Back to Journals » OncoTargets and Therapy » Volume 13
Upregulation of miR-211 Promotes Chondrosarcoma Development via Targeting Tumor Suppressor VHL
Authors Chen C, Tian A, Zhou H, Zhang X, Liu Z, Ma X
Received 25 November 2019
Accepted for publication 24 March 2020
Published 5 April 2020 Volume 2020:13 Pages 2935—2943
DOI https://doi.org/10.2147/OTT.S239887
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tohru Yamada
Changbao Chen,1,* Aixian Tian,2,* Hua Zhou,3 Xiaolin Zhang,1 Zhongjun Liu,3 Xinlong Ma1
1Department of Spinal Surgery, Tianjin Hospital, Tianjin 300211, People’s Republic of China; 2Department of Orthopedics Institute, Tianjin Hospital, Tianjin 300211, People’s Republic of China; 3Department of Orthopaedic Surgery, Peking University Third Hospital, Beijing 100191, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xinlong Ma
Department of Spinal Surgery, Tianjin Hospital, 406 Jiefang South Road, Tianjin 300211, People’s Republic of China
Email [email protected]
Introduction: miR-211 has been demonstrated to be aberrantly expressed and plays a pivotal role in human cancers. However, its expression profiles and potential roles in chondrosarcoma development remain still elusive. This study aims to determine the clinical values and underlying roles of miR-211 in chondrosarcoma.
Methods: miR-211 expression was analyzed by qRT-PCR in chondrosarcoma specimens and the matched adjacent non-tumor tissues. The relationships among miR-211 expression, clinicopathological factors and overall survival were also evaluated. Cell viability, colony formation, migration and invasion were further investigated in chondrosarcoma cells. Potential target of miR-211 was predicted using bioinformatics to delineate the molecular mechanisms.
Results: miR-211 was remarkably increased in chondrosarcoma compared with the matched adjacent non-tumor tissues. High miR-211 level was identified as 66.7% in chondrosarcoma specimens, which were significantly associated with histological grade and MSTS stage. miR-211 had significant influences on the prognosis of chondrosarcoma patients. Multivariate analysis demonstrated that miR-211 was an independent prognostic factor for overall survival of chondrosarcoma patients. We also found that overexpression or inhibitor of miR-211 promotes or suppresses chondrosarcoma cell proliferation, migration and invasion, respectively. Mechanistically, miR-211 binds to the 3ʹ-UTR of Von Hippel-Lindau (VHL) and suppresses its expression, while restoration of VHL suppressed the potentiated function of miR-211 on proliferation and invasion of chondrosarcoma cells.
Conclusion: miR-211 is identified as a potent oncogenic function in chondrosarcoma development, which can serve as a novel biomarker to predict the survival of chondrosarcoma patients. miR-211 potentiates chondrosarcoma growth via targeting VHL, highlighting a novel attractive target for chondrosarcoma treatment.
Keywords: miR-211, chondrosarcoma development, VHL, cell survival, therapeutic target
Introduction
Chondrosarcoma is recognized as the common bone sarcoma and accounts for recurrence and death.1 This malignancy also develops chemoresistance and radioresistance, which warrants further research.2 Thus, novel biomarkers and targeted therapy are in demand by exploring potential biological targets governing the development of chondrosarcoma.2,3
MicroRNAs are identified as a large family of noncoding RNAs of 19–25 nucleotides, negatively regulating gene expression by directly binding to the 3ʹ untranslated region (3ʹ-UTR) of target mRNAs, which induces mRNA degradation or protein translation repression.4 Emerging evidence has linked microRNAs to the pathogenesis and progression of human cancers.5 In addition, many reports have documented that several microRNAs were dysregulated in chondrosarcoma development and progression, such as microRNA-30a, −145, and −181.6 Notably, microRNA-211 (miR-211) is located on intron 6 of the Trpm1 gene at 15q13-q14, a locus frequently lost in many neoplasms.7 Thus, miR-211 is recognized as a tumor suppressor in melanoma and other cancers.8–12 However, miR-211 also functions as an oncogenic miRNA in human cancers such as colorectal cancer by targeting chromodomain helicase DNA binding protein 5 (CHD5),13 and breast cancer by downregulating several tumor suppressor genes.14 In addition, miR-211 promotes cell growth by targeting the p53-induced loc285194 long noncoding RNA in colorectal cancer.15 miR-211 can prevent the endoplasmic reticulum (ER) induction by transcriptional repression of C/EBP homologous protein (CHOP) to modulate the cell survival.16 To date, no related report has addressed the clinical value and potential role of miR-211 in chondrosarcoma, and molecular mechanisms whereby miR-211 regulates chondrosarcoma development are poorly understood.
In this study, we initially determined the expression profile and clinical significance of miR-211 in chondrosarcoma. We further explored the potential role and molecular mechanism of miR-211-mediated chondrosarcoma development. This study provides the basis for using miR-211 as a novel biomarker and therapeutic target in chondrosarcoma.
Methods
Clinical Specimens and Cell Line
Ten cases of fresh conventional chondrosarcoma tissues and the corresponding non-tumor tissues were selected in Tianjin Hospital. Sixty-three cases of conventional chondrosarcomas were collected as described previously.17 Histological grading and the Musculoskeletal Tumor Society (MSTS) staging were carefully investigated as reported previously.18,19 The median follow-up time was 30 months (range 4 to 98 months).
Human chondrosarcoma SW1353 cell line were obtained from the American Type Culture Collection (Bethesda, MD, USA) and grown in DMEM with 10% FBS. All cells were maintained in a humidified cell incubator with 5% CO2 at 37°C.
miRNA Targets Prediction, Oligonucleotide and Plasmid Transfection
The putative miRNA targets were predicted using the TargetScan (http://www.targetscan.org/), miRanda (http://www.microrna.org/) and PicTar (http://pictar.mdc-berlin.de/) algorithms. The miR-211 mimics, a nonspecific miRNA control (Ctrl mimic), miR-211 inhibitor, a nonspecific miRNA inhibitor control (Ctrl inhibitor) were purchased from Genepharma. The plasmid with Von Hippel-Lindau (VHL) overexpression was previously described.20 miRNAs, oligonucleotide and plasmid transfection were transfected with Lipofectamine 2000 (Invitrogen). The transfected cells were incubated at 37°C for 24 hours or 48 hours in complete medium and harvested at the indicated time points.
RNA Extraction and Real-Time PCR
For the quantification of miR-211 expression, a stem-loop qRT-PCR was performed. Briefly, 5μg of small RNA extracted from cells was reverse transcribed to cDNA with the stem-loop RT primer using the M-MLV reverse transcriptase (Takara, Japan). The cDNA was subsequently used for the amplification of miR-211 and an endogenous control, U6 snRNA, via PCR. PCR cycles were as follows: 95°C for 3 min, followed by 40 cycles of 95°C for 30 s, 58°C for 30 s and 72°C for 30 s. The mRNA expression of VHL was investigated using qRT-PCR, and GAPDH was used as an endogenous control. The primer sequences of VHL were previously validated.21 The fold changes were calculated through relative quantification with 2–ΔΔCt and were representative of at least three independent experiments.
Protein Extraction and Western Blot Analysis
Protein extraction and Western blot were performed using a standard protocol as previously presented.20 Primary VHL antibodies were described.21 β-actin was used as an internal loading control. The proteins were transferred to a PVDF membrane (Bio-Rad) and further incubated with the secondary antibodies.
Luciferase Reporter Assay
The 3ʹ-UTR sequence of VHL, containing the putative miR-211 binding site, was amplified by PCR using the primers. PCR products were cloned into the pcDNA3/GFP plasmid between the BamHI and EcoRI sites, which was designated as wildtype 3ʹ-UTR (wt 3ʹ-UTR). Moreover, a mutant fragment of 3ʹ-UTR VHL containing a mutated miR-211 binding site, was amplified using PCR site-directed mutagenesis and cloned into the pcDNA3/GFP plasmid between the same sites, resulting in mutated 3ʹ-UTR (mut 3ʹ-UTR). All insertions were confirmed by sequencing. The primers used to construct the luciferase reporters and mutations were followed as VHL wt (forward, GATCCGTGAACAAATGTCTTAAAGGGAA GCTTG; reverse, AATTCAAGCTTCCCTTTAAGACATTTGTTCACG) and VHL mut (forward, GATCCGTGAACAAATGTCTTTATCCGTTAAGCTTG; reverse, AATTCAAGCTTAACGGATAAAGACATTTGTTCACG); wt or mut 3ʹ-UTR vectors encoding Renilla luciferase were cotransfected with miR-211 mimics or negative control into HT-293T cells using Lipofectamine 2000 (Invitrogen). The luciferase activity was measured 48 hours later using the Dual-Luciferase Reporter Assay System (HITACHI, Tokyo, Japan). The Renilla luciferase activity was normalized to firefly luciferase activity, and the ratios of Renilla/firefly values were presented. The experiments were performed independently in triplicate.
Cell Viability and Clonogenic Assays
Cell viability and clonogenic assays were investigated as illustrated previously.20 After treatment, SW1353 cells were seeded in 96-well plates at 2×103 cells/well and incubated for 1, 2, 3, 4 and 5 days. At each time point, CCK-8 (Dojindo, Kumamoto, Japan) was added into each well for 4h of incubation and calculated the cell viability. Respect to clonogenic assays, after treatment, SW1353 cells were seeded in 6-well plates at 1×103 cells per well overnight, cell medium was then replaced. Cells were grown for an additional 10–14 days. Colonies were fixed with methanol/acetone (1:1), stained with Gentian Violet, counted, and photographed. All samples were assayed in triplicate, and experiments were also performed in triplicate.
Cell Migration and Invasion Assays
Cell migration and invasion assays were investigated as previously presented using a modified two chamber transwell migration assay.20 After treatment, 5×104 SW1353 cells were seeded into the upper part of each transwell chamber. After 12h incubation at 37°C, cells in the upper chamber were carefully removed and the cells traversed the membrane were then fixed in methanol and stained with leucocrystal violet. The number of invasive cells was evaluated through counting the leucocrystal violet-stained cells. For the invasion assay, after treatment, 5×104 SW1353 cells were placed on the upper chamber of each insert coated with 40μL of 2mg/mL Matrigel (Growth Factor Reduced BD Matrigel TM Matrix), and 600 μL DMEM/F12 with 20% FBS was added to the lower part of the chamber. After 24h incubation at 37°C, noninvasive cells were gently removed, and the invasive cells at the bottom of the matrigel were fixed in methanol, stained with a 2% crystal violet solution for 10min. Cells migrated across the membrane were counted in five random visual fields. All assays were performed three independent times in triplicate.
Statistical Analysis
Values are the Mean ± SD. Two group means and a contingency table were compared for significance using 2-sided t tests and Pearson’s Chi-square test/Fisher’s exact test, respectively. Overall survival probabilities were estimated by the Kaplan-Meier method and the significance of differences was assessed by the Log rank test. Univariate and multivariate survival analyses were performed with the Cox regression model. Statistical analyses were performed using SPSS version 19 and P<0.05 was identified statistically significant. Each experiment was repeated two or three times as mentioned in each figure legend.
Results
Upregulation of miR-211 Was Associated with High-Grade Chondrosarcoma and Poor Prognosis
We initially measured the expression level of miR-211 in 10 cases of fresh chondrosarcoma tissues and the matched adjacent tissues, and found miR-211 expression in chondrosarcomas was significantly increased compared with the matched adjacent non-tumor tissues (Figure 1A, **P<0.01); Next, we shown that high miR-211 expression was identified as 66.7% (42/63) in 63 cases of chondrosarcoma tissues (Table 1). In addition, we analyzed the relationships between miR-211 and clinicopathological factors, and revealed that miR-211 expression significantly increased with the advancement of histological grade (P=0.017, Table 1) and MSTS stage (P=0.003, Table 1). However, no significant differences were illustrated for gender, age and anatomical location as shown in Table 1.
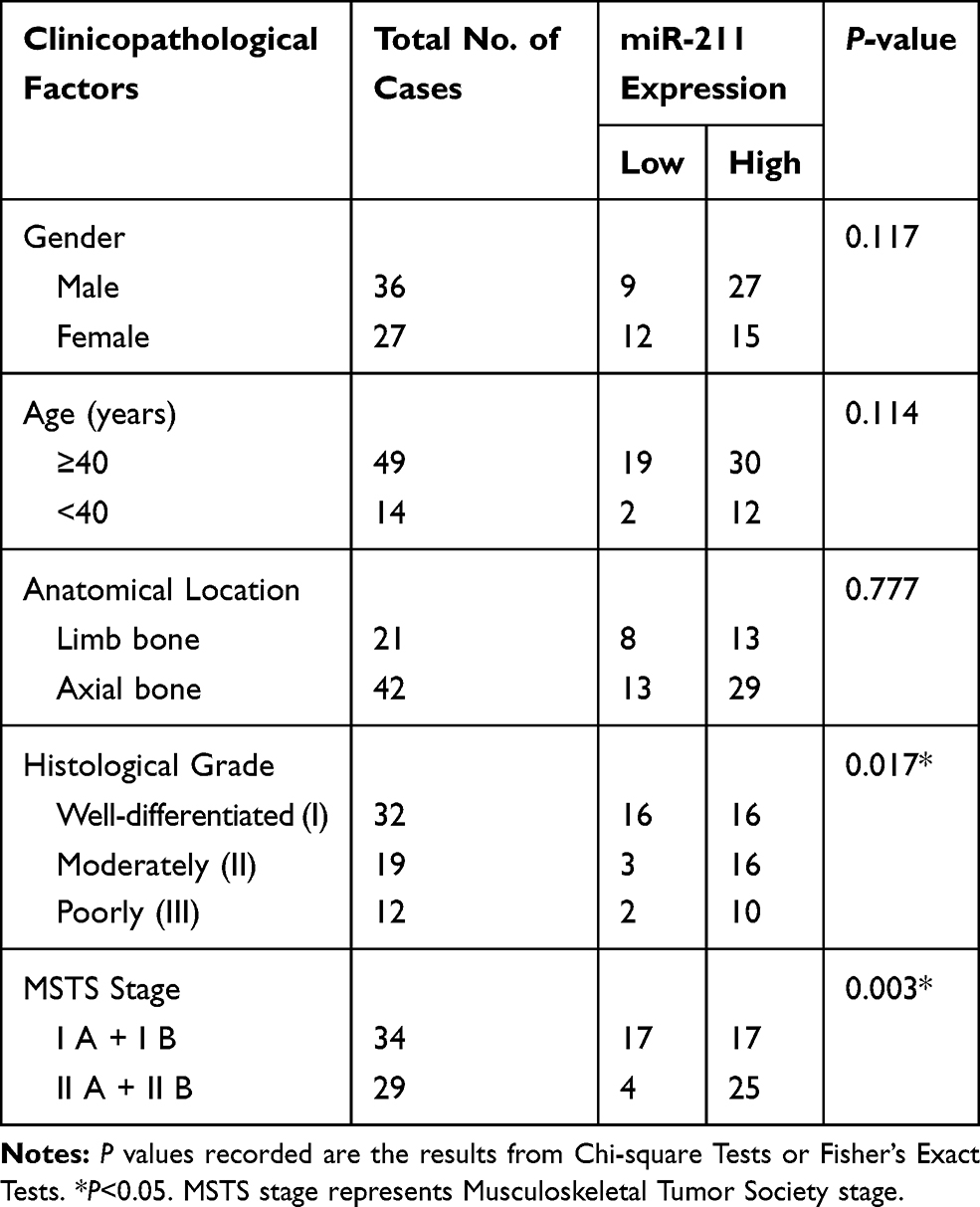 |
Table 1 Relationships Between miR-211 Expression and Clinicopathological Factors in 63 Cases of Chondrosarcomas |
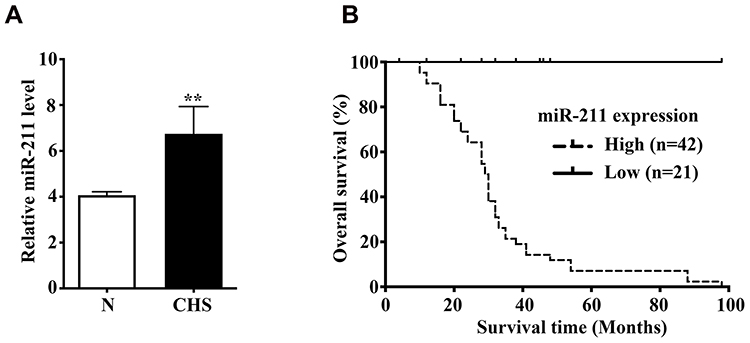 |
Figure 1 Upregulation of miR-211 is markedly correlated with poor prognosis in patients with chondrosarcoma. (A) The miR-211 expression level was analyzed by Real time PCR in 10 cases of chondrosarcoma tissues and the paired corresponding non-tumor tissues. N, the corresponding non-tumor tissues; CHS, chondrosarcoma tissues. CHS vs N, **P<0.01. (B) Prognostic significance of miR-211 expression in 63 patients with chondrosarcomas by Kaplan-Meier survival curves. High expression level of miR-211 was significantly associated with poor prognosis (P=0.024). |
To determine whether miR-211 can be predictive for the prognosis of chondrosarcoma patients, we had conducted the Kaplan-Meier analysis, and showed that the overall survival rate of patients with high-miR-211 tumors was significantly lower than that of patients with low-miR-211 tumors (P=0.024, Figure 1B). Univariate Cox regression analysis had demonstrated that histological grade, MSTS stage, and miR-211 expression were significantly associated with the overall survival (P=0.000, P=0.003, P=0.036, respectively). Multivariate analysis had presented that upregulation of miR-211 (HR, 6.115, P=0.006) was an independent prognostic marker for poor survival, similar to histological grade (HR, 3.150, P=0.028) and MSTS stage (HR, 5.006, P=0.004) as shown in Table 2. These findings suggested that miR-211 can serve as a novel independent predictor for the prognosis of chondrosarcoma patients.
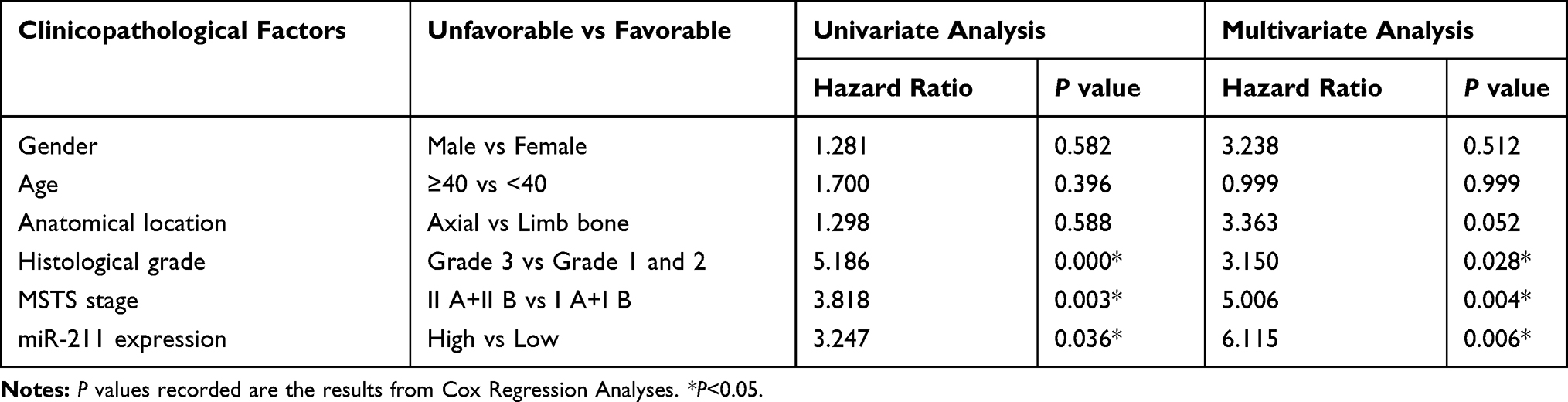 |
Table 2 Results of Cox Regression Model Analyses of the Prognostic Factors for Overall Survival in 63 Cases of Chondrosarcomas |
miR-211 Promotes Chondrosarcoma Cell Proliferation, Migration and Invasion
To illustrate the biological function of miR-211 in chondrosarcoma development, miR-211 expression was manipulated in SW1353 cells by transfecting miR-211 mimic or inhibitor, respectively, as validated by qRT-PCR (Figure 2A). Next, we found that miR-211 mimic significantly promoted cell viability and colony formation, whereas inhibitor of miR-211 markedly suppressed cell proliferation (Figure 2B and C). Notably, miR-211 mimic can promote the migration and invasion of SW1353 cells, whereas inhibitor of miR-211 significantly inhibited the migration and invasion (Figure 2D and E). Accordingly, these results suggested that miR-211 can potentiate chondrosarcoma cell proliferation, migration and invasion.
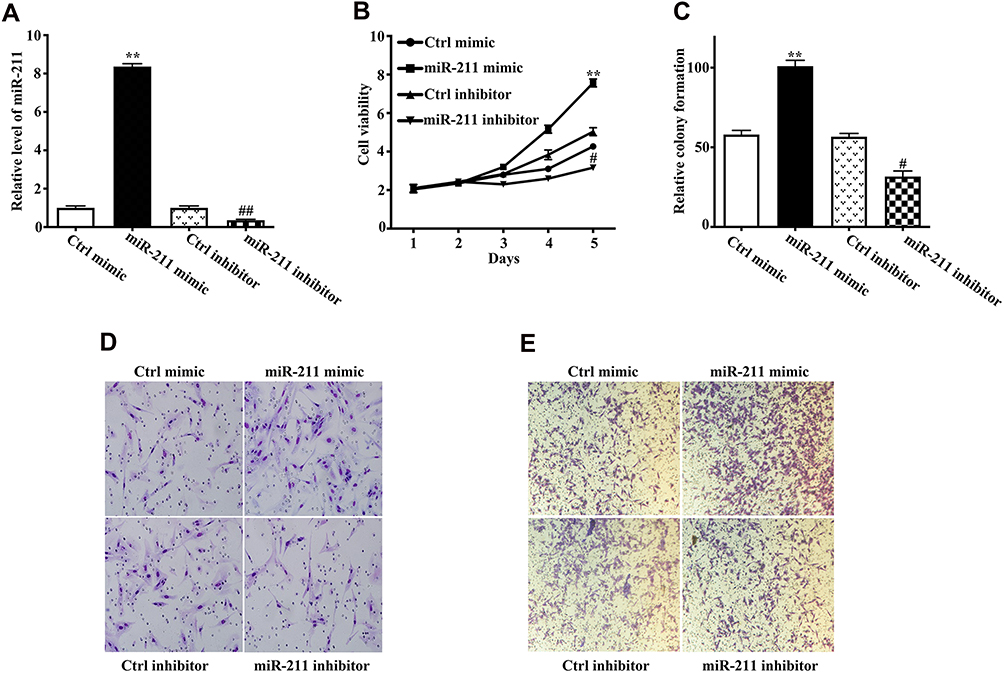 |
Figure 2 miR-211 potentiates cell viability, colony formation, migration and invasion of chondrosarcoma cells. (A) SW1353 cells were transfected with Ctrl mimic, miR-211 mimic, Ctrl inhibitor, and miR-211 inhibitor, respectively, and 48h later, the expression level of miR-211 was determined using RT-qPCR (three replicates per group, three independent experiments per group). miR-211 mimic vs Ctrl mimic, **P<0.01; miR-211 inhibitor vs Ctrl inhibitor, ##P<0.01. (B, C) SW1353 cell proliferation was evaluated by the CCK-8 assay at 1, 2, 3, 4 and 5d (B) and colony formation was evaluated by the soft agar assay (C). Data are plotted as the mean ± SD (miR-211 mimic vs Ctrl mimic, **P<0.01; miR-211 inhibitor vs Ctrl inhibitor, #P<0.05). (D, E) Effect of miR-211 on the migration and invasion abilities of chondrosarcoma cells. SW1353 cells were transfected with Ctrl mimic, miR-211 mimic, Ctrl inhibitor, and miR-211 inhibitor. Migration (D) and invasion (E) abilities were measured by the transwell assay as indicated in Materials and methods. All experiments were carried out in triplicate. |
VHL Identified as a Direct Downstream Target of miR- 211
To unravel the molecular mechanism of miR-211 in chondrosarcoma, we further determined a direct downstream target of miRNA-211. As predicted by miRanda, TargetScan and PicTar, the complementarity between miR-211 and VHL 3ʹ-UTR was illustrated (Figure 3A). To validate the direct downstream target of miR-211, we performed luciferase reporter assays, and found that miR-211 mimic or inhibitor significantly reduced or increased luciferase activity of the reporter gene in wild type, but not mutant type, respectively, indicating that miR-211 directly targeted the VHL 3ʹ-UTR (Figure 3B). This inhibitory effect was further validated by mRNA and protein expression of VHL as shown in Figure 3C and D. Taken together, these results presented that miR-211 directly recognizes the 3ʹ-UTR of VHL mRNA transcripts and inhibits its expression.
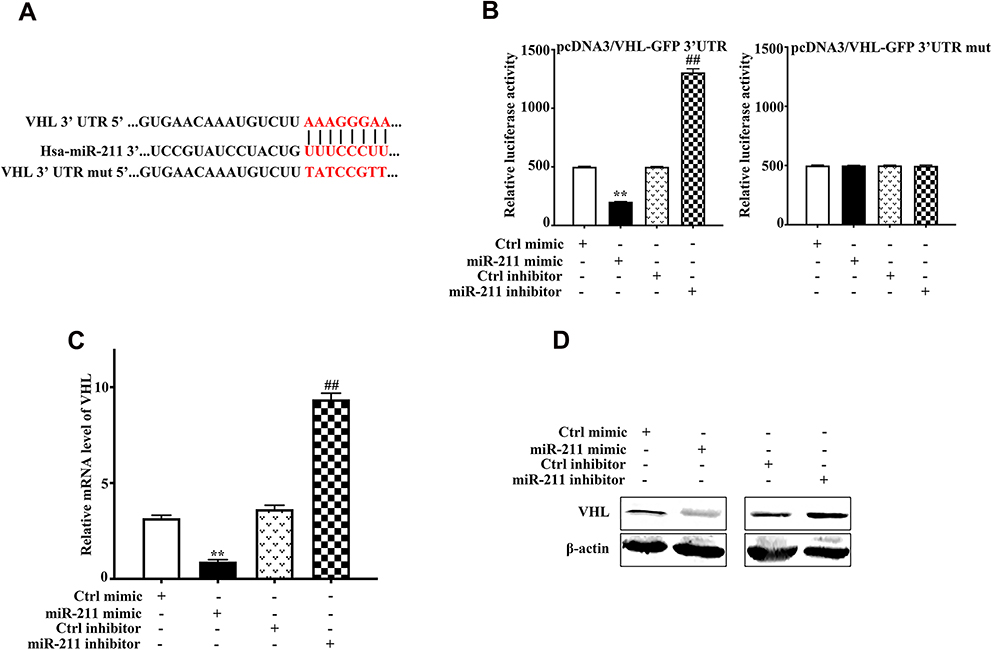 |
Figure 3 Identification of VHL as a target of miR-211. (A) Schematic representation of VHL 3ʹ-UTR demonstrating putative miRNA target site, luciferase activities of wild-type (wt-UTR) and mutant (mut-UTR) constructs. (B) Luciferase reporter activity following expression after transfections (Ctrl mimic, miR-211 mimic, Ctrl inhibitor, and miR-211 inhibitor) in SW1353 cells for 24h. Data are plotted as the mean ± SD (miR-211 mimic vs Ctrl mimic, **P<0.01, miR-211 inhibitor vs Ctrl inhibitor, ##P<0.05). (C) Real-time PCR analysis showed that miR-211 mimic can decrease VHL mRNA level, while miR-211 inhibitor can increase VHL mRNA level. Data are plotted as the mean ± SD (miR-211 mimic vs Ctrl mimic, **P<0.01, miR-211 inhibitor vs Ctrl inhibitor, ##P<0.05). (D) VHL protein expression in SW1353 cells after transfections. All results are representative of three independent experiments. |
Restoration of VHL Suppressed the Potentiated Function of miR-211 on Proliferation
We have presented that Ad5-VHL significantly increased VHL protein expression in chondrosarcoma as previously presented,20 whereas miR-211 mimic markedly decreased VHL protein expression (Figure 3D). Co-transfection of Ad5-VHL and miR-211 abrogated the inhibitory effects of miR-211 on VHL expression (Figure 4A). Importantly, miR-211 significantly increased while Ad5-VHL markedly suppressed SW1353 cell viability and colony formation, respectively; whereas co-transfection of Ad5-VHL and miR-211 suppressed the potentiated effects of miR-211 on viability and colony formation (Figure 4B and C). These results showed that overexpression of VHL by Ad5-VHL suppressed the potentiated function of miR-211 on proliferation, suggesting miR-211 can potentiate chondrosarcoma development via targeting VHL.
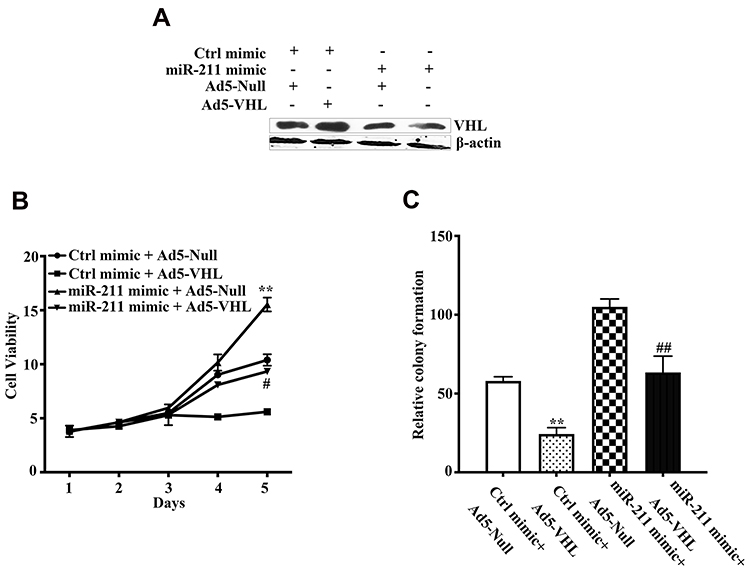 |
Figure 4 Restoration of VHL suppressed the potentiated proliferation function of miR-211. (A) SW1353 cells were transfected with the indicated vectors, and VHL protein level was detected by Western blot. β-actin protein regarded as endogenous normaliser. (B, C) Effect of VHL on proliferation function of miR-211 potentiated chondrosarcoma cells. SW1353 cells were transfected with the indicated vectors. SW1353 cell viability (B) and colony formation (C) were determined by CCK-8 and colony formation assays, respectively. CCK-8 and colony formation assays were performed to evaluate the cell proliferation and colony formation as indicated in Materials and methods. All experiments were carried out in triplicate. Data are plotted as the mean ± SD (miR-211 mimic+Ad5-Null vs Ctrl mimic+Ad5-Null, **P <0.01; miR-211 mimic+Ad5-VHL vs miR-211 mimic+Ad5-Null, #P<0.05, ##P <0.01). |
Discussion
Many reports have documented that microRNAs play a crucial role in human cancers.5 Recently, miR-211 was remarkably upregulated in colorectal cancer and breast cancer,13,14 whereas downregulated in melanomas and other tumors,8–12 indicating that miR-211 has distinct functions in multiple human cancers. However, its underlying biological role in chondrosarcoma remains poorly unravelled. In the present study, we first demonstrated that miR-211 was significantly upregulated in chondrosarcoma tissues as compared to the matched adjacent tissues, which would be consistent with a pro-oncogenic role for miR-211 in the pathogenesis of chondrosarcoma in vivo. To characterize the clinical significance of miR-211 in chondrosarcoma, we reported that upregulation of miR-211 is significantly associated with high-grade chondrosarcoma and poor prognosis, and miR-211 is recognized as an independent prognostic factor for overall survival of chondrosarcoma patients. These findings implicate that miR-211 can be identified as a prognostic biomarker for chondrosarcoma patients. However, our results were retrospectively analyzed and the small sample size was problematic, further research of larger cohorts is needed to validate its clinical value and prognosis in chondrosarcoma.
Accumulated evidence suggest that VHL has been identified as a tumor suppressor gene in human cancerogenesis.22 Specifically, we previously reported that reduced expression of VHL contributes to chondrosarcoma development,22 suggesting VHL is also identified as a tumor suppressor gene in chondrosarcoma development. Importantly, we also presented that restoration of VHL by adenoviral delivery suppresses chondrosarcoma cell growth through inhibition of Wnt/β-catenin signaling.20 In this study, using bioinformatics and a luciferase-based reporter assay, we have demonstrated that miR-211 can bind to the 3ʹ-UTR sequence of VHL, and decreased the mRNA and protein expression of VHL (Figure 3). Furthermore, ectopic expression of VHL by Ad5-VHL could effectively impede the ability of miR-211 to promote proliferation and metastasis (Figure 4). Our results presented stable evidence to support that miR-211 can promote proliferation and metastasis of chondrosarcoma by directly targeting VHL. VHL is a well characterized tumor suppressor gene, mainly inhibiting hypoxia-inducible factor (HIF)-α activity through the proteasome-dependent degradation via the ubiquitin-proteasome pathway.22,23 Therefore, reduced expression of VHL in chondrosarcoma as showed by our previous studies,21 in part due to increased level of miR-211 in chondrosarcoma tissues (Figure 1A), may prevent HIF-α from proteasome degradation, thereby leading to HIF-α accumulation in the nucleus that promotes chondrosarcoma cell proliferation and invasion. In deed, we have illustrated that dysregulation of HIF-1α and HIF-2α has previously been implicated in chondrosarcoma development.18,19 Therefore, further research that specifically explore the effect of miR-211 on HIF-α will be performed in the following studies implicated in chondrosarcoma development.
We presented for the first time that miR-211 can potentiate chondrosarcoma cell proliferation, migration and invasion in vitro (Figure 4), suggesting the crucial role of miR-211 in the development of chondrosarcoma. Intriguingly, VHL, a direct functional target of miR-211 in chondrosarcoma, would be an attractive therapeutic target.20 Notably, targeting dysregulated miRNA is expected to be a well-tolerated therapy, since differentially expressed miRNAs are often disease-dependent.24 Our main results have presented that increased levels of miR-211 as a post-transcriptional regulator of VHL could be a therapeutic strategy for chondrosarcoma treatment. However, this attractive therapy needs to be further validated in animal models, and strategies to apply miRNA precursors or mature miRNAs will be developed in the chondrosarcoma treatment.
Conclusions
Our work identifies upregulation of miR-211 positively associated with high-grade chondrosarcoma and poor prognosis in patients with chondrosarcoma, suggesting that miR-211 is a potent oncogenic function in chondrosarcoma. We have discovered that miR-211 potentiated growth effects in chondrosarcoma cells, in part, mediated through its downstream target gene, VHL, and highlighting a novel attractive target for chondrosarcoma treatment.
Data Sharing Statement
The datasets used and/or analyzed during this study are available from the corresponding author on reasonable request.
Acknowledgments
This work was supported by grants from the National Natural Sciences Foundation of China (81102037), and was also supported by grants from Tianjin Youth Medicine Talents Plan.
Author Contributions
Experiments Designed: Changbao Chen and Xinlong Ma. Experiments Performed: Changbao Chen, Aixian Tian, and Hua Zhou. Data Analysis: Xiaolin Zhang, Zhongjun Liu and Xinlong Ma. Facility and Reagents: Hua Zhou, Xiaolin Zhang and Zhongjun Liu. Manuscript Preparation: Changbao Chen, Aixian Tian and Xinlong Ma. All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
All authors declared that they have no competing interest in this work.
References
1. Nazeri E, Gouran Savadkoohi M, Majidzadeh AK, Esmaeili R. Chondrosarcoma: an overview of clinical behavior, molecular mechanisms mediated drug resistance and potential therapeutic targets. Crit Rev Oncol Hematol. 2018;131:102–109. doi:10.1016/j.critrevonc.2018.09.001
2. Brown HK, Schiavone K, Gouin F, Heymann MF, Heymann D. Biology of bone sarcomas and new therapeutic developments. Calcif Tissue Int. 2018;102(2):174–195. doi:10.1007/s00223-017-0372-2
3. Polychronidou G, Karavasilis V, Pollack SM, Huang PH, Lee A, Jones RL. Novel therapeutic approaches in chondrosarcoma. Future Oncol. 2017;13(7):637–648. doi:10.2217/fon-2016-0226
4. Shah MY, Ferrajoli A, Sood AK, Lopez-Berestein G, Calin GA. microRNA therapeutics in cancer - An emerging concept. EBioMedicine. 2016;12:34–42. doi:10.1016/j.ebiom.2016.09.017
5. Rupaimoole R, Slack FJ. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov. 2017;16(3):203–222. doi:10.1038/nrd.2016.246
6. Palmini G, Marini F, Brandi ML. What is new in the miRNA world regarding osteosarcoma and chondrosarcoma? Molecules. 2017;22(3). doi:10.3390/molecules22030417
7. Poetsch M, Kleist B. Loss of heterozygosity at 15q21.3 correlates with occurrence of metastases in head and neck cancer. Mod Pathol. 2006;19(11):1462–1469. doi:10.1038/modpathol.3800666
8. Mazar J, DeYoung K, Khaitan D, et al. The regulation of miRNA-211 expression and its role in melanoma cell invasiveness. PLoS One. 2010;5(11):e13779. doi:10.1371/journal.pone.0013779
9. Margue C, Philippidou D, Reinsbach SE, Schmitt M, Behrmann I, Kreis S. New target genes of MITF-induced microRNA-211 contribute to melanoma cell invasion. PLoS One. 2013;8(9):e73473. doi:10.1371/journal.pone.0073473
10. Mazar J, Qi F, Lee B, et al. MicroRNA 211 functions as a metabolic switch in human melanoma cells. Mol Cell Biol. 2016;36(7):1090–1108. doi:10.1128/MCB.00762-15
11. Xia B, Yang S, Liu T, Lou G. miR-211 suppresses epithelial ovarian cancer proliferation and cell-cycle progression by targeting Cyclin D1 and CDK6. Mol Cancer. 2015;14:57. doi:10.1186/s12943-015-0322-4
12. Deng B, Qu L, Li J, et al. MiRNA-211 suppresses cell proliferation, migration and invasion by targeting SPARC in human hepatocellular carcinoma. Sci Rep. 2016;6:26679. doi:10.1038/srep26679
13. Cai C, Ashktorab H, Pang X, et al. MicroRNA-211 expression promotes colorectal cancer cell growth in vitro and in vivo by targeting tumor suppressor CHD5. PLoS One. 2012;7(1):e29750. doi:10.1371/journal.pone.0029750
14. Lee H, Lee S, Bae H, Kang HS, Kim SJ. Genome-wide identification of target genes for miR-204 and miR-211 identifies their proliferation stimulatory role in breast cancer cells. Sci Rep. 2016;6:25287. doi:10.1038/srep25287
15. Liu Q, Huang J, Zhou N, et al. LncRNA loc285194 is a p53-regulated tumor suppressor. Nucleic Acids Res. 2013;41(9):4976–4987. doi:10.1093/nar/gkt182
16. Chitnis NS, Pytel D, Bobrovnikova-Marjon E, et al. miR-211 is a prosurvival microRNA that regulates chop expression in a PERK-dependent manner. Mol Cell. 2012;48(3):353–364. doi:10.1016/j.molcel.2012.08.025
17. Chen C, Zhou H, Zhang X, Ma X, Liu Z, Liu X. Elevated levels of Dickkopf-1 are associated with beta-catenin accumulation and poor prognosis in patients with chondrosarcoma. PLoS One. 2014;9(8):e105414. doi:10.1371/journal.pone.0105414
18. Chen C, Ma Q, Ma X, Liu Z, Liu X. Association of elevated HIF-2alpha levels with low Beclin 1 expression and poor prognosis in patients with chondrosarcoma. Ann Surg Oncol. 2011;18(8):2364–2372. doi:10.1245/s10434-011-1587-5
19. Chen C, Zhou H, Wei F, et al. Increased levels of hypoxia-inducible factor-1alpha are associated with Bcl-xL expression, tumor apoptosis, and clinical outcome in chondrosarcoma. J Orthop Res. 2011;29(1):143–151. doi:10.1002/jor.21193
20. Chen C, Tian A, Zhao M, Ma X. Adenoviral delivery of VHL suppresses bone sarcoma cell growth through inhibition of Wnt/beta-catenin signaling. Cancer Gene Ther. 2019;26(3–4):83–93. doi:10.1038/s41417-018-0041-2
21. Chen C, Zhou H, Liu X, Liu Z, Ma Q. Reduced expression of von hippel-lindau protein correlates with decreased apoptosis and high chondrosarcoma grade. J Bone Joint Surg Am. 2011;93(19):1833–1840. doi:10.2106/JBJS.I.01553
22. Gossage L, Eisen T, Maher ER. VHL, the story of a tumour suppressor gene. Nat Rev Cancer. 2015;15(1):55–64. doi:10.1038/nrc3844
23. Sanchez DJ, Simon MC. Transcriptional control of kidney cancer. Science. 2018;361(6399):226–227. doi:10.1126/science.aau4385
24. Catela Ivkovic T, Voss G, Cornella H, Ceder Y. microRNAs as cancer therapeutics: a step closer to clinical application. Cancer Lett. 2017;407:113–122. doi:10.1016/j.canlet.2017.04.007
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.