Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 13
Variations in the Wnt/β-Catenin Pathway Key Genes as Predictors of Cervical Cancer Susceptibility
Authors Wang B ![]() , Wang M
, Wang M ![]() , Li X, Yang M, Liu L
, Li X, Yang M, Liu L
Received 6 February 2020
Accepted for publication 22 April 2020
Published 20 May 2020 Volume 2020:13 Pages 157—165
DOI https://doi.org/10.2147/PGPM.S248548
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Bingqi Wang, Min Wang, Xianping Li, Min Yang, Lei Liu
Department of Laboratory Medicine, The Second Xiangya Hospital, Central South University, Changsha 410011, Hunan, People’s Republic of China
Correspondence: Min Wang
Department of Laboratory Medicine, The Second Xiangya Hospital, Central South University, 139 Middle Renmin Road, Changsha 410011, Hunan, People’s Republic of China
Tel/Fax +86 132 9869 7558
Email [email protected]
Background: Cervical cancer is the fourth most common and fatal tumor among women worldwide. The Wnt/β-catenin signaling pathway was etiologically involved in the cervical cancer model. Herein, we aimed to investigate whether germline genetic variations within the Wnt/β-catenin pathway can be genetic risk factors of cervical cancer.
Patients and Methods: A total of 305 samples (147 patients, 158 controls) were included. Eight genetic variations located in APC (rs454886), GSK3β (rs3755557), CTNNB1 (rs11564475, rs1798802, rs3864004, rs2293303, and rs4135385), and TCF7L2 (rs7903146) were genotyped via Sanger sequencing. The χ 2 test and non-conditional logistic regression were used in the single-locus analysis. Gene–gene interactions and haplotype construction in case–control samples were performed by the GMDR method and Haploview software, respectively.
Results: The frequency of CTNNB1 rs1798802 GA+AA genotype was significantly lower in cervical cancer patients adjusted for age (OR=0.626, 95% CI=0.398– 0.984). The mutant alleles of rs3864004 (A) and rs2293303 (T) located in CTNNB1 showed 1.513 (1.038– 2.207), and 1.654 (1.020– 2.683) fold increased risk of cervical cancer, respectively. Haplotype analysis showed no association between haplotypes of the CTNNB1 gene and cervical cancer risk. No significant contribution of interactions among genes in the Wnt pathway was identified.
Conclusion: We concluded that the genetic variants in the CTNNB1 gene might contribute to the development of cervical cancer.
Keywords: cervical cancer, Wnt/β-catenin signaling pathway, single nucleotide polymorphism, APC, GSK3β, CTNNB1, TCF7L2
Corrigendum for this paper has been published
Introduction
Cervical cancer is a common and fatal malignancy among women. In 2018, cervical cancer ranked as the fourth female diagnosed cancer and the fourth leading cause of cancer worldwide.1 In China, the incidence and mortality of cervical cancer were 12.0 and 3.4 per 100,000 women, respectively.2 Refractory infection with human papillomavirus (HPV) is the primary cause of cervical cancer. The viral E6 and E7 gene products can bind and inhibit tumor suppressor proteins p53 and pRb, respectively, which interrupt cell-cycle control and induce malignant transformation.3 According to the epidemiological investigation, the HPV infection rate might be as high as 90% in sexually active women. However, most of the viral infections are self-limited and cleared within several months; only a very few persist and ultimately contribute to cancer.4 Hence, HPV infection could merely be regarded as a fundamental step in cellular transformation, while an active contribution of other host or environmental alterations that are needed at the molecular level to represent as a “second hit” in the progression to malignancy.5
Multiple signaling pathways play an essential role in tumor development, such as the MAPK signaling pathway,6 JAK-STAT signaling pathway,7 Mitochondrial and PI3K/AKT signaling pathways,8,9 etc. One of the candidate molecular mechanisms involved in the cervical cancer model is the Wnt signaling pathway. Three Wnt signaling pathways have been characterized, and the heavily studied is the Wnt/β-catenin signaling pathway (also named the canonical Wnt signaling pathway). In mammals, the canonical Wnt signaling pathway mainly includes three steps: Wnt signals transduction in the membrane, stability of β-catenin in the cytoplasm, and Wnt target genes activation in the nucleus.10 The core of the Wnt/β-catenin pathway is the cytoplasm-nucleus translocation of β-catenin protein. Its stability is controlled by the so-called β-catenin destruction complex, composed of the scaffold protein Axin, the tumor suppressor adenomatous polyposis coli (APC), and two kinases casein kinase 1 (CK1) and glycogen synthase kinase 3β (GSK3β). In the Wnt “off” state, β-catenin is maintained at low levels in the cytoplasm. Mechanistically, free β-catenin is efficiently phosphorylated by GSK3β, recognized by β-TrCP, and targeted for proteasomal degradation.11 In the Wnt “on” state, the interactions between Wnt ligands and the receptors recruit Dishevelled (Dvl) proteins to the plasma membrane, subsequently inducing the docking of Axin and thereby deconstructing the destruction complex. Without the destruction complex, β-catenin enters the nucleus. There, it displaces Groucho from the T cell factor/lymphoid enhancer factor, upregulates Wnt target genes expression, and induces a change of multiple cellular processes.12
The Wnt/β-catenin pathway is a highly conserved cascade regulating embryonic development, tissue regeneration, stem cell maintenance, and adult homeostasis.13 Mutated Wnt pathway key components are tightly associated with multiple growth-related pathologies and with cancer. In more than half of all cancer cases, such as colorectal cancer,14 breast cancer,15 melanoma,16 and leukemia,17 aberrant Wnt signaling pathway has been reported. Also, a large number of studies have suggested that variants of Wnt signaling pathway genes contributed to cancer susceptibility.18–20 Genetic variants of CTNNB1 could be used as indicators of gastric cancer risk and prognosis.21 The APC polymorphism was a significant risk factor for colon cancer.20 In recent years, some studies have verified that Wnt signaling attribute to cervical cancer pathology at multiple levels, including tumor proliferation, invasion, metastasis, angiogenesis, and therapeutic resistance.22 Nevertheless, insufficient evidence suggests how various Wnt pathway genetic polymorphisms affect the risk of cervical cancer. Stated thus, we hypothesized that Wnt pathway gene polymorphisms were equally associated with cervical cancer risk. We conducted a case–control study and screened potentially functional variants within APC, GSK3β, CTNNB1, and TCF7L2 genes to determine whether they can be genetic risk factors of cervical cancer.
Patients and Methods
Study Population
A total of 305 people was enrolled from the Second Xiangya Hospital and then categorized into two groups. The inclusion criteria for the case group were: (a) histopathological biopsy and imaging examination to determine cervical cancer; (b) no other malignant tumors; (c) no family history of cancer; (d) no clear occupational carcinogenic factors exposure history. The clinical criterion of the International Federation of Gynecology and Obstetrics (FIGO) was carried out for tumor staging. We recorded the clinic-pathological parameters of all patients, consisting of age, menopause age, HPV status, and menopausal status. All the control participants were included in the study when they satisfied the following inclusion criteria: (a) no history of cancer or cancer-related diseases; (b) typical physical examination results after diagnostic exclusion of cancer; (c) no cervical intraepithelial neoplasia. All participants were genetically unrelated Han Chinese.
SNP Selection, DNA Extraction and Genotyping
SNPs in the Wnt/β-catenin pathway genes were selected based on the following standards: (a) located in the core genes of the Wnt/β-catenin pathway; (b) which have been reported in previous literature; (c) the minor allele frequency was higher than 0.05. We first chosen 102 SNP loci based on the above principles. Finally, a total of eight widely studied SNP candidates were included in our study (APC rs454886, GSK3β rs 3755557, CTNNB1 rs11564475, rs1798802, rs3864004, rs4135385, rs2293303, and TCF7L2 rs7903146).
Approximately 5 mL of venous blood were collected in vacutainers containing ethylene diamine tetraacetic acid (EDTA) from all subjects enrolled in the study. Genomic DNA was extracted from peripheral blood leukocytes using the phenol-chloroform method and then stored at −80°C before genotyping. The A260/A280 of the purified DNA was tested between 1.8 and 2.0, which indicated that no external DNA contamination. The isolated samples were sent to the Huada Gene Corporation for genotyping. The polymorphic regions of rs11564475 A/G, rs1798802 G/A, rs3864004 G/A, rs4135385 G/A, rs2293303 C/T, rs454886 G/A, rs3755557 T/A and rs7903146 C/T were amplified by PCR. SNPs genotyping was conducted using Sanger sequencing. Around 10% of the samples were randomly used to reconfirm the results.
Statistical Analysis
In the case–control study, differences in age between cases and control subjects were compared by the Student’s t-test. The Stata software (version 14.0) was used to check genotype and allelic frequencies for deviation from Hardy–Weinberg equilibrium (HWE). SNPs were excluded from the analysis if they did not obey HWE (P<0.05). Linkage disequilibrium (LD) analysis and haplotype construction was performed using Haploview software (version 4.2). The extent of LD was calculated by the standardized D’. Those haplotypes with a frequency under 3% were ignored. Genetic models used in association testing were the co-dominant, dominant, and recessive models. The reference variant was the homozygous form of the most frequent allele. χ2 or Fishers’ exact test was performed to compare genotype and allele distributions between two groups. We conducted the non-conditional logistic regression to assess the effect of individual SNP in different groups with adjustment for age. Chi-square (χ2) values, odds ratios (OR), 95% confidence intervals (95% CI), and P-value were carried out using SPSS software (version 24.0). All P-value were two-sided, and we considered a P-value of 0.05 as significant. The GMDR v0.9 program estimated all possible SNP interactions, cross-validation consistency, testing balance accuracy, and permutation testing with 1000 replications.
Results
Clinical Characteristics of Subjects
The age of patients ranged from 21 to 67 years, while the control group was between 21 and 71 years. The mean age of cases and controls was 46.0±9.0 and 45.6±9.8, respectively. No statistical difference was found between patients and controls in terms of age. The relevant experimental information, including histology, staging, HPV status of the cervical cancer subjects, has also been described. Menopause age, first birth age, menopausal status, family cancer history, and early symptoms were documented as medical history. The early stages of cervical cancer may be completely free of symptoms. Abnormal vaginal bleeding, contact bleeding (one most common form being bleeding after sexual intercourse), or (rarely) a vaginal mass was common in the case group. Most of the patients were HPV positive, and the dominant common high-risk HPV type was HPV-16. Also, it was observed HPV co-infection.
HWE Analysis
Table 1 presented the genomic information about the SNPs we studied and the results of HWE tests. The distributions of all eight SNPs in our study were consistent with the HWE (P>0.05), suggesting these study groups were from the same Mendelian population.
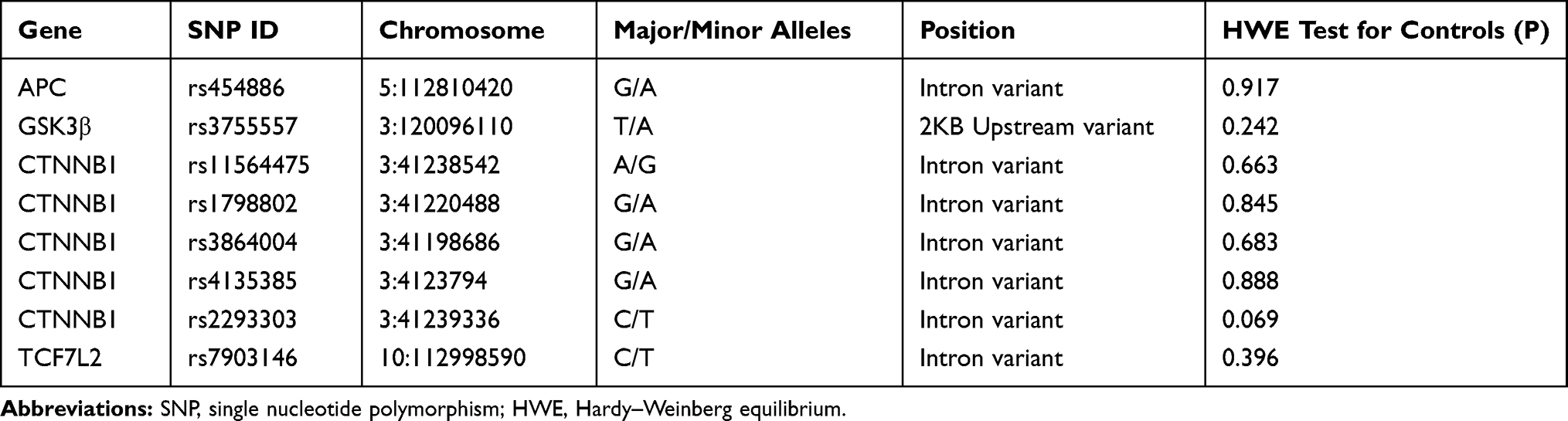 |
Table 1 SNP Information of Genes Analyzed in This Study |
Association Analysis of Eight Polymorphisms with Cervical Cancer
The independent correlations of all selected SNPs with risk of cervical cancer are concluded in Tables 2 and 3. Non-conditional logistic regression analysis with adjustment for age was used to determine the link between SNPs and cancer risk under three genetic models. As shown in Table 2, the risk of cervical cancer for people carrying GA+AA genotype in the CTNNB1 rs1798802 locus was 0.626 times than that of the GG genotype after adjustment with age (OR=0.626, 95% CI=0.398–0.984, P=0.042). Compared with the healthy control group, the rs3864004 A allele (OR=1.513, 95% CI=1.038–2.207, P=0.031) of the CTNNB1 gene showed significant differences in the cervical cancer group. The minor allele (T) for rs2293303 was positively associated with a higher risk for cervical cancer development (OR=1.654, 95% CI=1.020–2.683, P=0.040, Table 3). We did not find any statistical significance in the distributions of genetic models or alleles between the two groups for other individual SNPs.
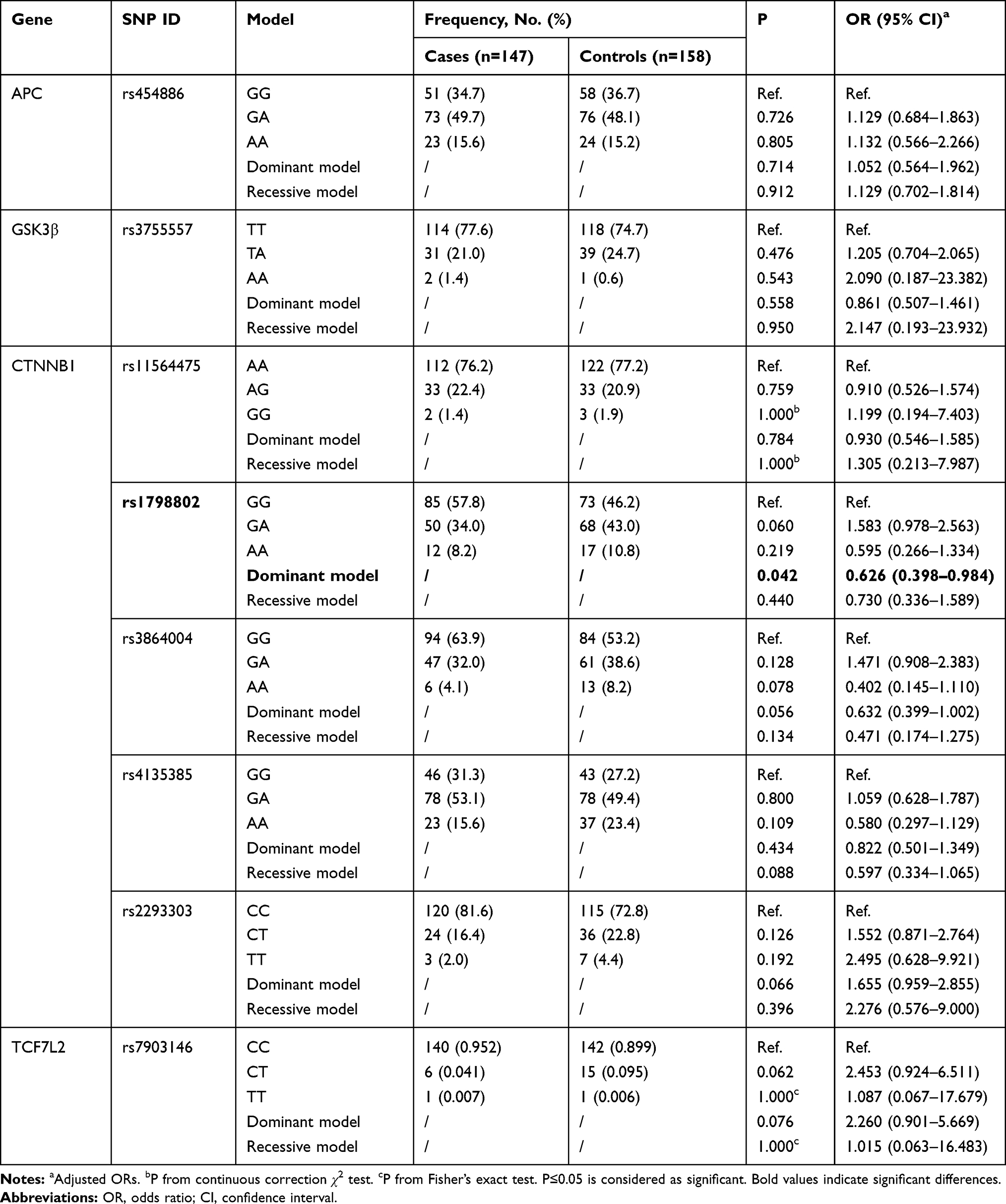 |
Table 2 Association Between Genotypes Frequencies and Cervical Cancer Risk |
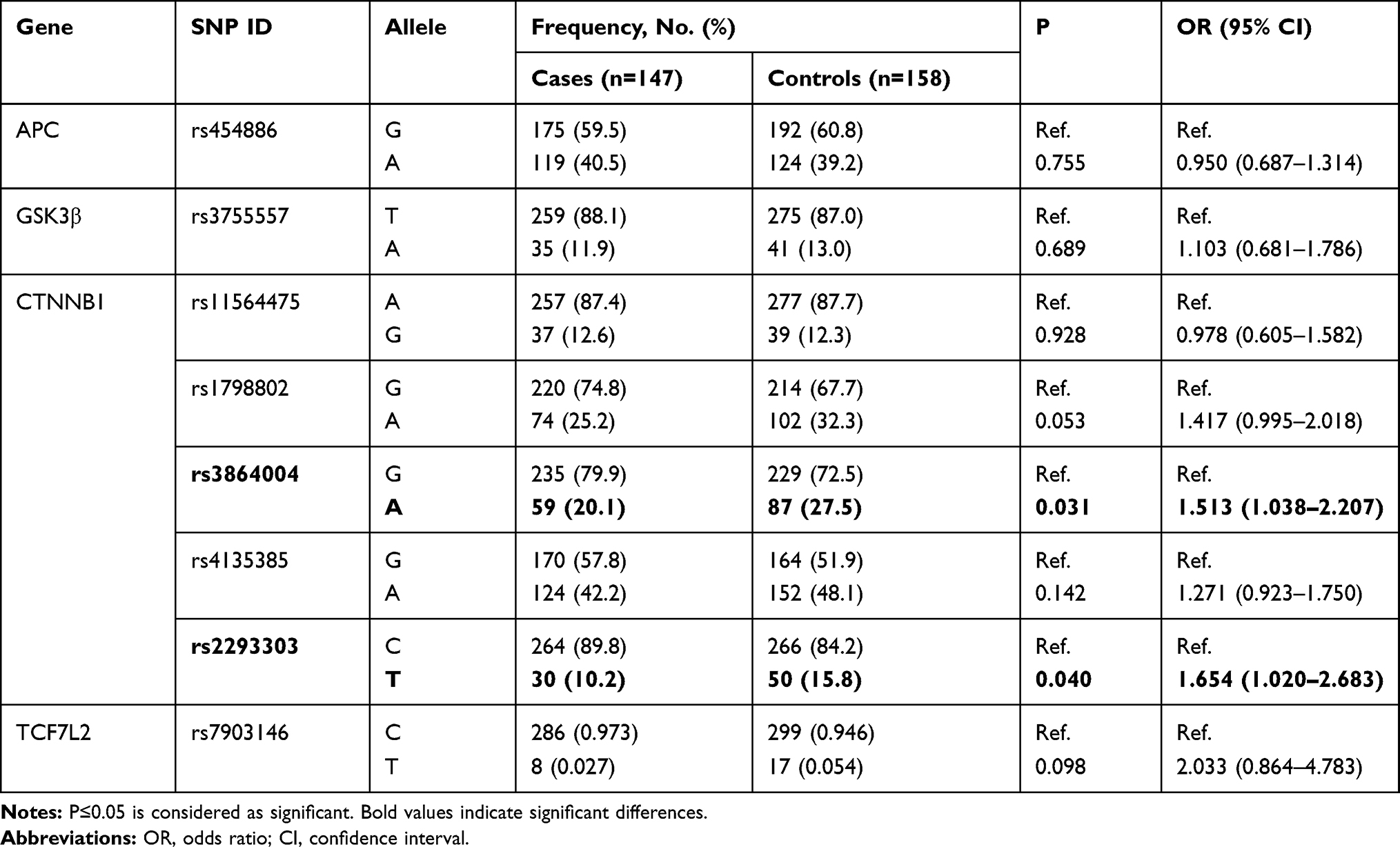 |
Table 3 Association Between Alleles Frequencies and Cervical Cancer Risk |
Haploview Analysis
The presence of a LD structure was analyzed for the tested CTNNB1 SNPs, and their haplotypes were constructed (Table 4, Figure 1). There was linkage disequilibrium between five SNPs, and the LD coefficient (D’) values ranged from 0.980 to 1.000. Only one LD block was identified, yielding six haplotypes with an estimated frequency >3%. No significant association between the haplotypes from rs3864004-rs1798802-rs4135385-rs11564475-rs2293303 in cases and controls was observed (P>0.05, Pc>0.05, Table 5).
 |
Table 4 The Pairwise Linkage Disequilibrium of SNPs in the CTNNB1 Gene |
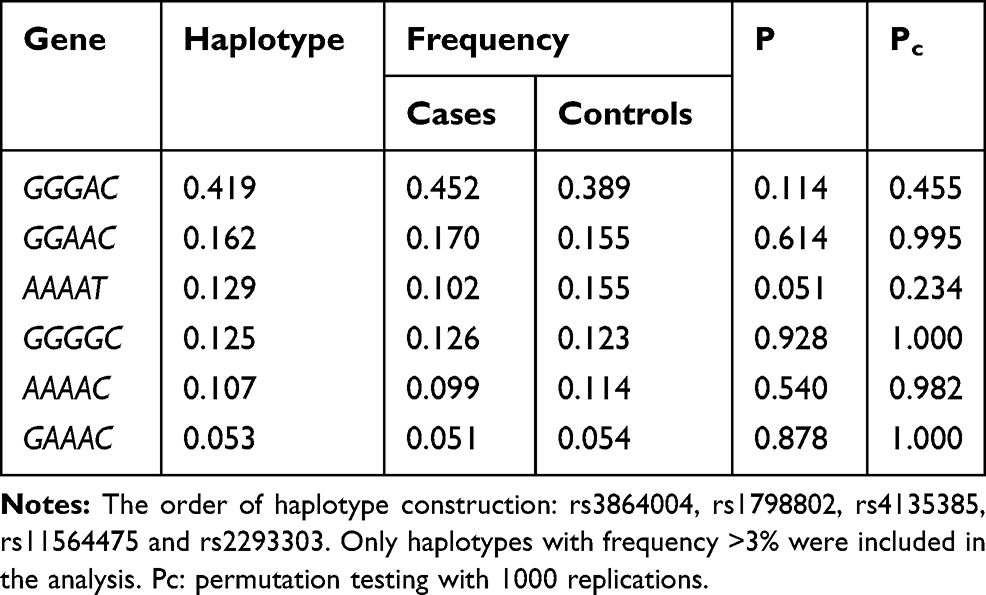 |
Table 5 Haplotype Analysis of the CTNNB1 Gene |
 |
Figure 1 The linkage disequilibrium estimates in the CTNNB1 gene. Notes: Each square represents the LD analysis results between two SNP loci. The color ranges from white to red, representing the linkage degree from low to high. |
SNP–SNP Interaction
We employed the GMDR analysis to evaluate the effect of SNP–SNP interaction among all enrolled SNPs on cervical cancer susceptibility, and age was selected as the covariate. Table 6 and Figure 2 summarize the results from GMDR analysis for each number of loci. One-locus model CTNNB1 rs1798802 had a maximum cross-validation consistency (8/10) and a maximum testing accuracy (0.5464). While all the models about gene–gene interaction among the eight loci on cervical cancer risk were no significant (P>0.05).
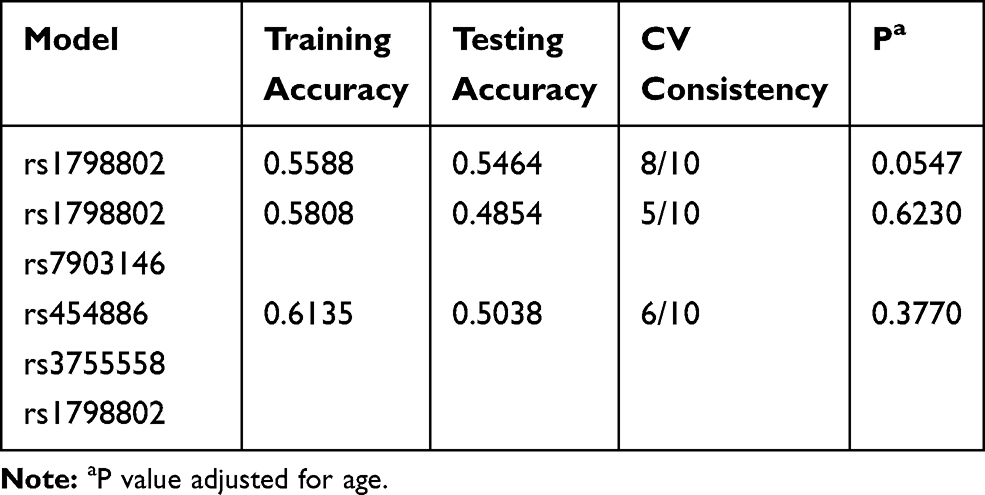 |
Table 6 The Best SNP–SNP Interaction Models Determined by GMDR |
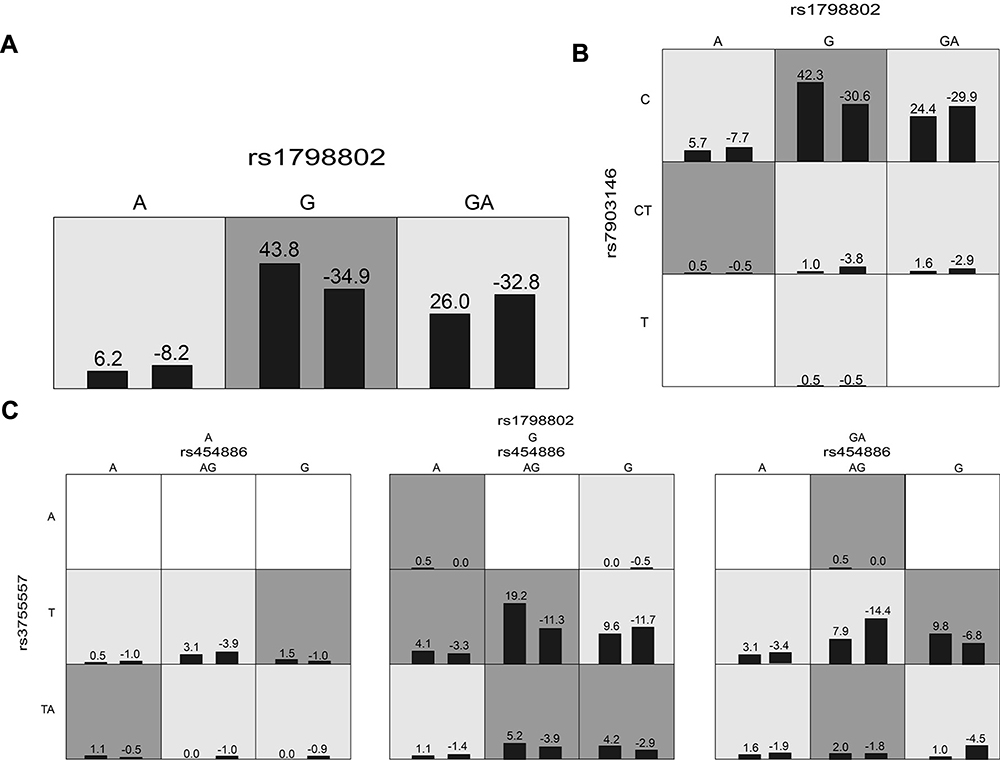 |
Figure 2 The best three models for predicting the occurrence of cervical cancer (A) one-locus model of rs1798802. (B) Two-loci model of rs1798802-rs7903146. (C) Three-loci model of rs454886-rs3755557-rs1798802. A grid represented an interactive combination. Dark gray and light gray grids presented the high- and low-risk factor combinations, respectively. Left bars within each grid represented case while the right bars represented control. |
Discussion
The Wnt/β-catenin signaling pathway is a critical regulatory system in the control of cell proliferation, migration, cell adhesion, and organogenesis.13 In the present study, eight SNPs from the APC, GSK3β, CTNNB1, and TCF7L2 were genotyped in a Chinese Han population. Three of them in the CTNNB1 gene (rs1798802, rs3864004, rs2293303) were observed statistically significant differences. There was no association between cervical cancer and either common haplotypes or SNP–SNP interactions among any of the genes associated with the Wnt/β-catenin signaling pathway. Altogether, our results provided pieces of evidence that Wnt pathway gene polymorphisms may alter gene functions or affect gene activity in patients with cervical cancer. These variations may act as novel genetic markers for cervical cancer screening and diagnosis.
Aberrant activation of the Wnt/β-catenin signaling pathway is a frequent event in human cancers. CTNNB1 is a gene coding β-catenin protein, dually functioning in the coordination of cell adhesion and gene transcription.21 In the single-locus analysis, we identified that mutant alleles of rs3864004 (A) and rs2293303 (T) might be risk factors for cervical cancer. Individuals carrying CTNNB1 rs1798802 dominant genetic model were harder to develop cervical cancer. These findings above were consistent with previous studies in other diseases. However, some of the results have been contradictory. A meta-analysis performed by Li et al mentioned that rs1798802 and rs2293303 polymorphisms were associated with overall cancer risk,23 as the common conclusion we have got. Genotype GA and allele A of rs3864004 were referred to hepatocellular carcinoma in the Chinese Han population.24 Still, Hu et al have reported that rs3864004 A allele decreased tuberculosis risk.25 In various types of cancers, deregulation of the Wnt/β-catenin signaling pathway occurs almost invariably via mutations in APC, dysfunction of GSK3β, or mutations of β-catenin itself.15 Nevertheless, variations in CTNNB1 have rarely been detected in cervical cancer. Aberrant protein level or abnormal distribution of β-catenin were frequently found in cervical carcinogenesis.26 It has been mentioned that all the significantly associated SNPs in the intron region, so we speculated that location in the intron region might influence gene transcription, shearing, altering CTNNB1 expression level, and exert effects on the downstream-cancer related molecules.27 Therefore, direct experiments focused on validating the functional consequences and exact roles of these SNPs are needed in the future.
As members of the destruction complex, APC and GSK3β have been implicated in regulating the canonical Wnt signaling. APC encodes a tumor suppressor protein and acts as an antagonist of the Wnt signaling pathway. GSK3β is a serine-threonine kinase phosphorylated β-catenin and creating a binding site for ubiquitination and rapid degradation. With the “Wnt” on, Wnt signals turn off β-catenin destruction, and the resulting stabilized β-catenin enters the nucleus and binds to TCF7L2 to modulate Wnt target gene transcription.28 The results for the case–control population indicated that three SNPs in APC (rs454886), GSK3β (3755557), and TCF7L2 (rs7903146) were not significantly associated with cervical cancer. A similar study of breast cancer reported the minor allele of rs454886 was related to an increased risk of breast cancer,29 which was inconsistent with ours. Zheng et al proposed that there was no significant association in the frequencies of alleles and genotypes of rs3755557 polymorphic sites with gastric carcinoma.30 In the Caucasian population, Silbernagel and colleagues have examined rs7903146 and noted that carriers with the risk alleles in rs7903146 increase type 2 diabetes susceptibility.31 The difference between our studies and other results could be attributed to various potential reasons, such as different disease types, different tumor stages, ethnicity, environment, sample size, and so on. Although we did not report any associations of the above three SNPs with cervical cancer, their functions cannot be ignored. Genotyping more SNP sites in these genes, detecting the alterations in protein level, exploring the epigenetic modification of gene expression still requires more research.
To date, the majority of single-locus analysis focuses on the identification of individual SNP effects, which are not sufficient to explain the complexity of disease causality. Haplotypes and SNP–SNP interactions are considered more informative in the etiology of complicated diseases. Herein, we conducted haplotypes analysis from different combinations of CTNNB1 five SNPs. The distribution frequency of all six haplotypes in the case group was not different from the control group. Conversely, Kim et al conducted investigated the correlation between polymorphisms of rs3755557 with the risk of hepatocellular carcinoma. They revealed a significantly higher frequency of CTNNB1 A-A haplotype at rs3864004 and rs4135385 positions in patients with hepatocellular carcinoma than in chronic HBV carriers.18 Furthermore, no significant SNP–SNP interaction has been found in our study, while using SNP–SNP interaction to identify potentially pathogenic genes of cervical cancer analyze the exact etiology more accurately.
Cervical cancer is mostly a preventable disease if early detection is achieved. Notably, before translating molecular epidemiology findings into clinical use, there is a need to consider inter-ethnic differences in genetic backgrounds due to the existence of heterogeneity.32 Because of the ethnic diversity in genetic polymorphisms, the same genotype may show different clinical significance in diverse populations. The impact of environmental factors should also be considered.
Conclusions
In summary, we took a pathway-based candidate gene approach to verify the association between cervical cancer predisposition with genetic variants in the pathway genes. Our data highlighted CTNNB1 rs1798802, rs2293303, and rs3864004 are associated with cervical cancer susceptibility. It’s significant to identify “hot spot” genetic factors that may contribute to the development of cervical cancer. Confirmation of our findings in multi-ethnic and multi-region can provide definitive evidence for the use of Wnt/β-catenin pathway gene polymorphisms as screening biomarkers in cervical cancer early detection.
Abbreviations
HPV, human papillomavirus; APC, adenomatous polyposis coli; CK1, casein kinase 1; GSK3β, glycogen synthase kinase 3β; Dvl, Dishevelled; FIGO, Federation of Gynecology and Obstetrics; EDTA, ethylene diamine tetraacetic acid; HWE, Hardy–Weinberg equilibrium; LD, Linkage disequilibrium; OR, odds ratios; 95% CI, 95% confidence intervals.
Ethics Approval and Informed Consent
The study complied with the Declaration of Helsinki Principles and was approved by the ethics committee of The Second Xiangya Hospital, Central South University (Changsha, China). All participants provided written informed consent.
Funding
This study was supported by the Natural Science Foundation of China [No. 81470133], the Natural Science Foundation of Hunan Province [Nos. 2019JJ40426, 2019JJ40418], and the Fundamental Research Funds for the Central South University [No.2017zzts844].
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Chen D, Gyllensten U. Lessons and implications from association studies and post-GWAS analyses of cervical cancer. Trends Genet. 2015;31(1):41–54. doi:10.1016/j.tig.2014.10.005
3. Zur Hausen H. Papillomaviruses causing cancer: evasion from host-cell control in early events in carcinogenesis. J Natl Cancer Inst. 2000;92(9):690–698. doi:10.1093/jnci/92.9.690
4. Smith JS, Lindsay L, Hoots B, et al. Human papillomavirus type distribution in invasive cervical cancer and high-grade cervical lesions: a meta-analysis update. Int J Cancer. 2007;121(3):621–632. doi:10.1002/ijc.22527
5. Perez-Plasencia C, Duenas-Gonzalez A, Alatorre-Tavera B. Second hit in cervical carcinogenesis process: involvement of wnt/beta catenin pathway. Int Arch Med. 2008;1(1):10. doi:10.1186/1755-7682-1-10
6. Peluso I, Yarla NS, Ambra R, Pastore G, Perry G. MAPK signalling pathway in cancers: olive products as cancer preventive and therapeutic agents. Semin Cancer Biol. 2019;56:185–195. doi:10.1016/j.semcancer
7. Groner B, von Manstein V. Jak Stat signaling and cancer: opportunities, benefits and side effects of targeted inhibition. Mol Cell Endocrinol. 2017;451:1–14. doi:10.1016/j.mce.2017.05.033
8. Zhang F, Zhang YY, Sun YS, et al. Asparanin A from Asparagus officinalis L. Induces G0/G1 Cell Cycle Arrest and Apoptosis in Human Endometrial Carcinoma Ishikawa Cells via Mitochondrial and PI3K/AKT Signaling Pathways. J Agric Food Chem. 2020;68(1):213–224. doi:10.1021/acs.jafc.9b07103
9. Zhang YY, Zhang F, Zhang YS, et al. Mechanism of Juglone-Induced Cell Cycle Arrest and Apoptosis in Ishikawa Human Endometrial Cancer Cells. J Agric Food Chem. 2019;67(26):7378–7389. doi:10.1021/acs.jafc.9b02759
10. Galluzzi L, Spranger S, Fuchs E, Lopez-Soto A. WNT Signaling in Cancer Immunosurveillance. Trends Cell Biol. 2019;29(1):44–65. doi:10.1016/j.tcb.2018.08.005
11. Duchartre Y, Kim YM, Kahn M. The Wnt signaling pathway in cancer. Crit Rev Oncol Hematol. 2016;99:141–149. doi:10.1016/j.critrevonc.2015.12.005
12. Valenta T, Hausmann G, Basler K. The many faces and functions of beta-catenin. EMBO J. 2012;31(12):2714–2736. doi:10.1038/emboj.2012.150
13. Nusse R, Clevers H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell. 2017;169(6):985–999. doi:10.1016/j.cell.2017.05.016
14. Kinzler KW, Nilbert MC, Su LK, et al. Identification of FAP locus genes from chromosome 5q21. Science. 1991;253(5020):661–665. doi:10.1126/science.1651562
15. Webster MT, Rozycka M, Sara E, et al. Sequence variants of the axin gene in breast, colon, and other cancers: an analysis of mutations that interfere with GSK3 binding. Genes Chromosomes Cancer. 2000;28(4):443–453. doi:10.1002/1098-2264(200008)28:4<443::aid-gcc10>3.3.co;2-4.
16. Webster MR, Weeraratna AT. A Wnt-er migration: the confusing role of beta-catenin in melanoma metastasis. Sci Signal. 2013;6(268):pe11. doi:10.1126/scisignal.2004114
17. Luis TC, Ichii M, Brugman MH, Kincade P, Staal FJ. Wnt signaling strength regulates normal hematopoiesis and its deregulation is involved in leukemia development. Leukemia. 2012;26(3):414–421. doi:10.1038/leu.2011.387
18. Kim SS, Cho HJ, Lee HY, et al. Genetic polymorphisms in the Wnt/beta-catenin pathway genes as predictors of tumor development and survival in patients with hepatitis B virus-associated hepatocellular carcinoma. Clin Biochem. 2016;49(10–11):792–801. doi:10.1016/j.clinbiochem.2016.01.025
19. Rosales-Reynoso MA, Saucedo-Sariñana AM, Contreras-Díaz KB, et al. Genetic Polymorphisms in APC, DVL2, and AXIN1 Are Associated with Susceptibility, Advanced TNM Stage or Tumor Location in Colorectal Cancer. Tohoku J Exp Med. 2019;249(3):173–183. doi:10.1620/tjem.249.173
20. Parine NR, Azzam NA, Shaik J, et al. Genetic variants in the WNT signaling pathway are protectively associated with colorectal cancer in a Saudi population. Saudi J Biol Sci. 2019;26(2):286–293. doi:10.1016/j.sjbs.2018.05.018
21. Wang S, Tian Y, Wu D, et al. Genetic variation of CTNNB1 gene is associated with susceptibility and prognosis of gastric cancer in a Chinese population. Mutagenesis. 2012;27(6):623–630. doi:10.1093/mutage/ges027
22. Yang M, Wang M, Li X, et al. Wnt signaling in cervical cancer? J Cancer. 2018;9(7):1277–1286. doi:10.7150/jca.22005
23. Li Y, Zhang F, Yang D. Comprehensive assessment and meta-analysis of the association between CTNNB1 polymorphisms and cancer risk. Biosci Rep. 2017;37:6. doi:10.1042/BSR20171121
24. Li QM, Zhang FQ, Li YF, Xian QJ, Zhang YQ, Li P. Influence of polymorphisms in the Wnt/beta-catenin pathway genes on hepatocellular carcinoma risk in a Chinese Han population. Medicine. 2017;96(12):e6127. doi:10.1097/MD.0000000000006127
25. Hu X, Zhou J, Chen X, et al. Pathway Analyses Identify Novel Variants in the WNT Signaling Pathway Associated with Tuberculosis in Chinese Population. Sci Rep. 2016;6:28530. doi:10.1038/srep28530
26. Rodríguez-Sastre MA, González-Maya L, Delgado R, et al. Abnormal distribution of E-cadherin and beta-catenin in different histologic types of cancer of the uterine cervix. Gynecol Oncol. 2005;97(2):330–336. doi:10.1016/j.ygyno.2004.12.062
27. Lv Z, Xu Q, Yuan Y. A systematic review and meta-analysis of the association between long non-coding RNA polymorphisms and cancer risk. Mutat Res. 2017;771:1–14. doi:10.1016/j.mrrev.2016.10.002
28. Stamos JL, Weis WI. The beta-catenin destruction complex. Cold Spring Harb Perspect Biol. 2013;5(1):a007898. doi:10.1101/cshperspect.a007898
29. Wang X, Goode EL, Fredericksen ZS, et al. Association of genetic variation in genes implicated in the beta-catenin destruction complex with risk of breast cancer.Cancer. Epidemiol Biomarkers Prev. 2008;17(8):2101–2108. doi:10.1158/1055-9965
30. Zheng HP, Zhou W, Xu LH, Si JM. [Correlation between gene polymorphisms of Wnt signalling pathway related components and risk of gastric carcinoma: a case-control study]. Zhonghua Yi Xue Yi Chuan Xue Za Zhi. 2006;23(6):647–651. Chinese.
31. Silbernagel G, Renner W, Grammer TB, et al. Association of TCF7L2 SNPs with age at onset of type 2 diabetes and proinsulin/insulin ratio but not with glucagon-like peptide 1. Diabetes Metab Res Rev. 2011;27(5):499–505. doi:10.1002/dmrr.1194
32. Tan SC, Ankathil R. Genetic susceptibility to cervical cancer: role of common polymorphisms in apoptosis-related genes. Tumour Biol. 2015;36(9):6633–6644. doi:10.1007/s13277-015-3868-2
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.