Back to Journals » Substance Abuse and Rehabilitation » Volume 17
What Do Thirty Years of Neuroimaging Research Tell Us About Recreational Cannabis Use and Brain Integrity? A Narrative Review of the Multimodal Neuroimaging Evidence to Date
Authors Murphy E, Suo C, Poudel G, Thomson H, Lorenzetti V
Received 27 November 2025
Accepted for publication 27 April 2026
Published 15 May 2026 Volume 2026:17 584766
DOI https://doi.org/10.2147/SAR.S584766
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rajendra Badgaiyan
Ethan Murphy,1 Chao Suo,1– 4 Govinda Poudel,5 Hannah Thomson,1 Valentina Lorenzetti1,6
1Neuroscience of Addiction and Mental Health Program, Australian Catholic University, Melbourne, VIC, Australia; 2Brain Park, Monash University, Melbourne, VIC, Australia; 3QIMR Berghofer, Brisbane, QLD, Australia; 4School of Psychology, University of Queensland, Brisbane, QLD, Australia; 5Mary MacKillop Institute for Health Research, Australian Catholic University, Melbourne, VIC, Australia; 6Clinical Psychopharmacology Unit, University College London, London, UK
Correspondence: Valentina Lorenzetti, Neuroscience of Addiction and Mental Health Program, Australian Catholic University, 115 Victoria Parade, Level 5 Daniel Mannix Building, Fitzroy, Melbourne, VIC, 3065, Australia, Email [email protected]
Abstract: The consumption of cannabinoids is highly prevalent and has been associated with altered structural, functional, and metabolic brain integrity, measured using PET and neuroimaging tools. However, the current neuroimaging evidence has been summarized by distinct modalities that measure different metrics of brain integrity, precluding a comprehensive understanding of the underlying neurobiology. A non-systematic narrative review method was used to summarize the multimodal neuroimaging evidence on brain integrity from experimental studies of cannabinoid intoxication and observational studies in non-intoxicated cannabis users. Consistent evidence showed that acute intoxication with delta-9-tetrahydrocannabinol (THC) was associated with greater brain activity in fronto-striatal pathways. For regular cannabis users compared to controls, there was consistent cross-sectional evidence of lower hippocampal volumetry and white matter microstructure of the superior longitudinal fasciculus, and of different fronto-striatal activity and connectivity during cue-reactivity tasks and resting-state. In cannabis users, there was emerging evidence from Positron Emission Tomography studies of altered neurochemistry in fronto-striatal pathways (eg, lower N-acetyl aspartate); lower glucose metabolism in the frontal cortex; and lower density of cannabinoid receptors, which may reverse with abstinence. Longitudinal multimodal neuroimaging studies are required to confirm if brain differences predate or follow the onset of cannabis use or cannabis use disorder, and whether changes in brain integrity in people who use cannabis dissipate with abstinence.
Keywords: marijuana, MRS, fMRI, sMRI, PET, magnetic resonance imaging
Introduction
Over 228 million individuals report consuming cannabinoids worldwide, making cannabis the most commonly used regulated psychoactive substance in the world.1 As liberal cannabis policies have spread internationally, cannabinoid-based products have become increasingly more accessible and diverse.2 Meanwhile, the amount of Δ-9-tetrahydrocannabinol (THC), the primary psychoactive cannabinoid of cannabis, has increased over time in different world regions, including from 4% in 1995 to 14% in 2019 in the United States3,4 and from 7% in 2010 to 11% in 2019 in Europe.5 These statistics are worrying, as THC has psychoactive, anxiogenic, psychotogenic and addictive properties.6–8
Relatedly, the consumption of cannabis can come with negative physical and psychosocial outcomes. For example, intoxication with cannabinoids has been associated with increased anxiety and psychotic-like symptoms,9,10 motor vehicle crashes resulting in injury or death,10–12 and reduced cognitive function, including working memory, verbal fluency, executive functioning, and time estimation.13,14 Regular cannabis consumption has also been linked to respiratory issues,15 cannabis craving and withdrawal,16 and greater symptoms and presence of psychiatric disorders, including cannabis use disorder (CUD),14,17 and psychotic, mood, and anxiety disorders.14,18
The adverse outcomes associated with acute intoxication with cannabinoids and the regular consumption of cannabinoids have been partly attributed to the impact of THC on the brain.8,19 Indeed, in vitro animal evidence shows that THC exerts its psychoactive effects via binding to cannabinoid type 1 (CB1) G protein-coupled receptors.7 In vitro human evidence demonstrates that CB1 receptors are densely innervated in multiple brain regions implicated in cognitive processes reportedly altered during cannabinoid intoxication and in regular cannabis users.20 The regions include: the prefrontal cortex (PFC), including the orbitofrontal cortex (OFC) and the anterior cingulate cortex (ACC), which are implicated in executive function, cognitive control, and disinhibition;21 the amygdala, which is implicated in craving;22 the hippocampus, which is involved in stress, learning, and memory;23 the cerebellum, which is implicated in salience, motivation, and executive function;24 and the basal ganglia, including the striatum and the globus pallidus, which are involved in reward processing.25 According to prominent neuroscientific theories of addiction, the chronic consumption of cannabinoids can be ascribed to neuroadaptations22 in pathways high in CB1 receptors outlined above, and implicated in addiction-related cognitive processes such as interoception (eg, insula).22
In the last three decades, in vivo neuroimaging studies in humans have begun to uncover the neurobiology of cannabinoid intoxication and regular cannabis use compared to non-use. While several high-quality reviews of the neuroimaging evidence in cannabis users to date have been published,26,27 the reviews focus either on outdated evidence (eg, up to year 2019)28 and therefore lack an up-to-date summary; or on individual neuroimaging techniques or two neuroimaging techniques, such as structural T1-weighted magnetic resonance imaging (sMRI)29,30 and functional MRI (fMRI).26,27 The neurobiological mechanisms of cannabis use and related problems are likely complex and may not be fully understood with an individual neuroimaging modality. A comprehensive synthesis of the evidence regarding the impact of cannabis exposure on brain integrity is warranted, especially in light of global decriminalization trends and the subsequent rise in cannabis availability2 and potency.3–5 Such an overview is essential to provide evidence-based guidance on cannabis use and brain health for the general community, vulnerable people who consume cannabinoids, clinical practitioners, and policy-makers. However, the current evidence should be considered in light of several methodological limitations, including but not limited to cross-sectional study designs, heterogeneous metrics and reporting of cannabis use patterns and related problems, variable cannabinoid dosing and product heterogeneity (eg, THC:CBD ratios, routes of administration), under-representation of females, and inconsistent collection and adjusting for confounding factors, such as polysubstance use, via means of design (eg, matching between groups) or statistical analyses (eg, covariate).
This narrative review aims to synthesize the evidence from multimodal human neuroimaging studies to date. The synthesis was achieved via examining the neural correlates of acute cannabinoid intoxication and of regular cannabis consumption in non-intoxicated participants (ie, henceforth termed cannabis users) compared to controls. The multimodal neuroimaging techniques included in the review are briefly described in Supplementary Materials Section 1.1, and illustrated in Figure 1, and include positron emission tomography (PET);31 fMRI;32 sMRI;32 diffusion-weighted imaging (DWI);33 and magnetic resonance spectroscopy (MRS).34 To that end, a series of searches of the electronic database Google Scholar were conducted to identify relevant studies between any time and October 2024. The following search terms were used: “cannabis”; OR “THC”; “tetrahydrocannabinol” OR “marijuana”; AND “neuroimaging”; OR “brain imaging”; OR “fMRI”; OR “functional magnetic resonance imaging”; OR “BOLD”; OR “brain function”; OR “magnetic resonance imaging”; OR “sMRI”; OR “structural MRI”; OR “structural magnetic resonance imaging”; OR “brain volume”; OR “brain structure”; OR “PET”; OR “positron emission tomography”; OR “MRS”; OR “magnetic resonance spectroscopy”. Studies were included if they: (i) examined human participants; (ii) were written in the English language; (iii) were full-text peer-reviewed articles; (iv) assessed a sample of people who consume cannabis as defined by each study, compared to non-using controls, or a sample who were acutely administered THC, compared to placebo. We excluded studies that (i) examined non-human participants; (ii) were published in languages other than English; (iii) were non-peer-reviewed (eg, conference abstracts); (iv) the sample selected, endorsed at a group level dependence/severe substance use disorders on substances other than cannabis or tobacco; (v) the sample selected, endorsed major psychiatric (eg, schizophrenia) or neurological (eg, epilepsy) or health disorders (eg, cancer) at a group level. Full-text articles were cross-referenced for additional relevant studies.
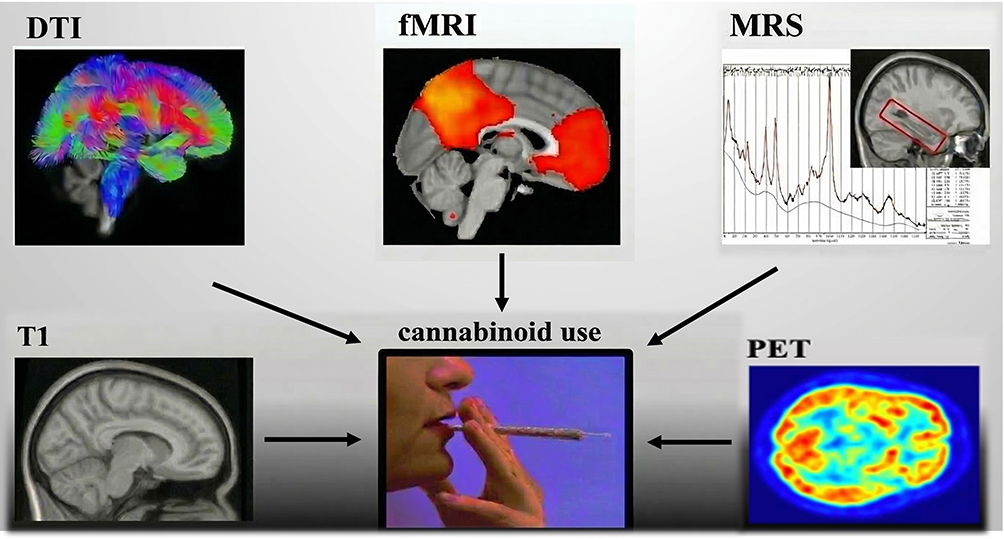 |
Figure 1 An Overview of the Non-invasive Neuroimaging Tools used to Assess Brain Integrity and Cannabis Use. Abbreviations: ACC, anterior cingulate cortex; BOLD, blood oxygen level-dependent; CB1, cannabinoid type 1 receptor; CBD, cannabidiol; CUD, cannabis use disorder; CUDIT, cannabis use disorder identification test; dlPFC, dorsolateral prefrontal cortex; DWI, diffusion-weighted imaging; fMRI, functional magnetic resonance imaging; MPS, marijuana problem scale; MRS, magnetic resonance spectroscopy; NAA, N-acetyl aspartate; OFC, orbitofrontal cortex; PCC, posterior cingulate cortex; PET, positron emission tomography; PFC, prefrontal cortex; ROI, region-of-interest; sMRI, structural T1-weighted magnetic resonance imaging; THC, delta-9-tetrahydrocannabinol; VBM, voxel-based morphometry. |
Neuroimaging Evidence on Brain Integrity During THC Intoxication
As overviewed in Table 1, a substantial number of PET and fMRI studies have examined brain metabolism, blood flow, neurotransmission, and function during acute intoxication with THC. Overall, the studies provided consistent evidence that THC intoxication affects distinct metrics of brain integrity, most commonly brain function within fronto-striatal regions.
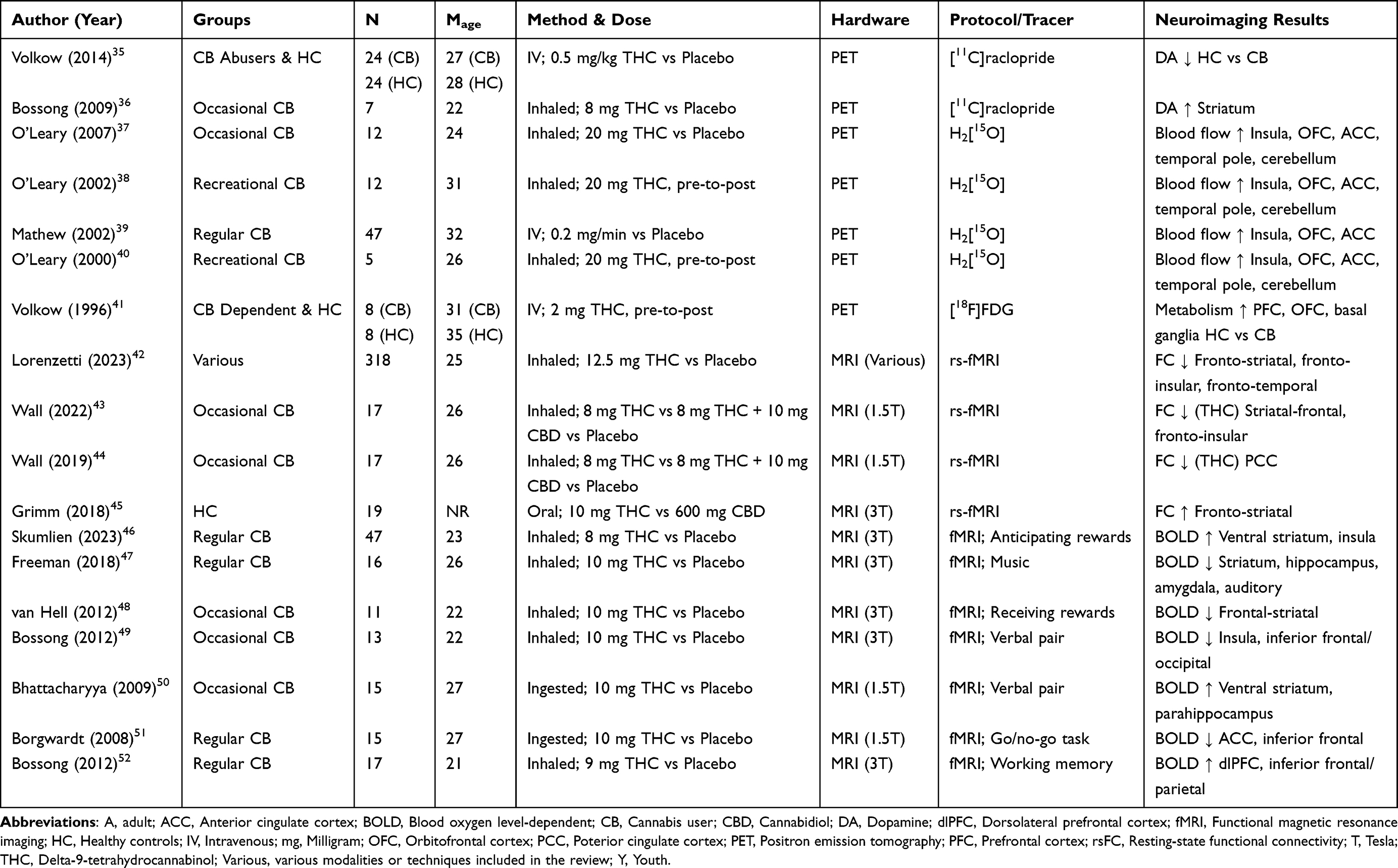 |
Table 1 Overview of Demographics, THC Administration, and Neuroimaging Results from THC Intoxication Studies |
PET Studies Investigating Changes in Brain Metabolism, Blood Flow, and Neurotransmission During THC Intoxication
Results from PET studies consistently demonstrated that acute THC intoxication changes brain metabolism and blood flow, and reduces dopamine release in frontal-insular regions in cannabis users compared to controls. A series of four studies employing H2[15O]-PET, administered 20 mg of inhaled THC vs placebo to 76 cannabis users (Mage = 29).37–40 Cannabis users intoxicated with THC vs placebo showed increased cerebral blood flow in the insula and several frontal regions, including the OFC and the ACC.37–40 Second, two PET studies examined how THC intoxication affects dopamine release using D2/D3 receptor tracer [11C]raclopride.35,36 Administration of 8 mg of inhaled THC vs placebo increased dopamine release in controls (n = 31; Mage = 26).35,36 In contrast, intravenous injection of 0.5 mg THC/kg vs placebo did not change dopamine release in cannabis users (n = 24; Mage = 27).35,36 Third, an individual 2-deoxy-2-(18F)fluoro-D-glucose PET study administered 2 mg of THC intravenously to 8 cannabis users (Mage = 31) and 8 controls (Mage = 35).41 THC intoxication increased cerebellar glucose metabolism in both cannabis users and controls.41 However, only the cannabis group demonstrated increased metabolism in the OFC, PFC, and basal ganglia.41 Overall, PET evidence shows THC intoxication-induced changes in brain metabolism, blood flow, and dopamine release.
fMRI Studies: Brain Functional Changes During THC Intoxication
The fMRI evidence to date examining brain function during acute intoxication is growing and shows consistent changes in brain function, particularly within fronto-striatal pathways.
Resting State Functional Connectivity fMRI During THC Intoxication
There was consistent evidence that acute intoxication with cannabis alters brain function at rest, particularly between striatal and frontal regions involved in reward processing and disinhibition, respectively.53 One review of 13 studies in young cannabis users (n = 318; Mage = 25) showed that the type of cannabinoid administered (eg, THC, cannabidiol [CBD] or cannabis plant matter) differentially changed resting state functional connectivity.42 For example, inhalation of 12.5 mg THC vs placebo reduced frontal-striatal connectivity (eg, ACC, frontal cortex, nucleus accumbens).42 Similarly, lower resting state functional connectivity emerged during intoxication with cannabis plant matter vs placebo, between fronto-insular, fronto-temporal and fronto-striatal pairs,42 and in correlation with greater self-report subjective intoxication.44,54,55 In young cannabis users (n = 19), oral administration of 10 mg of THC vs 600 mg of CBD increased fronto-striatal connectivity.45 Additionally, emerging evidence from two studies in occasional cannabis users (n = 17; Mage = 26) suggests that co-administering THC and CBD reduces the effect of THC on resting state functional connectivity. Indeed, inhaling 8 mg of THC reduced resting state functional connectivity between striatal-frontal and fronto-insular pairs, compared to inhaling 8 mg of THC concurrently with 10 mg of CBD.43,44 Overall, THC and cannabis intoxication were associated with functional changes in fronto-striatal pathways at rest.
Task-Based fMRI Studies: Brain Function During THC Intoxication
There were inconsistent results from a growing number of task-based fMRI studies that acute THC intoxication affects brain activity across reward, learning and memory, and additional fMRI tasks.
Reward Processing Tasks During THC Intoxication
During fMRI tasks of reward processing, there was emerging, inconsistent, and mixed evidence that acute THC intoxication changes brain activity, particularly in striatal regions during the receipt and anticipation of rewards.56 For example, a group of cannabis users (n = 11; Mage = 22) administered 10 mg of inhaled THC vs placebo, demonstrated reduced frontal-striatal activity when receiving monetary rewards.48 In separate samples of youth (n = 24; Mage = 17) and adult cannabis users (n = 23; Mage = 27), intoxication with 8 mg inhaled THC vs placebo increased ventral striatal and insular activity while anticipating monetary rewards.46 Further, in cannabis users (n = 16; Mage = 26), intoxication with 10 mg inhaled THC vs placebo lowered striatal, hippocampal, amygdalar, and auditory activity while listening to music.47 Overall, the direction and location of THC-induced changes were inconsistent during reward processing fMRI tasks. The inconsistent findings might be explained by the use of heterogeneous fMRI tasks.
Verbal Learning and Memory fMRI Tasks During THC Intoxication
Only two studies in occasional cannabis users examined the relationship between brain activity during verbal pair associate fMRI tasks following the oral administration of 10 mg THC vs placebo.49,50 Both studies provided mixed results for THC vs placebo; one study (n = 15; Mage = 27) demonstrated increased activity in the ventral striatum during memory retrieval and parahippocampus during memory encoding,50 while a second study (n = 13; Mage = 22) found lower insular and inferior frontal/occipital activity during memory encoding.49
Inhibitory and Working Memory fMRI Tasks During THC Intoxication
Only individual studies have examined how THC intoxication affects brain function during additional fMRI tasks. One study examined brain activity during a go/no-go inhibitory fMRI task in cannabis users (n = 15; Mage = 27) who ingested 10 mg of THC.51 Cannabis users showed decreased inhibitory-related ACC and inferior frontal activity during THC intoxication vs placebo.51 A separate study examined brain activity during a working memory fMRI task in young cannabis users (n = 17; Mage = 21) who inhaled 9 mg THC vs placebo.52 Young cannabis users showed increased working memory-related activity in dorsolateral PFC (dlPFC) and inferior frontal/parietal during THC vs placebo.52
Brain Integrity from Case Control Studies: Non-Intoxicated Cannabis Users Compared to Controls
As summarized in Table 2, this section overviews the multimodal neuroimaging evidence to date from PET, sMRI, DWI, MRS, and fMRI studies, investigating differences in brain integrity between non-intoxicated cannabis users and controls.
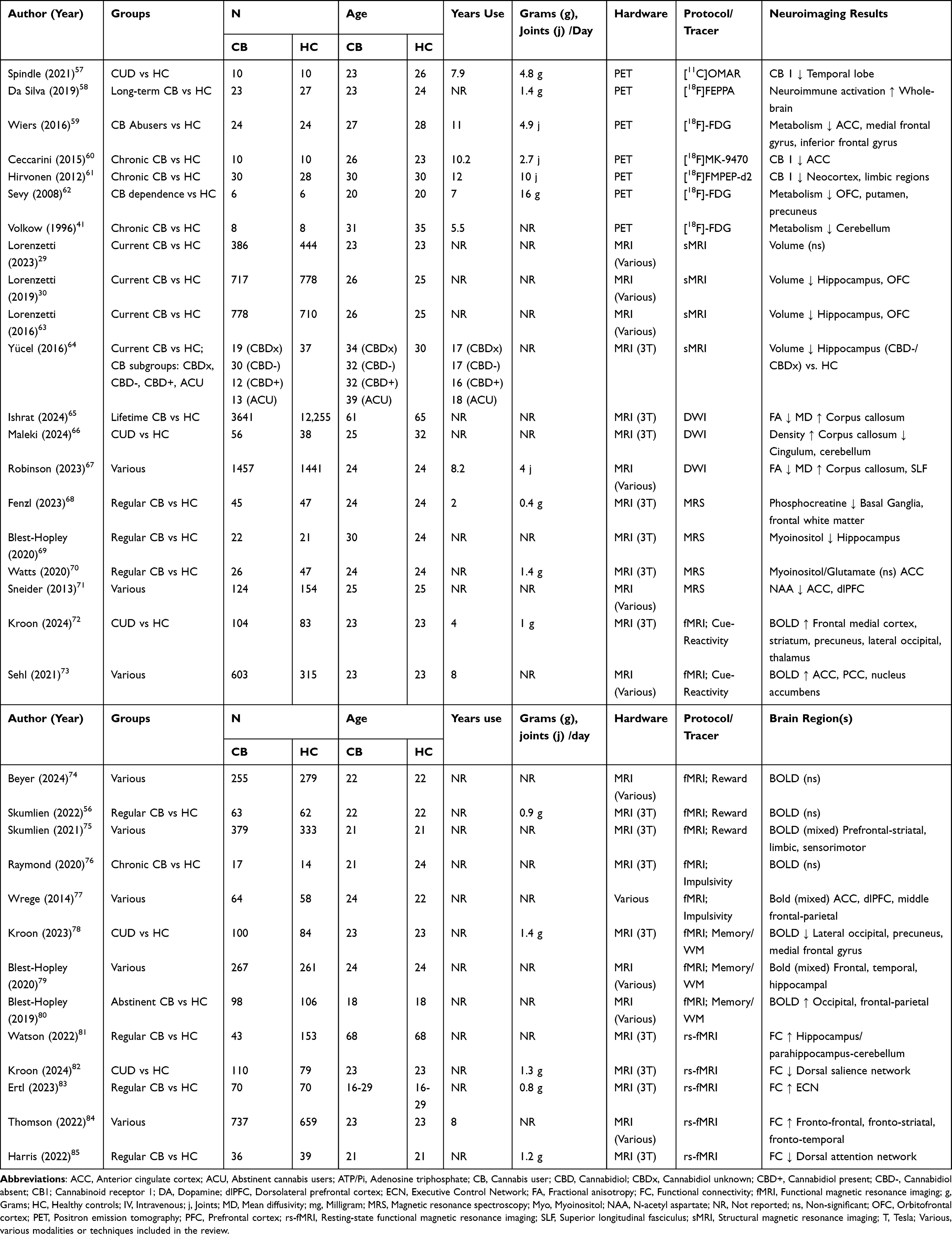 |
Table 2 Overview of Demographics, Cannabis Use, and Neuroimaging Results from Case-Control Studies |
PET Studies Investigating Group Differences in Brain Metabolism, Cannabinoid Receptor Density, and Neuroimmune Activation in Cannabis Users Compared to Controls
Emerging evidence from PET studies has examined group differences between cannabis users and controls in brain metabolism,59,62 CB1 receptor density,61 and neuroimmune activation58 using different techniques, such as 2-deoxy-2-(18F)fluoro-D-glucose (FDG), [18F]FMPEP-d2, and [18F]FEPPA PET. A seminal FDG PET study showed lower cerebellar glucose metabolism in cannabis users (n = 8; Mage = 31) compared to controls (n = 8; Mage = 35).41 Similarly, lower glucose metabolism was reported in two [18F]-FDG PET studies in cannabis users (n = 30; Mage = 26) compared to controls (n = 30; Mage = 26).59,62 Further, lower CB1 receptor density was demonstrated in the neocortex and limbic regions using [18F]FMPEP-d2 PET in cannabis users (n = 30; Mage = 30) compared to controls (n = 28; Mage = 30), in correlation with the duration of cannabis use;61 in the ACC using [18F]MK-9470 PET in cannabis users (n = 10; Mage = 26) compared to controls (n = 10; Mage = 23);57,60 and in the temporal lobe using [11C]OMAR PET in cannabis users (n = 10; Mage = 23) compared to controls (n = 10; Mage = 26).57,60 Notably, an individual study reported that CB1 receptor density normalized after a 4-week monitored abstinence.61 Additional [18F]FEPPA PET work demonstrated higher whole-brain neuroimmune activation (ie, translocator protein levels) in cannabis users, particularly those with a CUD (n = 23; Mage = 23), compared to controls (n = 27; Mage = 24).58 Overall, PET studies of cannabis users, compared to controls, demonstrated lower cerebellar metabolism, reduced CB1 receptor density in cortical and limbic regions that may be restored following a period of abstinence, and greater whole-brain neuroimmune activation.
T1-Weighted Studies in Cannabis Users Compared to Controls: Volumetry
A substantial number of T1-weighted MRI studies to date have compared volumetry between cannabis users and controls.29,63 There were no significant differences between cannabis users (n = 778; Mage = 26) and controls (n = 710; Mage = 25) for global brain metrics, including total brain volume, intracranial volume, total grey matter volume, and total white matter volume.63 Lower volumes in the OFC and hippocampus emerged in a review of 31 studies in cannabis users (n = 778; Mage = 26) compared to controls (n = 710; Mage = 25).63 Lower OFC and hippocampal volumes were confirmed in a meta-analysis of cannabis users (n = 717; Mage = 26) compared to controls (n = 778; Mage = 25).30 Age may moderate the relationship between cannabis use and volumetry. In younger samples aged up to 26 years, no volumetric differences emerged between cannabis users (n = 386; Mage = 23) and controls (n = 444; Mage = 23).29
Specific cannabinoids may moderate the association between cannabis use and volumetry. One study compared hippocampal NAA between 4 groups, including 3 groups of cannabis users and a control group (n = 37; Mage = 30).64 The cannabis groups were separated depending on cannabinoid profile based on toxicology analyses from urine and hair samples, and were: (i) presence of THC metabolites in urine, but unknown cannabinoids in hair analysis (CBDx, n = 19, Mage = 34); (ii) confirmed THC but no CBD from hair analysis (CBD−, n = 30; Mage = 32); (iii) presence of both THC and CBD in hair analysis (CBD+, n = 12; Mage = 32); and (iv) regular cannabis users who reported abstinence for an average of 2.4 years, corroborated by absence of cannabinoids in urine and hair analysis (n = 13; Mage = 39).64 Volumes were significantly lower in the cannabis groups: without CBD (CBD-) than controls (11% reduction), and with unknown hair cannabinoids (CBDx) than controls (7% reduction). There were no significant hippocampal volume differences between former regular users and controls.64 Therefore, CBD might mitigate the impact of cannabis use on hippocampal volumetry, and abstinence may recover volume reductions. Finally, the evidence to date provided inconsistent evidence for an association between specific cannabis use patterns and volumetry, and only a few studies reported significant correlations between lower volumes and greater cannabis dosage.63 However, no significant associations emerged in a meta-analytical evaluation of the evidence.30 Overall, T1-weighted evidence showed that reduced volumes were confined to specific brain regions, including the OFC and the hippocampus.
DWI Studies in Cannabis Users Compared to Controls: White Matter Microstructure
DWI evidence consistently demonstrated different white matter microstructure in cannabis users compared to controls, particularly within the corpus callosum and superior longitudinal fasciculus.65–67 A review of 30 DWI studies in cannabis users (n = 1457; Mage = 24) compared to controls (n = 1441; Mage = 24) consistently reported lower white matter microstructural integrity (ie, lower fractional anisotropy and greater mean diffusivity) in cannabis users.67 The more consistently altered white matter fiber tracts included the corpus callosum, which connects the right and left cerebral hemispheres; and the superior longitudinal fasciculus, which connects the frontal, parietal, and temporal lobes.67 A few DWI studies showed consistent associations between earlier age of cannabis use onset and a longer duration of use and lower white matter integrity in cannabis users.86–89 Two recent DWI studies also confirm lower white matter microstructural integrity. A study from the UK Biobank (https://www.ukbiobank.ac.uk/) reported worse corpus callosum integrity (eg, lower fractional anisotropy and higher mean diffusivity) in older participants who reported consuming cannabis in their lifetime (n = 3641; Mage = 61) compared to controls (n = 12255; Mage = 65).65 Notably, Mendelian randomization demonstrated no causal relationship between cannabis use in the same sample of older adults and white matter microstructure.65 Finally, a recent study combined connectome and Fixel-Based Analysis methodologies to compare CUD (n = 56; Mage = 25) and control groups (n = 38; Mage = 32).66 CUD individuals exhibited higher white matter density in the corpus callosum and lower white matter density in the cingulum and right cerebellum.66 Overall, DWI evidence points to consistently different white matter integrity in the corpus callosum in cannabis users compared to controls.
MRS Studies in Cannabis Users Compared to Controls: Neurochemistry
There was mixed MRS evidence that cannabis users demonstrated alterations in markers of neuronal viability (NAA) and glial cell metabolite markers (myoinositol), glutamate and other neurochemicals (eg, adenosine triphosphate, phosphocreatine).64,71 A review of eight MRS studies reported emerging evidence that cannabis users (n = 124; Mage = 25) compared to controls (n = 154; Mage = 25) had reduced NAA levels, most consistently in the ACC and dorsolateral PFC.71 There was also emerging evidence of associations between frontal NAA and duration of cannabis use.90–93 However, a more recent study found no differences in NAA levels within the ACC in cannabis users with adolescent onset (n = 22; Mage = 30) compared to controls (n = 21; Mage = 24).69 A further study found that hippocampal NAA was significantly lower in the cannabis groups: without CBD in hair analysis (CBD-), compared to controls (14% lower), and to former regular users (16% lower); with unknown hair cannabinoids (CBDx) than former regular users.64 There were no significant NAA differences between former regular users and controls.64 Thus, CBD might minimize the effect of cannabis use on hippocampal NAA, and abstinence may recover hippocampal NAA levels. Additional MRS research on brain metabolites other than NAA comprises largely individual studies. For example, myoinositol or glutamate markers in the ACC did not significantly differ between cannabis users (n = 26; Mage = 24) and controls (n = 47; Mage = 24).70 Similarly, levels of adenosine triphosphate and inorganic phosphate across five brain regions – frontal grey and white matter, thalamus, basal ganglia, and temporal lobe – were comparable between cannabis users (n = 45; Mage = 24) and controls (n = 47; Mage = 24).68 Yet, in the same sample, phosphocreatine was lower in the cannabis group in the left basal ganglia and left frontal white matter.68 Further, in a separate study, myoinositol but not glutamate levels in the hippocampus were lower in cannabis users with adolescent use onset (n = 22; Mage = 30) compared to controls (n = 21; Mage = 24).69 In sum, MRS studies showed that NAA levels in the hippocampus and ACC were inconsistently altered in cannabis users.
Cue-Reactivity fMRI Studies Investigating Brain Function in Cannabis Users Compared to Controls
Mounting fMRI cue-reactivity studies have investigated brain function in cannabis users and demonstrated differences while watching cannabis cues compared to neutral cues in several brain pathways implicated in decision-making and reward processing.72,73 A review of 18 fMRI studies comparing brain function between regular cannabis users (n = 603; Mage = 23) and controls (n = 315; Mage = 23) demonstrated a pattern of greater activity in frontal and striatal regions: ACC, posterior cingulate cortex [PCC], and nucleus accumbens.73 In two studies, cue-reactivity-related brain function in frontal regions (eg, OFC, ACC) and PCC correlated with cannabis use problems (eg, scores on the Marijuana Problem Scale [MPS] and Cannabis Use Disorder Identification Test [CUDIT])94,95 However, no association emerged between cue-reactivity-related brain function and cannabis frequency or dosage.73 A more recent multi-site study recruited 104 participants from Texas (USA) and 83 from the Netherlands.72 The CUD group (n = 104; Mage = 23) compared to controls (n = 83; Mage = 23) showed greater activity in the frontal medial cortex, striatum (eg, nucleus accumbens), precuneus, lateral occipital cortex, and thalamus.72 There were no correlations with cannabis dosage, but there were significant correlations between precuneus/occipital activity and CUDIT scores and cue-induced craving, and the direction of the correlations was positive for the Texan CUD group, and negative for participants from the Netherlands.72 Overall, cannabis cue reactivity was associated with greater brain activity and connectivity in prefrontal-striatal pathways, in some instances exacerbated with more problematic cannabis use and moderated by world region/culture.
Reward Processing fMRI Studies Investigating Differences in Brain Function in Cannabis Users Compared to Controls
The fMRI evidence comparing brain reward function between cannabis users and controls was emerging and largely non-significant. A recent review summarized the evidence from 13 fMRI studies comparing cannabis users (n = 379; Mage = 21) and controls (n = 333; Mage = 21),75 during varied fMRI tasks such as card-guessing and monetary incentive delay.74,75 About half of the fMRI evidence pointed to altered reward anticipatory activity in prefrontal-striatal regions and altered reward receipt activity in prefrontal, limbic, and sensorimotor regions. The remaining findings were non-significant.75 A separate review synthesized findings comparing brain activity between cannabis users (n = 255; Mage = 22) and controls (n = 279; Mage = 22) during the monetary incentive delay fMRI task.74 The review found emerging but inconsistent evidence of altered brain reward function.74 A more recent fMRI study found no significant differences between cannabis users (n = 63; Mage = 22) and controls (n = 62; Mage = 22) during the anticipation or receipt of monetary rewards.56 Overall, the findings on brain reward function in cannabis users were largely non-significant.
Impulsivity fMRI Studies Investigating Differences in Brain Function in Cannabis Users Compared to Controls
A handful of fMRI studies examined brain function during impulsivity-related fMRI tasks (eg, decision-making, response inhibition). In one review of 4 fMRI studies comparing cannabis users (n = 64; Mage = 24) and controls (n = 58; Mage = 22),77 about half of the studies showed group differences in prefrontal activity (eg, ACC, dlPFC and left/right middle frontal-parietal lobe).77 There was emerging evidence of significant correlations between brain activity during inhibitory processing and cannabis use patterns, such as duration of use.96 A recent study demonstrated no group differences between cannabis users (n = 17; Mage = 21) and controls (n = 14; Mage = 24) during the Balloon Analogue Risk fMRI task measuring risky behaviour.76 Overall, there is insufficient and mixed fMRI evidence on brain functional differences during impulsivity-related tasks between cannabis users and controls (eg, decision-making, response inhibition).77
Memory and Working Memory fMRI Studies Investigating Differences in Brain Function in Cannabis Users Compared to Controls
There was consistent evidence of altered brain activity during memory-related and working-memory tasks in cannabis users compared to controls.78–80 One review in cannabis users (n = 267; Mage = 24) and controls (n = 261; Mage = 24) consistently observed group differences in memory-related brain activity, most frequently in frontal and temporal/hippocampal regions.79 There was sparse evidence demonstrating correlations between memory-related brain activity and the age of cannabis use onset.79 A more recent study in cannabis users (n = 100; Mage = 23) and controls (n = 84; Mage = 23) investigated brain activity during a working memory task (N-back).78 The cannabis group displayed reduced working memory-load brain activity in the lateral occipital cortex, right precuneus and left and right medial frontal gyrus.78 An additional review examined the correlates of cannabis abstinence on brain activity in adolescents during various fMRI tasks, including spatial working memory.80 Three studies investigated group differences between cannabis users (n = 69; Mage = 18), cannabis users who were abstinent for at least 25 days (n = 98; Mage = 18) and controls (n = 106; Mage = 18).80 Both abstinent and current cannabis users consistently displayed differences compared to controls in activity in occipital areas and higher-order frontal-parietal regions. No significant differences were found between the abstinent and current cannabis-using groups.80 Overall, the fMRI evidence demonstrated altered brain activity during memory-related and working-memory tasks, particularly in the higher-order cognitive regions and the temporal lobe.78–80 In some cases, these differences in brain activity were associated with cannabis use patterns, including age of onset.79 The emerging evidence from abstinence studies suggests that memory-related differences in brain activity persist beyond at least 25 days of abstinence.80
Resting State Functional Connectivity Studies Investigating Differences in Brain Function in Cannabis Users Compared to Controls
There was consistent evidence that cannabis users, compared to controls, demonstrated altered patterns of resting state functional connectivity between frontal and other cortical regions, in correlation with cannabis use patterns. For instance, one review summarized the evidence from 21 studies in cannabis users (n = 737; Mage = 23) and controls (n = 659; Mage = 23).84 Studies consistently demonstrated greater connectivity between fronto-frontal, fronto-striatal, and fronto-temporal region pairings, and in correlation with cannabis use frequency and cannabis-related problems (eg, MPS and CUDIT scores).84 Additional evidence from three studies revealed that similar patterns of altered resting state functional connectivity within the dorsal salience network and temporal regions persist across cannabis using samples across different ages including youth (n = 70, Mage = 17), adults (n = 180, Mage = 25), and older adults (n = 43, Mage = 68).81–83 Resting state functional connectivity differences in cannabis users correlated with cannabis use patterns, including heavier cannabis use and participants’ perceived benefits and harms of cannabis.81–83 Finally, one individual study showed that young cannabis users (n = 36; Mage = 21) compared to controls (n = 39; Mage = 21), displayed lower resting state functional connectivity within the dorsal attention network after two weeks of monitored abstinence.85 In sum, different frontal-cortical resting state functional connectivity consistently emerged in cannabis users compared to controls and across distinct age groups.
Discussion
This review summarizes the multimodal neuroimaging evidence published over the past 3 decades, on the neurobiology of cannabinoid consumption, during cannabinoid intoxication and in non-intoxicated cannabis consumers. PET and fMRI evidence consistently showed that acute cannabis THC intoxication altered brain fronto-striatal function. Case-control PET and MRS studies consistently showed differences between cannabis users and controls, in glucose metabolism and CB1 receptor density, and the latter reversed after a 4-week abstinence.61 Case-control fMRI studies consistently showed that cannabis users, compared to controls, had different brain function in frontal pathways implicated in higher-order cognitive control.21 There was largely inconsistent MRS evidence on altered neurochemistry (eg, NAA level in frontal regions) in cannabis users. Consistent findings from sMRI and DTI studies demonstrated that cannabis users compared to controls showed lower volumetry in selected brain regions (eg, OFC/hippocampus), and altered white matter microstructure in specific tracts (eg, superior longitudinal fasciculus). There was inconsistent MRS evidence of altered brain metabolism. The evidence from abstinent samples was preliminary and suggests recovery of brain integrity, albeit inconsistently. Emerging evidence suggests that brain changes associated with cannabinoid consumption may be moderated by age, cannabis dependence/CUD, heavier cannabis use, amount of THC and other cannabinoids (eg, CBD) consumed. Overall, the most consistently affected brain metrics across both the evidence on THC intoxication and case–control studies were activity, connectivity and structure of selected brain pathways (eg, OFC, ACC) and lower volumes of selected regions (eg, OFC, hippocampus). Additionally, case–control studies demonstrated altered activity, connectivity, and metabolism of striatal regions implicated in the experience of reward.22
Early findings suggest that some variables may moderate the association between cannabis use and brain integrity. First, age may moderate the relationship between cannabis and brain anatomy but not brain function. Brain functional differences were consistently reported across adult and youth samples,97 but neuroanatomical differences were consistent in adult samples only.29 Perhaps, adult age or prolonged cannabis exposure or both might be required for volumetric differences to occur. Second, it remains unclear if neurobiological changes resolve with abstinence. There was early evidence from individual studies that CB1 receptor density normalized after 4 weeks of monitored abstinence,61 and hippocampal NAA and volumes recovered after periods of abstinence from regular use (eg, 24 years).61,64 However, individual fMRI studies indicated that differences in brain function between cannabis users and controls persist after periods of abstinence (range: 14-to-25 days).80,85 Multimodal neuroimaging research tracking participants who are abstinent from cannabis multiple times and over time is required to confirm if differences in brain integrity between cannabis users and controls dissipate over time and their time course. Third, chronicity of cannabis use patterns and the severity of cannabis use-related problems may moderate alterations in the integrity of prefronto-temporal-striatal pathways. Indeed, group differences emerged more consistently in samples with more chronic exposure to cannabis (eg, greater frequency, longer duration and greater dosage) and with greater cannabis use-related problems (eg, cannabis dependence or CUD). Consistently, prominent neuroscientific theories of addiction propose that neuroadaptations within the addiction neurocircuitry occur with chronic and problematic consumption of substances.22
This review is characterized by methodological limitations, including a focus on published studies in English that may be biased towards positive findings, and the lack of a systematic and meta-analytical approach. The multimodal neuroimaging literature reviewed herein also needs to be considered in light of several methodological limitations. First, most case-control studies were cross-sectional, and the temporality or causality of the observed neurobiological correlates cannot be established. Longitudinal neuroimaging cohort studies represent valuable tools to determine whether brain differences predate or confer risk for cannabis use or if they dissipate with abstinence. Several longitudinal cohort studies could prove useful to this end, including the Connectome Coordination Facility (https://www.humanconnectome.org) and the ABCD study (https://abcdstudy.org/). Second, cannabis use patterns (eg, frequency, occasions, dosage, age of onset, duration), cannabis use-related problems (eg, dependence/CUD), and cannabinoid dosing and products used (eg, THC:CBD ratios, routes of administration) were measured rarely and inconsistently. Thus, it cannot be established how specific patterns of use can be harmful to brain integrity.98,99 Future research should use consistent metrics for cannabis consumption (eg, iCannToolkit)100 and the Standard THC Unit101 to quantify cannabis use patterns and their link to neurobiological indices. Third, females were largely under-represented,102 and the role of sex was largely not accounted for in analyses.102,103 Importantly, males and females show different cannabinoid metabolism, CB1 receptors and endocannabinoids.104 Therefore, the findings to date may not generalize to female participants. Further, males consume greater quantities of cannabis and on more days than females.105–107 In case-control studies, sex was also often statistically unaccounted for. Therefore, it is unresolved whether findings were driven by cannabis use or by heavier consumption in males. Future research should include diverse samples and statistically account for the effect of sex (eg, sex as an interaction term with cannabis use).103 Fourth, the literature failed to consistently collect data, report, and statistically examine the contribution of mental health and the use of other substances to the observed neurobiological correlates. The presence and severity of psychopathology (eg, anxiety, mood, and psychotic disorders) and substances other than cannabis (eg, tobacco and alcohol) are entrenched with cannabis use and also impact brain integrity independently, and in interaction with cannabis use.108–110 Therefore, future work is required to confirm which of the emerging effects are specific to cannabinoids or to confounders. Outstanding gaps also exist in MRS studies during THC intoxication, which limit the understanding of the neurochemical correlates of cannabinoid consumption. Further, there was little evidence on the neurobiological correlates of cannabis withdrawal, and future work is required to confirm if the theorized mechanisms underlying drug withdrawal or problematic substance use apply to cannabinoid consumption, eg, amygdala, habenula involvement.22 New neuroimaging work is required to address the limitations described above and shed some light on the neurobiological correlates of cannabis use to inform the general community, vulnerable people who consume cannabinoids, clinical practitioners, and policy-makers. The reviewed evidence suggests that cannabis use is associated with changes in various metrics of brain integrity, particularly in acute THC intoxication studies and emerging case-control studies where changes in brain integrity correlated with heavier cannabis use and amount of THC use. As such, the early evidence implies that policy-makers could consider the current trend towards decrimininalization and increased availability of cannabis products2 and potency3–5 in light of the relationship between cannabis use and brain integrity. However, more work that addresses the limitations described above is required to fully understand the neurobiological mechanisms of cannabis use to confidently inform public health and policy-making. Further, characterizing the neurobiological correlates of CUD will inform targets for neuromodulation interventions that have been gathering interest in other substance use disorders. These interventions can include fMRI-neurofeedback and repetitive transcranial magnetic stimulation, which have shown promise to modulate brain activity in other substance-using cohorts (eg, alcohol, cocaine, tobacco).111,112 Indeed, novel work has begun to investigate the use of fMRI-neurofeedback to mitigate craving-related brain activity in individuals with CUD.113
Conclusion
We summarized the last thirty years of multimodal neuroimaging evidence on the neural correlates of cannabinoid use. During THC intoxication, consistent evidence showed changes in the function of frontal-striatal regions, including increased cerebral blood flow as measured with PET, as well as decreased rsFC and activity, as per fMRI. There were consistent findings that non-intoxicated cannabis users, compared to controls, showed: changes in glucose metabolism and CB1 receptor density (which reversed with a 4-week abstinence), as measured with PET; changes in brain function as per fMRI, most consistently increases during cue-reactivity; lower OFC/hippocampal volumetry measured by T1-weighted imaging; and lower superior longitudinal fasciculus white matter microstructure. The reviewed evidence came with several methodological limitations. Indeed, the evidence was largely cross-sectional and did not account for potentially confounders entrenched with the consumption of cannabis (eg, mental ill health, polysubstance use). Further, many studies were constrained by small sample sizes, particularly those investigating the brain integrity and cannabis abstinence. As such, the reviewed evidence should be considered as hypothesis-generating rather than hypothesis-testing. More longitudinal multimodal neuroimaging work that controls for confounds is required to confirm the time course and causal mechanisms of cannabinoid consumption. The lack of brain-behavior correlations restricts the behavioural and clinical relevance of the findings; however, alterations were most commonly reported in brain pathways implicated in disinhibition, stress and interoception pathways and in samples with more severe problems with cannabis use and chronicity of use. Therefore, the findings support the notion that severe problems with/chronicity of cannabis use might require the targeting of neurocognitive systems linked to inhibition, stress and affect, via individually tailored psychological approaches, exercise-based interventions, or non-invasive neuromodulation techniques (eg, neurofeedback, TMS).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by an Australian Government Research Training Program (RTP) Scholarship doi.org/10.82133/C42F-K220. Ethan Murphy was supported by Australian Government Research Training Program (RTP) Scholarship (doi.org/10.82133/C42F-K220). Valentina Lorenzetti was supported by an Al and Val Rosenstrauss Senior Research Fellowship (2022–2026) and by a National Health and Medical Research (NHMRC) Investigator Grant (2023–2027 ID: 2016833).
Disclosure
Govinda Poudel was employed by Braincast Neurotechnologies. Hannah Thomson contracts for Syneos Health Learning Solutions, with the Insights and Evidence Generation Team in Patient Insights and Assessment Research (Implementation Science). The funders did not influence the development, results/outcomes of the study, despite the authors’ affiliations with the funder. All other authors have no competing interests in this work.
References
1. UNODC. World drug report 2024. United Nations office on drugs and crime. Available from: https://www.unodc.org/unodc/data-and-analysis/world-drug-report-2024.html.
2. Spindle TR, Bonn-Miller MO, Vandrey R. Changing landscape of cannabis: novel products, formulations, and methods of administration. Curr Opin Psychol. 2019;30:98–17. doi:10.1016/j.copsyc.2019.04.002
3. ElSohly MA, Mehmedic Z, Foster S, Gon C, Chandra S, Church JC. Changes in cannabis potency over the last 2 decades (1995–2014): analysis of current data in the United States. Biol Psychiatry. 2016;79(7):613–619. doi:10.1016/j.biopsych.2016.01.004
4. ElSohly MA, Chandra S, Radwan M, Majumdar CG, Church JC. A comprehensive review of cannabis potency in the United States in the last decade. Biol Psychiatry Cogn Neurosci Neuroimag. 2021;6(6):603–606. doi:10.1016/j.bpsc.2020.12.016
5. Manthey J, Freeman TP, Kilian C, López-Pelayo H, Rehm J. Public health monitoring of cannabis use in Europe: prevalence of use, cannabis potency, and treatment rates. Lancet Reg Health Eur. 2021;10:100227. doi:10.1016/j.lanepe.2021.100227
6. Petrilli K, Ofori S, Hines L, Taylor G, Adams S, Freeman TP. Association of cannabis potency with mental ill health and addiction: a systematic review. Lancet Psychiatry. 2022;9(9):736–750. doi:10.1016/S2215-0366(22)00161-4
7. Baker D, Pryce G, Giovannoni G, Thompson AJ. The therapeutic potential of cannabis. Lancet Neurol. 2003;2(5):291–298. doi:10.1016/S1474-4422(03)00381-8
8. Bloomfield MA, Ashok AH, Volkow ND, Howes OD. The effects of Δ9-tetrahydrocannabinol on the dopamine system. Nature. 2016;539(7629):369–377. doi:10.1038/nature20153
9. Keung MY, Leach E, Kreuser K, et al. Cannabis-induced anxiety disorder in the emergency department. Cureus. 2023;15(4):e38158. doi:10.7759/cureus.38158
10. Solmi M, De Toffol M, Kim JY, et al. Balancing risks and benefits of cannabis use: umbrella review of meta-analyses of randomised controlled trials and observational studies. BMJ. 2023;382:e072348. doi:10.1136/bmj-2022-072348
11. Rogeberg O, Elvik R. The effects of cannabis intoxication on motor vehicle collision revisited and revised. Addiction. 2016;111(8):1348–1359. doi:10.1111/add.13347
12. White MA, Burns NR. The risk of being culpable for or involved in a road crash after using cannabis: a systematic review and meta-analyses. Drug Sci Policy Law. 2021;7:20503245211055381. doi:10.1177/20503245211055381
13. Dellazizzo L, Potvin S, Giguère S, Dumais A. Evidence on the acute and residual neurocognitive effects of cannabis use in adolescents and adults: a systematic meta-review of meta-analyses. Addiction. 2022;117(7):1857–1870. doi:10.1111/add.15764
14. Hoch E, Volkow ND, Friemel CM, Lorenzetti V, Freeman TP, Hall W. Cannabis, cannabinoids and health: a review of evidence on risks and medical benefits. Eur Arch Psychiatry Clin Neurosci. 2025;275(2):281–292. doi:10.1007/s00406-024-01880-2
15. Ghasemiesfe M, Barrow B, Leonard S, Keyhani S, Korenstein D. Association between marijuana use and risk of cancer: a systematic review and meta-analysis. JAMA Network Open. 2019;2(11):e1916318. doi:10.1001/jamanetworkopen.2019.16318
16. Norberg MM, Kavanagh DJ, Olivier J, Lyras S. Craving cannabis: a meta-analysis of self-report and psychophysiological cue—reactivity studies. Addiction. 2016;111(11):1923–1934. doi:10.1111/add.13472
17. Leung J, Chan GC, Hides L, Hall WD. What is the prevalence and risk of cannabis use disorders among people who use cannabis? A systematic review and meta-analysis. Addict Behav. 2020;109:106479. doi:10.1016/j.addbeh.2020.106479
18. Hasin D, Walsh C. Cannabis use, cannabis use disorder, and comorbid psychiatric illness: a narrative review. J Clin Med. 2021;10(1):15. doi:10.3390/jcm10010015
19. Ramaekers JG, Mason NL, Kloft L, Theunissen EL. The why behind the high: determinants of neurocognition during acute cannabis exposure. Nat Rev Neurosci. 2021;22(7):439–454. doi:10.1038/s41583-021-00466-4
20. Glass M, Faull RLM, Dragunow M. Cannabinoid receptors in the human brain: a detailed anatomical and quantitative autoradiographic study in the fetal, neonatal and adult human brain. Neuroscience. 1997;77(2):299–318. doi:10.1016/S0306-4522(96)00428-9
21. Goldstein RZ, Volkow ND. Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nat Rev Neurosci. 2011;12(11):652–669. doi:10.1038/nrn3119
22. Koob GF, Volkow ND. Neurobiology of addiction: a neurocircuitry analysis. Lancet Psychiatry. 2016;3(8):760–773. doi:10.1016/S2215-0366(16)00104-8
23. Kutlu MG, Gould TJ. Effects of drugs of abuse on hippocampal plasticity and hippocampus-dependent learning and memory: contributions to development and maintenance of addiction. Learn Mem. 2016;23(10):515–533. doi:10.1101/lm.042192.116
24. Moulton EA, Elman I, Becerra LR, Goldstein RZ, Borsook D. The cerebellum and addiction: insights gained from neuroimaging research. Addict Biol. 2014;19(3):317–331. doi:10.1111/adb.12101
25. Haber S. Parallel and Integrative processing through the Basal Ganglia reward circuit: lessons from addiction. Biol Psychiatry. 2008;64(3):173–174. doi:10.1016/j.biopsych.2008.05.033
26. Colyer-Patel K, Romein C, Kuhns L, Cousijn J, Kroon E. Recent evidence on the relation between cannabis use, brain structure, and function: highlights and challenges. Curr Addict Rep. 2024;11(3):371–383. doi:10.1007/s40429-024-00557-z
27. Chye Y, Kirkham R, Lorenzetti V, McTavish E, Solowij N, Yücel M. Cannabis, cannabinoids, and brain morphology: a review of the evidence. Biol Psychiatry Cogn Neurosci Neuroimag. 2021;6(6):627–635. doi:10.1016/j.bpsc.2020.07.009
28. Bloomfield MA, Hindocha C, Green SF, et al. The neuropsychopharmacology of cannabis: a review of human imaging studies. Pharmacol Ther. 2019;195:132–161. doi:10.1016/j.pharmthera.2018.10.006
29. Lorenzetti V, Kowalczyk M, Duehlmeyer L, et al. Brain anatomical alterations in young cannabis users: is it all hype? A meta-analysis of structural neuroimaging studies. Cannabis Cannabinoid Res. 2023;8(1):184–196. doi:10.1089/can.2021.0099
30. Lorenzetti V, Chye Y, Silva P, Solowij N, Roberts CA. Does regular cannabis use affect neuroanatomy? An updated systematic review and meta-analysis of structural neuroimaging studies. Eur Arch Psychiatry Clin Neurosci. 2019;269(1):59–71. doi:10.1007/s00406-019-00979-1
31. Berger A. How does it work? Positron emission tomography. BMJ. 2003;326(7404):1449. doi:10.1136/bmj.326.7404.1449
32. Suckling J, Nestor LJ. The neurobiology of addiction: the perspective from magnetic resonance imaging present and future. Addiction. 2017;112(2):360–369. doi:10.1111/add.13474
33. Le Bihan D, Mangin JF, Poupon C, et al. Diffusion tensor imaging: concepts and applications. J Magn Reson Imaging. 2001;13(4):534–546. doi:10.1002/jmri.1076
34. Tognarelli JM, Dawood M, Shariff MI, et al. Magnetic resonance spectroscopy: principles and techniques: lessons for clinicians. J Clin Exp Hepatol. 2015;5(4):320–328. doi:10.1016/j.jceh.2015.10.006
35. Volkow ND, Wang G-J, Telang F, et al. Decreased dopamine brain reactivity in marijuana abusers is associated with negative emotionality and addiction severity. Proc Natl Acad Sci. 2014;111(30):E3149–E3156. doi:10.1073/pnas.1411228111
36. Bossong MG, van Berckel BN, Boellaard R, et al. Delta 9-tetrahydrocannabinol induces dopamine release in the human striatum. Neuropsychopharmacology. 2009;34(3):759–766. doi:10.1038/npp.2008.138
37. O’Leary DS, Block RI, Koeppel JA, et al. Effects of smoking marijuana on focal attention and brain blood flow. Hum Psychopharmacol. 2007;22(3):135–148. doi:10.1002/hup.832
38. O’Leary DS, Block RI, Koeppel JA, et al. Effects of smoking Marijuana on brain perfusion and cognition. Neuropsychopharmacology. 2002;26(6):802–816. doi:10.1016/S0893-133X(01)00425-0
39. Mathew RJ, Wilson WH, Turkington TG, et al. Time course of tetrahydrocannabinol-induced changes in regional cerebral blood flow measured with positron emission tomography. Psychiatry Res Neuroimaging. 2002;116(3):173–185. doi:10.1016/s0925-4927(02)00069-0
40. O’Leary DS, Block RI, Flaum M, et al. Acute marijuana effects on rCBF and cognition: a PET study. NeuroReport. 2000;11(17):3835–3840. doi:10.1097/00001756-200011270-00047
41. Volkow ND, Gillespie H, Mullani N, et al. Brain glucose metabolism in chronic marijuana users at baseline and during marijuana intoxication. Psychiatry Res. 1996;67(1):29–38. doi:10.1016/0925-4927(96)02817-x
42. Lorenzetti V, Gaillard A, Thomson D, Englund A, Freeman TP. Effects of cannabinoids on resting state functional brain connectivity: a systematic review. Neurosci Biobehav Rev. 2023;145:105014. doi:10.1016/j.neubiorev.2022.105014
43. Wall MB, Freeman TP, Hindocha C, et al. Individual and combined effects of cannabidiol and Δ9-tetrahydrocannabinol on striato-cortical connectivity in the human brain. J Psychopharmacol. 2022;36(6):732–744. doi:10.1177/02698811221092506
44. Wall MB, Pope R, Freeman TP, et al. Dissociable effects of cannabis with and without cannabidiol on the human brain’s resting-state functional connectivity. J Psychopharmacol. 2019;33(7):822–830. doi:10.1177/0269881119841568
45. Grimm O, Löffler M, Kamping S, et al. Probing the endocannabinoid system in healthy volunteers: cannabidiol alters fronto-striatal resting-state connectivity. Eur Neuropsychopharmacol. 2018;28(7):841–849. doi:10.1016/j.euroneuro.2018.04.004
46. Skumlien M, Freeman TP, Hall D, et al. The effects of acute cannabis with and without cannabidiol on neural reward anticipation in adults and adolescents. Biol Psychiatry Cogn Neurosci Neuroimag. 2023;8(2):219–229. doi:10.1016/j.bpsc.2022.10.004
47. Freeman TP, Pope RA, Wall MB, et al. Cannabis dampens the effects of music in brain regions sensitive to reward and emotion. Int J Neuropsychopharmacol. 2018;21(1):21–32. doi:10.1093/ijnp/pyx082
48. van Hell HH, Jager G, Bossong MG, et al. Involvement of the endocannabinoid system in reward processing in the human brain. Psychopharmacology. 2012;219(4):981–990. doi:10.1007/s00213-011-2428-8
49. Bossong MG, Jager G, van Hell HH, et al. Effects of Δ9-tetrahydrocannabinol administration on human encoding and recall memory function: a pharmacological fMRI study. J Cogn Neurosci. 2012;24(3):588–599. doi:10.1162/jocn_a_00156
50. Bhattacharyya S, Fusar-Poli P, Borgwardt S, et al. Modulation of mediotemporal and ventrostriatal function in humans by Delta9-tetrahydrocannabinol: a neural basis for the effects of Cannabis sativa on learning and psychosis. Arch Gen Psychiatry. 2009;66(4):442–451. doi:10.1001/archgenpsychiatry.2009.17
51. Borgwardt SJ, Allen P, Bhattacharyya S, et al. Neural basis of Δ-9-Tetrahydrocannabinol and Cannabidiol: effects during response inhibition. Biol Psychiatry. 2008;64(11):966–973. doi:10.1016/j.biopsych.2008.05.011
52. Bossong MG, Jansma JM, van Hell HH, et al. Effects of Δ9-Tetrahydrocannabinol on human working memory function. Biol Psychiatry. 2012;71(8):693–699. doi:10.1016/j.biopsych.2012.01.008
53. Mato S, Chevaleyre V, Robbe D, Pazos A, Castillo PE, Manzoni OJ. A single in-vivo exposure to delta 9THC blocks endocannabinoid-mediated synaptic plasticity. Nat Neurosci. 2004;7(6):585–586. doi:10.1038/nn1251
54. Mason NL, Theunissen EL, Hutten NR, et al. Reduced responsiveness of the reward system is associated with tolerance to cannabis impairment in chronic users. Addict Biol. 2021;26(1):e12870. doi:10.1111/adb.12870
55. Zaytseva Y, Horáček J, Hlinka J, et al. Cannabis-induced altered states of consciousness are associated with specific dynamic brain connectivity states. J Psychopharmacol. 2019;33(7):811–821. doi:10.1177/0269881119849814
56. Skumlien M, Mokrysz C, Freeman TP, et al. Neural responses to reward anticipation and feedback in adult and adolescent cannabis users and controls. Neuropsychopharmacology. 2022;47(11):1976–1983. doi:10.1038/s41386-022-01316-2
57. Spindle TR, Kuwabara H, Eversole A, et al. Brain imaging of cannabinoid type I (CB1) receptors in women with cannabis use disorder and male and female healthy controls. Addict Biol. 2021;26(6):e13061. doi:10.1111/adb.13061
58. Da Silva T, Hafizi S, Watts JJ, et al. In vivo imaging of translocator protein in long-term cannabis users. JAMA Psychiatry. 2019;76(12):1305–1313. doi:10.1001/jamapsychiatry.2019.2516
59. Wiers CE, Shokri-Kojori E, Wong CT, et al. Cannabis abusers show hypofrontality and blunted brain responses to a stimulant challenge in females but not in males. Neuropsychopharmacology. 2016;41(10):2596–2605. doi:10.1038/npp.2016.67
60. Ceccarini J, Kuepper R, Kemels D, van Os J, Henquet C, Van Laere K. [18F]MK-9470 PET measurement of cannabinoid CB1 receptor availability in chronic cannabis users. Addict Biol. 2015;20(2):357–367. doi:10.1111/adb.12116
61. Hirvonen J, Goodwin R, Li C-T, et al. Reversible and regionally selective downregulation of brain cannabinoid CB 1 receptors in chronic daily cannabis smokers. Mol Psychiatry. 2012;17(6):642–649. doi:10.1038/mp.2011.82
62. Sevy S, Smith GS, Ma Y, et al. Cerebral glucose metabolism and D2/D3 receptor availability in young adults with cannabis dependence measured with positron emission tomography. Psychopharmacology. 2008;197(4):549–556. doi:10.1007/s00213-008-1075-1
63. Lorenzetti V, Solowij N, Yucel M. The role of cannabinoids in neuroanatomic alterations in cannabis users. Biol Psychiatry. 2016;79(7):E17–E31. doi:10.1016/j.biopsych.2015.11.013
64. Yücel M, Lorenzetti V, Suo C, et al. Hippocampal harms, protection and recovery following regular cannabis use. Transl Psychiatry. 2016;6(1):e710–e710. doi:10.1038/tp.2015.201
65. Ishrat S, Levey DF, Gelernter J, Ebmeier K, Topiwala A. Association between cannabis use and brain structure and function: an observational and Mendelian randomisation study. BMJ Ment Health. 2024;27(1):e301065. doi:10.1136/bmjment-2024-301065
66. Maleki S, Hendrikse J, Richardson K, et al. White matter alterations associated with chronic cannabis use disorder: a structural network and fixel-based analysis. Transl Psychiatry. 2024;14(1):429. doi:10.1038/s41398-024-03150-0
67. Robinson EA, Gleeson J, Arun AH, et al. Measuring white matter microstructure in 1457 cannabis users and 1441 controls: a systematic review of diffusion-weighted MRI studies. Front Neuroimag. 2023;2:1129587. doi:10.3389/fnimg.2023.1129587
68. Fenzl M, Backens M, Bodea S, et al. Impact of cannabis use on brain metabolism using 31P and 1H magnetic resonance spectroscopy. Neuroradiology. 2023;65(11):1631–1648. doi:10.1007/s00234-023-03220-y
69. Blest-Hopley G, O’Neill A, Wilson R, et al. Adolescent-onset heavy cannabis use associated with significantly reduced glial but not neuronal markers and glutamate levels in the hippocampus. Addict Biol. 2020;25(6):e12827. doi:10.1111/adb.12827
70. Watts JJ, Garani R, Da Silva T, Lalang N, Chavez S, Mizrahi R. Evidence that cannabis exposure, abuse, and dependence are related to glutamate metabolism and glial function in the anterior cingulate cortex: a 1H-magnetic resonance spectroscopy study. Front Psychiatry. 2020;11:746. doi:10.3389/fpsyt.2020.00764
71. Sneider JT, Mashhoon Y, Silveri M. A review of magnetic resonance spectroscopy studies in Marijuana using adolescents and adults. Addict Res Ther. 2013;S4(010). doi:10.4172/2155-6105.S4-010
72. Kroon E, Kuhns L, Cousijn J, Filbey F. Cannabis cue-reactivity in cannabis use disorder: diverging evidence in two distinct cannabis cultures. J Psychiatr Res. 2024;179:341–350. doi:10.1016/j.jpsychires.2024.09.030
73. Sehl H, Terrett G, Greenwood L-M, et al. Patterns of brain function associated with cannabis cue-reactivity in regular cannabis users: a systematic review of fMRI studies. Psychopharmacology. 2021;238(10):2709–2728. doi:10.1007/s00213-021-05973-x
74. Beyer E, Poudel G, Antonopoulos S, Thomson H, Lorenzetti V. Brain reward function in people who use cannabis: a systematic review. Front Behav Neurosci. 2024;17:1323609. doi:10.3389/fnbeh.2023.1323609
75. Skumlien M, Langley C, Lawn W, et al. The acute and non-acute effects of cannabis on reward processing: a systematic review. Neurosci Biobehav Rev. 2021;130:512–528. doi:10.1016/j.neubiorev.2021.09.008
76. Raymond DR, Paneto A, Yoder KK, et al. Does chronic cannabis use impact risky decision-making: an examination of fMRI activation and effective connectivity? Front Psychiatry. 2020;11:599256. doi:10.3389/fpsyt.2020.599256
77. Wrege J, Schmidt A, Walter A, et al. Effects of cannabis on impulsivity: a systematic review of neuroimaging findings. Curr Pharm Des. 2014;20(13):2126–2137. doi:10.2174/13816128113199990428
78. Kroon E, Kuhns L, Colyer-Patel K, Filbey F, Cousijn J. Working memory-related brain activity in cannabis use disorder: the role of cross-cultural differences in cannabis attitudes. Addict Biol. 2023;28(6):e13283. doi:10.1111/adb.13283
79. Blest-Hopley G, Giampietro V, Bhattacharyya S. A systematic review of human neuroimaging evidence of memory-related functional alterations associated with cannabis use complemented with preclinical and human evidence of memory performance alterations. Brain Sci. 2020;10(2):102. doi:10.3390/brainsci10020102
80. Blest-Hopley G, Giampietro V, Bhattacharyya S. Regular cannabis use is associated with altered activation of central executive and default mode networks even after prolonged abstinence in adolescent users: results from a complementary meta-analysis. Neurosci Biobehav Rev. 2019;96:45–55. doi:10.1016/j.neubiorev.2018.10.026
81. Watson KK, Bryan AD, Thayer RE, Ellingson JM, Skrzynski CJ, Hutchison KE. Cannabis use and resting state functional connectivity in the aging brain. Front Aging Neurosci. 2022;14:804890. doi:10.3389/fnagi.2022.804890
82. Kroon E, Toenders YJ, Kuhns LN, Cousijn J, Filbey F. Resting state functional connectivity in dependent cannabis users: the moderating role of cannabis attitudes. Drug Alcohol Depend. 2024;256:111090. doi:10.1016/j.drugalcdep.2024.111090
83. Ertl N, Lawn W, Mokrysz C, et al. Associations between regular cannabis use and brain resting-state functional connectivity in adolescents and adults. J Psychopharmacol. 2023;37(9):904–919. doi:10.1177/02698811231189441
84. Thomson H, Labuschagne I, Greenwood LM, et al. Is resting-state functional connectivity altered in regular cannabis users? A systematic review of the literature. Psychopharmacology. 2022;239(5):1191–1209. doi:10.1007/s00213-021-05938-0
85. Harris JC, Wallace AL, Thomas AM, Wirtz HG, Kaiver CM, Lisdahl KM. Disrupted resting state attentional network connectivity in adolescent and young adult cannabis users following two-weeks of monitored abstinence. Brain Sci. 2022;12(2):287. doi:10.3390/brainsci12020287
86. Gruber SA, Silveri MM, Dahlgren MK, Yurgelun-Todd D. Why so impulsive? White matter alterations are associated with impulsivity in chronic marijuana smokers. Exp Clin Psychopharmacol. 2011;19(3):231–242. doi:10.1037/a0023034
87. Gruber SA, Sagar KA, Dahlgren MK, Racine M, Lukas SE. Age of onset of marijuana use and executive function. Psychol Addict Behav. 2012;26(3):496. doi:10.1037/a0026269
88. Orr JM, Paschall CJ, Banich MT. Recreational marijuana use impacts white matter integrity and subcortical (but not cortical) morphometry. NeuroImage Clin. 2016;12:47–56. doi:10.1016/j.nicl.2016.06.006
89. Cousijn J, Toenders YJ, van Velzen LS, Kaag AM. The relation between cannabis use, dependence severity and white matter microstructure: a diffusion tensor imaging study. Addict Biol. 2022;27(1):e13081. doi:10.1111/adb.13081
90. Chang L, Cloak C, Yakupov R, Ernst T. Combined and independent effects of chronic marijuana use and HIV on brain metabolites. J Neuroimmune Pharmacol. 2006;1(1):65–76. doi:10.1007/s11481-005-9005-z
91. Cowan RL, Joers JM, Dietrich MS. N-acetylaspartate (NAA) correlates inversely with cannabis use in a frontal language processing region of neocortex in MDMA (Ecstasy) polydrug users: a 3 T magnetic resonance spectroscopy study. Pharmacol Biochem Behav. 2009;92(1):105–110. doi:10.1016/j.pbb.2008.10.022
92. Silveri MM, Jensen JE, Rosso IM, Sneider JT, Yurgelun-Todd DA. Preliminary evidence for white matter metabolite differences in marijuana-dependent young men using 2D J-resolved magnetic resonance spectroscopic imaging at 4 Tesla. Psychiatry Res Neuroimaging. 2011;191(3):201–211. doi:10.1016/j.pscychresns.2010.10.005
93. Sung Y-H, Carey PD, Stein DJ, et al. Decreased frontal N-acetylaspartate levels in adolescents concurrently using both methamphetamine and marijuana. Behav Brain Res. 2013;246:154–161. doi:10.1016/j.bbr.2013.02.028
94. Filbey FM, Schacht JP, Myers US, Chavez RS, Hutchison KE. Marijuana craving in the brain. Proc Natl Acad Sci. 2009;106(31):13016–13021. doi:10.1073/pnas.0903863106
95. Vingerhoets W, Koenders L, Van Den Brink W, et al. Cue-induced striatal activity in frequent cannabis users independently predicts cannabis problem severity three years later. J Psychopharmacol. 2016;30(2):152–158. doi:10.1177/0269881115620436
96. Tapert SF, Schweinsburg AD, Drummond SP, et al. Functional MRI of inhibitory processing in abstinent adolescent marijuana users. Psychopharmacology. 2007;194(2):173–183. doi:10.1007/s00213-007-0823-y
97. Lorenzetti V, Alonso-Lana S, J Youssef G, et al. Adolescent cannabis use: what is the evidence for functional brain alteration? Curr Pharml Des. 2016;22(42):6353–6365. doi:10.2174/1381612822666160805155922
98. Temple EC, Brown RF, Hine DW. The ‘grass ceiling’: limitations in the literature hinder our understanding of cannabis use and its consequences. Addiction. 2011;106(2):238–244. doi:10.1111/j.1360-0443.2010.03139.x
99. Solowij N, Lorenzetti V, Yücel M. Effects of cannabis use on human behavior: a call for standardization of cannabis use metrics. JAMA Psychiatry. 2016;73(9):995–996. doi:10.1001/jamapsychiatry.2016.1329
100. Lorenzetti V, Hindocha C, Petrilli K, et al. The international cannabis toolkit (iCannToolkit): a multidisciplinary expert consensus on minimum standards for measuring cannabis use. Addiction. 2022;117(6):1510–1517. doi:10.1111/add.15702
101. Freeman TP, Lorenzetti V. ‘Standard THC units’: a proposal to standardize dose across all cannabis products and methods of administration. Addiction. 2019;115(7):1207–1216. doi:10.1111/add.14842
102. Sterling E, Pearl H, Liu Z, Allen JW, Fleischer CC. Demographic reporting across a decade of neuroimaging: a systematic review. Brain Imaging Behav. 2022;16(6):2785–2796. doi:10.1007/s11682-022-00724-8
103. Lorenzetti V, Christensen E, Valentino R. Understanding sex differences in substance use and substance use disorders: a moving target. J Addict Med. 2025;10:1097. doi:10.1097/adm.0000000000001584
104. Calakos KC, Bhatt S, Foster DW, Cosgrove KP. Mechanisms underlying sex differences in cannabis use. Curr Addict Rep. 2017;4(4):439–453. doi:10.1007/s40429-017-0174-7
105. Preston P. Marijuana use as a coping response to psychological strain: racial, ethnic, and gender differences among young adults. Deviant Behav. 2006;27(4):397–421. doi:10.1080/01639620600721353
106. Greaves L, Hemsing N. Sex and gender interactions on the use and impact of recreational cannabis. Int J Environ Res Public Health. 2020;17(2):509. doi:10.3390/ijerph17020509
107. Cuttler C, Mischley LK, Sexton M. Sex differences in cannabis use and effects: a cross-sectional survey of cannabis users. Cannabis Cannabinoid Res. 2016;1(1):166–175. doi:10.1089/can.2016.0010
108. Hasin D, Walsh C. Trends over time in adult cannabis use: a review of recent findings. Curr Opin Psychol. 2021;38:80–85. doi:10.1016/j.copsyc.2021.03.005
109. Yurasek AM, Aston ER, Metrik J. Co-use of alcohol and cannabis: a review. Curr Addict Rep. 2017;4(2):184–193. doi:10.1007/s40429-017-0149-8
110. Hines LA, Freeman TP, Gage SH, et al. Association of high-potency cannabis use with mental health and substance use in adolescence. JAMA Psychiatry. 2020;77(10):1044–1051. doi:10.1001/jamapsychiatry.2020.1035
111. Murphy E, Poudel G, Ganesan S, et al. Real-time fMRI-based neurofeedback to restore brain function in substance use disorders: a systematic review of the literature. Review. Neurosci Biobehav Rev. 2024;165:105865. doi:10.1016/j.neubiorev.2024.105865
112. Mahoney JJ, Hanlon CA, Marshalek PJ, Rezai AR, Krinke L. Transcranial magnetic stimulation, deep brain stimulation, and other forms of neuromodulation for substance use disorders: review of modalities and implications for treatment. J Neurol Sci. 2020;418:117149. doi:10.1016/j.jns.2020.117149
113. Murphy E, Dakhili AH, Ganesan S, et al. CannChange: a protocol for a feasibility study using fMRI-based neurofeedback to change the neurobiology of craving in cannabis use disorder. BMJ Open. 2025;15(8):e105854. doi:10.1136/bmjopen-2025-105854
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.