")
Back to Journals » International Journal of Nanomedicine » Volume 9 » Supplement 2
Undetactable levels of genotoxicity of SiO2 nanoparticles in in vitro and in vivo tests
Authors Kwon JY, Kim HL, Lee JY, Ju YH, Kim JS, Kang SH, Kim Y, Lee JK, Jeong J, Kim M, Meang E, Seo YR
Received 20 November 2013
Accepted for publication 7 February 2014
Published 15 December 2014 Volume 2014:9(Supplement 2) Pages 173—181
DOI https://doi.org/10.2147/IJN.S57933
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 6
Jee Young Kwon,1,* Hye Lim Kim,1,* Jong Yun Lee,2 Yo Han Ju,2 Ji Soo Kim,2 Seung Hun Kang,1 Yu-Ri Kim,3 Jong-Kwon Lee,4 Jayoung Jeong,4 Meyoung-Kon Kim,3 Eun Ho Maeng,2 Young Rok Seo1
1Department of Life Science, Institute of Environmental Medicine, Dongguk University, Seoul, 2Korea Testing and Research Institute, 3Department of Biochemistry and Molecular Biology, Korea University College of Medicine, Seoul, 4Toxicological Research Division, National Institute of Food and Drug Safety Evaluation, Ministry of Food and Drug Safety, Chungcheongbuk-do, Republic of Korea
*These authors contributed equally to this work and should be considered co-first authors
Background: Silica dioxide (SiO2) has been used in various industrial products, including paints and coatings, plastics, synthetic rubbers, and adhesives. Several studies have investigated the genotoxic effects of SiO2; however, the results remain controversial due to variations in the evaluation methods applied in determining its physicochemical properties. Thus, well characterized chemicals and standardized methods are needed for better assessment of the genotoxicity of nanoparticles.
Methods: The genotoxicity of SiO2 was evaluated using two types of well characterized SiO2, ie, 20 nm (-) charge (SiO2EN20(-)) and 100 nm (-) charge (SiO2EN100(-)). Four end point genotoxicity tests, ie, the bacterial mutation assay, in vitro chromosomal aberration test, in vivo comet assay, and in vivo micronucleus test, were conducted following the test guidelines of the Organization for Economic Cooperation and Development (OECD) with application of Good Laboratory Practice.
Results: No statistically significant differences were found in the bacterial mutation assay, in vitro chromosomal aberration test, in vivo comet assay, and in vivo micronucleus test when tested for induction of genotoxicity in both two types of SiO2 nanoparticles.
Conclusion: These results suggest that SiO2 nanoparticles, in particular SiO2EN20(-) and SiO2EN100(-), are not genotoxic in both in vitro and in vivo systems under OECD guidelines. Further, the results were generated in accordance with OECD test guidelines, and Good Laboratory Practice application; it can be accepted as reliable information regarding SiO2-induced genotoxicity.
Keywords: genotoxicity test, Organization for Economic Cooperation and Development test guideline, Good Laboratory Practice, silica dioxide
Introduction
The field of nanotechnology has been growing rapidly within various industries over the last decade. Various types of nanoparticles, including titanium oxide, zinc oxide, and silica dioxide (SiO2), are used in cosmetics, sports equipment, and building materials.1 Due to their widespread applications in various industries, use of nanoparticles continues to increase. However, nanoparticles may be hazardous to human health because of their unusual physicochemical properties, eg, small size, high surface to volume ratio, chemical composition, crystallinity, electronic properties, surface structure reactivity and functional groups, inorganic or organic coatings, solubility, shape, and aggregation behavior.2–4 The risk to human health arising from exposure to nanoparticles through ingestion, inhalation, and dermal absorption may increase as the applications of nanoparticles continue to increase.5–7
The Organization for Economic Cooperation and Development (OECD) has also paid attention to this problem and organized a special session in a joint meeting on the Potential Implications of Manufactured Nanomaterials for Human Health and Environmental Safety in 2005. The OECD established the Working Party on Manufactured Nanomaterials in 2006 for dealing with human and environmental safety issues associated with manufactured nanomaterials.8
SiO2 nanoparticles have been widely used in a variety of industrial fields, including the plastics, rubber, ceramics, coatings, adhesives, and medical industries.9,10 Numerous studies have reported the cytotoxicity, genotoxicity, and alterations in protein expression associated with SiO2 nanoparticles in both in vitro and in vivo systems.11,12 Further, it has been suggested that the genotoxicity caused by SiO2 nanoparticles may be due to proinflammatory effects, modification of chromatin structure, and liberation of DNase or a potent inducer of cytogenetic damage.13,14 However, these studies were controversial due to the slightly different physicochemical features involved in the synthesis, dispersion, and stability of nanoparticles in biological media.15–18 Nanoparticle-related studies reported that these differences were generated by the size and surface charge of the specific nanoparticles.19–22 The size of nanoparticles could influence their absorption, distribution, metabolism, and excretion.23–25 The hydrodynamic size distributions of nanoparticles could be also changed by surface charge and properties, affecting agglomeration. Moreover, the surface charge of nanoparticles could affect uptake and translocation within organisms.26,27 Several nanotoxicology reports have proposed that more detailed characterization of nanoparticles would yield a better understanding of their properties and interactions in vivo.28–30 Therefore, the physicochemical features of nanoparticles would be the critical factor in studying nanotoxicology.
In the present study, four genotoxicity tests were conducted with well characterized SiO2 nanoparticles 20 nm (SiO2EN20(−)) and 100 nm (SiO2EN100(−)) in size, each having a negative surface charge. In order to accurately evaluate toxicity, physicochemical characterization of nanoparticles and their behavior under physiological conditions were determined by following experimental protocols similar to those used in our previous study.31 For accurate evaluation of genotoxicity, four types of in vitro and in vivo genotoxicity tests, ie, the bacterial mutation assay, in vitro chromosomal aberration test, in vivo comet assay, and in vivo micronucleus test, were performed according to OECD guidelines32–34 or international validation study guideline35 with the Good Laboratory Practice (GLP).
Materials and methods
Animal care
All animals used in this study were cared for in accordance with the Guide for the Care and Use of Laboratory Animals issued by the Animal Care and Use Committee of the National Veterinary Research and Quarantine Service.
Preparation and characterization of nanoparticles
SiO2 colloids 20 nm (SiO2EN20(−)) and 100 nm (SiO2EN100(−)) in size were obtained from E&B Nanotech Co Ltd (Gyeonggi-do, Republic of Korea). The preparation process and physicochemical characterizations were conducted as described in a previous report.31 Briefly, a JSM-6700F field emission scanning electron microscope (JOEL Ltd, Tokyo, Japan) and Zetasizer NanoZS (Malvern Instruments Ltd, Malvern, UK) were used to determine the particle size, morphology, and surface charge of SiO2. Distilled water was used as the solvent control in this study.
Bacterial reverse mutation test
The bacterial reverse mutation test was conducted in compliance with the Korea Food and Drug Administration Notification No. 2009-116 testing guideline and OECD testing guideline 471.32 The tester strains used in this study were Ames Salmonella typhimurium TA98, TA100, TA1535, and TA1537, and Escherichia coli WP2uvrA in the absence and presence of metabolic activation system. All of the test strains were purchased from Molecular Toxicology Inc. (Boone, NC, USA). The metabolic activation system was prepared by mixing S9 metabolic activation (Molecular Toxicology Inc.) with Cofactor 1 from Wako Pure Chemical Industries Ltd (Osaka, Japan), giving a final concentration of 10% (volume/volume) S9. The tester strains were cultured in 2.5% nutrient broth No 2 (Oxoid Ltd, Basingstoke, UK) in a 37°C shaking incubator (120 rpm) for approximately 11 hours. The mutagenicity test was performed by mixing test substance and tester strains, which was cultured overnight in the presence and absence of the S9 mix. Next, the mixture was incubated in a water bath for 20 minutes at 37°C, mixed with top agar and a minimal amount of histidine-biotin (for S. typhimurium strains) or tryptophan (for E. coli strain), and then poured onto the surface of a gamma-ray sterile Falcon® Petri dish (Thermo Fisher Scientific, Waltham, MA, USA) containing about 15 mL of solidified bottom agar. The finished plates were incubated for 48–72 hours at 37°C. The number of revertant colonies was then counted. All plates were prepared in triplicate, and the results were tabulated as the mean ± standard deviation for each condition.
In vitro chromosomal aberration test
The in vitro chromosomal aberration test was performed according to the Korea Food and Drug Administration Notification No. 2009-116 testing guideline and OECD testing guideline 473.33 The clastogenicity of SiO2 nanoparticles was evaluated for its ability to induce chromosomal aberrations in Chinese hamster lung (CHL) fibroblast cells. A clonal subline of CHL cells was obtained from the American Type Culture Collection (Rockville, MD, USA). The karyotype of the CHL cells consisted of 25 chromosomes. The CHL cells were grown in Minimum Essential Eagle’s Medium, supplemented with 10% fetal bovine serum, 50 U/mL penicillin, and 50 μg/mL streptomycin (all from Gibco BRL Life Technologies Inc., Gaithersburg, MD, USA) at 37°C in a humidified atmosphere containing 5% CO2. Mitomycin C (CAS No. 50-07-7; Sigma-Aldrich Co., St Louis, MO, USA) was used as a positive control, both with and without the S9 mixture. After 22 hours of incubation, colcemid was added to the cultures at a final concentration of 0.2 μg/mL, and metaphase cells were harvested by trypsinization and centrifugation. The cells were swelled by adding hypotonic (0.075 M) KCl solution for 20 minutes at 37°C, and then washed three times in ice-cold fixative (methanol to glacial acetic acid, 3:1). A few drops of cell pellet suspension were dropped onto precleaned glass microscope slides and air-dried. The slides were stained with 5% Giemsa buffer solution (Thermo Fisher Scientific, Waltham, MA, USA). The number of cells with chromosomal aberrations was recorded on 200 well spread metaphases. The classification of aberration types referred to JEMS-MMS (Japanese Environmental Mutagen Society-Mammalian Mutagenesis Study group). Aberration frequencies, defined as aberrations observed, were divided by the number of cells counted, and were analyzed using Fisher’s exact test with Dunnett’s adjustment, and compared with results from the solvent controls.
In vivo alkaline comet assay
The alkaline (pH >13) comet assay was conducted according to international validation of the in vivo rodent alkaline comet assay for the detection of genotoxic carcinogens (version 14.2).35 Male Crl:CD (Sprague Dawley [SD]) rats (aged 6 weeks) were used for the in vivo alkaline comet assay. The SD rats were randomized into groups containing five rats each. After a 7-day acclimation period, the test substance was administered three times by gavage at 0, 24, and 45 hours. The test doses (500, 1,000, and 2,000 mg/kg) were selected by a range-finding experiment. Following the guideline, ethyl methanesulfonate was used for the positive control at a dose of 200 mg/kg. The liver and stomach were collected from each animal and maintained in cold mincing buffer (Mg2+-free and Ca2+-free Hanks’ Balanced Salt Solution (Gibco, Carlsbad, CA, USA) with 20 mM ethylenediaminetetraacetic acid (EDTA)-Na2 and 10% volume/volume dimethylsulfoxide (Sigma-Aldridge, St Louis, MO, USA). The liver was minced using fine scissors in cold mincing buffer. The stomach was opened and washed free of food using cold mincing buffer. The forestomach was discarded, and the glandular stomach was placed into cold mincing buffer and incubated on ice for 15–30 minutes. After incubation, the surface mucosa was gently scoured using a scraper. This layer was discarded subsequently, and the stomach epithelium was carefully scoured with a scraper to release the cells. The cell suspension was stored on ice for 15–30 seconds to allow large clumps to settle, and the supernatant was used to prepare the comet slides. A 10 μL aliquot of single cell suspension was mixed with 0.5% low melting agarose (Invitrogen, Carlsbad, CA, USA) and spread on comet assay slides (Travigen, Gaithersburg, MD, USA). The slides were immersed in cold lysis solution (2.5 M NaCl, 100 mM EDTA-Na2, 10 mM Tris-base, 10% dimethylsulfoxide, 1% Triton-X [pH10]) overnight. After this incubation, the slides were placed in electrophoresis solution (0.3 M NaOH, 1 mM EDTA [pH >13]) for 20 minutes to allow for unwinding of DNA. Electrophoresis was subsequently conducted at 25 V and 300 mA for 20 minutes. The slides were then immersed in neutralization solution (0.4 M Tris-base [pH 7.5]) for at least 5 minutes and then dehydrated with absolute ethanol to fix. The cells were stained with SYBR Gold (Invitrogen) according to the manufacturer’s specifications. The comet was observed via fluorescence microscopy (Nikon, Tokyo, Japan) at a magnification of 200× and analyzed by Comet Assay IV software (Perceptive Instruments Ltd, Bury St Edmunds, UK). For each sample (animal/tissue), 50 comets per slide were analyzed, with two slides scored per sample. A positive response is defined as a statistically significant change in the percent tail DNA in at least one dose group in comparison with the vehicle control value using Dunnett’s test (two-sided, P<0.05) as well as a statistically significant linear trend test (two-sided, P<0.05). The positive control should produce a statistically significant increase as determined by the Student’s t-test (one-sided, P<0.025).
In vivo micronucleus test
The in vivo micronucleus test was performed in compliance with the Korea Food and Drug Administration Notification No. 2009-116 testing guideline and OECD testing guideline 474.34 Out-bred 6–7-week-old mice of strain ICR were used in this study. The ICR mice were randomized into groups containing five mice each. The test substance was administered orally in three doses in volumes of 10 mL/kg. It was given twice with a 24-hour interval in between, and test subjects were sacrificed by cervical dislocation. Preparation and staining of bone marrow was carried out according to the method described by Schmid.36 In scoring the preparations, micronuclei were counted in polychromatic erythrocytes and separately in monochromatic erythrocytes. The rate of micronucleated cells, expressed as a percentage, was based on the total of polychromatic erythrocytes present in the scored optic fields. This method of scoring, which must always be followed where the test substance markedly influences the proliferation rate in bone marrow, prevented distortion of the results by the influx of peripheral blood into the damaged marrow. The scoring of micronucleated normocytes was used to recognize the presence of artifacts (which is rare in mouse preparations), which provided additional interesting information on the mode of action of the test substances. Generally, an incidence of more than one micronucleated normocyte per 1,000 polychromatic erythrocytes indicates an effect on cell stages, especially post-S-phase. The result of the statistical evaluation was deemed to be statistically significant when the P-value was less than 0.05. We used the Kruskal–Wallis H test and Dunnett’s test for differences in numbers of micronucleated polychromatic erythrocytes between the treated and negative control groups; the Mann–Whitney U test for differences in numbers of micronucleated polychromatic erythrocytes between the positive and negative control groups; analysis of variance and Dunnett’s test for differences in the polychromatic erythrocyte (PCE)/(PCE + normochromatic erythrocyte [NCE]) ratio between the treated and negative control groups; the Student’s t-test for differences in the PCE/(PCE + NCE) ratio between the positive and negative control groups; and analysis of variance and Dunnett’s test for comparison of animal body weight at the time of euthanasia.
Results
Characterization of nanoparticles
Particle size and surface charge are significant factors when determining the biological behavior of nanoparticles.31 The physicochemical properties of SiO2 nanoparticles were investigated in a previous study, as described briefly below.31
The average size and morphology of the SiO2 nanoparticles was investigated by field emission scanning electron microscopy. The results showed a monodispersed size distribution and spherical morphology with a mean size of ~33 nm and ~90 nm for SiO2EN20(−) and SiO2EN100(−), respectively. Dynamic light scattering analysis was determined to be 23±0.1 nm and 91.6±0.5 nm for SiO2EN20(−) and SiO2EN100(−), respectively. The surface charge on the SiO2 nanoparticles in distilled water suspension was determined to be negative (around −40 mV).
Bacterial reverse mutation test
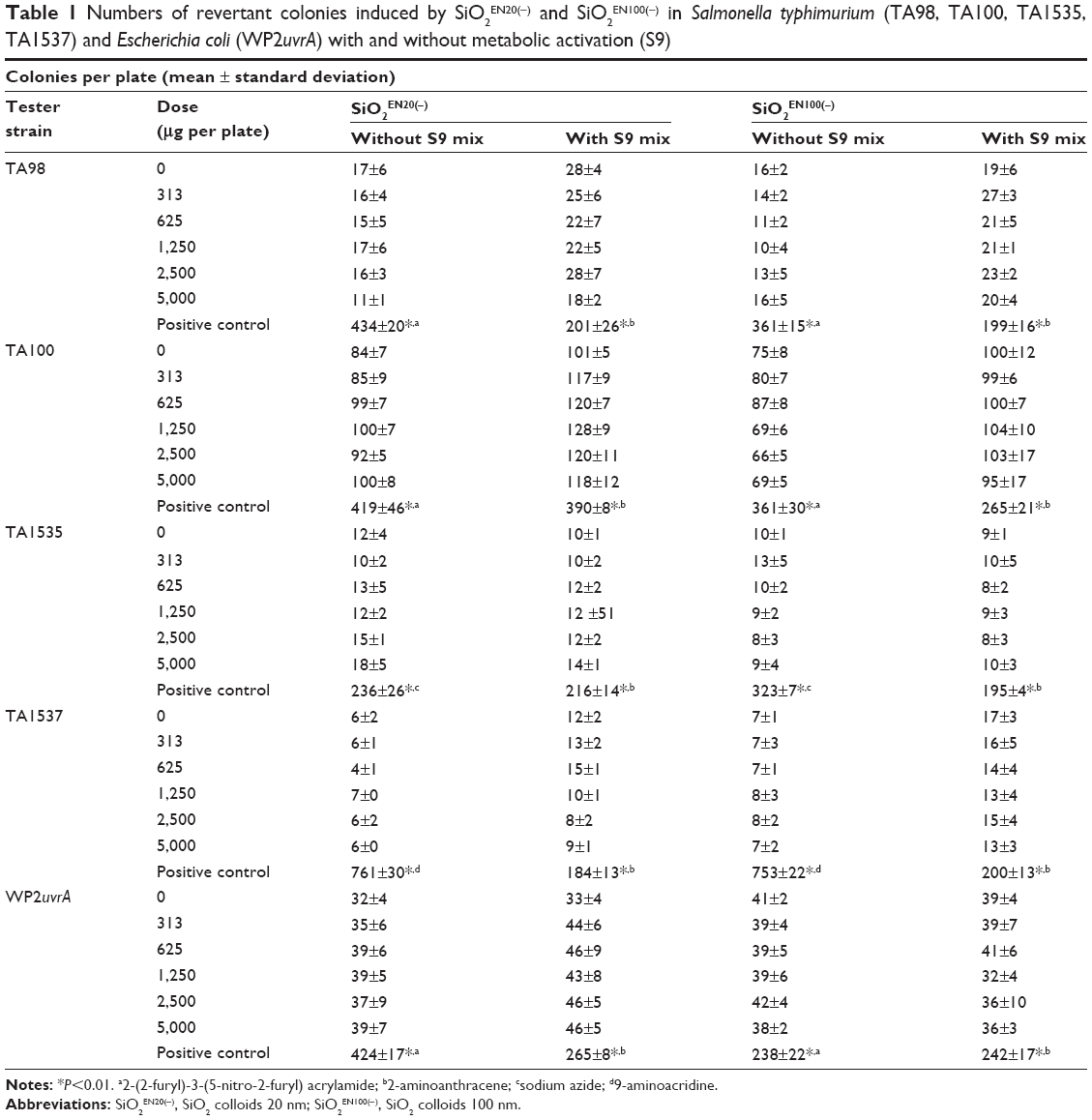
In the preliminary dose-ranging tests (data not shown), two kinds of SiO2 nanoparticles, ie, SiO2EN20(−) and SiO2EN100(−), were found to have a nontoxic effect in all tester strains of S. typhimurium (TA98, TA100, TA1535, and TA1537) and in E. coli WP2uvrA at a dose of 5,000 μg per plate in the presence and absence of S9. Based on the data from the preliminary dose-ranging tests, we selected 5,000 μg per plate as the highest dose. As shown in Table 1, none of the tester strains showed any increase in the number of revertant colonies in comparison with the solvent control (0 μg per plate) when the bacteria were treated with the two types of SiO2 nanoparticles at 313, 625, 1,250, 2,500, and 5,000 μg per plate, regardless of metabolic activation. On the other hand, the positive control group resulted in more revertant colonies than the solvent control or other SiO2 treatment groups.
|
Table 1 Numbers of revertant colonies induced by SiO2EN20(−) and SiO2EN100(−) in Salmonella typhimurium (TA98, TA100, TA1535, TA1537) and Escherichia coli (WP2uvrA) with and without metabolic activation (S9)Notes: *P<0.01. a2-(2-furyl)-3-(5-nitro-2-furyl) acrylamide; b2-aminoanthracene; csodium azide; d9-aminoacridine.Abbreviations: SiO2EN20(−), SiO2 colloids 20 nm; SiO2EN100(−), SiO2 colloids 100 nm. |
In vitro chromosome aberration test
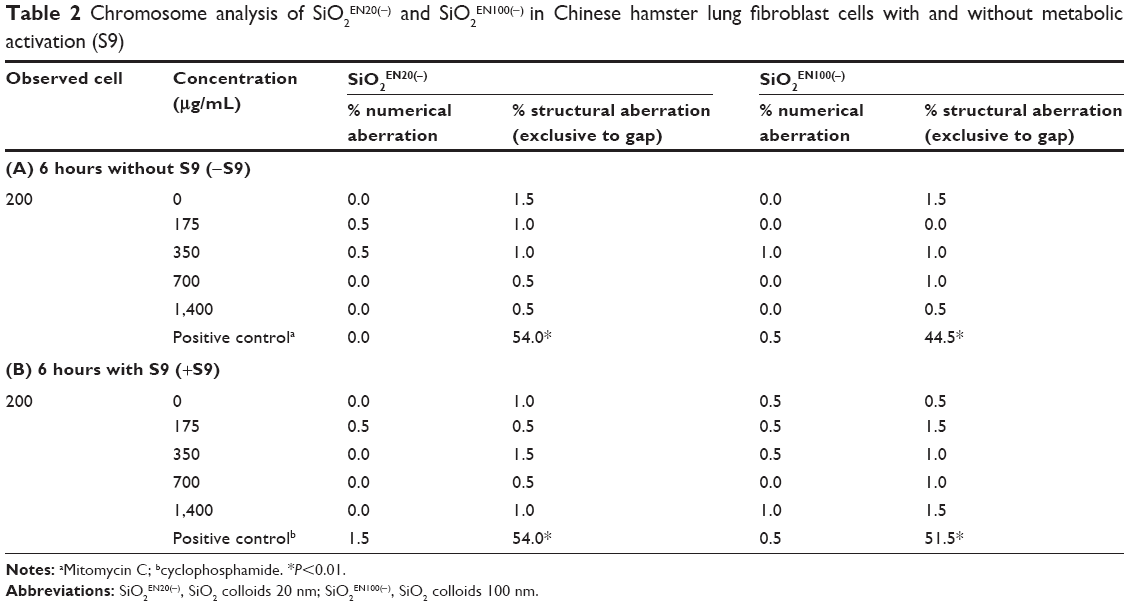
Initially, a dose-ranging test was performed to determine the test doses for use in the in vitro chromosome aberration test. The highest concentration was determined to be 1,400 μg/mL, as it showed less than 50% cytotoxicity in CHL cells (data not shown). The in vitro chromosome aberration test was conducted with SiO2EN20(−) and SiO2EN100(−) at concentrations of 175, 350, 700, and 1,400 μg/mL, both with and without metabolic activation (S9). Except for the positive control group, the metaphase arrested cells with structural aberrations were less than 5%. The results indicated no significant increase in SiO2 nanoparticles in the treatment groups in comparison with the solvent control at the four concentrations, regardless of metabolic activation (Table 2). On the other hand, the positive control group showed significantly increased structural aberrations in comparison with the solvent control and with the other SiO2 treatment groups.
|
Table 2 Chromosome analysis of SiO2EN20(−) and SiO2EN100(−) in Chinese hamster lung fibroblast cells with and without metabolic activation (S9)Notes: aMitomycin C; bcyclophosphamide. *P<0.01.Abbreviations: SiO2EN20(−), SiO2 colloids 20 nm; SiO2EN100(−), SiO2 colloids 100 nm. |
In vivo alkaline comet assay
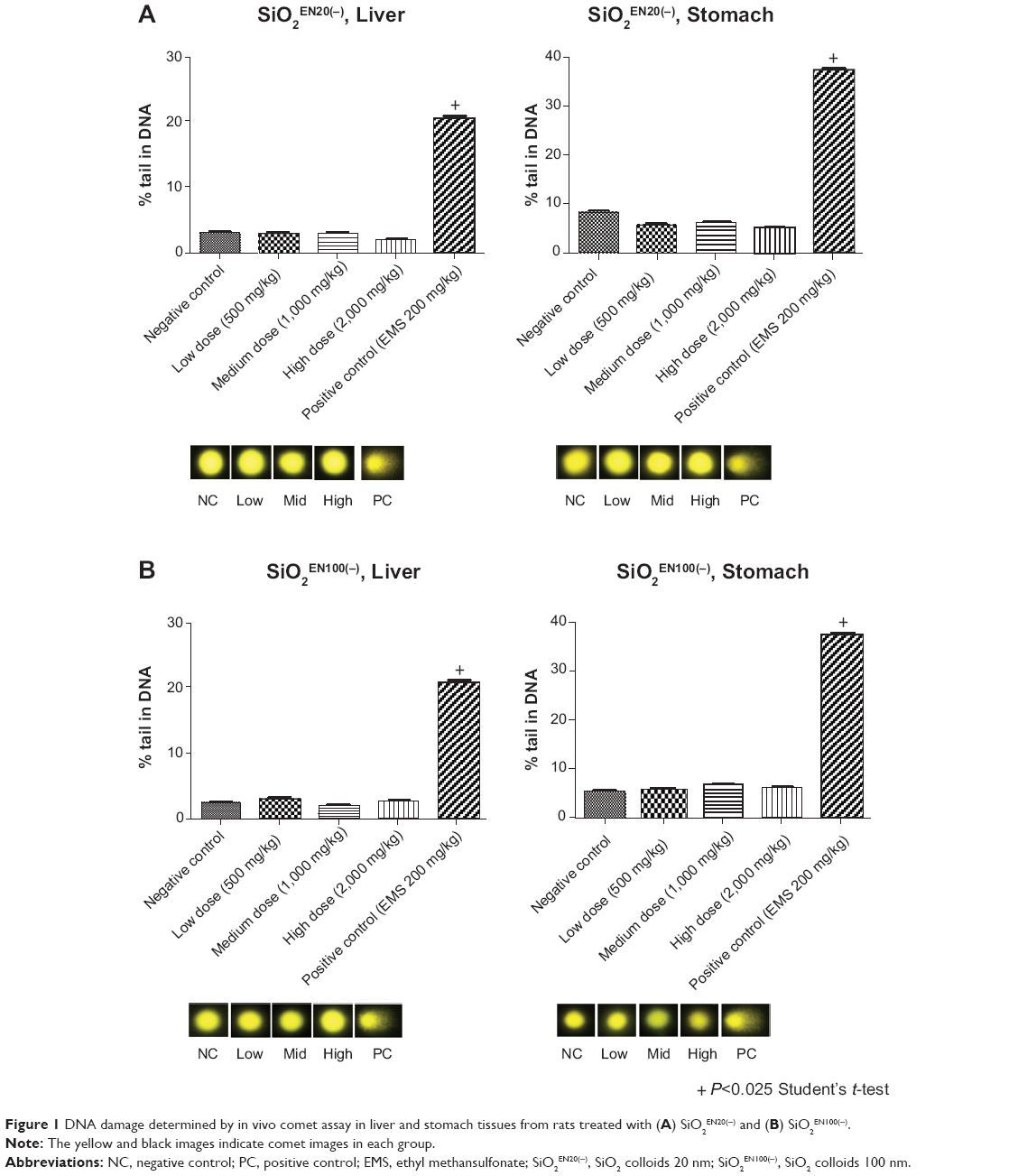
A dose-ranging study was conducted in SD rats (data not shown), and the test doses were determined as 500, 1,000, and 2,000 mg/kg body weight. DNA damage was measured by the in vivo alkaline comet assay, following the international validation study protocol (version 14.2).35 Our data showed no significant differences in percent tail DNA in single cells from the liver and stomach treated with SiO2EN20(−) (Figure 1A) and SiO2EN100(−) (Figure 1B). The positive control showed a significant increase in tail DNA intensity (percent tail DNA) in comparison with the negative control. Therefore, no genotoxic effect of SiO2 nanoparticles (SiO2EN20(−) and SiO2EN100(−)) was observed using the in vivo comet assay.
|
Figure 1 DNA damage determined by in vivo comet assay in liver and stomach tissues from rats treated with (A) SiO2EN20(−) and (B) SiO2EN100(−). |
In vivo micronucleus test
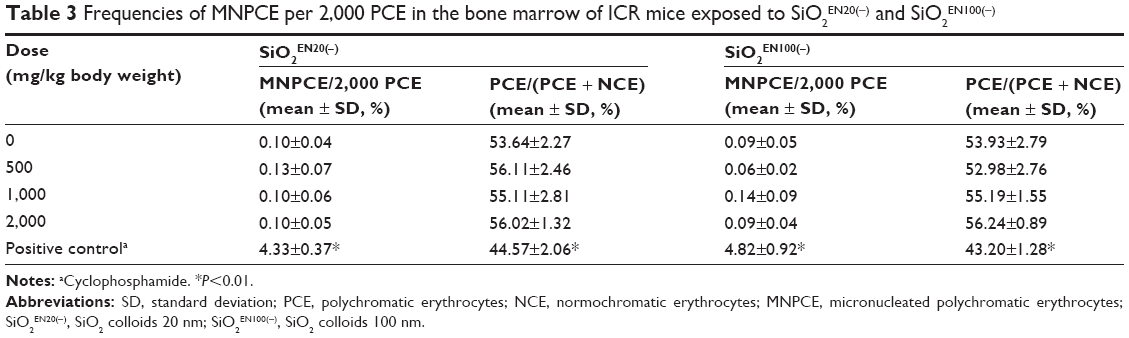
In the preliminary dose-ranging test, acute oral toxicity, death, or apparent abnormality in appearance could not be determined in mice at doses ranging from 500 mg/kg to 2,000 mg/kg body weight. The PCE/(NCE + PCE) ratios were used as an index of bone marrow cytotoxicity. The ratios did not show any significant difference in the SiO2 treatment groups in comparison with the solvent control group, for either size of SiO2 nanoparticle. The micronucleated PCE frequencies were not statistically significant and did not yield any dose-dependent pattern among the three treatment groups in comparison with the solvent control group for either size of SiO2 nanoparticle. On the other hand, the positive control group showed a significantly increased micronucleated PCE frequency in comparison with the solvent control and SiO2 treatment groups (Table 3). Therefore, both types of SiO2 nanoparticles were regarded to have no genotoxic or cytotoxic potential using the current in vivo system.
|
Table 3 Frequencies of MNPCE per 2,000 PCE in the bone marrow of ICR mice exposed to SiO2EN20(−) and SiO2EN100(−)Notes: aCyclophosphamide. *P<0.01.Abbreviations: SD, standard deviation; PCE, polychromatic erythrocytes; NCE, normochromatic erythrocytes; MNPCE, micronucleated polychromatic erythrocytes; SiO2EN20(−), SiO2 colloids 20 nm; SiO2EN100(−), SiO2 colloids 100 nm. |
Discussion
Nanotoxicity studies have been necessary, although their results are still controversial due to variations in the physicochemical properties of the materials under investigation.19–21,37–39 Further, their results could differ according to the protocol used for evaluation. Hence, well characterized nanoparticles and approved methods are necessary when conducting nanotoxicity studies. In this study, nanogenotoxicity was investigated using well characterized SiO2 nanoparticles, ie, SiO2EN20(−) and SiO2EN100(−). Characterization of these nanoparticles and their behavior was done under physiological conditions, as described in our previous study.31 Next, four different end point genotoxic tests, ie, the bacterial mutation assay, in vitro chromosomal aberration test, in vivo alkaline comet assay, and in vivo micronucleus test, were conducted in accordance with OECD guidelines and an international validation study protocol using the GLP system.
The bacterial mutation assay is used to evaluate the mutagenicity of a chemical compound.1 It is also reported to be an essential test within the current battery of assays required for evaluation of genotoxicity.40 To date, there have been a few reports on the mutagenicity of nanoparticles such as aluminum oxide, cobalt oxide, titanium oxide, and zinc oxide based on the bacterial mutation assay. Hence, we performed this assay using strains of S. typhimurium TA98, TA100, TA1535, and TA1537, and E. coli WP2uvrA, with and without metabolic activation by S9 mixture.41–43 It has recently been reported that no mutagenic potential is observed in the Ames test with and without metabolic activation.44–46 In addition, Li et al investigated the genotoxic potential of exfoliated silicate nanoclay.47 Our data also showed no significantly increased mutagenicity in any of the strains exposed to SiO2EN20(−) and SiO2EN100(−) with and without S9 (Table 1).
The in vitro chromosomal aberration test is another standard genotoxicity test, and is commonly used to observe variations in chromosomal structure induced by toxic chemicals.33 Our data show that SiO2EN20(−) and SiO2EN100(−) did not induce any structural chromosomal aberrations in CHL cells, with or without S9 mixture (Table 2). Another previous report showed no induction of chromosomal aberrations in mammalian cells.44–46 Although different types of SiO2 were used in the previous studies, similar results were obtained. Hence, SiO2 nanoparticles might not induce clastogenesis in mammalian cells. In vitro genotoxicity tests could afford a better understanding of their genotoxic potential by investigating various compounds. The in vivo alkaline comet assay and in vivo micronucleus test were included in the current study in order to conduct and evaluate genotoxicity tests accurately. The comet assay, a single cell gel electrophoresis assay, has been widely used for detection of DNA damage due to its simplicity, low cost, and high sensitivity.48 The in vivo comet assay has been regarded as a potential replacement for the in vivo rodent hepatocyte unscheduled DNA synthesis assay, and the protocol for the in vivo comet assay has been standardized and published by an international expert group. In the present study, the in vivo comet assay was conducted using SiO2EN20(−) and SiO2EN100(−) and followed the standard protocol, revealing no genotoxic effect of SiO2EN20(−) and SiO2EN100(−) in rat liver and stomach cells (Figure 1A and 1B). Even though the majority of the studies have reported negative results for the genotoxicity of SiO2,11,17,47 a few investigations have indicated otherwise.49 Downs et al investigated the genotoxic effects of exposure to three consecutive intravenous injections of SiO2 nanoparticles by in vivo comet assay in rat liver and lung tissues, and observed the induction of DNA damage by SiO2.50 Hence, the different administration route can lead to different results for genotoxicity evaluation.
The micronucleus test was used to evaluate genotoxicity by scoring the number of micronuclei under chemical exposure. Regarding the genotoxicity induced by SiO2 nanoparticles, several publications have reported on in vitro micronucleus tests.11,15,47,51,52 In contrast, few in vivo micronucleus tests have been performed. Downs et al conducted in vivo micronucleus testing of SiO2 nanoparticles, where they analyzed micronuclei in rat blood.50 In the present study, in vivo micronucleus testing was performed with rat bone marrow cells for the first time to determine whether SiO2EN20(−) and SiO2EN100(−) could induce chromosome abnormalities or mitotic apparatus in mouse model. Based on the frequencies of micronucleated PCE, no genotoxic effect of SiO2 was found (Table 3). Nanotoxicology study results can be influenced by physicochemical properties, and the contrast between our results and the previous data could be due to the different administration methods used or different organs investigated, as mentioned above.
Conclusion
In the present study, the genotoxic effects of SiO2EN20(−) and SiO2EN100(−) were elucidated using four genotoxicity tests, ie, the bacterial mutation assay, in vitro chromosomal aberration test, in vivo comet assay, and in vivo micronucleus test, under standardized protocols with GLP system. Although the different exposure routes from our study can induce SiO2 genotoxicity in different organs in in vivo systems, our data suggest that SiO2EN20(−) and SiO2EN100(−) are not genotoxic substances based on the OECD test guidelines.
Acknowledgment
This research was supported by a grant (10182MFDS991) from the Ministry of Food and Drug Safety in 2011–2012.
Disclosure
The authors report no conflicts of interest in this work.
References
Ng CT, Li JJ, Bay BH, Yung LY. Current studies into the genotoxic effects of nanomaterials. J Nucleic Acids. 2010;2010:947859. | ||
Schrand AM, Rahman MF, Hussain SM, Schlager JJ, Smith DA, Syed AF. Metal-based nanoparticles and their toxicity assessment. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2010;2(5):544–568. | ||
Yang H, Liu C, Yang D, Zhang H, Xi Z. Comparative study of cytotoxicity, oxidative stress and genotoxicity induced by four typical nanomaterials: the role of particle size, shape and composition. J Appl Toxicol. 2009;29(1):69–78. | ||
Donaldson K, Brown D, Clouter A, et al. The pulmonary toxicology of ultrafine particles. J Aerosol Med. 2002;15(2):213–220. | ||
Rim KT, Kim SJ, Song SW, Park JS. Effect of cerium oxide nanoparticles to inflammation and oxidative DNA damages in H9c2 cells. Mol Cell Toxicol. 2012;8(3):271–280. | ||
Rim KT, Kim SJ, Han JH, Kang MG, Kim JK, Yang JS. Effects of carbon black to inflammation and oxidative DNA damages in mouse macrophages. Mol Cell Toxicol. 2011;7(4):415–423. | ||
Kim YJ, Yang SI, Ryu JC. Cytotoxicity and genotoxicity of nano-silver in mammalian cell lines. Mol Cell Toxicol. 2010;6(2):119–125. | ||
OECD environment, health and safety publications series on the safety of manufactured nanomaterials. 2010; No. 27, Paris. Available from: http://www.nanotechia.org/node/17752. Accessed July 16, 2014. | ||
Rothen-Rutishauser BM, Schurch S, Haenni B, Kapp N, Gehr P. Interaction of fine particles and nanoparticles with red blood cells visualized with advanced microscopic techniques. Environ Sci Technol. 2006;40(14):4353–4359. | ||
Choi JE, Park YH, Lee EY, et al. A safety assessment of phototoxicity and sensitization of SiO2 nanoparticles. Mol Cell Toxicol. 2011;7(2):171–176. | ||
Wang JJ, Sanderson BJ, Wang H. Cytotoxicity and genotoxicity of ultrafine crystalline SiO2 particulate in cultured human lymphoblastoid cells. Environ Mol Mutagen. 2007;48(2):151–157. | ||
Yang X, Liu J, He H, et al. SiO2 nanoparticles induce cytotoxicity and protein expression alteration in HaCaT cells. Part Fibre Toxicol. 2010;7:1. | ||
Eom HJ, Choi J. Oxidative stress of silica nanoparticles in human bronchial epithelial cell, Beas-2B. Toxicol In Vitro. 2009;23(7):1326–1332. | ||
Mohamed BM, Verma NK, Prina-Mello A, et al. Activation of stress-related signalling pathway in human cells upon SiO2 nanoparticles exposure as an early indicator of cytotoxicity. J Nanobiotechnology. 2011;9:29. | ||
Sergent JA, Paget V, Chevillard S. Toxicity and genotoxicity of nano- SiO2 on human epithelial intestinal HT-29 cell line. Ann Occup Hyg. 2012;56(5):622–630. | ||
Doak SH, Manshian B, Jenkins GJ, Singh N. In vitro genotoxicity testing strategy for nanomaterials and the adaptation of current OECD guidelines. Mutat Res. 2012;745(1–2):104–111. | ||
Barnes CA, Elsaesser A, Arkusz J, et al. Reproducible comet assay of amorphous silica nanoparticles detects no genotoxicity. Nano Lett. 2008;8(9):3069–3074. | ||
Chu Z, Huang Y, Li L, Tao Q, Li Q. Physiological pathway of human cell damage induced by genotoxic crystalline silica nanoparticles. Biomaterials. 2012;33(30):7540–7546. | ||
Gurr JR, Wang AS, Chen CH, Jan KY. Ultrafine titanium dioxide particles in the absence of photoactivation can induce oxidative damage to human bronchial epithelial cells. Toxicology. 2005;213(1–2):66–73. | ||
Pan Y, Leifert A, Ruau D, et al. Gold nanoparticles of diameter 1.4 nm trigger necrosis by oxidative stress and mitochondrial damage. Small. 2009;5(18):2067–2076. | ||
Carlson C, Hussain SM, Schrand AM, et al. Unique cellular interaction of silver nanoparticles: size-dependent generation of reactive oxygen species. J Phys Chem B. 2008;112(43):13608–13619. | ||
Park YH, Bae HC, Jang YS, et al. Effect of the size and surface charge of silica nanoparticles on cutaneous toxicity. Mol Cell Toxicol. 2013;9(1):67–74. | ||
Renwick LC, Donaldson K, Clouter A. Impairment of alveolar macrophage phagocytosis by ultrafine particles. Toxicol Appl Pharmacol. 2001;172(2):119–127. | ||
Borm PJ, Robbins D, Haubold S, et al. The potential risks of nanomaterials: a review carried out for ECETOC. Part Fibre Toxicol. 2006;3:11. | ||
Choi HS, Liu W, Misra P, et al. Renal clearance of quantum dots. Nat Biotechnol. 2007;25(10):1165–1170. | ||
Hoshino A, Fujioka K, Oku T, et al. Physicochemical properties and cellular toxicity of nanocrystal quantum dots depend on their surface modification. Nano Lett. 2004;4(11):2163–2169. | ||
Lockman PR, Koziara JM, Mumper RJ, Allen DD. Nanoparticle surface charges alter blood-brain barrier integrity and permeability. J Drug Target. 2004;12(9–10):635–641. | ||
Oberdorster G, Maynard A, Donaldson K, et al. Principles for characterizing the potential human health effects from exposure to nanomaterials: elements of a screening strategy. Part Fibre Toxicol. 2005;2:8. | ||
Powers KW, Brown SC, Krishna VB, Wasdo SC, Moudgil BM, Roberts SM. Research strategies for safety evaluation of nanomaterials. Part VI. Characterization of nanoscale particles for toxicological evaluation. Toxicol Sci. 2006;90(2):296–303. | ||
Powers KW, Palazuelos M, Moudgil BM, Roberts SM. Characterization of the size, shape, and state of dispersion of nanoparticles for toxicological studies. Nanotoxicology. 2007;1(1):42–51. | ||
Kim K-M, Kim H-M, Choi M-H, et al. Colloidal properties of surface coated colloidal silica nanoparticles in aqueous and physiological solutions. Sci Adv Mater. 2014;6:1573–1581. | ||
The Organisation for Economic Co-operation and Development (OECD). OECD guideline for testing of chemicals, Bacterial reverse mutation test. Paris: The Organisation for Economic Co-operation and Development (OECD); 1997. Available from: http://www.oecd.org/chemicalsafety/risk-assessment/1948418.pdf. Accessed July 16, 2014. | ||
The Organisation for Economic Co-operation and Development (OECD). OECD guideline for testing of chemicals, In vitro mammalian chromosome aberration test. The Organisation for Economic Co-operation and Development (OECD); 1997. Available from: http://www.oecd.org/chemicalsafety/risk-assessment/1948434.pdf. Accessed July 16, 2014. | ||
The Organisation for Economic Co-operation and Development (OECD). OECD guideline for testing of chemicals, Mammalian erythrocyte micronucleus test. The Organisation for Economic Co-operation and Development (OECD); 1997. Available from: http://www.oecd.org/chemicalsafety/risk-assessment/1948442.pdf. Accessed July 16, 2014. | ||
The Validation Management Team. International validation of the in vivo rodent alkaline comet assay for the detection of genotoxic carcinogens (version 14.2), 2009. Available from: http://cometassay.com/JaCVAM.pdf. Accessed May 22, 2014. | ||
Schmid W. The micronucleus test. Mutat Res. 1975;31(1):9–15. | ||
Goodman CM, McCusker CD, Yilmaz T, Rotello VM. Toxicity of gold nanoparticles functionalized with cationic and anionic side chains. Bioconjug Chem. 2004;15(4):897–900. | ||
Oh WK, Kim S, Choi M, et al. Cellular uptake, cytotoxicity, and innate immune response of silica-titania hollow nanoparticles based on size and surface functionality. ACS Nano. 2010;4(9):5301–5313. | ||
Ryman-Rasmussen JP, Riviere JE, Monteiro-Riviere NA. Surface coatings determine cytotoxicity and irritation potential of quantum dot nanoparticles in epidermal keratinocytes. J Invest Dermatol. 2007;127(1):143–153. | ||
Kirkland D, Reeve L, Gatehouse D, Vanparys P. A core in vitro genotoxicity battery comprising the Ames test plus the in vitro micronucleus test is sufficient to detect rodent carcinogens and in vivo genotoxins. Mutat Res. 2011;721(1):27–73. | ||
Pan X, Redding JE, Wiley PA, Wen L, McConnell JS, Zhang B. Mutagenicity evaluation of metal oxide nanoparticles by the bacterial reverse mutation assay. Chemosphere. 2010;79(1):113–116. | ||
Yoshida R, Kitamura D, Maenosono S. Mutagenicity of water-soluble ZnO nanoparticles in Ames test. J Toxicol Sci. 2009;34(1):119–122. | ||
Kumar A, Pandey AK, Singh SS, Shanker R, Dhawan A. Cellular uptake and mutagenic potential of metal oxide nanoparticles in bacterial cells. Chemosphere. 2011;83(8):1124–1132. | ||
European Centre for Ecotoxicology and Toxicology of Chemicals. Synthetic Amorphous Silica (CAS No. 7631-86-9). JACC No. 51. Brussels. 2006. | ||
US Environmental Protection Agency. Screening-level hazard characterization of silane, dichlorodimethyl-, reaction product with silica (CASRN 68611-44-9), 2011. Available from: http://www.epa.gov/chemrtk/hpvis/hazchar/68611-44-9_dimethyldichlorosilane%20silica_June_2011.pdf. Accessed May 22, 2014. | ||
Organisation for Economic Cooperation and Development. SIDS dossier on synthetic amorphous silica and silicates, 2004. Available from: http://www.chem.unep.ch/irptc/sids/oecdsids/Silicates.pdf. Accessed May 22, 2014. | ||
Li PR, Wei JC, Chiu YF, Su HL, Peng FC, Lin JJ. Evaluation on cytotoxicity and genotoxicity of the exfoliated silicate nanoclay. ACS Appl Mater Interfaces. 2010;2(6):1608–1613. | ||
Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp Cell Res. 1988;175(1):184–191. | ||
Choi HS, Kim Y-J, Song M, Song M-K, Ryu J-C. Genotoxicity of nano-silica in mammalian cell lines. Toxicol Environ Health Sci. 2010;3(1):7–13. | ||
Downs TR, Crosby ME, Hu T, et al. Silica nanoparticles administered at the maximum tolerated dose induce genotoxic effects through an inflammatory reaction while gold nanoparticles do not. Mutat Res. 2012;745(1–2):38–50. | ||
Park MV, Verharen HW, Zwart E, et al. Genotoxicity evaluation of amorphous silica nanoparticles of different sizes using the micronucleus and the plasmid lacZ gene mutation assay. Nanotoxicology. 2011;5(2):168–181. | ||
Uboldi C, Giudetti G, Broggi F, Gilliland D, Ponti J, Rossi F. Amorphous silica nanoparticles do not induce cytotoxicity, cell transformation or genotoxicity in Balb/3T3 mouse fibroblasts. Mutat Res. 2012;745(1–2):11–20. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.