Back to Journals » OncoTargets and Therapy » Volume 12
Upregulated RACK1 attenuates gastric cancer cell growth and epithelial–mesenchymal transition via suppressing Wnt/β-catenin signaling
Authors Zhu L, Chen W, Li G, Chen H, Liao W, Zhang L, Xiao X
Received 18 February 2019
Accepted for publication 29 April 2019
Published 20 June 2019 Volume 2019:12 Pages 4795—4805
DOI https://doi.org/10.2147/OTT.S205869
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Geoffrey Pietersz
This paper has been retracted.
Lihui Zhu,* Wen Chen,* Guoqing Li, Honghui Chen, Wenqiu Liao, Li Zhang, Xiaoli Xiao
Department of Gastroenterology, The Second Hospital Affiliated to the University of South China, Hengyang, Hunan Province, People’s Republic of China
*These authors contributed equally to this work
Purpose: As there have been few studies on the effects of the receptor for activated C kinase 1 (RACK1) on gastric cancer (GC), we aimed to explore such effects and the mechanism that may be involved.
Patients and methods: Normal gastric epithelial cells and six GC cell lines were used to detect the mRNA expression of RACK1. Overexpressing RACK1 was transfected in HGC27 and MGC803 cells. The effects of overexpressing RACK1 on cell viability, migration, and invasion were determined by cell counting kit-8, wound scratch, and Transwell assay, respectively. The expressions of epithelial–mesenchymal transition (EMT) and Wnt/β-catenin signaling related genes were detected using quantitative real-time PCR or Western blot. Wnt pathway agonist LiCl was added into RACK1 overexpressing GC cells, and then cell viability, migration, and invasion were also detected.
Results: RACK1 was downregulated in GC cell lines. Under the circumstance that overexpressing RACK1 was successfully transfected in the two lowest RACK1-expressing GC cells, significant inhibition of cell viability, migration, and invasion, promotion to the mRNA and protein expression of E-cadherin, as well as a decrease in the N-cadherin and Snail expressions could be observed. Overexpressing RACK1 also enhanced the protein level of phosphorylation-β-catenin/β-catenin and attenuated c-Jun protein expression. Additionally, LiCl could partially reverse the inhibitory effects of cell viability, migration and invasion by overexpressing RACK.
Conclusion: We found RACK1 possibly inhibited epithelial–mesenchymal transition of GC cells through limitation of the Wnt/β-catenin pathway, thereby suppressing cell migration and invasion; RACK1 could also suppress cell growth.
Keywords: RACK1, gastric cancer, LiCl, Wnt/β-catenin signaling, epithelial-mesenchymal transition
Introduction
Gastric cancer (GC) is a heterogeneous, multifactorial disease.1,2 It poses a serious threat to the physical and mental health of human beings and brings heavy social and economic burden in both developed and developing countries.3 According to the GLOBOCAN series of the International Agency for Research on Cancer report, in 2012 there were about 951,000 new cases of GC and 723,000 deaths in the world, making it the fifth in morbidity and third in mortality of malignant tumor worldwide.4 Among them, more than 70% of GC cases occurred in developing countries, half of the world total cases occurred in East Asia, and the mortality rate of GC in East Asia was also the highest in the world.5 China was the main high incidence and high mortality area of GC.4 According to statistics of cancer in China in 2015, the number of new cases of GC was about 679,100, and the death toll was about 489,000.6 Studies have shown that environmental and genetic factors play a key role in the development of GC, with highly relevant environmental factors such as smoking, Helicobacter pyloriinfection, high salt and smoked food intake, and non-steroidal anti-inflammatory drug abuse.7 However, the incomplete early screening and the limitation of current cancer treatment are the main reasons for the high mortality rate of GC.8,9 At present, the main treatment of GC is radical surgery combined with radiotherapy and chemotherapy.10 More than half of patients with GC undergoing radical resection had a local recurrence or distant metastasis or were diagnosed when the tumor spread, and the median survival time was rarely more than 12 months.11 In the case of metastatic GC, the 5-year survival rate was lower than 10%, and in GC without metastasis, the 5-year survival rate was only about 20%.11
Recent studies have found that early diagnosis combined with advanced surgical treatment can effectively improve the prognosis and survival rate of patients with GC, and early diagnosis is the most critical factor in treatment.12–15 Receptor for activated C kinase 1 (RACK1), as a scaffold protein interacting with protein kinase C (PKC), was first discovered as a receptor of PKCβ II.16,17 RACK1 can bind a variety of kinases and receptors through different WD40 sites and participate in many signal transduction pathways, including PKC signaling, cAMP/PKA signaling, MAPK signaling and Src signaling, and play an important role in immune response, cell growth, migration, and differentiation.18–23 In addition, studies have shown that RACK1 plays a role in regulating cell proliferation, apoptosis, and migration in many cancers.24–27
To the best of our knowledge, the main mechanisms of RACK1 in GC are involved in nuclear factor-κB pathway and Wnt/β-catenin pathway.28–31 In the present study, we aimed to explore the effects of RACK1 on the biological function of GC cells and verify whether the effects of RACK1 on GC was through the Wnt/β-catenin pathway using Wnt pathway agonist LiCI.
Materials and methods
Cell culture
Normal gastric epithelial cells (GES-1 cells) and GC cell lines (SGC7901, BGC823, MKN45,AGS,HGC27,MGC803 cells) were purchased from Cell Bank of the Chinese Academy of Sciences (Shanghai, China). GES-1 and MKN45 cells were cultured in DMEM (Beijing,Solarbio, China), and others were cultured in RPMI1640 medium (Solarbio, Beijing, China). All of them were incubated with 10% FBS (Thermo Fisher Scientific, Waltham, MA, USA), 100 U/mL penicillin and 50 µg/mL streptomycin (Gibco, Thermo Fisher Scientific) at 37°C with 5% CO2.
HGC27 and MGC803 cells were selected to establish a LiCl-disposed cell model. Twenty mM LiCl was added into the medium after cell adherence and then cells were continuously cultured. The medium was changed every 1 d with 20 mM LiCl.
Cell transfection
HGC27 and MGC803 cells were seeded in 6-well plates (1.0×105) for 24 h before transfection. Overexpressing RACK1 and negative control (NC) plasmids were synthesized by Invitrogen (Thermo Fisher Scientific). Lipofectamine 2000 (Invitrogen) was performed to determine transient transfection according to the manufacturer’s protocol. A total of overexpressing RACK1 or NC and lipofectamine 2000 were respectively added to Opti-MEM medium. Lipofectamine/overexpressing RNA mixtures were cultured at 20°C for 10 min and then added into Opti-MEM RPMI1640 medium. After 6 h culturing, the fluid was changed back to RPMI1640 medium containing 10% FBS.
Cell counting kit-8 (CCK-8)
Cell viability of GC cells after treatment or transfection was performed by CCK-8 assay. Cells were plated into 96-well plates at a seeding density of 1×104 cells per well for 24 h. After the cells had been transfected for 12, 24, 48 h, 10 μL/well CCK-8 solution was added to each well and incubated for another 3 h at 37°C. OD was recorded at 450 nm by a microplate reader (Thermo Fisher Scientific).
Cell wound scratch assay
Cells were seeded in 12-well plates at 37°C incubation for 24 h. A wound was drawn using the sterile 10 ul pipette tip in the center of the plate, PBS was used to gently wash the cells 3 times, and then the serum-free medium was then added. Cell migration was observed by inverted microscope at 0 and 24 h. Scratch area was measured using Image J software.
Cell invasion assay
After transfection treatment, cells were resuspended in serum-free medium and 3×104 cells/well were added into the upper chamber coated with matrigel.32 RPMI1640 medium containing 10% FBS was added in the lower 24-well chamber, and the cells were incubated for 24 h at 37°C with 5% CO2. The cells were fixed with 4% formaldehyde for 20 min at 25°C and stained with 1% crystal violet for another 15 min. Invasion cells were counted at 200× magnification.
Quantitative real-time polymerase chain reaction (qRT-PCR)
RNA was isolated from cultured cells by use of Trizol reagent (Invitrogen) according to the manufacturer’s protocol. qRT-PCR was performed to detect RACK1, E-cadherin, N-cadherin, and Snail in different groups. Reverse transcription was performed by OrimeScript TM RT reagent kit (Takara, Otsu, Japan) following the manufacturer’s instructions at 42°C for 60 min and 70°C for 15 min. qRT-PCR was performed by SYBR Fast qPCR Mix (Invitrogen) and primer sequences are listed in Table 1. RACK1 ran at 94°C for 7 min, 94°C for 1 min, 55°C for 45 s followed by 28 cycles of 72°C for 2 min and 72°C for 10 min; other samples ran with the following cycling parameters: 95°C for 2 min, 95°C for 15 s, 60°C for 1 min followed by 40 cycles of 72°C for 30 s and 72°C for 10 min. The above primers were synthesized by Sangon Biotech (Shanghai, China). Amplified products were electrophoresed through 2% agarose gels. The amount of RNA was calculated using the 2−ΔΔCT method.
 | Table 1 Primers used in qRT-PCR |
Western blotting analysis
Proteins were collected from cultured cells in Radio Immuno Precipitation Assay (RIPA) lysis buffer (Beyotime, Shanghai, China). The concentrations of proteins were detected via BCA protein kit (Beyotime). Aliquots of protein were separated by 12% SDS-PAGE and resolved proteins were transferred to polyvinylidene fluoride (PVDF) membranes (EMD Millipore, Billerica, MA, USA). Membranes were blocked with 5% milk in PBS with 0.1% Triton X-100 and incubated with different primary antibodies: anti-RACK1 antibody (ab62735, 1:1,000, 30 kDa, Abcam, Cambridge, MA, USA), anti-E-cadherin antibody (ab15148, 1:500, 135 kDa, Abcam, USA), anti-N-cadherin antibody (ab18203, 1:1,000, 125 kDa, Abcam), anti-Snail antibody (ab82846, 1:500, 68 kDa, Abcam), anti-phosphorylation (p)-β-catenin antibody (ab27798, 1:500, 86 kDa, Abcam), anti-β-catenin antibody (ab32572, 1:5,000, 92 kDa, Abcam), anti-c-Jun antibody (ab32137, 1:1,000, 43 kDa, Abcam) and anti-GAPDH (ab181602, 1:10,000, 36 kDa, Abcam, USA) overnight at 4°C. The membranes were then incubated with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibody (Proteintech, Rosemont, IL, USA). Protein bands were detected with enhanced chemiluminescence (ECL, Thermo Fisher Scientific) and visualized by Quantity one (Bio-Rad Laboratories Inc., Hercules, CA, USA).
Statistical analysis
Statistical analysis was performed by Prism Graphpad version 6.0 software. All data were presented as mean ± SD. Differences were performed using one-way ANOVA following Turkey’s multiple comparison. A P <0.05 was considered as significant.
Results
Expression of RACK1 in gastric normal cells and cancer cell lines
In order to explore the expression of RACK1 in GC, we detected mRNA level of RACK1 in gastric normal cells (GES-1 cells) and six cancer cell lines (SGC7901, BGC823, MKN45, AGS, HGC27, and MGC803 cell lines). As Figure 1 shows, the mRNA expression of RACK1 was significantly downregulated in the GC cell lines (BGC823 and MKN45, P<0.05; others, P<0.01) compared with GES-1 cells. HGC27 and MGC803 cell lines were selected to conduct the following experiments as RACK1 was expressed at a lower level in the two cell lines.
 | Figure 1 Downregulation of RACK1 in GC cell lines. qRT-PCR was performed to detect the mRNA expression of RACK1 in gastric normal cells (GES-1 cells) and cancer cell lines (SGC7901, BGC823, MKN45, AGS, HGC27, and MGC803 cells). Data were shown as mean ± SD in three independent experiments. Note: Compared with gastric normal cells, *P<0.05, **P<0.01. Abbreviations: GC, gastric cancer; qRT-PCR, quantitative real-time polymerase chain reaction. |
Upregulated expression of RACK1 inhibits cell viability, migration, and invasion in HGC27 and MGC803 cells
When overexpressing RACK1 was transfected into HGC27 (Figure 2A and B) and MGC803 cells (Figure 2D and E), mRNA and protein levels of RACK1 were determined to detect transfection efficiency of overexpressing RACK1. The results showed that both in HGC27 and MGC803 cells, the mRNA and protein levels of RACK1 were noticeably high expressed in RACK1 group in comparison to control or NC (P<0.01, Figure 2A, B and D, E). Compared to control or NC, overexpressing RACK1 could decrease the cell viability at 12, 24 and 48 h in the two cell lines. However, in HGC27 cells, the significantly decreased cell viability was mainly presented at 12 (P<0.05) and 48 h (P<0.01) (Figure 2C). In MGC803 cells, the remarkably decreased cell viability was mainly presented at 24 h (P<0.05) and 48 h (P<0.01) (Figure 2F). In regard to cell migration and invasion, wound scratch (Figure 3A and E) and Transwell assay (Figure 3C and G) were performed. In HGC27 cells, overexpressing RACK1 significantly decreased cell migration and invasion compared with control or NC (migration, P<0.01, Figure 3B; invasion, P<0.05, Figure 3D). In MGC803 cells, a similar tendency of an inhibitory effect of overexpressing RACK1 on migration and invasion was observed as in HGC27 cells (P<0.01, Figure 3F and H).
 | Figure 2 The inhibitory effects of overexpressing RACK1 on cell viability in HGC27 and MGC803 cells. qRT-PCR (A) and Western blot (B) were used to detect the transfection efficiency of overexpressing RACK1 in HGC27 cells. (C) The effect of overexpressing RACK1 on cell viability at 12, 24, and 48 h was detected by CCK-8 assay in HGC27 cells. qRT-PCR (D) and Western blot (E) were used to detect the transfection efficiency of overexpressing RACK1 in MGC803 cells. (F) The effect of overexpressing RACK1 on cell viability at 12, 24, and 48 h was detected by CCK-8 assay in MGC803 cells. Expression of each protein in cells was following normalization with a loading control GAPDH. Data are shown as mean ± SD in three independent experiments. Notes: Compared with control, *P<0.05, **P<0.01; compared with NC ^P<0.05, ^^P<0.01. Abbreviations: qRT-PCR, quantitative real-time polymerase chain reaction; CCK-8, cell counting kit-8; NC, negative control. |
 | Figure 3 The inhibitory effects of overexpressing RACK1 on cell migration and invasion in HGC27 and MGC803 cells. (A) Wound scratch assay was performed to detect the effect of overexpressing RACK1 on HGC27 cell migration at 0, 24 h. (B) Wound closure is shown as bar diagrams in HGC27 cells. (C) Transwell assay was used to detect the effect of overexpressing RACK1 on HGC27 cell invasion. (D) Invasion capability is shown as bar diagrams in HGC27 cells. (E) Wound scratch assay was performed to detect the effect of overexpressing RACK1 on MGC803 cell migration at 0, 24 h. (F) Wound closure is shown as bar diagrams in MGC803 cells. (G) Transwell assay was used to detect the effect of overexpressing RACK1 on MGC803 cell invasion. (H) Invasion capability is shown as bar diagrams in MGC803 cells. Data are shown as mean ± SD from three independent experiments. Notes: Compared with control, *P<0.05, **P<0.01; compared with NC, ^P<0.05, ^^P<0.01. Abbreviation: NC, negative control. |
Upregulated expression of RACK1 regulates the expressions of epithelial–mesenchymal transition (EMT) related genes in HGC27 and MGC803 cells
The above experiments confirmed that upregulation of RACK1 could inhibit cell migration and invasion, so we further detected the expressions of EMT related genes. The results showed that there were no obvious differences in the expressions of EMT related genes in control and NC groups. Overexpressing RACK1 could noticeably increase the mRNA and protein levels of E-cadherin in HGC27 cells (P<0.01, Figure 4A, D and E). Meanwhile, both mRNA and protein expressions of N-cadherin and Snail showed a rapid reduction (P<0.01, Figure 4B–E) in RACK1 overexpressing HGC27 cells. In MGC803 cells, overexpressing RACK1 significantly increased the expression of E-cadherin and suppressed the expressions of N-cadherin and Snail in both mRNA and protein levels (P<0.01, Figure 4F–J).
 | Figure 4 The effects of overexpressing RACK1 on EMT-related genes. qRT-PCR was used to detect the overexpressing RACK1 on mRNA expressions of E-cadherin (A), N-cadherin (B) and Snail (C) in HGC27 cells. (D) Western blot was used to assess the overexpressing RACK1 on protein expressions of E-cadherin, N-cadherin, and Snail in HGC27 cells. (E) The three proteins were quantitative as bar diagrams in HGC27 cells. qRT-PCR was used to detect the overexpressing RACK1 on mRNA expressions of E-cadherin (F), N-cadherin (G) and Snail (H) in MGC803 cells. (I) Western blot was used to assess the overexpressing RACK1 on protein expressions of E-cadherin, N-cadherin, and Snail in MGC803 cells. (J) The three proteins were quantitative as bar diagrams in MGC803 cells. Expression of each protein in cells was following normalization with a loading control GAPDH. Data are shown as mean ± SD in three independent experiments.Notes: Compared with control, *P<0.05, **P<0.01; ^compared with NC, ^P<0.05, ^^P<0.01. Abbreviations: EMT, epithelial–mesenchymal transition; qRT-PCR, quantitative real-time polymerase chain reaction; NC, negative control. |
Upregulated expression of RACK1 regulates the related proteins expressions of Wnt/β-catenin signaling
In order to explore the possible mechanism of the inhibitory effects of overexpressing RACK1 on cell viability, migration and invasion, the related proteins of Wnt/β-catenin signaling in HGC27 (Figure 5A) and MGC803 cells (Figure 5D) were detected by Western blot. The results showed that in both the cell lines, overexpressing RACK1 could increase p-β-catenin/β-catenin (P<0.01, Figure 5B and E) expression and dramatically decrease c-Jun expression (P<0.01, Figure 5C and F) at the protein level. To further verify whether the inhibitory effects of overexpressing RACK1 on the two cell lines were through Wnt/β-catenin signaling, we applied the Wnt signaling agonist, LiCl. As shown in Figure 6, no obvious difference in cell viability in HGC27 (Figure 6A) and MGC803 cells (Figure 6E) was observed in LiCI group and in the control or NC group; however, in the LiCI group, cell viability in HGC27 (Figure 6A) and MGC803 cells (Figure 6E) were increased, compared to group overexpressing RACK1 (P<0.01). Overexpressing RACK1 could reverse the increase of cell viability induced by LiCl (P<0.01). Wound scratch and Transwell assay were performed to detect cell migration and invasion, respectively (Figure 6D and H). In HGC27 cells, the results showed that LiCl could significantly induce cell migration and invasion in comparison to RACK1 (P<0.01, Figure 6B and C). In the case of RACK1 combined with LiCl, a significant inhibitory tendency was observed in cell migration compared to that under LiCl treatment alone (P<0.01), while no noticeable reduction of invasion was observed compared to that under LiCl treatment alone (P>0.05). In MGC803 cells, compared to the RACK1 group, LiCl could also remarkably increase cell migration and invasion (P<0.01, Figure 6F and G). Overexpressing RACK1 could significantly decrease cell migration induced by LiCl (P<0.01). In the case of RACK1 combined with LiCl, a slight reduction of cell invasion was observed in comparison to that under LiCl treatment alone (P>0.05). All the above results indicated that Wnt/β-catenin signaling was the possible underlying mechanism of the inhibitory effects of overexpression RACK1 on cell growth and metastasis in GC cells.
 | Figure 5 The effects of overexpressing RACK1 on the Wnt/β-catenin signaling-related proteins in HGC27 and MGC803 cells. (A) p-β-catenin, β-catenin, and c-Jun were detected by Western blot to obverse the effects of overexpressing RACK1 on protein expressions in HGC27 cells. (B) p-β-catenin/β-catenin protein level is shown as bar diagrams in HGC27 cells. (C) c-Jun protein level is shown as bar diagrams in HGC27 cells. (D) p-β-catenin, β-catenin, and c-Jun were detected by Western blot to obverse the effects of overexpressing RACK1 on protein expressions in MGC803 cells. (E) p-β-catenin/β-catenin protein level is shown as bar diagrams in MGC803 cells. (F) c-Jun protein level is shown as bar diagrams in MGC803 cells. Expression of each protein in cells was following normalization with a loading control GAPDH. Data are shown as mean ± SD in three independent experiments.Notes: Compared with control, *P<0.05, **P<0.01; compared with NC, ^P<0.05, ^^P<0.01. Abbreviations: p-β-catenin, phosphorylation-β-catenin; NC, negative control. |
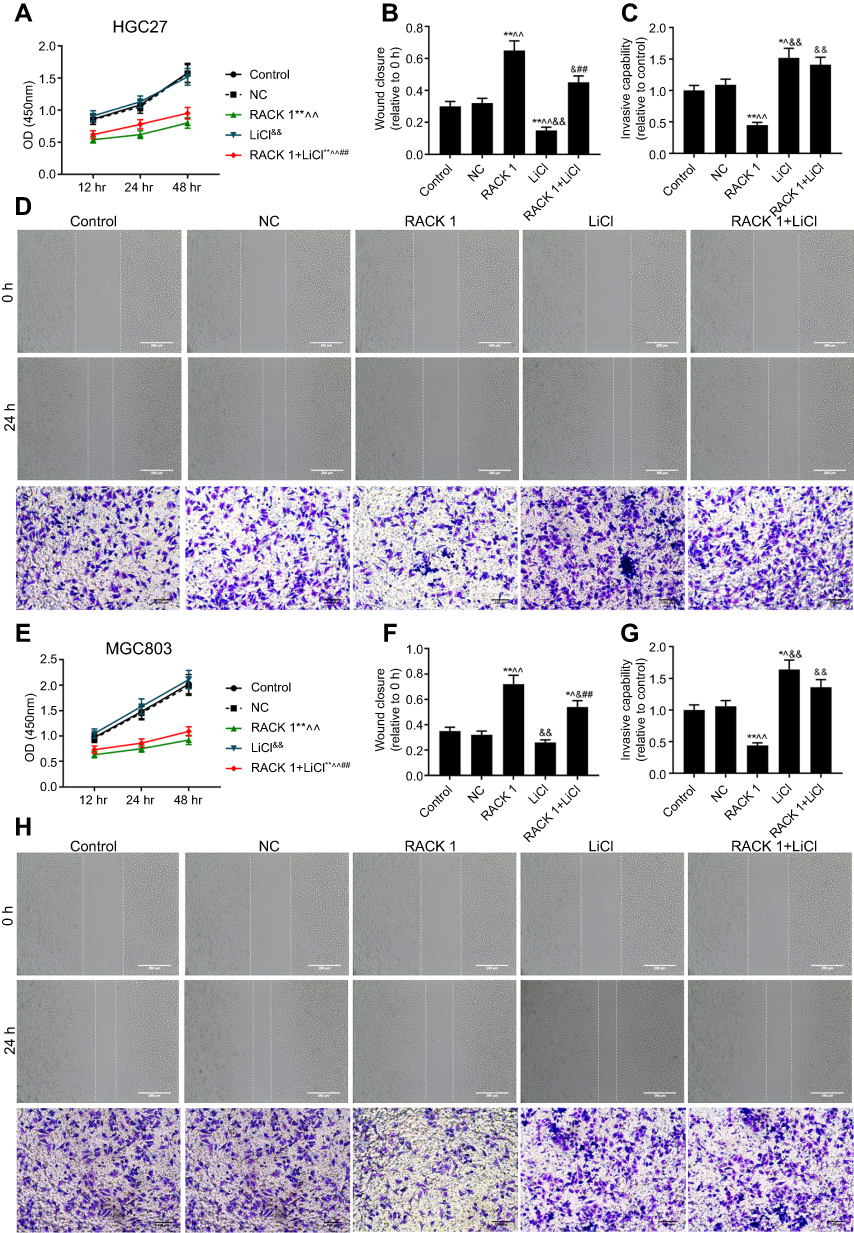 | Figure 6 The verification experiments of Wnt/β-catenin in GC cells. (A) The suppression effects of overexpressing RACK1 on increasing cell viability induced by LiCl in HGC27 cells. (B) The suppression effects of overexpressing RACK1 on increasing cell migration induced by LiCl in HGC27 cells. (C) The suppression effects of overexpressing RACK1 on increasing cell invasion induced by LiCl in HGC27 cells. (D) Cell migration and invasion in HGC27 cells were respectively performed by wound scratch and Transwell assay. (E) The suppression effects of overexpressing RACK1 on increasing cell viability induced by LiCl in MGC803 cells. (F) The suppression effects of overexpressing RACK1 on increasing cell migration induced by LiCl in MGC803 cells. (G) The suppression effects of overexpressing RACK1 on increasing cell invasion induced by LiCl in MGC803 cells. (H) Cell migration and invasion in MGC803 cells were respectively performed by wound scratch and Transwell assay. Data are shown as mean ± SD in three independent experiments. Notes: Compared with control, *P<0.05, **P<0.01; compared with NC, ^P<0.05, ^^P<0.01; compared with RACK1, &P<0.05, &&P<0.01; compared with LiCl, #P<0.05, ##P<0.01. Abbreviations: GC, gastric cancer; NC, negative control. |
Discussion
Increasing research is carried out on RACK1, especially in relation to cancer.33 Studies have shown that RACK1 can promote tumor invasion and metastasis and multiple cellular functions by PKC.34 At the same time, RACK1 binds to PKC and performs corresponding translation and regulation of ribosomes, thereby activating factors related to biological functions such as tumor invasion and metastasis.34 The abnormal expression of RACK1 is closely associated with the occurrence and development of tumor, and increased expression of RACK1 occurred in most tumors such as breast cancer, non-small cell lung cancer, liver cancer, and melanoma, indicating that it acts as an oncogene.35–38 In the present study, the finding showed that RACK1 was downregulated in several GC cell lines compared with normal gastric epithelial cells. The result was in accordance with Deng's report that RACK1 was a tumor suppressor gene of GC.39 Furthermore, it was expressed at a low level in GC and was associated with tumor invasion and low differentiation.39 Although Liu reported that RACK1 proteins were expressed at a low level in GC patients,40 and Chen reported that low expression of RACK1 predicted poor prognosis for overall five-year survival in all GC cases,29 our study lacked the information about RACK1 prognosis from TCGA database. We are planning to launch this analysis in a future study.
In addition, overexpressing RACK1 was successfully transfected into HGC27 and MGC803 cells, and showed significant inhibitory effects on cell viability, migration, and invasion. We also detected the effects of overexpressing RACK on EMT genes in GC. To the best of our knowledge, EMT is a critical and necessary process in the early stage of tumor metastasis.41 In this process, tumor cells lose epithelial cell characteristics, including E-cadherin expression, intercellular adhesion, and epithelial cell polarity; and acquire mesenchymal cell characteristics, including migration and invasion.42–45 Our results found that overexpressing RACK1 regulated markers of EMT. Specifically, it inhibited the expression of N-cadherin and promoted E-cadherin expression. Meanwhile, overexpressing RACK1 suppressed Snail expression, which could induce the occurrence of EMT and downregulate E-cadherin expression in the progression of cancer.46,47
In regard to the underlying mechanism of RACK1 on the above biological function, we explored the Wnt signaling. There is a study reported that RACK1 contributed to the occurrence of GC through activation of the Wnt pathway by increasing dissociation of the β-catenin complex.39 In our work, overexpression RACK1 could significantly increase the protein expression of p-β-catenin/β-catenin and decrease the protein expression of c-Jun. β-catenin is a key step in the Wnt pathway.48 The abnormal activation of Wnt/β-catenin signaling pathway leads to a significant increase in the accumulation of β-catenin in the nucleus, triggering excessive proliferation, and malignant transformation of cells.49,50 C-Jun is a member of the Jun gene family and is an oncogene.51 It encodes the components of transcription factor AP-1 in dimer form.52 Takeda reported that in the activated Wnt pathway, β-catenin/TCF can bind to the c-Jun promoter and induce c-Jun transcription, which plays an important role in the development and progression of colorectal cancer.53 Therefore, we inferred the effect of overexpressing RACK1 on GC cells through inhibition of Wnt/β-catenin signaling.
Furthermore, to verify the correlation between this pathway and the anti-cancer effect of RACK1, we added Wnt pathway agonist LiCl to RACK1 overexpressing GC cells. The results presented that LiCl could noticeably increase cell viability, induce cell migration and invasion in comparison to control or NC. Meanwhile, LiCl could also partially reverse the inhibitory effects of overexpressing RACK1 on cell viability, migration, and invasion. RACK1 may inhibit the proliferation, migration, and invasion of GC cells by blocking the Wnt/β-catenin signaling pathway. There may be other pathways at the same time, and more in-depth research is needed.
In this study, there are still many limitations, for instance, a lack of research on cell mortality, experiments regarding the relation between cell migration/invasion and cell viability, as well as JNK activator, to validate the involved mechanism of Wnt/β-catenin signaling. With regard to this, we are planning to launch a comprehensive, confirmative and in-depth study to make up the limitations.
Conclusion
In this study, we found that RACK1 possibly inhibited EMT of GC cells through limitation of the Wnt/β-catenin pathway, thereby suppressing cell migration and invasion; and also inhibited cell growth. This study provides a theoretical basis for determining the anti-cancer effects of RACK1 and its application in the clinical study and targeted therapy of GC.
Acknowledgments
This work was supported by the National Natural Science Foundation of China [81372378].
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gigek CO, Calcagno DQ, Rasmussen LT, et al. Genetic variants in gastric cancer: risks and clinical implications. Exp Mol Pathol. 2017;103(1):101–111. doi:10.1016/j.yexmp.2017.07.004
2. Ratti M, Lampis A, Hahne JC, Passalacqua R, Valeri N. Microsatellite instability in gastric cancer: molecular bases, clinical perspectives, and new treatment approaches. Cell Mol Life Sci. 2018;75(22):4151–4162. doi:10.1007/s00018-018-2906-9
3. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127(12):2893–2917. doi:10.1002/ijc.25516
4. Ferlay J, Soerjomataram I, Dikshit R, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136(5):E359–E386. doi:10.1002/ijc.29210
5. Rahman R, Asombang AW, Ibdah JA. Characteristics of gastric cancer in Asia. World J Gastroenterol. 2014;20(16):4483–4490. doi:10.3748/wjg.v20.i16.4483
6. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
7. Karimi P, Islami F, Anandasabapathy S, Freedman ND, Kamangar F. Gastric cancer: descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol Biomarkers Prev. 2014;23(5):700–713. doi:10.1158/1055-9965.EPI-13-1057
8. Takahashi T, Saikawa Y, Kitagawa Y. Gastric cancer: current status of diagnosis and treatment. Cancers. 2013;5(1):48–63. doi:10.3390/cancers5010048
9. Chen XZ, Zhang WH, Hu JK. A difficulty in improving population survival outcome of gastric cancer in mainland China: low proportion of early diseases. Med Oncol. 2014;31(12):315. doi:10.1007/s12032-014-0374-0
10. Orditura M, Galizia G, Sforza V, et al. Treatment of gastric cancer. World J Gastroenterol. 2014;20(7):1635–1649. doi:10.3748/wjg.v20.i7.1635
11. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
12. Gill RS, Al-Adra DP, Nagendran J, et al. Treatment of gastric cancer with peritoneal carcinomatosis by cytoreductive surgery and HIPEC: a systematic review of survival, mortality, and morbidity. J Surg Oncol. 2011;104(6):692–698. doi:10.1002/jso.22017
13. Bairamov RB, Abdullaeva RT. [The impact of early gastric cancer diagnosis on indices of survival in patients after radical surgical intervention]. Klinichna Khirurhiia. 2013;6:18–21.
14. Smyth EC, Verheij M, Allum W, Cunningham D, Cervantes A, Arnold D. Gastric cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann Oncol. 2016;27(suppl 5):v38–v49. doi:10.1093/annonc/mdw350
15. Tartaglia D, Bertolucci A, Galatioto C, et al. Incidental appendectomy? Microscopy tells another story: a retrospective cohort study in patients presenting acute right lower quadrant abdominal pain. Int J Surg. 2016;28:149–152. doi:10.1016/j.ijsu.2016.02.085
16. Wang Z, Zhang B, Jiang L, et al. RACK1, an excellent predictor for poor clinical outcome in oral squamous carcinoma, similar to Ki67. Eur J Cancer. 2009;45(3):490–496. doi:10.1016/j.ejca.2008.11.012
17. Adams DR, Ron D, Kiely PA. RACK1, A multifaceted scaffolding protein: structure and function. Cell Commun Signal. 2011;9:22. doi:10.1186/1478-811X-9-22
18. Berns H, Humar R, Hengerer B, Kiefer FN, Battegay EJ. RACK1 is up-regulated in angiogenesis and human carcinomas. Faseb J. 2000;14(15):2549–2558. doi:10.1096/fj.99-1038com
19. Mamidipudi V, Cartwright CA. A novel pro-apoptotic function of RACK1: suppression of Src activity in the intrinsic and Akt pathways. Oncogene. 2009;28(50):4421–4433. doi:10.1038/onc.2009.293
20. Yan Y, Jiang Y. RACK1 affects glioma cell growth and differentiation through the CNTN2-mediated RTK/Ras/MAPK pathway. Int J Mol Med. 2016;37(1):251–257. doi:10.3892/ijmm.2015.2421
21. Liu Q, Wang X, Liu Y, et al. RACK1 inhibits morphine re-exposure via inhibition of src. Neurol Res. 2011;33(1):56–62. doi:10.1179/016164110X12714125204236
22. Li J, Guo Y, Feng X, et al. Receptor for activated C kinase 1 (RACK1): a regulator for migration and invasion in oral squamous cell carcinoma cells. J Cancer Res Clin Oncol. 2012;138(4):563–571. doi:10.1007/s00432-011-1097-7
23. Ruan Y, Sun L, Hao Y, et al. Ribosomal RACK1 promotes chemoresistance and growth in human hepatocellular carcinoma. J Clin Invest. 2012;122(7):2554–2566. doi:10.1172/JCI58488
24. Hu F, Tao Z, Wang M, et al. RACK1 promoted the growth and migration of the cancer cells in the progression of esophageal squamous cell carcinoma. Tumour Biol. 2013;34(6):3893–3899. doi:10.1007/s13277-013-0977-7
25. Li JJ, Xie D. RACK1, a versatile hub in cancer. Oncogene. 2015;34(15):1890–1898. doi:10.1038/onc.2014.127
26. Shen F, Yan C, Liu M, Feng Y, Chen Y. RACK1 promotes prostate cancer cell proliferation, invasion and metastasis. Mol Med Rep. 2013;8(4):999–1004. doi:10.3892/mmr.2013.1612
27. Wu J, Meng J, Du Y, et al. RACK1 promotes the proliferation, migration and invasion capacity of mouse hepatocellular carcinoma cell line in vitro probably by PI3K/Rac1 signaling pathway. Biomed Pharmacother = Biomedecine & Pharmacotherapie. 2013;67(4):313–319. doi:10.1016/j.biopha.2013.01.011
28. Yong-Zheng X, Wan-Li M, Ji-Ming M, Xue-Qun R. Receptor for activated protein kinase C 1 suppresses gastric tumor progression through nuclear factor-kB pathway. Indian J Cancer. 2015;52(Suppl 3):E172–E175. doi:10.4103/0019-509X.186572
29. Chen L, Min L, Wang X, et al. Loss of RACK1 promotes metastasis of gastric cancer by Inducing a miR-302c/IL8 signaling loop. Cancer Res. 2015;75(18):3832–3841. doi:10.1158/0008-5472.CAN-14-3690
30. Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004;20:781–810. doi:10.1146/annurev.cellbio.20.010403.113126
31. Sareddy GR, Challa S, Panigrahi M, Babu PP. Wnt/beta-catenin/Tcf signaling pathway activation in malignant progression of rat gliomas induced by transplacental N-ethyl-N-nitrosourea exposure. Neurochem Res. 2009;34(7):1278–1288. doi:10.1007/s11064-008-9906-3
32. Wang L, Li B, Zhang L. et al. miR-664a-3p functions as an oncogene by targeting hippo pathway in the development of gastric cancer. Cell Prolif;2019. e12567. doi:10.1111/cpr.12567
33. Brina D, Miluzio A, Ricciardi S, Biffo S. eIF6 anti-association activity is required for ribosome biogenesis, translational control and tumor progression. Biochim Biophys Acta. 2015;1849(7):830–835. doi:10.1016/j.bbagrm.2014.09.010
34. Lopez-Bergami P, Huang C, Goydos JS, et al. Rewired ERK-JNK signaling pathways in melanoma. Cancer Cell. 2007;11(5):447–460. doi:10.1016/j.ccr.2007.03.009
35. Cao XX, Xu JD, Xu JW, et al. RACK1 promotes breast carcinoma migration/metastasis via activation of the RhoA/Rho kinase pathway. Breast Cancer Res Treat. 2011;126(3):555–563. doi:10.1007/s10549-010-0955-3
36. Nagashio R, Sato Y, Matsumoto T, et al. Expression of RACK1 is a novel biomarker in pulmonary adenocarcinomas. Lung Cancer. 2010;69(1):54–59. doi:10.1016/j.lungcan.2009.09.015
37. Egidy G, Jule S, Bosse P, et al. Transcription analysis in the MeLiM swine model identifies RACK1 as a potential marker of malignancy for human melanocytic proliferation. Mol Cancer. 2008;7:34. doi:10.1186/1476-4598-7-34
38. Park SM, Gaur AB, Lengyel E, Peter ME. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008;22(7):894–907. doi:10.1101/gad.1640608
39. Deng YZ, Yao F, Li JJ, et al. RACK1 suppresses gastric tumorigenesis by stabilizing the beta-catenin destruction complex. Gastroenterology. 2012;142(4):812–823.e815. doi:10.1053/j.gastro.2011.12.046
40. Liu C, Ren L, Wang Y, Liu Y, Xiao J. The interaction between RACK1 and WEE1 regulates the growth of gastric cancer cell line HGC27. Oncol Lett. 2017;14(4):4784–4792. doi:10.3892/ol.2017.6741
41. Kraljevic Pavelic S, Sedic M, Bosnjak H, Spaventi S, Pavelic K. Metastasis: new perspectives on an old problem. Mol Cancer. 2011;10:22. doi:10.1186/1476-4598-10-93
42. Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139(5):871–890. doi:10.1016/j.cell.2009.11.007
43. Thiery JP, Sleeman JP. Complex networks orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell Biol. 2006;7(2):131–142. doi:10.1038/nrm1835
44. Savagner P, Boyer B, Valles AM, Jouanneau J, Thiery JP. Modulations of the epithelial phenotype during embryogenesis and cancer progression. Cancer Treat Res. 1994;71:229–249.
45. Tam WL, Weinberg RA. The epigenetics of epithelial-mesenchymal plasticity in cancer. Nat Med. 2013;19(11):1438–1449. doi:10.1038/nm.3336
46. Wang X, Wang B, Xie J, Hou D, Zhang H, Huang H. Melatonin inhibits epithelialtomesenchymal transition in gastric cancer cells via attenuation of IL1beta/NFkappaB/MMP2/MMP9 signaling. Int J Mol Med. 2018;42(4):2221–2228. doi:10.3892/ijmm.2018.3788
47. Cano A, Perez-Moreno MA, Rodrigo I, et al. The transcription factor snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat Cell Biol. 2000;2(2):76–83. doi:10.1038/35000025
48. Li VS, Ng SS, Boersema PJ, et al. Wnt signaling through inhibition of beta-catenin degradation in an intact Axin1 complex. Cell. 2012;149(6):1245–1256. doi:10.1016/j.cell.2012.05.002
49. Ueno K, Hirata H, Hinoda Y, Dahiya R. Frizzled homolog proteins, microRNAs and Wnt signaling in cancer. Int J Cancer. 2013;132(8):1731–1740. doi:10.1002/ijc.27746
50. Ripple MJ, Parker Struckhoff A, Trillo-Tinoco J, et al. Activation of c-Myc and cyclin D1 by JCV T-antigen and beta-catenin in colon cancer. PLoS One. 2014;9(9):e106257. doi:10.1371/journal.pone.0106257
51. Tanner B, Grimme S, Schiffer I, et al. Nuclear expression of apurinic/apyrimidinic endonuclease increases with progression of ovarian carcinomas. Gynecol Oncol. 2004;92(2):568–577. doi:10.1016/j.ygyno.2003.10.037
52. Turpaev KT. [Role of transcription factor AP-1 in integration of cellular signalling systems]. Mol Biol (Mosk). 2006;40(6):945–961.
53. Takeda K, Kinoshita I, Shimizu Y, et al. Clinicopathological significance of expression of p-c-Jun, TCF4 and beta-Catenin in colorectal tumors. BMC Cancer. 2008;8:328. doi:10.1186/1471-2407-8-172
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.