Back to Journals » Infection and Drug Resistance » Volume 11
Widespread amphotericin B-resistant strains of Aspergillus fumigatus in Hamilton, Canada
Authors Ashu EE, Korfanty GA, Samarasinghe H, Pum N, You M, Yamamura D ![]() , Xu J
, Xu J ![]()
Received 12 April 2018
Accepted for publication 14 July 2018
Published 20 September 2018 Volume 2018:11 Pages 1549—1555
DOI https://doi.org/10.2147/IDR.S170952
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Eta E Ashu,1 Gregory A Korfanty,1 Himeshi Samarasinghe,1 Nicole Pum,1 Man You,1 Deborah Yamamura,2 Jianping Xu1
1Department of Biology, McMaster University, Hamilton, ON, Canada; 2Department of Pathology and Molecular Medicine, McMaster University, Hamilton, ON, Canada
Purpose: Amphotericin B (AMB) is one of the major antifungal drugs used in the management of aspergillosis and is especially recommended for treating triazole-resistant strains of Aspergillus fumigatus. However, relatively little is known about the AMB susceptibility patterns of A. fumigatus in many parts of the world. This study aims to describe the AMB susceptibility patterns in Hamilton, Ontario, Canada.
Methods: The in vitro susceptibilities of 195 environmental and clinical A. fumigatus isolates to AMB were tested by the broth microdilution method as per the Clinical and Laboratory Standards Institute’s guidelines. Catalase-generated oxygen bubbles trapped by Triton X-100 were used to quantify catalase activity in a representative group of isolates.
Results: Of the 195 isolates, 188 (96.4%) had the minimum inhibitory concentration (MIC) of AMB ≥2 mg/L, with approximately 80% and 20% of all clinical and environmental isolates having MICs of ≥ 4 mg/L. Overall, the clinical isolates were less susceptible to AMB than environmental isolates (P-value <0.001). The strain with the highest AMB MIC (16 mg/L) had one of the highest catalase activities. However, there was no correlation between AMB MIC and catalase activity in our sample.
Conclusion: The widespread AMB resistance suggests that using AMB in the management of A. fumigatus infections in Hamilton would likely result in treatment failure. Although high catalase activity may have contributed to AMB resistance in some isolates, the mechanism(s) for the observed AMB resistance in Hamilton is unknown and likely complex.
Keywords: Aspergillus fumigatus, resistance, amphotericin B, catalase activity, Hamilton, Canada
Plain language summary
There is a growing spread of resistance against triazoles, the main antifungal drugs used in the treatment of infections by Aspergillus fumigatus. In order to circumvent this problem, a panel of global experts recommended the use of amphotericin B (AMB) for the treatment of infections to which the triazole antifungal drugs have become ineffective. However, very little is known about the AMB susceptibility patterns of A. fumigatus populations in many parts of the world. Because antifungal drug susceptibility in pathogen populations can be geography-specific, this study examined three ecological samples (agricultural soil, urban park, and hospital patients) of A. fumigatus from around Hamilton, Ontario, Canada, for AMB susceptibility. Our results showed widespread AMB resistance, suggesting that the aforementioned recommendations should not be applied to A. fumigatus infections in Hamilton. Instead, alternative recommendations should be established.
Introduction
Aspergillus fumigatus is a saprophytic fungus commonly found in decaying organic matter and soil. Primarily due to its thermotolerance and its ubiquitous and abundant asexual spores, A. fumigatus is one of the most common human fungal pathogens.1 A. fumigatus is the primary causative agent of a group of infections collectively known as aspergillosis. Allergic bronchopulmonary, chronic pulmonary, and invasive aspergillosis are the three most common forms of aspergillosis. Allergic bronchopulmonary aspergillosis results from immune hypersensitivity to the presence of A. fumigatus in the lungs and is common among asthma and cystic fibrosis patients. Allergic bronchopulmonary aspergillosis can exacerbate respiratory conditions such as cystic fibrosis and can eventually progress to chronic pulmonary aspergillosis if not treated properly. Chronic pulmonary aspergillosis is a long-term A. fumigatus infection that can lead to lung scarring and/or fungal masses (aspergillomas) in lung cavities. Both allergic bronchopulmonary and chronic aspergillosis can have symptoms ranging from close to asymptomatic to low-grade fevers, generalized malaise, wheezing, headaches, and hemoptysis.2,3 Unlike allergic bronchopulmonary and chronic aspergillosis where only the respiratory system is affected, invasive aspergillosis can affect a large number of organs belonging to the digestive, urinary, and nervous systems and has mortality rates of up to 90% in immunocompromised patients.1 However, in most cases, aspergillosis can be managed with antifungals including voriconazole, itraconazole, amphotericin B (AMB), caspofungin, micafungin, and so on.
Triazoles are currently the first-line antifungals in the treatment of aspergillosis.4,5 However, there is increasing evidence for triazole resistance in natural and clinical populations of A. fumigatus.6 We recently investigated whether the use of agricultural triazole fungicide causes clinical triazole resistance in Hamilton, Ontario, Canada, and noted that there was evidence of reduced itraconazole susceptibility in both clinical and agricultural A. fumigatus isolates (where the modal minimum inhibitory concentration [MIC] for both populations was 0.5 mg/L) when compared with urban park isolates (modal MIC =0.25 mg/L).7 The results suggested that continued triazole selection pressure could drive some of the Hamiltonian isolates to become resistant to triazole antifungal drugs. Additionally, based on genotype results at nine microsatellite loci, we showed evidence for widespread sexual recombination and frequent gene flow, both of which could have a significant impact on the initiation and spread of antifungal-resistant genes throughout the Hamiltonian A. fumigatus population.7
It is currently recommended that AMB should be administered in cases of triazole-resistant invasive pulmonary and chronic aspergillosis.8 Furthermore, AMB is recommended as the core therapy for central nervous system aspergillosis.9 However, the AMB susceptibility patterns remain to be investigated in many parts of the world, including Hamilton.
AMB is a broad-spectrum polyene antifungal that can cause cell membrane depolarization and leakage by binding to ergosterol, the main sterol in fungal cell membranes. AMB has also been suggested to induce oxidative cell death in Aspergillus terreus, a species closely related to A. fumigatus.9,10 Although resistance to AMB has been frequently observed in A. terreus and to some extent in Aspergillus flavus,10,11 AMB resistance has seldom been reported in clinical and environmental A. fumigatus samples. For example, AMB resistance was absent in over 100 sequential A. fumigatus isolates obtained from 18 patients who had been solely treated with AMB or AMB and itraconazole for periods of up to 4 months.12 Likewise, Moosa et al13 showed that acquired AMB resistance is uncommon during therapy and that laboratory-acquired AMB resistance is also extremely infrequent. Nonetheless, it has been previously shown that AMB resistance can be induced in the laboratory following a single-step UV-initiated mutagenesis.14 Interestingly, a recent study of samples from Brazil showed that 27% of the A. fumigatus isolates were resistant to AMB (MIC >2 mg/L), the highest reported so far in the literature.15 This result suggests that there might be some geographic specificity in the AMB susceptibility pattern across the world. If true, the current clinical recommendation will need to be modified to reflect such geographic specificity.
The objective of this study is to investigate the AMB susceptibility pattern among ecological populations of A. fumigatus in Hamilton. We hypothesize that strains of A. fumigatus from Hamilton should be susceptible to AMB, similar to the majority of A. fumigatus populations from other geographic regions in the world. In addition, if AMB resistance were found, we expected that the mechanism of resistance might be similar to what has been reported in A. terreus and that strains with high AMB MICs should have higher catalase activities than those with lower AMB MICs.10
Methods
Environmental and clinical isolates
A total of 195 A. fumigatus isolates were obtained from in and around Hamilton as previously described.7 Briefly, 62 A. fumigatus isolates each were, respectively, obtained from urban parks within Hamilton and from farm fields located ~35 km west of Hamilton between September 2014 and January 2015. Fungicides with trade names Stratego, Prosaro, and Headline were used on these farms every other year for at least 10 years prior to our sampling. However, none of these fungicides contained AMB or other polyenes. Seventy-one clinical isolates were obtained between January and October 2015 at the Clinical Microbiology Laboratory of Hamilton Health Sciences, located at the Hamilton General Hospital, Ontario, Canada. These isolates were obtained as part of the routine hospital laboratory procedure in the identification of infectious agents for these patients. All 71 patients provided informed written consent about the use of these clinical isolates. A retrospective chart review approved by the Hamilton Integrated Research Ethics Board (project No 3328-C) was conducted to obtain patient sample prior to antifungal use. None of the patients from whom the clinical A. fumigatus samples were obtained had taken AMB. Thus, none of the A. fumigatus isolates are known to have been exposed to AMB. All clinical and environmental A. fumigatus isolates included in this study were identified microscopically by means of conidial head shape and seriation, macroscopically by colony color and texture, and by their vigorous growth at 50°C. Molecular identification was done using A. fumigatus mating type-specific primers for PCR amplification. In addition, all 195 isolates were subjected to genotyping using nine A. fumigatus-specific and highly polymorphic microsatellite markers. The details of their isolation and identification have been described in the study by Ashu et al.7
AMB susceptibility testing
The in vitro susceptibilities of all 195 A. fumigatus isolates to AMB (Selleckchem, San Francisco, CA, USA) were tested following the M38-A2 guideline of the Clinical and Laboratory Standards Institute.16 Briefly, spores were harvested from cultures grown on Sabouraud’s dextrose agar at 37°C for 48 hours, and the spore densities were adjusted to a range of approximately 5×105–5×106 CFU/mL. The standardized spore suspensions were then inoculated in RPMI-1640 medium (Sigma-Aldrich Co., St Louis, MO, USA) containing varying AMB concentrations and incubated at 37°C for 48 hours. The range of AMB concentrations tested here was from 0.03 mg/L to 16 mg/L. Candida parapsilosis (ATCC 22019) and Candida krusei (ATCC 6258) were used as reference strains. As an independent confirmation to our susceptibility test results, five representative isolates from our collection were further tested at the Public Health of Ontario Laboratory in Toronto. Susceptibility tests carried out at the Public Health of Ontario Laboratory were also done following the M38-A2 guideline of the Clinical and Laboratory Standards Institute.16 Although a clinical resistance breakpoint has not been established for AMB in A. fumigatus, MIC >2 mg/L is the recommended epidemiological cutoff value (ECV) for resistance as per CLSI M59.17–19
Test for catalase activity
To investigate whether the different AMB susceptibilities of A. fumigatus samples observed in this study were related to their catalase activities, we selected a set of 31 isolates representing the full range of AMB MICs obtained in this study (1–16 mg/L). The protocol for measuring catalase activities followed that previously described by Iwase et al.20 This method was chosen because of its ease of use, precision, and reproducibility. For each isolate, A. fumigatus was incubated in Sabouraud’s dextrose broth at 37°C for 4 days, and a cell suspension was made by mixing approximately 15 mg of cells obtained from the incubation in 100 µL of saline. Subsequently, 100 µL of undiluted hydrogen peroxide (30% v/v in H2O; Thermo Fisher Scientific, Waltham, MA, USA) was added to a transparent glass tube (13 mm in diameter ×100 mm in height) containing 100 µL of Triton X-100 (1% v/v in H2O; Sigma-Aldrich Co.) and the cell suspension. The tube content was then mixed and incubated at room temperature for 20 minutes. Following reaction completion, the height of trapped oxygen gas, visualized as foam, was measured using a ruler. Catalase from bovine liver (Sigma-Aldrich Co. [product number C1345]) was used as a control.
Correlation between AMB susceptibility and multilocus microsatellite genotype as well as triazole susceptibility patterns
Cross-resistance between azoles and polyenes drugs has been reported in various fungi including Candida species, Saccharomyces cerevisiae, and Cryptococcus neoformans.21–23 In this study, we tested whether such cross-resistance exists in the Hamiltonian population of A. fumigatus. In addition, we also examined if there was any correlation between AMB susceptibility and genetic similarity among the isolates. Both the microsatellite genotype and the triazole susceptibility data were from a previous study.7 The correlational tests were performed using the Mantel and Pearson correlation tests as implemented in R version 3.0 and Microsoft Excel 2010, respectively.
Results
AMB susceptibility patterns
Of the 195 isolates obtained and analyzed here, 188 (96.4%) had AMB MICs of ≥2 mg/L and are considered resistant. Independent testing by the Public Health of Ontario Laboratory for five separate isolates showed identical AMB MIC results to what was observed in our own laboratory for these isolates. Specifically, all AMB-resistant isolates found in our testing also showed AMB-resistant phenotype in the independent test by the Public Health of Ontario Laboratory. The AMB susceptibility results for all 195 isolates are summarized in Table 1. Of the 71 clinical isolates, 57 (80.2%) had AMB MICs of ≥4 mg/L. Among all the environmental samples, 9 (14.5%) and 15 (24.2%) in urban and agricultural populations, respectively, had MICs >4 mg/L. Compared to environmental (urban and agricultural) A. fumigatus isolates, clinical isolates were significantly less susceptible to AMB (P-value <0.001 [Mann–Whitney test]). Nonetheless, the isolate with the highest AMB MIC (>16 mg/L) was obtained from an agricultural field.
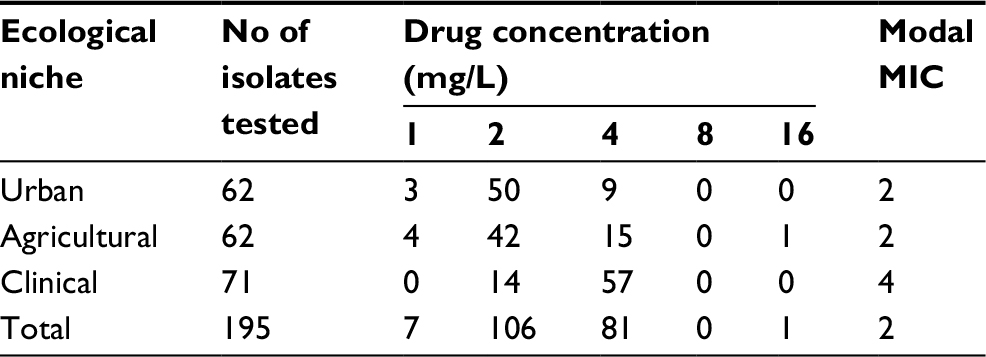 | Table 1 In vitro susceptibility profiles of Hamiltonian Aspergillus fumigatus populations to AMB Abbreviations: AMB, amphotericin B; MIC, minimum inhibitory concentration. |
Differences in catalase activity
Of the 195 isolates, the catalase activities of 31 isolates representing the broad range of obtained AMB MICs were tested. These included all seven strains with MICs of 1 mg/L; 13 of the 106 strains with MICs of 2 mg/L; 12 of the 81 strains with MICs of 4 mg/L; and only one strain with an MIC of 16 mg/L. Our analyses showed no statistically significant difference in the mean catalase activities among the three sampled MIC populations (ie, 1 mg/L, 2 mg/L, and 4 mg/L) (P-values of all three pairwise t-test results were greater than 0.1). The MIC 16 mg/L category had only one strain and thus was not compared with other three groups. Within each of the three MIC groups, there was a range of catalase activities among strains. The strain AV56 with the highest catalase activity (26.6 units) was from an urban park in Hamilton and had an MIC of 2 mg/L. Two other strains, AV29 from an urban park and CM11 from an agricultural field, shared the next highest catalase activities at 24 units each. Strains AV29 and CM11 had AMB MICs of 4 mg/L and 16 mg/L, respectively. The remaining 28 tested strains produced between 10.8 and 18.7 catalase units.
Relationship among AMB susceptibility, pairwise strain genetic distance, and triazole susceptibility patterns
There was a statistically insignificant correlation between genetic dissimilarity among the 195 strains and their differences in AMB susceptibility (r=0.035 and P-value =0.16). Similarly, there was a statistically insignificant correlation between AMB and voriconazole susceptibilities (r=0.13 and P-value =0.09). However, there was a weak positive and statistically significant correlation between AMB susceptibility and itraconazole susceptibility (r=0.20 and P-value =0.005).
Discussion
There has been an increase in the global occurrence of resistance to triazoles that are the first-line antifungals used in the management of diverse forms of aspergillosis. In cases of triazole-resistant chronic and invasive aspergillosis, AMB has been the recommended drug of choice for treatment. Furthermore, voriconazole–AMB combination therapy is recommended for initial treatment in countries/regions with high environmental triazole resistance rates (≥10%).8 However, given that the AMB susceptibility patterns can vary among geographic/ecological populations, it is necessary for local physicians and public health practitioners to understand the limitations of the aforementioned recommendations. Our results indicate that the frequencies of isolates with AMB MIC at or above the ECV for A. fumigatus in both environmental and clinical isolates are substantial, at 92% (114/124) and 100% (71/71), respectively. This result has significant implications for the empiric use of AMB in the treatment of invasive aspergillosis caused by A. fumigatus. We further show that unlike in A. terreus, catalase activity was not associated with AMB MIC at the population level. However, as shown in the “Results” section, the strain with the highest AMB MIC had the second highest catalase activity. Thus, at present, we cannot rule out the possibility that for some strains, a high catalase activity may contribute to AMB resistance through an oxidative stress response pathway in A. fumigatus. Subsequently, we discuss the implication of our results in Hamilton and Canada at large.
Over 96% of all isolates analyzed in this study had AMB MICs at or exceeding the ECV of resistance in A. fumigatus. Previous studies have reported varying frequencies of AMB-resistant A. fumigatus in clinical and environmental samples.15,17,24 However, the extent of AMB resistance reported in this study is much higher than what has been found in other parts of the world. For example, Dannaoui et al24 showed that none of the 156 clinical A. fumigatus isolates they tested were resistant to AMB. The highest percentage of AMB-resistant A. fumigatus reported so far was from a hospital in Brazil where 27.4% (46/168) of the isolates had MIC >2 mg/L.15 In a large-scale study involving eight medical centers in Canada, Spain, USA, and UK, 5.6% of (225/3,988) the clinical A. fumigatus isolates were found to be resistant to AMB.17 Furthermore, of the 3,988 isolates, only 11 (0.3%) had AMB MICs of ≥4 mg/L.17 In contrast, 82 of the 195 (42.1%) isolates in this study showed MICs of ≥4 mg/L. This percentage was a lot higher in the clinical sample where 57 of the 71 (80.2%) isolates had MICs of ≥4 mg/L. Similarly, previous studies have also shown that resistance to AMB was uncommon among environmental A. fumigatus isolates.25,26 However, 117 of the 124 (94.4%) environmental isolates analyzed here were resistant to AMB.
At present, the reason for such widespread AMB resistance in the Hamiltonian A. fumigatus population is not known. However, there are several possibilities for this phenomenon. First, the observed high AMB MIC in our sample might be due to unknown variables in our testing procedure. Lab-to-lab variability in susceptibility testing is common for many microbial pathogens and antibiotics. To eliminate this possibility, we tested each of our isolates, including the two reference strains, twice and requested an independent laboratory to test a small representative set of isolates. All tests confirmed the results presented here.
Second, the observed widespread AMB resistance might be due to a recent origin and invasion of an AMB-resistant genotype, due to the selective advantage(s) that the AMB-resistant genes conferred to progeny genotypes in Hamiltonian environments. At present, the mechanism of AMB resistance in A. fumigatus is largely unknown. However, in the related species A. terreus and other species, reduced absorption of AMB, reduced content of ergosterol in cytoplasmic membrane, enhanced expression of efflux pumps for AMB, and/or increased catalase activity to reduce oxidative stress caused by AMB have all been found to be associated with AMB resistance.10,27 If strains with mutations related to one or more of the abovementioned pathways are involved in establishing the Hamiltonian A. fumigatus population, then all strains in this population would have intrinsically elevated AMB MICs. Alternatively, mutant alleles with such effects could have recently invaded the Hamiltonian A. fumigatus population and spread to the rest of the population via sexual mating. The frequent gene flow and evidence for recombination observed in the Hamilton A. fumigatus populations are consistent with this possibility.7 The lack of an association between multilocus microsatellite genotypes and AMB MICs is also consistent with the microsatellite markers being selectively neutral and that the resistant genes are in linkage equilibrium with the microsatellite loci. If this hypothesis were true, it would be imperative to understand the genetic basis for AMB resistance in A. fumigatus, its potential selective advantage in the environments in Hamilton, and how we could prevent its spread to other geographic regions.
The third possibility is that the observed high frequency of AMB resistance in Hamilton might be a byproduct of other adaptation(s) such as to oxidative stress. Generally speaking, toxic ROS, such as superoxide and hydrogen peroxide, can be produced in nature or a human environment. In fact, in a human environment the production of ROS by innate immune cells is an important fungicidal mechanism. Catalase is a key enzyme involved in oxidative stress response in a broad range of cellular organisms as it can catalyze the formation of oxygen and water from hydrogen peroxide. This is possible due to the fact that catalase contains a heme group; oxidization of iron(III) [Fe(III)] in the heme group to Fe(IV), followed by a reduction of Fe(IV) back to Fe(III) is essential to the role of catalase in oxidative stress response. Interestingly, iron was recently shown to be essential to oxidative stress response in A. fumigatus.28 In this study, although there was no significant association between catalase activity and AMB susceptibility at the population level, all tested strains produced abundant catalase. It is entirely possible that once the catalase activity reached a certain level, there was little selective advantage to further increase its activity in nature. At present, the levels of oxidative stress that A. fumigatus strains are exposed to in vivo and in vitro and the potential oxidative damage caused by different concentrations of AMB are not known. Further investigations into these issues will allow a better understanding of the variations in catalase activities and AMB MICs among strains analyzed here.
Our results indicated a significant difference in AMB susceptibility between clinical and environmental A. fumigatus samples in Hamilton, with the clinical sample being more resistant to AMB than environmental samples. Our results differ from those obtained in the study by Araujo et al,25 where no difference in AMB susceptibility was observed between clinical and environmental A. fumigatus isolates. Given that AMB was not prescribed to treat any of the patients in this study, the decreased susceptibility to AMB in the clinical population of A. fumigatus was not due to prolonged AMB therapy. Similarly, polyene antifungals were not used on the farms that we sampled and thus prior exposure to AMB could not be used to explain the observed widespread environmental AMB resistance. Furthermore, the lack of triazole resistance among our set of 195 isolates7 suggests that the widespread AMB resistance is not due to triazole resistance in either the Hamiltonian clinical or the environmental A. fumigatus populations.
Regardless of the reason(s) for the differences in AMB susceptibility among the Hamiltonian A. fumigatus ecological populations, our results clearly suggest that using AMB in the management of triazole-resistant or even triazole-susceptible aspergillosis in Hamilton may cause treatment failure. However, further research into the therapeutic implications of MIC ≥2 mg/L to AMB is required. On the other hand, we note that all five isolates tested at the Public Health of Ontario Laboratory were very sensitive to echinocandins (MICs ≤0.03 mg/L for all isolates), including anidulafungin, caspofungin, and micafungin. Unlike the polyenes and triazoles that either directly interact with membrane ergosterol (the polyenes including AMB) or target a key enzyme involved in the synthesis of ergosterol (the triazole drugs), the echinocandins target a different part of the fungal cell, the cell wall biosynthesis. As a result, unlike correlations observed between AMB MIC and triazole MIC in fungi,21–23 cross-resistance has not been observed between the echinocandins and the other two types of drugs. From a health policy perspective, these results suggest that echinocandins should be the therapy of choice in the management of triazole-resistant A. fumigatus in Hamilton.
Whether AMB resistance is also common in other parts of Canada remains to be investigated. However, although the highest rate of AMB resistance reported so far in the literature was significantly lower than ours (27.4% vs 96%), similar to our patient samples, none of the patients in the study by Reichert-Lima et al15 had taken any AMB. Altogether, these results suggest that greater efforts are needed to survey the prevalence of AMB resistance across geographic populations of A. fumigatus in both clinical and environmental samples and to modify the therapeutic recommendations accordingly based on geography-specific antifungal drug-susceptibility data.
Acknowledgment
Research in our laboratory on fungal infectious diseases was supported by the Natural Science and Engineering Research Council (NSERC; CRDPJ474638-14) of Canada.
Author contributions
EA and JX conceived and designed the study. Experiments were performed by EA, GK, HS, NP, and MY. Data were analyzed by EA and JX. DY provided patient samples and significantly contributed toward drafting and revising the paper. All authors read and approved the final manuscript. All authors contributed toward data analysis, drafting and critically revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Dagenais TR, Keller NP. Pathogenesis of Aspergillus fumigatus in Invasive Aspergillosis. Clin Microbiol Rev. 2009;22(3):447–465. | ||
Maturu VN, Agarwal R. Itraconazole in chronic pulmonary aspergillosis: In whom, for how long, and at what dose? Lung India. 2015;32(4):309–312. | ||
Agarwal R. Allergic bronchopulmonary aspergillosis. Chest. 2009; 135(3):805–826. | ||
Segal BH, Walsh TJ. Current approaches to diagnosis and treatment of invasive aspergillosis. Am J Respir Crit Care Med. 2006;173(7):707–717. | ||
Patterson TF, Thompson GR, Denning DW, et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clin Infect Dis. 2016;63(4):e1–e60. | ||
Verweij PE, Chowdhary A, Melchers WJ, Meis JF. Azole resistance in Aspergillus fumigatus: can we retain the clinical use of mold-active antifungal azoles? Clin Infect Dis. 2016;62(3):362–368. | ||
Ashu EE, Kim GY, Roy-Gayos P, et al. Limited evidence of fungicide-driven triazole-resistant Aspergillus fumigatus in Hamilton, Canada. Can J Microbiol. 2018;64(2):119–130. | ||
Verweij PE, Ananda-Rajah M, Andes D, et al. International expert opinion on the management of infection caused by azole-resistant Aspergillus fumigatus. Drug Resist Updat. 2015;21-22:30–40. | ||
vanden Bossche H, Warnock DW, Dupont B, et al. Mechanisms and clinical impact of antifungal drug resistance. J Med Vet Mycol. 1994;32 Suppl 1:189–202. | ||
Blum G, Perkhofer S, Haas H, et al. Potential basis for amphotericin B resistance in Aspergillus terreus. Antimicrob Agents Chemother. 2008;52(4):1553–1555. | ||
Gonçalves SS, Stchigel AM, Cano J, Guarro J, Colombo AL. In vitro antifungal susceptibility of clinically relevant species belonging to Aspergillus section Flavi. Antimicrob Agents Chemother. 2013;57(4):1944–1947. | ||
Dannaoui E, Meletiadis J, Tortorano AM, et al. Susceptibility testing of sequential isolates of Aspergillus fumigatus recovered from treated patients. J Med Microbiol. 2004;53(Pt 2):129–134. | ||
Moosa MY, Alangaden GJ, Manavathu E, Chandrasekar PH. Resistance to amphotericin B does not emerge during treatment for invasive aspergillosis. J Antimicrob Chemother. 2002;49(1):209–213. | ||
Manavathu EK, Alangaden GJ, Chandrasekar PH. In-vitro isolation and antifungal susceptibility of amphotericin B-resistant mutants of Aspergillus fumigatus. J Antimicrob Chemother. 1998;41(6):615–619. | ||
Reichert-Lima F, Lyra L, Pontes L, et al. Surveillance for azoles resistance in Aspergillus spp. highlights a high number of amphotericin B-resistant isolates. Mycoses. 2018;61(6):360-365. | ||
Clinical and Laboratory Standards Institute. Reference method for broth dilution antifungal susceptibility testing of filamentous fungi. Approved standard. 2nd ed. Wayne, PA, USA: CLSI; 2002:M38–A2. | ||
Espinel-Ingroff A, Cuenca-Estrella M, Fothergill A, et al. Wild-type MIC distributions and epidemiological cutoff values for amphotericin B and Aspergillus spp. for the CLSI broth microdilution method (M38-A2 document. Antimicrob Agents Chemother. 2011;55(11):5150–5154. | ||
Elefanti A, Mouton JW, Verweij PE, Zerva L, Meletiadis J. Susceptibility breakpoints for amphotericin B and Aspergillus species in an in vitro pharmacokinetic-pharmacodynamic model simulating free-drug concentrations in human serum. Antimicrob Agents Chemother. 2014;58(4):2356–2362. | ||
Clinical and Laboratory Standards Institute. Epidemiological Cutoff Values for Antifungal Susceptibility Testing. CLSI supplement M59. 2nd ed. Wayne, PA: Clinical and Laboratory Standards Institute; 2018. | ||
Iwase T, Tajima A, Sugimoto S, et al. A simple assay for measuring catalase activity: a visual approach. Sci Rep. 2013;3:3081. | ||
Kelly SL, Lamb DC, Kelly DE, et al. Resistance to fluconazole and cross-resistance to amphotericin B in Candida albicans from AIDS patients caused by defective sterol delta5,6-desaturation. FEBS Lett. 1997;400(1):80–82. | ||
Joseph-Horne T, Hollomon D, Loeffler RS, Kelly SL. Cross-resistance to polyene and azole drugs in Cryptococcus neoformans. Antimicrob Agents Chemother. 1995;39(7):1526–1529. | ||
Forastiero A, Mesa-Arango AC, Alastruey-Izquierdo A, et al. Candida tropicalis antifungal cross-resistance is related to different azole target (Erg11p) modifications. Antimicrob Agents Chemother. 2013;57(10):4769–4781. | ||
Dannaoui E, Persat F, Monier MF, Borel E, Piens MA, Picot S. In-vitro susceptibility of Aspergillus spp. isolates to amphotericin B and itraconazole. J Antimicrob Chemother. 1999;44(4):553–555. | ||
Araujo R, Pina-Vaz C, Rodrigues AG. Susceptibility of environmental versus clinical strains of pathogenic Aspergillus. Int J Antimicrob Agents. 2007;29(1):108–111. | ||
Guinea J, Peláez T, Alcalá L, Ruiz-Serrano MJ, Bouza E. Antifungal susceptibility of 596 Aspergillus fumigatus strains isolated from outdoor air, hospital air, and clinical samples: analysis by site of isolation. Antimicrob Agents Chemother. 2005;49(8):3495–3497. | ||
Purkait B, Kumar A, Nandi N, et al. Mechanism of amphotericin B resistance in clinical isolates of Leishmania donovani. Antimicrob Agents Chemother. 2012;56(2):1031–1041. | ||
Brandon M, Howard B, Lawrence C, Laubenbacher R. Iron acquisition and oxidative stress response in Aspergillus fumigatus. BMC Syst Biol. 2015;9:19. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.