Back to Journals » Infection and Drug Resistance » Volume 16
18β-Glycyrrhetinic Acid Alleviates P. multocida-Induced Vascular Endothelial Inflammation by PARP1-Mediated NF-κB and HMGB1 Signalling Suppression in PIEC Cells
Authors Lu Q ![]() , Han W, Wen D, Guo P
, Han W, Wen D, Guo P ![]() , Liu Y, Wu Z, Fu S, Ye C, Wang X, Qiu Y
, Liu Y, Wu Z, Fu S, Ye C, Wang X, Qiu Y
Received 12 April 2023
Accepted for publication 21 June 2023
Published 29 June 2023 Volume 2023:16 Pages 4201—4212
DOI https://doi.org/10.2147/IDR.S413242
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Qirong Lu,1,* Wantong Han,1,* Defeng Wen,1,* Pu Guo,1 Yu Liu,1 Zhongyuan Wu,1 Shulin Fu,1 Chun Ye,1 Xu Wang,2 Yinsheng Qiu1
1Hubei Key Laboratory of Animal Nutrition and Feed Science, School of Animal Science and Nutritional Engineering, Wuhan Polytechnic University, Wuhan, 430023, People’s Republic of China; 2National Reference Laboratory of Veterinary Drug Residues (HZAU) and Ministry of Agriculture Key Laboratory for the Detection of Veterinary Drug Residues in Foods, Huazhong Agricultural University, Wuhan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Pu Guo; Yinsheng Qiu, Wuhan Polytechnic University, Wuhan, Hubei, 430023, People’s Republic of China, Email [email protected]; [email protected]
Background: At present, the treatment and prevention of Pasteurella multocida infections in pigs mainly rely on antibiotics and vaccines, but inflammatory injury cannot be eliminated. The compound 18β-glycyrrhetinic acid (GA), a pentacyclic triterpenoid extracted from Glycyrrhiza glabra L. root (liquorice) and with a chemical structure similar to that of steroidal hormones, has become a research focus because of its anti-inflammatory, antiulcer, antimicrobial, antioxidant, immunomodulatory, hepatoprotective and neuroprotective effects, but its potential for the treatment of vascular endothelial inflammatory injury by P. multocida infections has not been evaluated. This study aimed to investigate the effects and mechanisms of GA intervention in the treatment of vascular endothelial inflammatory injury by P. multocida infections.
Materials and Methods: Putative targets of GA intervention in the treatment of vascular endothelial inflammatory injury by P. multocida infections were identified using network pharmacological screening and molecular docking simulation. The cell viability of PIEC cells was investigated via the CCK-8 assay. The mechanism of GA intervention in the treatment of vascular endothelial inflammatory injury by P. multocida infections were investigated using cell transfection and western blot.
Results: Through network pharmacological screening and molecular docking simulation, this study found that PARP1 may be a core target for GA to exert anti-inflammatory effects. Mechanistically, GA alleviates P. multocida-induced vascular endothelial inflammation by PARP1-mediated NF-κB and HMGB1 signalling suppression.
Conclusion: These findings, for the first time, demonstrate the potential therapeutic relationship among GA, PARP1 and inflammatory injury, providing a candidate drug, therapeutic targets and explanation for treating vascular endothelial inflammatory injury caused by P. multocida infection.
Keywords: network pharmacology, GA, PARP1, inflammatory injury, P. multocida infection
Introduction
Pasteurella multocida is a zoonotic pathogen with the ability to infect a wide range of hosts including humans as well as domestic, wild and companion animals.1,2 Infection with P. multocida is responsible for significant morbidity and mortality in humans and animals.1,3 The pathogen is one of the culprits causing major economic losses in the pig industry worldwide because of its persistently high prevalence: 8% prevalence with pneumonia or progressive atrophic rhinitis (PAR) between 2003 and 2007 in China,4 3.4% prevalence of 44,175 samples between 2013 and 2017 in China,5 15.6% prevalence of 33,813 isolated swine respiratory cases between 2003 and 2010 in the United States,6 15.8% prevalence between 2009 and 2010 in South Korea,7 16.8% prevalence between 2008 and 2016 in Korea.8 In clinically diseased pigs, most P. multocida strains are characterised as capsular type A and D, causing respiratory disorder in pigs, such as PAR, pneumonia, pleurisy and septicaemia.9–11 The haemorrhagic pneumonia caused by P. multocida infection suggests severe injury to the vascular endothelial cells at the site of pathogenesis and is accompanied by increased vascular endothelial permeability. At present, clinical prevention and treatment of P. multocida infection are mainly based on the application of antimicrobials and vaccines,9,12 which do not eliminate the vascular endothelial inflammation injury caused by P. multocida infection but also cause the development of drug resistance. In addition, antimicrobials, glucocorticoids or non-steroidal anti-inflammatory drugs often cause adverse effects such as bacterial endotoxin release, immune suppression or gastric ulcers. Therefore, elucidating the molecular mechanisms of vascular endothelial inflammation injury caused by P. multocida infection and developing adjuvant therapies or effective drugs to control inflammation P. multocida infection are of great scientific interest and application to reduce the harm of P. multocida infection.
Poly (ADP-ribose) polymerases-1 (PARP1) is a direct predictor of inflammatory diseases, and its central role in inflammation is the regulation of nuclear factor-kappa B (NF-κB) activation and high mobility group box 1 (HMGB1) secretion.13,14 The PARP1 itself promotes the transcription of NF-κB-dependent genes of inflammatory cytokines, such as interleukins 6 and 1β (IL-6, IL-1β) and tumour necrosis factor α (TNF-α).15 Moreover, in in vivo models, PARP1 is involved in the regulation of inflammation in bacterial infections. In a model of S. typhimurium-induced murine colitis, loss of PARP1 reduced NF-κB-mediated proinflammatory gene expression, contributing to the alleviation of intestinal inflammation.16 The PARP1 mediates bacterial endotoxin-induced HMGB1 release from macrophages, and PARP1-dependent translocation of HMGB1 from the nucleus to the cytoplasm can increase the activation of cellular inflammatory pathways.14 However, the effect of P. multocida infection on PARP1 expression and whether PARP1 alleviates P. multocida-induced vascular endothelial inflammatory injury by regulating NF-κB and HMGB1 remain unclear.
According to traditional medicine, Glycyrrhiza glabra L. root (liquorice) is one of the oldest and most widely used herbs in the world and applied to moisten the lung and relieve cough and toxicity, dispel phlegm, promote meridians and harmonise the nature of other herbal medicines.17 The compound 18β-glycyrrhetinic acid (GA), a pentacyclic triterpenoid extracted from liquorice and with a chemical structure similar to that of steroidal hormones, has become a research focus because of its anti-inflammatory, antiulcer, antimicrobial, antioxidant, immunomodulatory, hepatoprotective and neuroprotective effects.18 In a previous study, GA inhibited Staphylococcus aureus-induced RAW264.7 cell inflammation by inhibiting the activation of NF-κB and the expression of HMGB1;19 in addition, it has a protective effect on Helicobacter pylori-infected gastric mucosa through alleviating the expression levels of inflammation-related cytokines (IL-1β, TNF-α).20 However, the protective role of GA against P. multocida-induced vascular endothelial inflammatory injury via PARP1 is still largely unclear.
In this study, drug target prediction and experimental validation were used to explore the mechanism by which GA alleviates vascular endothelial inflammatory injury caused by P. multocida infection through regulating the PARP1-NF-κB/HMGB1 pathway, with the aims to prove that GA has potential to serve as a novel antibiotic alternative and to provide a basis for clinically alleviating vascular endothelial inflammatory injury in P. multocida-infected piglets.
Materials and Method
Reagent and Chemicals
The GA (CAS NO. 471-53-4; Catalog # HY-N0180) was purchased from MedChemExpress, USA.
Network Pharmacological Analysis
The potential protein target of GA was identified by the drug target database, such as PharmMapper,21 SwissTargetPrediction22 and TCMSP.23 The targets of vascular endothelial injury were obtained from the PubMed database (https://pubmed.ncbi.nlm.nih.gov) and the GeneCards database (https://www.genecards.org). The Veen software (www.bioinformatics.com.cn) was employed to select the common target of the potential protein target of GA and the targets of vascular endothelial injury in pigs. The core target was selected by the protein-protein interaction (PPI) network of the String database (https://cn.string-db.org), using the Cytoscape 3.7.2 software.24 Subsequently, GO enrichment analysis and the Kyoto encyclopedia of genes and genomes (KEGG) pathway enrichment analysis were used to analyse the functions and the involved signalling pathways of core genes in the database for annotation, visualisation and integrated discovery (DAVID) (https://david.ncifcrf.gov), with the species limited to “Sus scrofa”. Finally, the results of GO and KEGG analysis were visualized using a bioinformatics tool (www.bioinformatics.com.cn).
Molecular Docking
The SWISS-MODEL (https://swissmodel.expasy.org) was employed to construct the protein structure homology-modelling of PARP1.25 Subsequently, molecular docking between PARP1 and GA was conducted using the software SYBYL-X 2.0. After molecular docking, the results were visualised by the PyMOL software.26
Bacterial Strains and Cell Culture
Pasteurella multocida strain HB03 (serogroups/genotypes A) was kindly provided by Professor Bin Wu and Dr. Zhong Peng (Huazhong Agricultural University, Wuhan). Strain HB03 was cultured in trypticase soy broth medium (Hopebio, China) supplemented with 5% bovine serum (NEWZERUM Ltd, New Zealand, Catalog # NCS-500) at 37°C for 8–12 h.
Porcine vascular endothelial cell line (PIEC) cells were kindly provided by Professor Hongkui Wei and Professor Menghong Dai at Huazhong Agricultural University (Wuhan, China). For culturing PIEC cells, RPMI medium modified (Cytiva, USA, Catalog # SH30809.01) with 1% penicillin-streptomycin solution (Gibco, USA, Catalog # 15070063) and 10% foetal bovine serum (PAN, Germany, Catalog # ST30-3302) was used. The cultures were cultured in a humidified incubator set at 37°C and 5% CO2. After inoculating the PIEC cells in the cell culture plate for approximately 12 h, the cultures were preincubated with GA for 2 h and then co-treated with strain HB03 (MOI = 50) for another 10 h. All experiments were performed at least in triplicate on three separate occasions.
Cell Viability Assay
The cell viability of PIEC cells was investigated via the CCK-8 assay. For this, PIEC monolayer cells in each well of a 96-well plate were incubated with a series of GA (5–160 μg/mL) for 24 h, followed by the addition of 10 μL CCK-8 solution (Vazyme, China, Catalog # A311-02) into each well of the plate for 2 h. The absorbance of formazan was measured at 450 nm using a microplate reader (Spectra MIX i3x, Molecular Devices, China).
Cell Transfection
The PIEC cells were transiently transfected using the lipo8000™ transfection reagent (Beyotime, China, Catalog # C0533). For the PARP1 overexpression plasmids, PIEC monolayer cells in each well of a 6-well plate were transfected with 125 μL Opti-MEM (Gibco, USA, Catalog # 31985070) diluted in 3.75 μg of DNA (pcDNA3.0-PARP1 or pcDNA3.0) and 3.75 μL of lipo8000™ transfection reagent. For the RNAi assay, PIEC monolayer cells in each well of a 6-well plate were transfected with 125 μL Opti-MEM diluted with 7.5 μL of siRNA (siRNA of PARP1 or negative control siRNA (GenePharma, China) and 5 μL of lipo8000™ transfection reagent.
Western Blot
Challenged PIEC cells were lysed in RIPA buffer (Beyotime, China, Catalog # P0013B) with a protease and phosphatase inhibitor cocktail (Beyotime, China, Catalog # P1045), sonicated and centrifuged at 10,000 × g for 10 min at 4°C. The protein concentration of the supernatant was measured by the enhanced BCA protein assay kit (Beyotime, China, Catalog # P1010). Briefly, 20 µg of protein from each sample was separated via 10% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (Sangon Biotech, China, Catalog # C671102-0125) and then transferred to polyvinylidene difluoride membranes (Millipore, USA, Catalog # SE4M039I09). After being blocked with 5% BSA in tris-buffered saline with Tween 20 for 2 h at room temperature, the blots were incubated overnight with the primary antibodies-anti-PARP1 polyclonal antibody (Proteintech Group, China, Catalog # 13371-1-AP, 1:1000 dilution) anti-β-actin polyclonal antibody (Proteintech Group, China, Catalog # 20536-1-AP, 1:1000 dilution), anti-phospho-NF-kappa-B transcription factor p65 monoclonal antibody (p-NF-κB p65) (Thermo Fisher Scientific, USA, Catalog # MA5-15160, 1:1000 dilution), anti-IL-1β polyclonal antibody (Abclonal, China, Catalog # A1112, 1:1000 dilution), anti-IL-18 polyclonal antibody (Abclonal, China, Catalog # A16737, 1:1000 dilution), anti-HMGB1 polyclonal antibody (Abclonal, China, Catalog # A2553, 1:1000 dilution) and anti-NF-κB p65 polyclonal antibody (Abclonal, China, Catalog # A11204, 1:1000 dilution). After washing, the blots were incubated with secondary anti-IgG antibody (Abclonal, China, Catalog # A11204, 1:5000 dilution) and then visualised with enhanced chemiluminescence reagents (Abclonal, China, Catalog # RM00021) by the FluorChem E (ProteinSimple, China). All western blots were densitometrically quantified using the ImageJ software (ImageJ, NIH).
Statistical Analysis
Statistical analysis was performed using SPSS 18.0 for Windows. All results were presented as mean ± standard deviation. Results were statistically analysed by one-way ANOVA followed by Duncan’s post hoc analysis. Statistical significance was set at p < 0.05, and a value of p < 0.01 was considered to be strongly significant.
Results
Identification of the Core Targets from Network Pharmacological Analysis
To explore the potential core targets of GA for attenuating P. multocida-mediated vascular endothelial inflammatory injury, we explored the core targets from the target database. Target database analysis yielded 216 and 749 targets for GA and vascular endothelial injury, respectively (Figure 1A and B, Supplementary Table 1). The 69 common targets of GA and vascular endothelial injury targets were analysed by the Venn diagram (Figure 1C, Supplementary Table 1). To further reveal the core targets of GA and vascular endothelial injury, all targets were mapped into the STRING database and conducted PPI network. As shown in Figure 2A and B, blue nodes represent vascular endothelial injury targets, orange nodes represent GA targets, and green nodes represent common targets. The core targets of GA and vascular endothelial injury were further selected, including PARP1, HMGB1, IL-1β, IL-18, RelA (NF-κB p65), PTPN11 and MAPK1 (Figure 2C). In addition, this study also compared the GA docking scores with niraparib (FDA-approved PARP1 inhibitor) docking scores, the docking scores of niraparib and GA with PARP1 are 5.9030 and 5.4304, respectively, indicating that both niraparib and GA have the potential to bind with PARP1 (Figure 3 and Table 1). Collectively, these data suggest that GA alleviated P. multocida-induced vascular endothelial inflammatory injury, possibly by altering the expression of core targets such as PARP1, HMGB1, IL-1β, IL-18 and NF-κB p65.
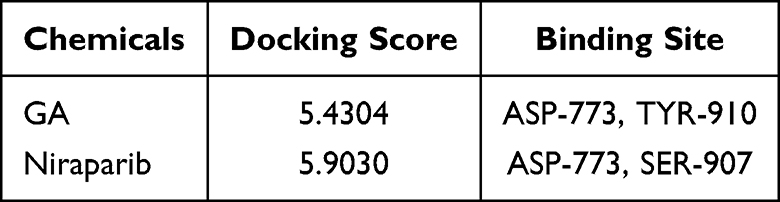 |
Table 1 The Docking Value of 18β-Glycyrrhetinic Acid (GA) and Niraparib with PARP1 |
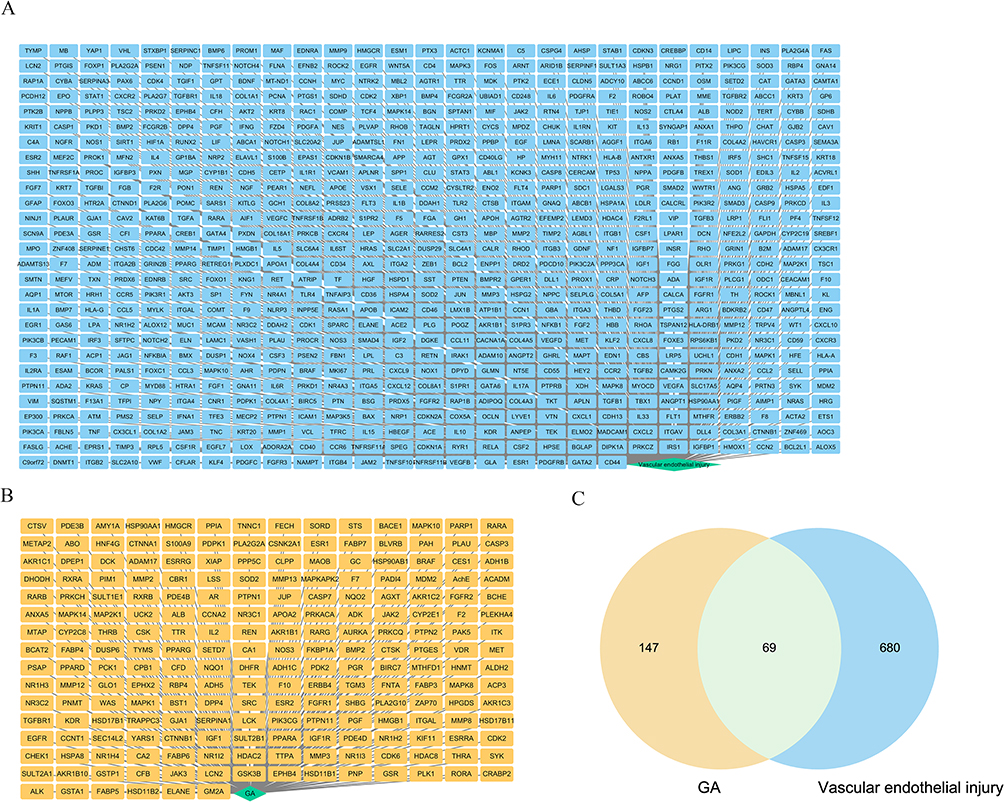 |
Figure 1 Target databases were applied to summarise the targets of 18β-glycyrrhetinic acid (GA) and vascular endothelial injury. (A) Summary of vascular endothelial injury targets using PubMed database and GeneCards databases; (B) Summary of GA using the drug target database; (C) The common vascular endothelial injury targets and GA targets were summarised by the Veen software. |
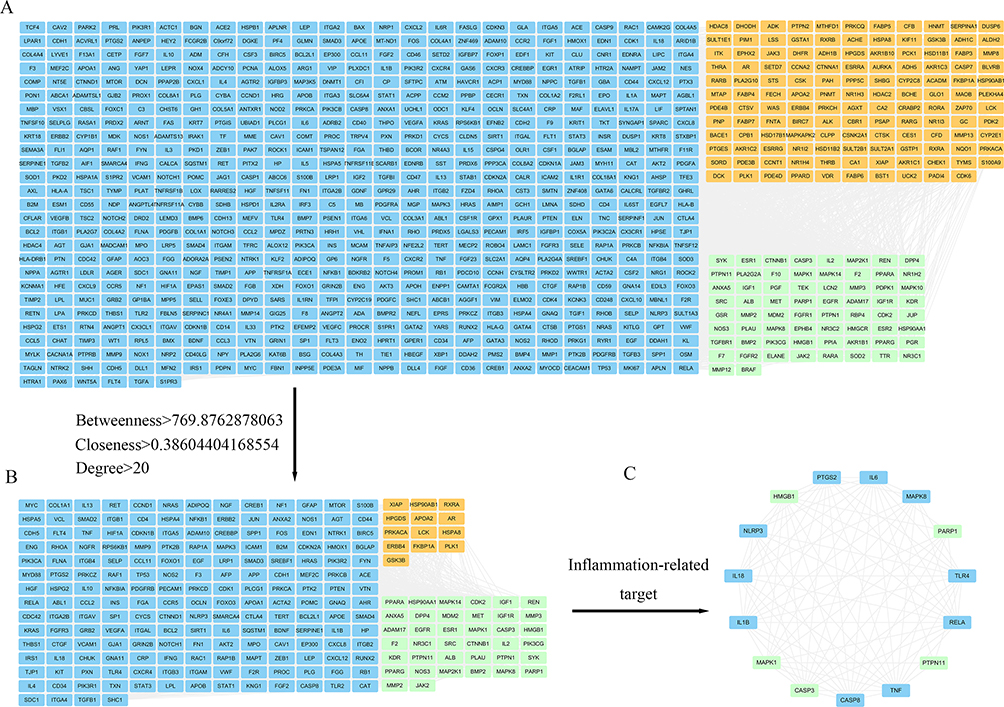 |
Figure 2 The protein-protein interaction (PPI) network and Cytoscape analyses were applied to screen the core targets of 18β-glycyrrhetinic acid (GA) in alleviating vascular endothelial injury. (A) PPI network diagram after String database screening; (B) Diagram of core targets for further screening based on the Cytoscape database. Orange nodes represent GA targets, blue nodes represent vascular endothelial injury targets, and green nodes represent common targets of vascular endothelial injury and GA. (C) Screening of core targets related to inflammation. |
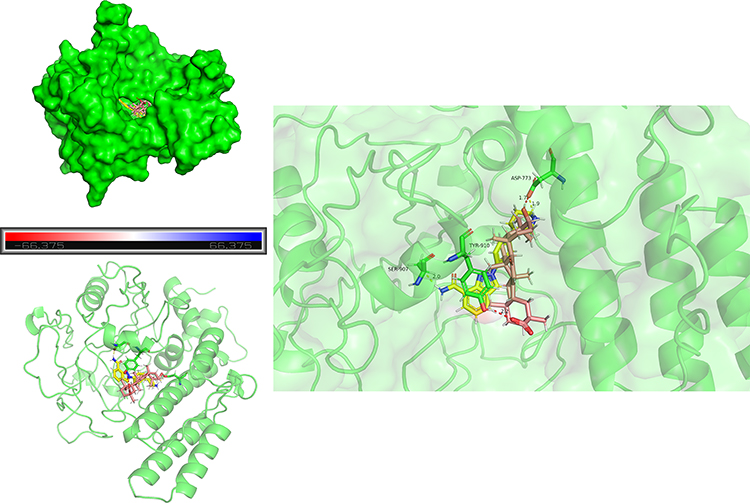 |
Figure 3 Molecular docking was applied to model the potential of 18β-glycyrrhetinic acid (GA) and niraparib to bind PARP1. Brown stick molecules represent GA, yellow stick molecules represent niraparib and green stick molecules represent amino acid residues of PARP1. |
Core Targets Participate in Inflammatory Disorders
To understand the putative functions of the core targets, they were mapped to the GO and KEGG databases for analyses. Among the enriched biological processes, some processes related to the regulation of inflammatory injury were mainly enriched, such as positive regulation of interleukin-8 production, cellular response to lipopolysaccharide (LPS), positive regulation of IκBα kinase/NF-κB signalling and inflammatory response (Figure 4A). According to the KEGG analysis, core targets involved in the top 40 signalling pathways were mainly involved in the PI3K-AKT signalling pathway, the MAPK signalling pathway, coronavirus disease-COVID-19 and the JAK-STAT signalling pathway (Figure 4B). Based on the results of the comprehensive GO and KEGG analysis, core targets of GA and vascular endothelial injury may participate in the inflammatory disorders of vascular endothelial cells.
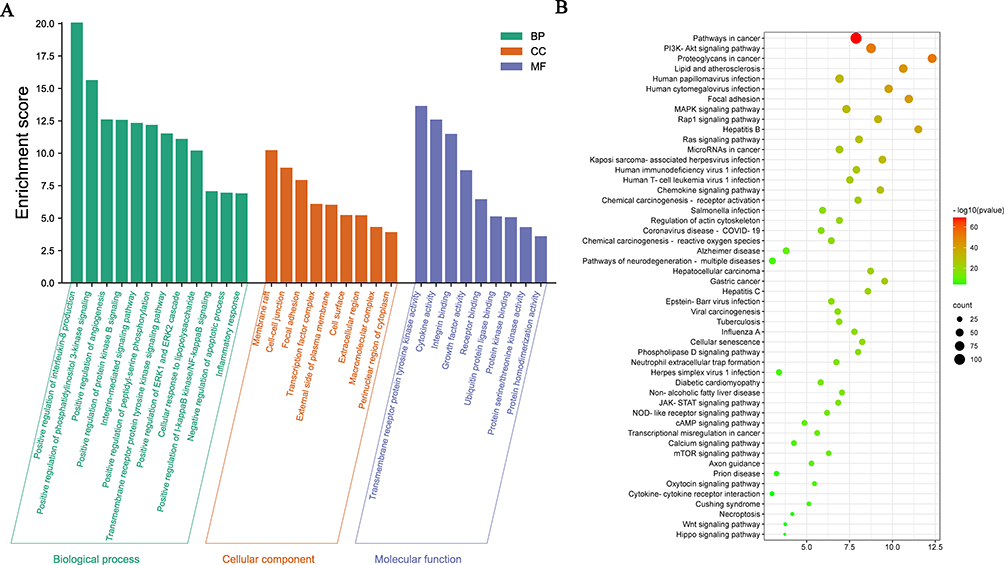 |
Figure 4 GO enrichment analysis and the Kyoto encyclopedia of genes and genomes (KEGG) analysis of the core targets of 18β-glycyrrhetinic acid (GA) in alleviating vascular endothelial inflammatory injury using the database for annotation, visualisation and integrated discovery (DAVID). (A) GO analysis of core targets; (B) KEGG analysis of core targets. |
Dose-Effect of GA on the Cytotoxicity of PIEC Cells
The CCK-8 assay was used to evaluate the dose-effect of GA on the cytotoxicity of PIEC cells. As presented in Figure 5A, below 40 μg/mL, GA was almost non-cytotoxic, with a cell viability > 80%. However, at concentrations of 80–160 μg/mL, GA exhibited significant cytotoxicity to PIEC cells (p<0.01). Consequently, 10, 20 and 40 40 μg/mL were used as the working concentrations of GA for the whole experiment.
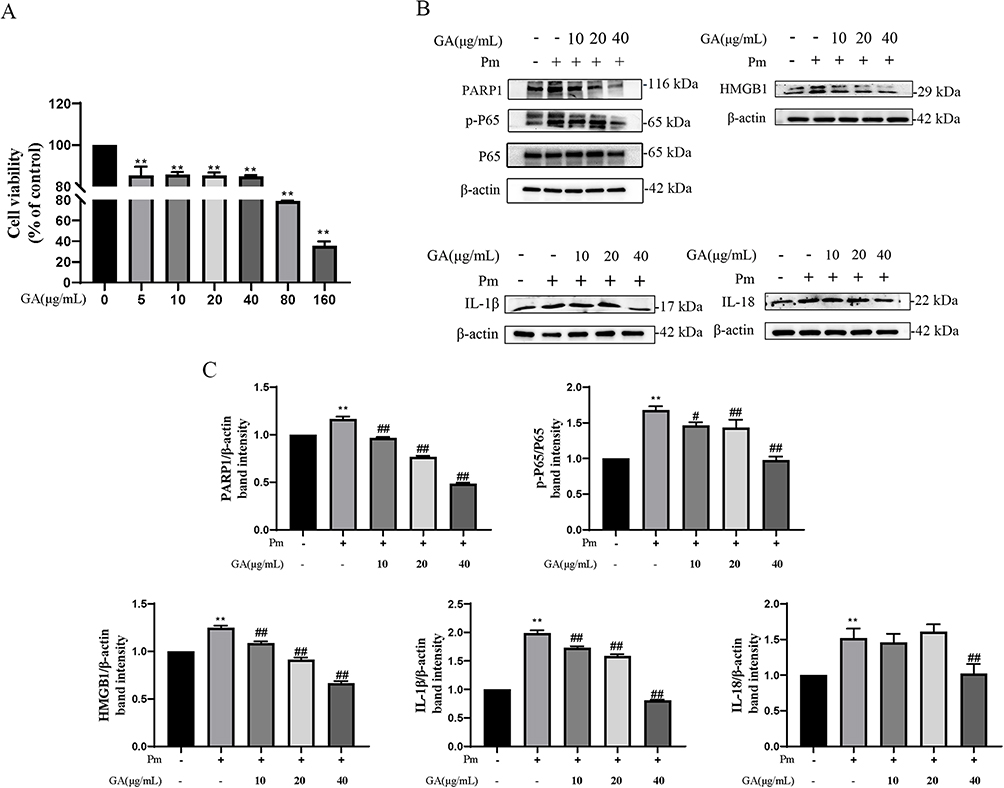 |
Figure 5 18β-glycyrrhetinic acid (GA) attenuates vascular endothelial inflammatory injury induced by P. multocida infection in PIEC cells. (A) Cytotoxicity of GA on PIEC cells using the CCK-8 assay; (B) GA reduced the expression of inflammatory proteins induced by P. multocida infection in PIEC cells; (C) Protein density quantified using the ImageJ software. **represents the control group vs P. multocida infection group, p < 0.01; #And ##Represent other groups vs P. multocida infection group, p < 0.05 and p < 0.01, respectively. |
GA Attenuates P. multocida-Induced Vascular Endothelial Inflammatory Injury
To evaluate whether GA can attenuate endothelial inflammatory injury, this study modelled vascular endothelial inflammatory injury using PIEC cells induced by P. multocida. The minimum inhibitory concentration (MIC) of GA against P. multocida was 1024 μg/mL. In addition, the western blot results showed that the expression levels of PARP1, p-NF-κB p65, HMGB1, IL-1β and IL-18 were upregulated after P. multocida infection (p<0.01), with a significant dose-dependent reversal of P. multocida-induced inflammatory protein (PARP1, p-NF-κB p65, HMGB1 and IL-1β) expression by 10–40 μg/mL of GA (p<0.01) (Figure 5B and C); 10 μg/mL (p=0.950) and 20 μg/mL (p=0.869) concentrations of GA did not produce a significant effect on P. multocida -induced expression of IL-18. However, the concentration of 40 μg/mL of GA significantly reduced the expression of IL-18. These findings indicate that P. multocida infection can cause an abnormal increase in inflammatory markers of vascular endothelial cells, whereas GA can effectively alleviate P. multocida infection-induced vascular endothelial inflammatory injury.
GA Attenuates P. multocida-Induced Vascular Endothelial Inflammatory Injury by Inhibiting the PARP1 Pathway
To gain insight into how GA alleviates the cellular inflammatory damage induced by P. multocida infection, a comprehensive analysis was performed in combination with PARP1 overexpression and interference assays, bacterial infection and GA treatment. As shown in Figure 6A and B, overexpressed PARP1 significantly promoted the expression of PARP1, p-NF-κB p65, HMGB1, IL-1β and IL-18 (p<0.01). However, GA was able to alleviate inflammatory protein elevation induced by PARP1 overexpression in a concentration-dependent manner. Knockdown of PARP1 also down-regulated the expression of inflammatory proteins such as PARP1, p-NF-κB p65, HMGB1, IL-1β and IL-18 (p<0.01) and inhibited the increase in inflammatory proteins induced by P. multocida (Figure 7A and B). The results showed that GA alleviates P. multocida-induced vascular endothelial inflammatory injury through the PARP1-HMGB1 or the PARP1- NF-κB p65 signalling pathway.
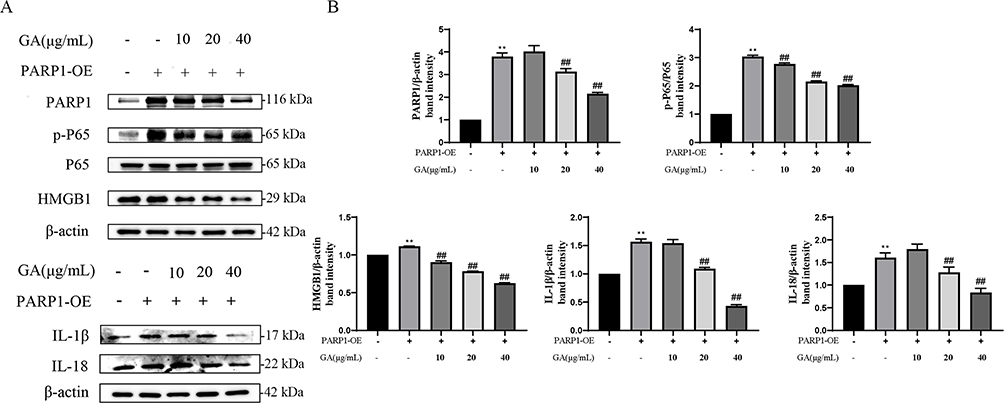 |
Figure 6 18β-glycyrrhetinic acid (GA) alleviates overexpressed PARP1-induced vascular endothelial inflammatory injury in PIEC cells. (A) Pre-treatment with GA could alleviate the overexpressed PARP1-induced expression of inflammation-related protein, such as PARP1, HMGB1, p-P65, IL-18 and IL-1β; (B) Protein density quantified using the ImageJ software. PARP1-OE represents overexpressed PARP1. **Represents the control group vs the PARP1-OE group, p < 0.01; ##Represents other groups vs PARP1-OE group, p < 0.01. |
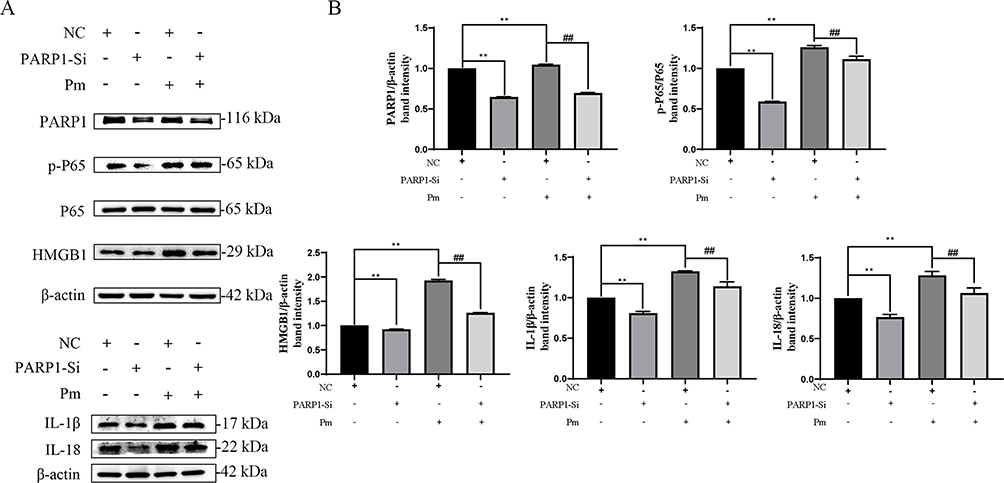 |
Figure 7 Knockdown of PARP1 decreased vascular endothelial inflammatory injury induced by P. multocida infection in PIEC cells. (A) Knockdown of PARP1 decreased P. multocida infection-induced expression of inflammation-related protein, such as PARP1, HMGB1, p-P65, IL-18 and IL-1β; (B) Protein density quantified using the ImageJ software. PARP1-Si represents Knockdown of PARP1. NC represents negative control siRNA. ##Represents NC group+ P. multocida infection group, p < 0.01; **Represents other groups vs NC group, p < 0.01. |
Discussion
As one of the main pathogenic bacteria causing respiratory diseases in humans and animals, the study of the interaction between P. multocida and vascular endothelial cells plays an important role in exploring the pathogenesis of hemorrhagic pneumonia. The clinical treatment of P. multocida infections mainly occurs via the application of antimicrobials. However, antimicrobials do not solve the problem of inflammatory injury of vascular endothelial cells caused after P. multocida infection, and the massive and illegal use of antimicrobials is often accompanied by the occurrence of antimicrobial resistance.27 Therefore, it is of great significance to explore the mechanism of vascular endothelial inflammatory injury induced by P. multocida infection and to search for new, effective drugs for adjuvant treatment or the control of inflammation in the disease to reduce the harm of P. multocida infection. In the present study, we used PIEC cells as a model and found that GA alleviates P. multocida-induced vascular endothelial inflammation by PARP1-mediated NF-κB and HMGB1 signalling suppression (Figure 8), suggesting that GA and PARP1 are potential alternatives to antibiotics and candidate therapeutic targets for the adjuvant treatment of P. multocida infection, respectively.
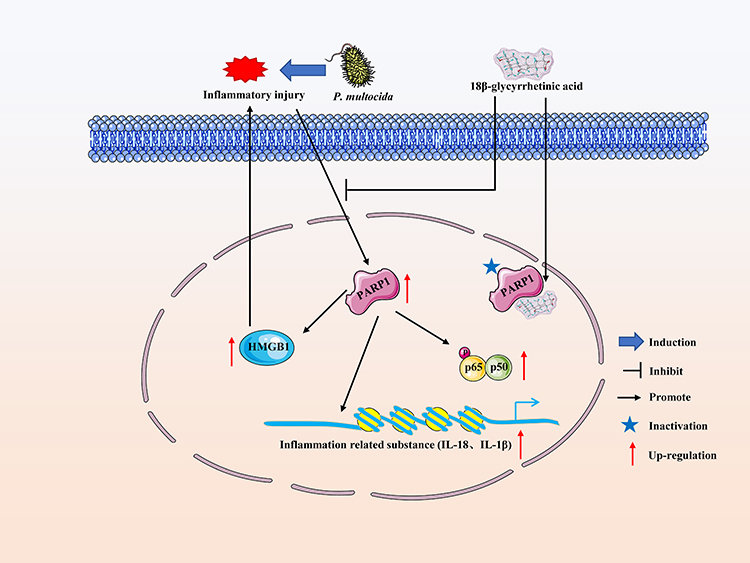 |
Figure 8 18β-glycyrrhetinic acid (GA) alleviates P. multocida-induced vascular endothelial inflammation by PARP1-mediated NF-κB and HMGB1 signalling suppression in PIEC cells. P. multocida-infected PIEC cells could significantly active the PARP1-mediated NF-κB and HMGB1 signalling pathways and induce the PARP1-mediated expression of inflammatory-related substances, such as IL-1β and IL-18. On the one hand, GA may inhibit the activity of PARP1 by targeting it, on the other hand, it may inhibit the NF-κB and HMGB1 signalling pathways by reducing the expression of PARP1, thus inhibiting P. multocida infection-induced vascular endothelial inflammatory injury in PIEC cells. |
Network pharmacology is a new discipline to study the mechanism of drug-target-disease interactions in the context of larger biological networks.28 To explore the possible target of GA in host cells that are responsible for the vascular endothelial inflammatory injury induced by P. multocida infection, network pharmacology analysis was conducted. Some inflammation-related core targets of GA and vascular endothelial injury were enriched by PPI network and Cytoscape analysis,29,30 such as PARP1, HMGB1, IL-1β, IL-18, NF-κB p65, indicating that inflammatory pathways play an important role in GA alleviating P. multocida-induced vascular endothelial inflammatory injury. Moreover, GO analysis showed that core targets are mainly enriched in biological functions related to the regulation of inflammatory injury, such as positive regulation of interleukin-8 production, cellular response to LPS, positive regulation of IκBα kinase/NF-κB signalling and inflammatory response. Based on the results of the KEGG enrichment analysis, GA involves multiple signalling pathways in the treatment of P. multocida infection, mainly including the PI3K-AKT signalling pathway, the MAPK signalling pathway, coronavirus disease-COVID-19 and the JAK-STAT signalling pathway. In addition, studies have shown that the NF-κB/MAPK signalling pathway is involved in the regulation of LPS-induced inflammatory responses in IPEC-J2 cells.31 According to another study, HMGB1 can enhance the inflammatory response by activating the p38 MAPK signalling pathway.32,33 Meanwhile, PARP1 could serve as a regulator of inflammatory diseases through promoting NF-κB activation and HMGB1 secretion.13,14 The network pharmacology results shown herein revealed that the activation of PARP1 might contribute to the activation of NF-κB and HMGB1, which may finally be beneficial for vascular endothelial inflammatory injury after P. multocida infection.
The GA can be an anti-inflammatory drug candidate for the treatment of inflammatory responses evoked by various external stimuli. In a previous study, it inhibited Staphylococcus aureus-induced RAW264.7 cell inflammation by inhibiting the activation of NF-κB and the expression of HMGB119 and showed a protective effect on Helicobacter pylori-infected gastric mucosa through alleviating the expression levels of inflammation-related cytokines (IL-1β, TNF-α).20 Further, GA suppressed allergic airway inflammation through NF-κB and Nrf2/HO-1 signalling pathways in an ovalbumin-induced asthma mouse model.34 In LPS- or TNF-α-induced inflammatory cell models and collagen-induced arthritis animal models, GA exerts its anti-inflammatory activity through the MAPK/NF-κB signalling pathway.35 The above studies demonstrate that GA presents great anti-inflammatory activity in vivo or in vitro by inhibiting pathways such as NF-κB and HMGB1. This study found that GA could inhibit the NF-κB and HMGB1 pathways to alleviate the vascular endothelial inflammatory injury induced by P. multocida infection. In addition, in this study, GA potentially targeted PARP1 by network pharmacology and molecular docking technology, which in turn exerted the inhibition of NF-κB and HMGB1 pathway activation.
The PARP1 is involved in the regulation of inflammation in bacterial infections. In LPS-induced mouse septic shock and S. typhimurium-induced mouse colitis models, the absence of PARP1 decreased NF-κB-mediated inflammatory disorders.16,36 Moreover, PARP1 mediates LPS-induced HMGB1 secretion from macrophages, and PARP1 activity-dependent HMGB1 translocation from the nucleus to the cytoplasm can promote the activation of cellular inflammatory pathways.14 However, loss of PARP1 activity impedes the cytosolic translocation of HMGB1, which can prevent the HMGB1-mediated TLR4- NF-κB signalling pathway to attenuate LPS-induced myocardial inflammation.37 In the present study, either PARP1 overexpression or PARP1 deletion demonstrated the ability of PARP1 to promote the expression of NF-κB and HMGB1, which in turn were involved in the progression of vascular inflammatory injury induced by P. multocida infection. In addition, the molecular docking score is greater than 4,38 implying that PARP1 might be a key target of GA. And GA at lower antimicrobial concentrations (10 ~ 40 μg/mL) on the inflammatory response of endothelial cells induced by P. multocida infection. Thus, we speculate that GA may play a role in alleviating vascular endothelial inflammatory injury induced by P. multocida infection by targeting PARP1 and inhibiting PARP1 activity. The above studies demonstrate that GA and PARP1 could serve as a novel antibiotic alternative and therapeutic target for the prevention and treatment of vascular endothelial inflammatory injury induced by P. multocida infection, respectively.
This study has, however, some limitations. First, some unconfirmed and unrecorded targets of the target database may not be enriched in the present study, implying that the target database of network pharmacology needs to be further improved. Second, the molecular docking results showed that PARP1 may have the potential to bind with GA, but its specific binding situation awaits further investigation. Third, the results of the present study showed that PARP1 could regulate the protein expression of NF-κB and HMGB1, but whether GA inhibits inflammation by targeting PARP1 to regulate the nuclear translocation of NF-κB and HMGB1 during P. multocida infection awaits further exploration. Fourth, pharmacodynamic and animal therapeutic trials are needed to further verify the potential of GA as an alternative to antibiotics in the treatment of vascular endothelial inflammatory injury caused by P. multocida infection.
Conclusion
Collectively, through the mining of network pharmacology studies and the verification of in vitro experiments, this study proves the powerful potential of network pharmacology in disease and drug target screening and that PARP1 is a potential target for GA for the treatment of vascular endothelial inflammatory injury caused by P. multocida infection. Further, PARP1-mediated NF-κB and HMGB1 signalling suppression may be a critical signalling pathway for treating vascular endothelial inflammatory injury caused by P. multocida infection.
Ethics Statement
Part of the target data for this manuscript was obtained from a public database, such as GeneCards. The ethics committee of Wuhan Polytechnic University has reviewed the data in this manuscript, which does not violate relevant ethical issues and agrees to publish it.
Acknowledgments
We sincerely acknowledge Professor Hongkui Wei and Professor Menghong Dai at Huazhong Agricultural University (Wuhan, China) for the gift of PIEC cells. We also thank the Professor Bin Wu and Dr. Zhong Peng at Huazhong Agricultural University (Wuhan, China) for the gift of P. multocida strain HB03.
Funding
This work was supported by National Natural Science Foundation of China (NSFC) (32202867) and Key Research and Development Plan of Hubei Province, China (2022BBA0055).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Smith E, Miller E, Aguayo JM, et al. Genomic diversity and molecular epidemiology of Pasteurella multocida. PLoS One. 2021;16(4):e0249138. doi:10.1371/journal.pone.0249138
2. Peng Z, Wang X, Zhou R, et al. Pasteurella multocida: genotypes and genomics. Microbiol Mol Biol Rev. 2019;83(4):e00014–e00019. doi:10.1128/mmbr.00014-19
3. Giordano A, Dincman T, Clyburn BE, Steed LL, Rockey DC. Clinical features and outcomes of Pasteurella multocida infection. Medicine. 2015;94(36):e1285. doi:10.1097/md.0000000000001285
4. Tang X, Zhao Z, Hu J, et al. Isolation, antimicrobial resistance, and virulence genes of Pasteurella multocida strains from swine in China. J Clin Microbiol. 2009;47(4):951–958. doi:10.1128/jcm.02029-08
5. Zhang B, Ku X, Yu X, et al. Prevalence and antimicrobial susceptibilities of bacterial pathogens in Chinese pig farms from 2013 to 2017. Sci Rep. 2019;9(1):9908. doi:10.1038/s41598-019-45482-8
6. Opriessnig T, Giménez-Lirola LG, Halbur PG. Polymicrobial respiratory disease in pigs. Anim Health Res Rev. 2011;12(2):133–148. doi:10.1017/s1466252311000120
7. Lee KE, Jeoung H-Y, Lee J-Y, et al. Phenotypic characterization and random amplified polymorphic DNA (RAPD) analysis of Pasteurella multocida isolated from Korean Pigs. J Vet Med Sci. 2012;74(5):567–573. doi:10.1292/jvms.11-0418
8. Kim J, Kim JW, Oh S-I, et al. Characterisation of Pasteurella multocida isolates from pigs with pneumonia in Korea. BMC Vet Res. 2019;15(1):119. doi:10.1186/s12917-019-1861-5
9. Oh YH, Moon DC, Lee YJ, Hyun BH, Lim SK. Genetic and phenotypic characterization of tetracycline-resistant Pasteurella multocida isolated from pigs. Vet Microbiol. 2019;233:159–163. doi:10.1016/j.vetmic.2019.05.001
10. Peng Z, Wang H, Liang W, et al. A capsule/lipopolysaccharide/MLST genotype D/L6/ST11 of Pasteurella multocida is likely to be strongly associated with swine respiratory disease in China. Arch Microbiol. 2018;200(1):107–118. doi:10.1007/s00203-017-1421-y
11. Oliveira Filho JX, Morés MAZ, Rebellato R, et al. Pathogenic variability among Pasteurella multocida type A isolates from Brazilian pig farms. BMC Vet Res. 2018;14(1):244. doi:10.1186/s12917-018-1565-2
12. Mostaan S, Ghasemzadeh A, Sardari S, et al. Pasteurella multocida vaccine candidates: a systematic review. Avicenna J Med Biotechnol. 2020;12(3):140–147.
13. Rosado MM, Bennici E, Novelli F, Pioli C. Beyond DNA repair, the immunological role of PARP-1 and its siblings. Immunology. 2013;139(4):428–437. doi:10.1111/imm.12099
14. Wang Y, Pleasure D, Deng W, Guo F. Therapeutic potentials of poly (ADP-Ribose) polymerase 1 (PARP1) inhibition in multiple sclerosis and animal models: concept revisiting. Adv Sci. 2022;9(5):e2102853. doi:10.1002/advs.202102853
15. Maluchenko NV, Feofanov AV, Studitsky VM. PARP-1-associated pathological processes: inhibition by natural polyphenols. Int J Mol Sci. 2021;22(21):11441. doi:10.3390/ijms222111441
16. Altmeyer M, Barthel M, Eberhard M, et al. Absence of poly(ADP-ribose) polymerase 1 delays the onset of Salmonella enterica serovar typhimurium-induced gut inflammation. Infect Immun. 2010;78(8):3420–3431. doi:10.1128/iai.00211-10
17. Ding Y, Brand E, Wang W, Zhao Z. Licorice: resources, applications in ancient and modern times. J Ethnopharmacol. 2022;298:115594. doi:10.1016/j.jep.2022.115594
18. Kowalska A, Kalinowska-Lis U. 18β-Glycyrrhetinic acid: its core biological properties and dermatological applications. Int J Cosmet Sci. 2019;41(4):325–331. doi:10.1111/ics.12548
19. Rao C, Hong Z, Yao Y, Zheng G, Wang S. 18-beta-glycyrrhetinic acid protects against Staphylococcus aureus infection by regulating the NF-κB pathway. Indian J Pharm Educ. 2019;53(2s):s151–s158. doi:10.5530/ijper.53.2s.60
20. Cao D, Jiang J, You L, et al. The protective effects of 18 β-glycyrrhetinic acid on Helicobacter pylori -infected gastric mucosa in Mongolian gerbils. Biomed Res Int. 2016;2016:4943793. doi:10.1155/2016/4943793
21. Liu X, Ouyang S, Yu B, et al. PharmMapper server: a web server for potential drug target identification using pharmacophore mapping approach. Nucleic Acids Res. 2010;38(Web Server issue):W609–W614. doi:10.1093/nar/gkq300
22. Daina A, Michielin O, Zoete V. SwissTargetPrediction: updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019;47(W1):W357–W364. doi:10.1093/nar/gkz382
23. Ru J, Li P, Wang J, et al. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. J Cheminform. 2014;6(1):13. doi:10.1186/1758-2946-6-13
24. Shannon P, Markiel A, Ozier O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–2504. doi:10.1101/gr.1239303
25. Waterhouse A, Bertoni M, Bienert S, et al. Swiss-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 2018;46(W1):W296–W303. doi:10.1093/nar/gky427
26. LLC, S. The {PyMOL} molecular graphics system Version 1.80 LLC, New York, NY; 2015.
27. Dayao DA, Gibson JS, Blackall PJ, Turni C. Antimicrobial resistance in bacteria associated with porcine respiratory disease in Australia. Vet Microbiol. 2014;171(1–2):232–235. doi:10.1016/j.vetmic.2014.03.014
28. Li X, Wei S, Niu S, et al. Network pharmacology prediction and molecular docking-based strategy to explore the potential mechanism of Huanglian Jiedu Decoction against sepsis. Comput Biol Med. 2022;144:105389. doi:10.1016/j.compbiomed.2022.105389
29. Zhang JN, Ma Y, Wei X-Y, et al. Remifentanil protects against lipopolysaccharide-induced inflammation through PARP-1/NF- κ B signaling pathway. Mediators Inflamm. 2019;2019:3013716. doi:10.1155/2019/3013716
30. Noguchi T, Sekiguchi Y, Kudoh Y, et al. Gefitinib initiates sterile inflammation by promoting IL-1β and HMGB1 release via two distinct mechanisms. Cell Death Dis. 2021;12(1):49. doi:10.1038/s41419-020-03335-7
31. Dong N, Li X, Xue C, et al. Astragalus polysaccharides alleviates LPS-induced inflammation via the NF-κB/MAPK signaling pathway. J Cell Physiol. 2020;235(7–8):5525–5540. doi:10.1002/jcp.29452
32. Liang Y, Hou C, Kong J, et al. HMGB1 binding to receptor for advanced glycation end products enhances inflammatory responses of human bronchial epithelial cells by activating p38 MAPK and ERK1/2. Mol Cell Biochem. 2015;405(1–2):63–71. doi:10.1007/s11010-015-2396-0
33. Chen Z, Chen Y, Pan L, et al. Dachengqi decoction attenuates inflammatory response via inhibiting HMGB1 mediated NF-κB and P38 MAPK signaling pathways in severe acute pancreatitis. Cell Physiol Biochem. 2015;37(4):1379–1389. doi:10.1159/000430403
34. Liu J, Xu Y, Yan M, Yu Y, Guo Y. 18β-Glycyrrhetinic acid suppresses allergic airway inflammation through NF-κB and Nrf2/HO-1 signaling pathways in asthma mice. Sci Rep. 2022;12(1):3121. doi:10.1038/s41598-022-06455-6
35. Feng Y, Mei L, Wang M, Huang Q, Huang R. Anti-inflammatory and pro-apoptotic effects of 18beta-glycyrrhetinic acid in vitro and in vivo models of rheumatoid arthritis. Front Pharmacol. 2021;12:681525. doi:10.3389/fphar.2021.681525
36. Hassa PO, Hottiger MO. The functional role of poly(ADP-ribose)polymerase 1 as novel coactivator of NF-kappaB in inflammatory disorders. Cell Mol Life Sci. 2002;59(9):1534–1553. doi:10.1007/s00018-002-8527-2
37. Ni S-Y, Zhong X-L, Li Z-H, et al. Puerarin alleviates lipopolysaccharide-induced Myocardial Fibrosis by inhibiting PARP-1 to prevent HMGB1-mediated TLR4-NF-κB signaling pathway. Cardiovasc Toxicol. 2020;20(5):482–491. doi:10.1007/s12012-020-09571-9
38. Ning J, Ahmed S, Cheng G, et al. Analysis of the stability and affinity of BlaR-CTD protein to β-lactam antibiotics based on docking and mutagenesis studies. J Biol Eng. 2019;13(1):27. doi:10.1186/s13036-019-0157-4
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.