Back to Journals » Biologics: Targets and Therapy » Volume 15
Virus Neutralization by Human Intravenous Immunoglobulin Against Influenza Virus Subtypes A/H5 and A/H7
Authors Kubota-Koketsu R ![]() , Yunoki M
, Yunoki M ![]() , Okuno Y, Ikuta K
, Okuno Y, Ikuta K
Received 12 November 2020
Accepted for publication 25 March 2021
Published 13 April 2021 Volume 2021:15 Pages 87—94
DOI https://doi.org/10.2147/BTT.S291808
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Doris Benbrook
Ritsuko Kubota-Koketsu,1,2,* Mikihiro Yunoki,3,* Yoshinobu Okuno,4 Kazuyoshi Ikuta1
1Department of Virology, Research Institute for Microbial Diseases, Osaka University, Osaka, Japan; 2Surveillance Section, Research and Production Technology Department, The Research Foundation for Microbial Diseases of Osaka University, Kagawa, Japan; 3Research and Development Division, Japan Blood Products Organization, Tokyo, Japan; 4Kanonji Institute, The Research Foundation for Microbial Diseases of Osaka University, Kagawa, Japan
*These authors contributed equally to this work
Correspondence: Ritsuko Kubota-Koketsu
Department of Viral Infections, Research Institute for Microbial Diseases, Osaka University, 3-1 Yamadaoka, Suita, Osaka, 565-0871, Japan
Tel +1-6-6879-8348
Fax +1-6-6879-8347
Email [email protected]
Mikihiro Yunoki
Research and Development Division, Japan Blood Products Organization, 15F Tamachi Station Tower N, 3-1-1 Shibaura, Minato-ku, Tokyo, 108-0023, Japan
Tel +81-3-6435-6500
Fax +81-3-6435-6274
Email [email protected]
Purpose: Highly pathogenic avian influenza viruses are a threat to human health. Although donor populations have not experienced pandemic, they have been immunized by natural infections and/or vaccinations of influenza viruses such as A/H1N1, A/H3N2, and B. Therefore, it is considered that human intravenous immunoglobulin (IVIG) derived from healthy donors does not include IgG against avian influenza viruses. However, cross-reactivity has not been evaluated yet. In this study, cross-reactivity against the avian influenza virus A/H5N1, A/H7N1, A/H7N2, A/H7N7, A/H7N9, and A/H10N9 was evaluated.
Materials and Methods: Several lots of IVIG derived from healthy donors in Japan were tested for virus neutralization using single- or multi-cycle virus neutralizing (S-VN or M-VN) assays that evaluate the infection-step associated with HA or the infection and propagation steps associated with HA and NA, respectively. In addition, anti-NA activities were evaluated by inhibiting the enzymatic activity in NAI assays.
Results: IVIG lots showed high neutralizing activities against three A/H5N1 strains in M-VN assays, whereas activities in S-VN assays were unstable. In addition, A/H7N2 was also neutralized in S-VN and M-VN assays, with higher activity in M-VN than in S-VN assays. A/H7N1 was neutralized in S-VN and M-VN assays. In contrast, weak or no activity against A/H7N7, A/H7N9, and A/H10N9 was observed in S-VN and M-VN assays. NAI assay results show that IVIG lots had inhibitory activities against N1 and N2; however, N2 activities differed depending on the strain. In contrast, no activities were observed against N7 and N9.
Conclusion: These results suggest that IVIG lots have neutralizing activity against avian influenza viruses during the virus propagation step, except for one strain, although no or weak activity was observed during the infection step.
Keywords: avian influenza virus, A/H5, A/H7, neutralizing, intravenous immunoglobulin
Introduction
Highly pathogenic avian influenza viruses are a threat to human health. These avian influenza viruses circulate in birds worldwide, and most human infections occur through contact with infected birds or contaminated materials. Avian influenza virus infections in humans are reported and the risk is assessed by public health agencies, with several types being described, such as A/H5, A/H7, and A/H9.1 To date, human-to-human infection is considered to be very limited, and the infection route seems to be the close contact that occurs when living in the same home or nursing care.
The blood donor population has been immunized by natural infections and/or vaccinations of influenza viruses such as A/H1N1, A/H3N2, and B. The influenza virus type A/H2N2 has been eradicated,2 and only older populations have anti- A/H2N2 IgG. Therefore, the current donor population has neutralizing IgG against A/H2N2; however, donors who have anti–A/H2N2 IgG will soon retire from the blood donor pool. Intravenous immunoglobulin (IVIG) is manufactured from pooled plasma derived from >10,000 healthy donors. Neutralizing activity against influenza viruses of IVIG lots derived from donors in Japan has been evaluated,3–5 and IVIG lots show hemagglutinin (HA) inhibition (HI) and virus neutralizing (VN) activities against A/H1N1, A/H2N2, A/H3N2, and B viruses. The HI, microneutralization (MN), and neuraminidase inhibition (NAI) activities against pandemic A/H1N1 virus of IVIG lots derived from donors in US, donated before and after pandemic, were also evaluated.6,7 These studies indicated that HI and MN titers of IVIG lots derived from donor plasmas collected after the pandemic have increased compared to lots derived from before the pandemic. In addition, a similar tendency of NAI titers derived from both before and after the pandemic was observed. Regarding avian influenza, a study indicated that IVIG derived from donor plasma collected before the pandemic shows a high HI titer against the seasonal A/H1N1 virus, low titer against pandemic A/H1N1, and very weak titer against A/H5N1. In addition, neuraminidase (N1) activities derived from pandemic A/H1N1 and A/H5N1 are inhibited by the globulin of IVIG.8 Another study suggested that cross-reactive antibodies against avian N1 and seasonal N1 antigen of IVIG lots neutralize the A/H5N1 virus.9 IVIG lots derived from donors in selected areas also show neutralizing activity against A/H5N1 but not against A/H7N7 viruses.10 These studies suggested that some IVIG lots could have cross-reactive antibodies against pandemic influenza viruses, including avian influenza A/HxN1 and A/HxN2 in the future. The neutralizing activities against avian flu of IVIG lots derived from donors in Japan are unknown. In this study, the neutralizing activity against low pathogenic avian influenza viruses as models of highly pathogenic viruses such as A/H5Nx and A/H7Nx of IVIG lots derived from donors in non-pandemic/endemic avian flu regions in Japan was evaluated.
Materials and Methods
Study Design
We aimed to evaluate the VN capacity of IVIG by two different methods. A major neutralizing point in the virus life cycle is the virus attachment and entry to the host cell associated with HA. Other neutralizing points include virus propagation and budding out, which are associated with NA. We tested the neutralizing capacity of IVIG using a single-cycle viral neutralization test (S-VN), and multiple-cycle viral neutralization test (M-VN), based on a previously described method.11,12 S-VN and M–VN assays focus on virus attachment and entry associated with HA and on virus propagation and budding associated with NA, respectively.
Samples and Viruses
This study was approved by the ethical committee of Japan Blood Products Organization, former Benesis Corporation. Human plasma, used as control in experiments, was approved by the Institutional Review Board of Research Institute for Microbial Diseases, Osaka University.
Human intravenous immunoglobulin (IVIG) (Venoglobulin IH 5% I.V. (V–IH); Japan Blood Products Organization, Tokyo, Japan; former the Benesis Corporation, Osaka, Japan) lots derived from Japanese donors, manufactured from 1993 to 2013, were used in this study. V-IH lots derived from donors in US manufactured in 1993 and 1999 were also used. IgG concentration in IVIG was 50 mg/mL at neat dilution, 4–5 times the concentration of healthy human plasma. The influenza virus strains used in this study are summarized in Table 1. Note that, low pathogenic avian influenza viruses were employed as models of highly pathogenic avian influenza viruses such as A/H5N1, A/H7Nx, and A/H10N9. The viruses were examined in biosafety level 2 or 3 facility of Research Institute for Microbial Diseases, Osaka University, with approval from the biosafety committee of the safety management committee on pathogens of Osaka University (Table 1).
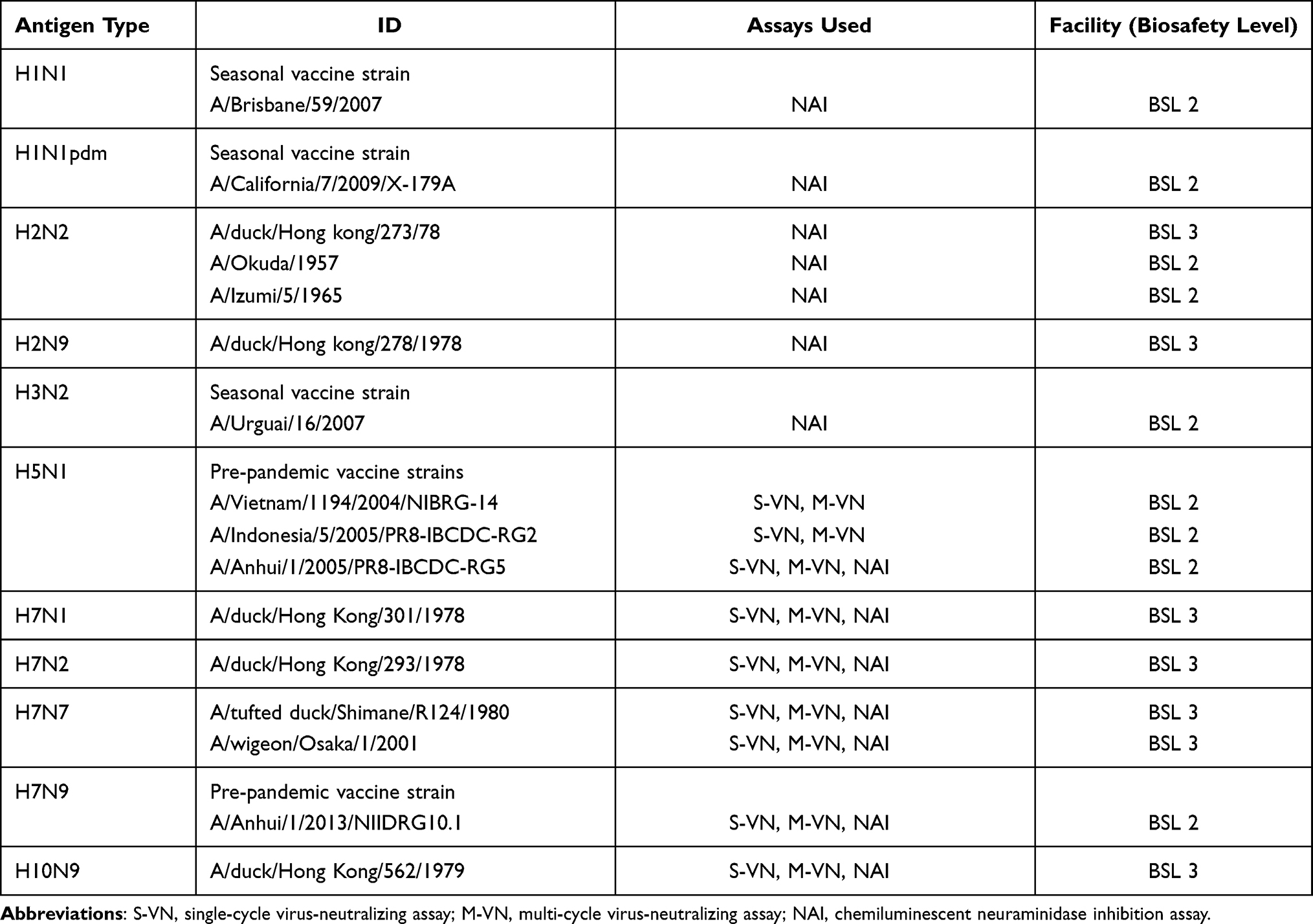 |
Table 1 Influenza Virus Strains Used in This Study |
Methods
The viruses were propagated as described previously.4 Briefly, the viruses were propagated in Madin–Darby canine kidney (MDCK) cells or allantoic cavity of 10 days-old embryonic chicken eggs. Culture supernatants and allantoic fluids were stored at −80 °C prior to use. Viral infectivity (FFU – focus forming unit/mL) was titrated in MDCK cells using an indirect immunofluorescence assay (IFA). For IFA, virus-infected cells were fixed with ethanol and incubated with 1 µg/mL of mouse monoclonal antibody (mMAb) against influenza A NP, C43 (Bio Academia, Osaka, Japan) for 1 h at room temperature. Goat anti-mouse IgG (H+L) Alexa Fluor 488 conjugate (Invitrogen) was used for detection of mMAb binding.
S-VN was carried out as follows. IVIGs were diluted four-fold with minimum essential medium (MEM) and mixed with an equal volume of 100 FFU of influenza viruses. After incubation for 1 h at 37 °C, 30 µL of IVIG-virus mixtures were transferred onto MDCK cells in a 96-well plate and incubated for 1 h at 37 °C. The IVIG-virus mixtures were aspirated from cells, which were washed once with serum-free MEM. Virus culture medium (MEM) supplemented with 2% fetal bovine serum (FBS) was added to each well. After incubation for 16 h at 37 °C, the cells were fixed with ethanol and subjected to IFA. M-VN was carried out as follows. IVIGs were diluted four-fold with MEM and mixed with an equal volume of 100 FFU of influenza viruses. After incubation for 1 h at 37 °C, 50 µL of IVIG-virus mixtures were transferred onto MDCK cells in a 96-well plate and incubated for 1 h at 37 °C. Equal volumes of MEM containing 0.5% of methyl cellulose and 4 µg/mL acetyl trypsin overlay was added to each well and the cells were incubated for 28 h at 37 °C. After washing three times with PBS, the cells were fixed with ethanol and subjected to IFA. Human plasma containing cross-reactive anti-HA or NA antibody and anti-H5N1 mouse monoclonal antibody C179 (1 mg/mL at neat dilution) was used as the positive control,13 and culture medium alone was used as the negative control. The neutralizing titers were determined as the reciprocal of the highest IVIG dilution required to neutralize over 50% of virus. The experiments were conducted in duplicate, and standard error (SE) was calculated from two replicates. Therefore, neutralization was categorized as none or unlikely (≤4 or below the detection limit), probable (8–16), and clear (≥32) in this study.
To evaluate anti-NA activity, strains summarized in Table 1 were used. Chemiluminescent neuraminidase inhibition (NAI) assay was carried out using a commercially available kit, NA-XTD (Applied Biosystems, CA, USA), as recommended by the manufacturer, with minor modifications. Viral NA activity was determined before the NAI assay was performed by serial two-fold dilution of the virus. Viruses were diluted in NA-XTD assay buffer so as to have 40 relative fluorescent units (RFUs). Each well contained 25 µL of diluted IVIG, to which 25 µL of each virus dilution was then added. After incubation for 30 min at 37 °C, 25 µL of NA-XTD substrate was added to each well, and the plate was incubated for 30 min at room temperature. Luminescence was read for 5 min after adding 60 µL of NA-XTD accelerator to each well, using a Grating Microplate Reader SH-9000Lab, (CORONA ELECTRIC, Ibaraki, Japan). NAI activity of IVIG was determined as follows: NAI activity (%) = {(RFUs of virus– RFUs of virus with IVIG)/RFUs of virus} × 100. Higher NAI activity (%) was associated with higher anti-NA activity of the sample. The experiments were conducted in triplicate, and SE was calculated from two replicates.
Results
Neutralizing titers against three strains of A/H5N1 viruses were determined using the S-VN and M-VN assays. IVIG from the manufacturing years of 1993 to 2010 showed none/unlikely, probable, and none/unlikely VN activity against A/Indonesia/05/2005, A/Anhui/01/2005, and A/Vietnam/1194/2004 strains, respectively, in S-VN assay. In contrast, M-VN assay showed clear VN activity of the lots against the virus strains, which was more effective than that of S-VN assay (Figure 1). Neutralizing titers against A/H7N1 (A/duck/Hong Kong/301/1978), A/H7N2 (A/duck/Hong Kong/293/1978), A/H7/N7 (A/tufted duck/Shimane/R124/1980 and A/wigeon/Osaka/1/2001), A/H7N9 (A/Anhui/1/2013/NIIDRG10.1), and A/H10N9 (A/duck/Hong Kong/562/1979) strains were also evaluated. Antigens H7, H10, N7, and N9 were not found in the seasonal influenza vaccine. Only A/H7N1 strain showed clear VN activity in S-VN assay, whereas none/unlikely or probable VN activity was observed with other virus strains. Out of the six strains, A/H7N2 showed clear VN activity in M-VN assay. In contrast, A/H7N7 (A/wigeon/Osaka/1/2001) showed no VN activity. Other virus strains A/H7N1, A/H7N7, A/H7N9, and A/H10N9 showed probable VN activities (Figure 2). These results suggest that IVIG lots also contained cross-reactive IgG with neutralizing activities against the infection and propagation steps. However, the neutralizing effects on A/H5N1, A/H7Nx, or A/H10N9 were not universal in this study (Figure 2).
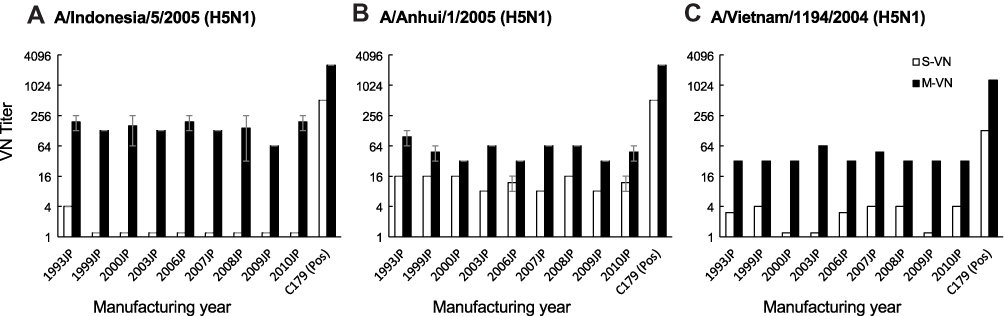 |
Figure 1 Neutralizing activities of IVIG lots against A/H5N1 viruses using S-VN and M-VN assays. (A) A/Indonesia/5/2005/PR8-IBCDC-RG2, (B) A/Anhui/1/2005/PR8-IBCDC-RG5, (C) A/Vietnam/1194/2004/NIBRG-14. Open bar: S-VN; closed bar: M-VN. Error bar: standard error. Abbreviations: IVIG, intravenous immunoglobulin; S-VN, single-cycle virus-neutralizing assay; M-VN, multi-cycle virus-neutralizing assay; Pos, positive control. |
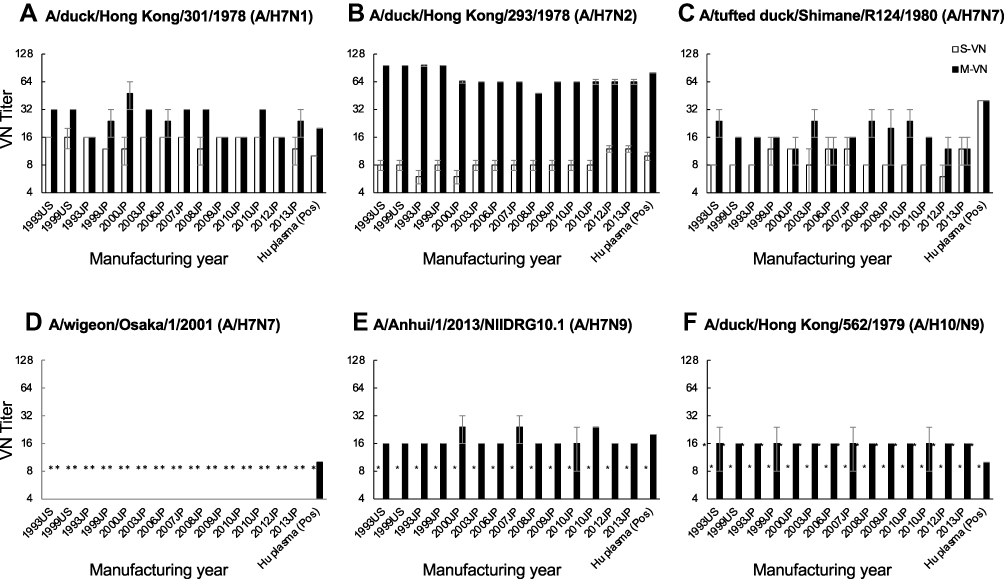 |
Figure 2 Neutralizing activities of IVIG lots against A/H7Nx and A/H10N9 viruses using S-VN and M-VN assays. (A) A/duck/Hong Kong/301/1978 (A/H7N1), (B) A/duck/Hong Kong/293/1978 (A/H7N2), (C) A/tufted duck/Shimane/R124/1980 (A/H7N7), (D) A/wigeon/Osaka/1/2001 (A/H7N7), (E) A/Anhui/1/2013/NIIDRG10.1 (A/H7N9), (F) A/duck/Hong Kong/562/1979 (A/H10/N9). Open bar: S-VN; closed bar: M-VN. Dots: no activity was detected (less than 8). Error bar: standard error. Abbreviations: IVIG, intravenous immunoglobulin; S-VN, single-cycle virus-neutralizing assay; M-VN, multi-cycle virus-neutralizing assay; Pos, positive control. |
Anti-NA activities of IVIG lots were also evaluated. The IVIG lots showed high and stable anti-NA activity against the A/H1N1, A/H5N1, and A/H7N1 strains. These results suggest that IVIG contains anti-N1 IgG (Figure 3A). Notably, IVIG lots showed various anti-NA activities against A/H2N2 (three strains), A/H3N2, and A/H7N2 strains. The result of the A/H3N2 strain, which is used in vaccines, showed a middle range of inhibition. The results of the A/H7N2 strain, which has not spread to humans, also showed a middle range of inhibition. In addition, the results of the A/H2N2 strains that have previously spread in humans showed various inhibiting activities. The activity of anti-N2 was different from that of anti-N1 (Figure 3B). In contrast to the activity against the A/H1N1, A/H5N1, and A/H7N1 strains, the IVIG lots showed low anti-NA activity against A/H2N9, A/H7N7, A/H7N9, and A/H10N9 strains (Figure 3C and D).
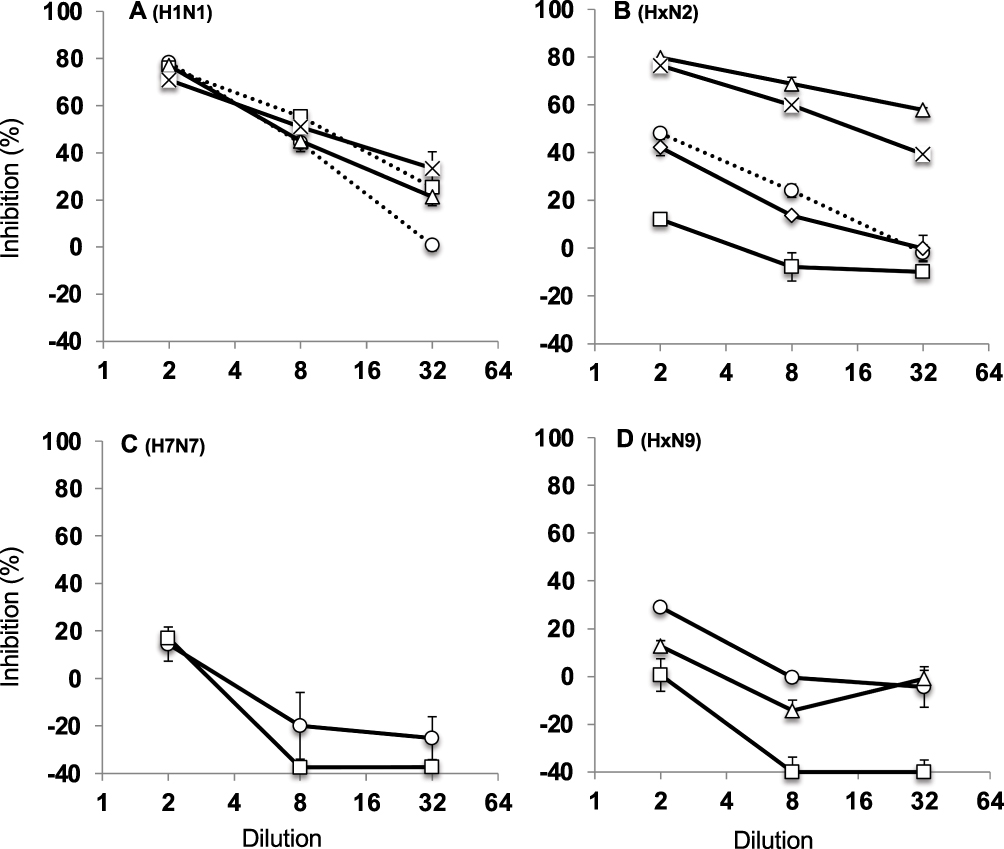 |
Figure 3 Comparison of neuraminidase inhibition activities of IVIG lots against A/H1N1, A/HxH2, A/H7N7, and A/HxN9 viruses. (A) A/HxN1 viruses. Circle: A/California/7/2009/X-179A (A/H1N1pdm); square: A/Brisbane/59/2007 (A/H1N1); triangle: A/Anhui/1/2005/PR8-IBCDC-RG5 (A/H5N1); cross: A/duck/Hong Kong/301/1978 (A/H7N1). (B) A/HxN2 viruses. Circle: A/Urguai/16/2007 (H3N2); square: A/Okuda/1957 (A/H2N2); triangle: A/Izumi/5/1965 (A/H2N2); diamond: A/duck/Hong Kong/293/1978 (A/H7N2); cross: A/duck/Hong kong/273/78 (A/H2N2). (C) A/H7N7 viruses. Circle: A/tufted duck/Shimane/R124/1980; square: A/wigeon/Osaka/1/2001. (D) A/HxN9 viruses. Circle: A/duck/Hong kong/278/1978 (A/H2N9); square: A/Anhui/1/2013/NIIDRG10.1 (A/H7N9); triangle: A/duck/Hong Kong/562/1979 (A/H10N9). Error bar: standard error. Abbreviation: IVIG, intravenous immunoglobulin. |
Discussion
The neutralizing activity of IVIG against A/H5N1, A/H7Nx, and A/H10N9 influenza viruses was evaluated as there are limited reports regarding neutralizing activity of IVIGs against avian influenza virus. The donor population was immunized by vaccination with A/H1, A/H3 and B antigens or by natural infection by seasonal influenza viruses A/H1N1, A/H3/N2, and B viruses. In addition, some older populations were also previously exposed to influenza A/H2N2. However, it is considered that most donor populations have not been exposed to the highly pathogenic avian influenza virus. IVIG is obtained from pooled plasma derived from >10,000 healthy donors. Therefore, IVIG included antibodies against seasonal influenza A/H1N1, A/H2N2, A/H3/N2, and B viruses.4 After the pandemic in 2009, there is still a possibility of outbreak of avian influenza virus in the human population. Transmission of avian influenza A viruses from animals to humans may result from two major two ways: directly from birds or avian influenza A virus-contaminated environment to humans or through an intermediate host, such as a pig.14 Therefore, WHO performed monthly assessments of the transmission of avian Influenza at the human-animal interface.1 In particular, A/H5N1 and A/H7N9 are being continuously monitored.
IVIG products derived from US or European populations show anti-A/H5N1 activity, but no activity against A/H7N7.9 S-VN is considered to evaluate the suppression of attachment and entry of the virus in host cell as a preventive measure, whereas M-VN is expected to evaluate the suppression of infection and proliferation stages as a treatment. However, previous studies did not focus on these differences. In this study, IVIG lots derived from donors in Japan also showed clear anti-A/H5N1 VN activity (with a titer ≥32) in M-VN assays. In contrast, the lots showed none/unlikely or probable (with a titer ≤16) VN activities in S-VN assays. In addition, the IVIG lots showed clear anti-A/H7N1 VN activity of half of tested lots and anti-A/H7N2 VN activities of all lots in M-VN assays, whereas S-VN assays showed probable VN activities. In contrast, IVIG lots contain cross-reactive IgG against H7. Other viruses, namely A/H7N7, A/A7N9, and A/A10N9, which have not been exposed to the donor population, showed probable VN activities against the viruses in S-VN and M-VN assays. Thus, IVIG lots may contain some cross-reactive IgG against H7 depending on the virus strain and NA subtypes. The steric hindrance of anti-head HA antibodies may have inhibited the binding of some anti-stem HA antibodies. Thus, the anti-stem HA antibodies may remain unbound after consumption of the anti-head HA antibodies. Therefore, M-VN assays would also evaluate the activity of the anti-stem HA antibodies. IVIG lots showed higher levels of anti-NA activity against A/H1N1pdm, A/H1N1, A/H5N1, A/H7N1, and some A/H2N2 strains, and moderate anti-NA activity against A/H3N2 and A/H7N2. Notably, a weak anti-NA activity was observed against an A/H2N2 strain. Weak or no anti-NA activity was observed against the strains A/H7N7, A/H2N9, A/H7N9, and A/H10N9. Sialidase activity of neuraminidase of some avian influenza viruses is intact at low pH.15 Therefore, it seems that the obtained results might be false negative since pH of IVIG was 3.9–4.4.
Different VN capacities were observed from the same sub-type virus strains such as A/H5N1 and A/H7N7 (Figures 1 and 2). A similar phenomenon was observed previously when A/Osaka/168/2009 (H1N1 pandemic strain), A/NC/20/99 (vaccine strain before pandemic 2009), and A/Osaka/16/2008 of A/H1N1 showed different VN activities.3 Additionally, different anti-NA activities were observed from strains of the same N2 type (Figure 3). These results suggested that each strain has unique properties, despite the same sub-type. This phenomenon seems to result from complex factors such as host species, receptor binding avidity of HA, NA activity, and propagation characteristics in host cell. If a pandemic were to occur in the future, VN activity against the virus should be evaluated, because of the difference in VN activity against each strain of the virus.
The results suggested that IVIG lots contained cross-reacting IgG against H5 or H7 antigen, depending on the virus strain, and contained IgG against N1 or N2 antigen. Highly pathogenic avian influenza virus causes a fatal infectious disease, capable of a pandemic. It is believed that the IVIG lots contain no-effective IgG against avian influenza viruses.10 This study revealed that IVIG is only weakly effective or ineffective against viral attachment and entry in the host cell but is effective against virus propagation in some strains with N1 and N2 antigens. The results suggest a potential treatment against the highly pathogenic influenza virus infections; however, further investigation is needed, particularly to evaluate the differences between low pathogenic strains and high pathogenic viruses, such as structure and alignment of neutralizing epitope.
Conclusion
The results of this study suggest that, although donors have not experienced an epidemic or pandemic of highly pathogenic avian influenza in humans, IVIG lots have a neutralizing activity by M-VN against some avian influenza viruses of the N1 and some N2 types, whereas have no neutralizing activity by S-VN. However, the neutralizing activities against other avian influenza viruses, such as A/H7Nx, were dependent on strains. The results of this study suggest that IVIG has the ability to suppress the growth of avian influenza virus infections with N1 or N2 antigens, but it is premature to determine whether it has a therapeutic effect against highly pathogenic avian influenza viruses. Further studies to evaluate the mechanism of activity during the entire viral life cycle using highly pathogenic viruses and the therapeutic effect are needed.
Acknowledgments
The authors thank Dr. Takeru Urayama and Mr. Shoji Ideno (Japan Blood Products Organization) and Dr. Hironori Yoshii (The Research Foundation for Microbial Diseases of Osaka University) for technical support.
Author Contributions
All authors have made a significant contribution to the work reported, that is, in the conception, study design, execution, acquisition of data, analysis and interpretation, and in drafting, revising or critically reviewing the article. All authors have given approval to the final version to be published; have agreed on the journal to which the article is being submitted; and have agreed to be accountable for all aspects of the work.
Disclosure
This study was funded by the Benesis Corporation (Currently Japan Blood Products Organization) and was conducted in part as a collaborative research project between Osaka University, The Research Foundation for Microbial Diseases of Osaka University, and the Benesis Corporation. Current affiliations are as follows: RK Koketsu, Department of Viral Infections, Research Institute for Microbial Diseases, Osaka University; M Yunoki, Japan Blood Products Organization; Y Okuno and K Ikuta, Osaka Institute of Public Health. The authors declare no competing interests.
References
1. World Health Organization. Influenza at the human-animal interface. Summary and assessment; 2021. Available from: https://www.who.int/influenza/human_animal_interface/HAI_Risk_Assessment/en/.
2. Kilbourne ED. Influenza pandemics of the 20th century. Emerg Infect Dis. 2006;12:9–14. doi:10.3201/eid1201.051254
3. Yunoki M, Kubota-Koketsu R, Urayama T, et al. Significant neutralizing activity of human immunoglobulin preparations against pandemic 2009 H1N1. Br J Haematol. 2010;148:953–955. doi:10.1111/j.1365-2141.2009.08016.x
4. Kubota-Koketsu R, Yunoki M, Okuno Y, Ikuta K. Significant neutralizing activities against H2N2 influenza A viruses in human intravenous immunoglobulin lots manufactured from 1993 to 2010. Biologics. 2012;6:245–247. doi:10.2147/BTT.S33495
5. Onodera H, Urayama T, Hirota K, et al. Neutralizing activities against seasonal influenza viruses in human intravenous immunoglobulin. Biologics. 2017;11:23–30. doi:10.2147/BTT.S123831
6. Hohenadl C, Wodal W, Kerschbaum A, et al. Hyperimmune intravenous immunoglobulin containing high titers of pandemic H1N1 hemagglutinin and neuraminidase antibodies provides dose-dependent protection against lethal virus challenge in SCID mice. Virol J. 2014;11:70. doi:10.1186/1743-422X-11-70
7. Kreil TR, Mc Vey JK, Lei LS, et al. Preparation of commercial quantities of a hyperimmune human intravenous immunoglobulin preparation against an emerging infectious disease: the example of pandemic H1N1 influenza. Transfusion. 2012;52:803–809. doi:10.1111/j.1537-2995.2011.03347.x
8. Rockman S, Lowther S, Camuglia S, et al. Intravenous immunoglobulin protects against severe pandemic influenza infection. EBioMedicine. 2017;19:119–127. doi:10.1016/j.ebiom.2017.04.010
9. Pichyangkul S, Jongkaewwattana A, Thitithanyanont A, et al. Cross-reactive antibodies against avian influenza virus A (H5N1). Emerg Infect Dis. 2009;15:1537–1539. doi:10.3201/eid1509.090471
10. Sullivan JS, Selleck PW, Downton T, et al. Heterosubtypic anti-avian H5N1 influenza antibodies in intravenous immunoglobulins from globally separate populations protect against H5N1 infection in cell culture. J Mol Genet Med. 2009;3:217–224. doi:10.4172/1747-0862.1000038
11. Okuno Y, Tanaka K, Baba K, Maeda A, Kunita N, Ueda S. Rapid focus reduction neutralization test of influenza A and B viruses in microtiter system. J Clin Microbiol. 1990;28:1308–1313. doi:10.1128/JCM.28.6.1308-1313.1990
12. Ohshima N, Kubota-Koketsu R, Iba Y, Okuno Y, Kurosawa Y. Two types of antibodies are induced by vaccination with A/California/2009 pdm virus: binding near the sialic acid-binding pocket and neutralizing both H1N1 and H5N1 viruses. PLoS One. 2014;9:e87305. doi:10.1371/journal.pone.0087305
13. Ohshima N, Iba Y, Kubota-Koketsu R, Asano Y, Okuno Y, Kurosawa Y. Naturally occurring antibodies in humans can neutralize a variety of influenza virus strains, including H3, H1, H2, and H5. J Virol. 2011;85(21):11048–11057. doi:10.1128/JVI.05397-11
14. Centers for Disease Control and Prevention. Transmission of avian influenza A viruses between animals and people; 2015. Available from: https://www.cdc.gov/flu/avianflu/virus-transmission.htm#:~:text=Avian%20influenza%20A%20viruses%20may,host%2C%20such%20as%20a%20pig.
15. Takahashi T, Suzuki Y, Nishinaka D, et al. Duck and human pandemic influenza A viruses retain sialidase activity under low pH conditions. J Biochem. 2001;130:279–283. doi:10.1093/oxfordjournals.jbchem.a002983
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.