues and PVTT tissues might reveal different underlying molecular mechanisms. Furthermore, the phosphorylation of 4E-BP1 was elevated in HCC tissues and PVTT tissues comparing with surrounding non-cancerous tissues, it might be closely related to HCC recurrence and metastasis. Therefore, the high phosphorylation level of 4E-BP1 on Thr46 might serve as a biomarker for the diagnosis of early recurrence of HCC. These results would help us better understanding of the mechanisms of PVTT, which was essential to HCC metastasis and recurrence.
Abbreviations
iTRAQ, isobaric tags for relative and absolute quantification; IARC, international agency for research on cancer; PVTT, portal vein tumor thrombus; TNM, tumor node metastasis; LC-MS/MS, liquid chromatography-tandem mass spectrometry; AFP, alpha fetoprotein; HBV, Hepatitis B virus; BCA, bicinchoninic acid; TEAB, triethylammonium bicarbonate; DTT, DL-Dithiothreitol; IAA, Iodoacetamide; ACN, acetonitrile; HCD, high energy collisional dissociation; ppm, parts per million; FDR, false discovery rate; GO, Gene Ontology.
Acknowledgments
This work is supported by the National Natural Science Foundation of China (Grant No. 81672376 and Grant No. 81702910), the Natural Science Foundation of Fujian Province (Grant No. 2017J01159 and Grant No. 2016J01417), and the Backbone Talents Training Project of Fujian Health Department (Grant No. 2019-ZQN-65).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.v68.6
2. Zhou XD, Tang ZY, Yang BH, et al. Experience of 1000 patients who underwent hepatectomy for small hepatocellular carcinoma. Cancer. 2001;91(8):1479–1486. doi:10.1002/1097-0142(20010415)91:8<1479::AID-CNCR1155>3.0.CO;2-0
3. Ng KM, Yan TD, Black D, et al. Prognostic determinants for survival after resection/ablation of a large hepatocellular carcinoma. HPB (Oxford). 2009;11(4):311–320. doi:10.1111/j.1477-2574.2009.00044.x
4. Lee YY, McKinney KQ, Ghosh S, et al. Subcellular tissue proteomics of hepatocellular carcinoma for molecular signature discovery. J Proteome Res. 2011;10(11):5070–5083. doi:10.1021/pr2005204
5. Iizuka N, Oka M, Yamada-Okabe H, et al. Oligonucleotide microarray for prediction of early intrahepatic recurrence of hepatocellular carcinoma after curative resection. Lancet. 2003;361(9361):923–929. doi:10.1016/S0140-6736(03)12775-4
6. Poon RT. Prevention of recurrence after resection of hepatocellular carcinoma: a daunting challenge. Hepatology. 2011;54(3):757–759. doi:10.1002/hep.24569
7. Portolani N, Coniglio A, Ghidoni S, et al. Early and late recurrence after liver resection for hepatocellular carcinoma: prognostic and therapeutic implications. Ann Surg. 2006;243(2):229–235. doi:10.1097/01.sla.0000197706.21803.a1
8. Chau GY, Lui WY, Wu CW. Spectrum and significance of microscopic vascular invasion in hepatocellular carcinoma. Surg Oncol Clin N Am. 2003;12(1):25–34, viii. doi:10.1016/S1055-3207(02)00077-7
9. Xu X, Wei X, Ling Q, et al. Identification of two portal vein tumor thrombosis associated proteins in hepatocellular carcinoma: protein disulfide-isomerase A6 and apolipoprotein A-I. J Gastroenterol Hepatol. 2011;26(12):1787–1794. doi:10.1111/jgh.2011.26.issue-12
10. Guo W, Xue J, Shi J, et al. Proteomics analysis of distinct portal vein tumor thrombi in hepatocellular carcinoma patients. J Proteome Res. 2010;9(8):4170–4175. doi:10.1021/pr100412w
11. Bromberg-White JL, Andersen NJ, Duesbery NS. MEK genomics in development and disease. Brief Funct Genomics. 2012;11(4):300–310. doi:10.1093/bfgp/els022
12. Otero C, Penaloza JP, Rodas PI, et al. Temporal and spatial regulation of cAMP signaling in disease: role of cyclic nucleotide phosphodiesterases. Fundam Clin Pharmacol. 2014;28(6):593–607. doi:10.1111/fcp.2014.28.issue-6
13. Chen J, Xu W, Chen Y, et al. Matrix metalloproteinase 9 facilitates hepatitis B virus replication through binding with type i interferon (IFN) receptor 1 to repress IFN/JAK/STAT signaling. J Virol. 2017;91(8). doi:10.1128/JVI.01824-16.
14. Luo Q, Wang C, Jin G, et al. LIFR functions as a metastasis suppressor in hepatocellular carcinoma by negatively regulating phosphoinositide 3-kinase/AKT pathway. Carcinogenesis. 2015;36(10):1201–1212. doi:10.1093/carcin/bgv108
15. Breunig C, Mueller BJ, Umansky L, et al. BRaf and MEK inhibitors differentially regulate cell fate and microenvironment in human hepatocellular carcinoma. Clin Cancer Res. 2014;20(9):2410–2423. doi:10.1158/1078-0432.CCR-13-1635
16. Xing X, Huang Y, Wang S, et al. Comparative analysis of primary hepatocellular carcinoma with single and multiple lesions by iTRAQ-based quantitative proteomics. J Proteomics. 2015;128:262–271. doi:10.1016/j.jprot.2015.08.007
17. Xing X, Yuan H, Sun Y, et al. ANXA2(Tyr23) and FLNA(Ser2152) phosphorylation associate with poor prognosis in hepatic carcinoma revealed by quantitative phosphoproteomics analysis. J Proteomics. 2019;200:111–122. doi:10.1016/j.jprot.2019.03.017
18. Liu H, Wang Y, Xing X, et al. Comparative proteomics of side population cells derived from human hepatocellular carcinoma cell lines with varying metastatic potentials. Oncol Lett. 2018;16(1):335–345. doi:10.3892/ol.2018.8666
19. Livingstone M, Bidinosti M. Rapamycin-insensitive mTORC1 activity controls eIF4E:4E-BP1 binding. F1000Res. 2012;1:4. doi:10.12688/f1000research
20. Gingras AC, Gygi SP, Raught B, et al. Regulation of 4E-BP1 phosphorylation: a novel two-step mechanism. Genes Dev. 1999;13(11):1422–1437. doi:10.1101/gad.13.11.1422
21. Huang X, Zeng Y, Xing X, et al. Quantitative proteomics analysis of early recurrence/metastasis of huge hepatocellular carcinoma following radical resection. Proteome Sci. 2014;12:22. doi:10.1186/1477-5956-12-22
22. Reinke H, Asher G. Circadian clock control of liver metabolic functions. Gastroenterology. 2016;150(3):574–580. doi:10.1053/j.gastro.2015.11.043
23. Fortenberry Y. The role of serpins in tumor cell migration. Biol Chem. 2015;396(3):205–213. doi:10.1515/hsz-2014-0254
24. Aparicio LA, Castosa R, Haz-Conde M, et al. Role of the microtubule-targeting drug vinflunine on cell-cell adhesions in bladder epithelial tumour cells. BMC Cancer. 2014;14:507. doi:10.1186/1471-2407-14-507
25. Saias L, Gomes A, Cazales M, et al. Cell-cell adhesion and cytoskeleton tension oppose each other in regulating tumor cell aggregation. Cancer Res. 2015;75(12):2426–2433. doi:10.1158/0008-5472.CAN-14-3534
26. Wu W, Zhang X, Qin H, et al. Modulation of tumor cell migration, invasion and cell-matrix adhesion by human monopolar spindle-one-binder 2. Oncol Rep. 2015;33(5):2495–2503. doi:10.3892/or.2015.3855
27. Vivanco I, Sawyers CL. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev Cancer. 2002;2(7):489–501. doi:10.1038/nrc839
28. Gherardi E, Birchmeier W, Birchmeier C, et al. Targeting MET in cancer: rationale and progress. Nat Rev Cancer. 2012;12(2):89–103. doi:10.1038/nrc3205
29. Fruman DA, Rommel C. PI3K and cancer: lessons, challenges and opportunities. Nat Rev Drug Discov. 2014;13(2):140–156. doi:10.1038/nrd4204
30. Fruman DA, Chiu H, Hopkins BD, et al. The PI3K pathway in human disease. Cell. 2017;170(4):605–635. doi:10.1016/j.cell.2017.07.029
31. Li W, Saud SM, Young MR, et al. Targeting AMPK for cancer prevention and treatment. Oncotarget. 2015;6(10):7365–7378. doi:10.18632/oncotarget.3629
32. Zheng L, Yang W, Wu F, et al. Prognostic significance of AMPK activation and therapeutic effects of metformin in hepatocellular carcinoma. Clin Cancer Res. 2013;19(19):5372–5380. doi:10.1158/1078-0432.CCR-13-0203
33. Dong J, Feldmann G, Huang J, et al. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell. 2007;130(6):1120–1133. doi:10.1016/j.cell.2007.07.019
34. Song H, Mak KK, Topol L, et al. Mammalian Mst1 and Mst2 kinases play essential roles in organ size control and tumor suppression. Proc Natl Acad Sci U S A. 2010;107(4):1431–1436. doi:10.1073/pnas.0911409107
35. Bailey CL, Kelly P, Casey PJ. Activation of Rap1 promotes prostate cancer metastasis. Cancer Res. 2009;69(12):4962–4968. doi:10.1158/0008-5472.CAN-08-4269
36. Yamazaki D, Kurisu S, Takenawa T. Regulation of cancer cell motility through actin reorganization. Cancer Sci. 2005;96(7):379–386. doi:10.1111/j.1349-7006.2005.00062.x
37. Kooistra MR, Dube N, Bos JL. Rap1: a key regulator in cell-cell junction formation. J Cell Sci. 2007;120(Pt 1):17–22. doi:10.1242/jcs.03306
38. Goto M, Mitra RS, Liu M, et al. Rap1 stabilizes beta-catenin and enhances beta-catenin-dependent transcription and invasion in squamous cell carcinoma of the head and neck. Clin Cancer Res. 2010;16(1):65–76. doi:10.1158/1078-0432.CCR-09-1122
39. Arthur WT, Quilliam LA, Cooper JA. Rap1 promotes cell spreading by localizing Rac guanine nucleotide exchange factors. J Cell Biol. 2004;167(1):111–122. doi:10.1083/jcb.200404068
40. Gerard A, Mertens AE, van der Kammen RA, et al. The Par polarity complex regulates Rap1- and chemokine-induced T cell polarization. J Cell Biol. 2007;176(6):863–875. doi:10.1083/jcb.200608161
41. Friedl P, Alexander S. Cancer invasion and the microenvironment: plasticity and reciprocity. Cell. 2011;147(5):992–1009. doi:10.1016/j.cell.2011.11.016
42. Tzivion G, Hay N. PI3K-AKT-FoxO axis in cancer and aging. Biochim Biophys Acta. 2011;1813(11):1925. doi:10.1016/j.bbamcr.2011.08.014
43. Piazza GA, Thompson WJ, Pamukcu R, et al. Exisulind, a novel proapoptotic drug, inhibits rat urinary bladder tumorigenesis. Cancer Res. 2001;61(10):3961–3968.
44. Thompson WJ, Piazza GA, Li H, et al. Exisulind induction of apoptosis involves guanosine 3ʹ,5ʹ-cyclic monophosphate phosphodiesterase inhibition, protein kinase G activation, and attenuated beta-catenin. Cancer Res. 2000;60(13):3338–3342.
45. Blaj C, Schmidt EM, Lamprecht S, et al. Oncogenic effects of high MAPK activity in colorectal cancer mark progenitor cells and persist irrespective of RAS mutations. Cancer Res. 2017;77(7):1763–1774. doi:10.1158/0008-5472.CAN-16-2821
46. McCubrey JA, Steelman LS, Chappell WH, et al. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim Biophys Acta. 2007;1773(8):1263–1284. doi:10.1016/j.bbamcr.2006.10.001
47. Brown NR, Lowe ED, Petri E, et al. Cyclin B and cyclin A confer different substrate recognition properties on CDK2. Cell Cycle. 2007;6(11):1350–1359. doi:10.4161/cc.6.11.4278
48. Meggio F, Pinna LA. One-thousand-and-one substrates of protein kinase CK2? FASEB J. 2003;17(3):349–368. doi:10.1096/fj.02-0473rev
49. Wang X, Bian Y, Cheng K, et al. A large-scale protein phosphorylation analysis reveals novel phosphorylation motifs and phosphoregulatory networks in Arabidopsis. J Proteomics. 2013;78:486–498. doi:10.1016/j.jprot.2012.10.018
50. White RR, Kwon YG, Taing M, et al. Definition of optimal substrate recognition motifs of Ca2+-calmodulin-dependent protein kinases IV and II reveals shared and distinctive features. J Biol Chem. 1998;273(6):3166–3172. doi:10.1074/jbc.273.6.3166
51. Swanton C. Cell-cycle targeted therapies. Lancet Oncol. 2004;5(1):27–36. doi:10.1016/S1470-2045(03)01321-4
52. Senderowicz AM. Targeting cell cycle and apoptosis for the treatment of human malignancies. Curr Opin Cell Biol. 2004;16(6):670–678. doi:10.1016/j.ceb.2004.09.014
53. Litchfield DW. Protein kinase CK2: structure, regulation and role in cellular decisions of life and death. Biochem J. 2003;369(Pt 1):1–15. doi:10.1042/bj20021469
54. Hudmon A, Schulman H. Structure-function of the multifunctional Ca2+/calmodulin-dependent protein kinase II. Biochem J. 2002;364(Pt 3):593–611. doi:10.1042/bj20020228
55. Karlsson E, Perez-Tenorio G, Amin R, et al. The mTOR effectors 4EBP1 and S6K2 are frequently coexpressed, and associated with a poor prognosis and endocrine resistance in breast cancer: a retrospective study including patients from the randomised Stockholm tamoxifen trials. Breast Cancer Res. 2013;15(5):R96. doi:10.1186/bcr3557
56. Qin X, Jiang B, Zhang Y. 4E-BP1, a multifactor regulated multifunctional protein. Cell Cycle. 2016;15(6):781–786. doi:10.1080/15384101.2016.1151581
57. Li F, Wang Q, Xiong X, et al. Expression of 4E-BP1 and phospho-4E-BP1 correlates with the prognosis of patients with clear cell renal carcinoma. Cancer Manag Res. 2018;10:1553–1563. doi:10.2147/CMAR
58. Fang Z, Lu L, Tian Z, et al. Overexpression of phosphorylated 4E-binding protein 1 predicts lymph node metastasis and poor prognosis of Chinese patients with hilar cholangiocarcinoma. Med Oncol. 2014;31(5):940. doi:10.1007/s12032-014-0940-5
59. Korkolopoulou P, Levidou G, El-Habr EA, et al. Phosphorylated 4E-binding protein 1 (p-4E-BP1): a novel prognostic marker in human astrocytomas. Histopathology. 2012;61(2):293–305. doi:10.1111/his.2012.61.issue-2
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.