Back to Journals » OncoTargets and Therapy » Volume 13
Tumor Necrosis Factor Alpha-Induced Proteins in Malignant Tumors: Progress and Prospects
Received 6 December 2019
Accepted for publication 4 March 2020
Published 20 April 2020 Volume 2020:13 Pages 3303—3318
DOI https://doi.org/10.2147/OTT.S241344
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Fang Guo,1,2 Yuan Yuan1
1Liaoning Provincial Education Department, Tumor Etiology and Screening Department of Cancer Institute and General Surgery, The First Hospital of China Medical University, and Key Laboratory of Cancer Etiology and Prevention (China Medical University), Shenyang City, Liaoning Province, People’s Republic of China; 2Department of Oncology, PLA Cancer Center, General Hospital of Northern Theater Command, Shenyang City, Liaoning Province, People’s Republic of China
Correspondence: Yuan Yuan
Tumor Etiology and Screening Department of Cancer Institute and General Surgery, The First Hospital of China Medical University, Shenyang City, Liaoning Province 110001, People’s Republic of China
Tel/Fax +86-24-28851539
Email [email protected]
Abstract: Tumor necrosis factor (TNF) is the first cytokine used in tumor biotherapy, but TNF-related drugs are limited by the lack of specific targets. Tumor necrosis factor alpha-induced proteins (TNFAIPs), derived from TNF, is a protein family and participates in proliferation, invasion and metastasis of tumor cells. In order to better understand biological functions and potential roles of TNFAIPs in malignant tumors, this paper in the form of “Gene–Protein–Tumor correlation” summarizes the biological characteristics, physiological functions and mechanisms of TNFAIPs by searching National Center of Biotechnology Information, GeneCards, UniProt and STRING databases. The relationship between TNFAIPs and malignant tumors is analyzed, and protein–protein interaction diagram in members of TNFAIPs is drawn based on TNF for the first time. We find that TNF as a key factor is related to TNFAIP1, TNFAIP3, TNFAIP5, TNFAIP6, TNFAIP8 and TNFAIP9, which can be directly involved in activating TNFAIP1, TNFAIP5, TNFAIP8 and TNFAIP9. We confirm that the mechanism of TNFAIP1, TNFAIP2 and TNFAIP3 inducing tumors may be related to NF-κB signaling pathway, but the mechanism of tumor induction by other members of TNFAIPs is not clear. In the future, translational studies are needed to explore the mechanisms of TNF-TNFAIPs-tumors.
Keywords: tumor necrosis factor alpha-induced proteins, malignant tumor, biological characteristics, physiological functions
Introduction
Tumor necrosis factor (TNF) is one of the well-known bioactive factors. In 1975, Carswell et al found that there were a class of substances in serum of mice, which could kill tumor cells and cause hemorrhagic necrosis of tumor tissues, called TNF or Cachectin.1 Previous studies showed that the sources of TNF were diverse, and it could be released by multiple cells in mammals, such as macrophages, fibroblasts, lymphocytes, endothelial cells, smooth muscle cells and epidermal cells.2 According to genetic structure, TNF is divided into TNF-α produced by activated macrophages and TNF-β rooted in activated T cells and NK cells.3 Both TNF-α and TNF-β may encode different proteins that perform specific functions. Further studies have found that the biological activity of TNF-α dominates in nature and is closely related to malignant tumors and autoimmune diseases. However, how to explore TNF-related specific targets has always been a hot and difficult topic.
Tumor necrosis factor alpha-induced proteins (TNFAIPs) is a protein family that is induced by TNF-α. The members of TNFAIPs family mainly include TNFAIP1, TNFAIP2, TNFAIP3, TNFAIP4, TNFAIP5, TNFAIP6, TNFAIP8 and TNFAIP9, with the exception of TNFAIP7 which has been removed from the National Center of Biotechnology Information (NCBI) databases by HUGO Gene Nomenclature Committee (HGNC). Recently, we compare the similarity of amino acid sequences among members of TNFAIPs' family by using UniProt database (https://www.uniprot.org) to confirm amino acid sequences, CLUSTALW software (https://www.genome.jp/tools-bin/clustalw) for amino acid sequence comparison, ESPript 3.0 (http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi) and Adobe PhotoShop CS6 software for data processing and image beautification. Our study confirmed that all members of TNFAIPs' family can be induced by TNF, but they have less than 15% amino acid sequence similarity with each other, indicating that they may have different biological characteristics (Supplementary Figure 1). Previous studies have found that the expression of TNFAIPs may regulate cell differentiation, apoptosis, immune response, inflammatory response, signal transduction and material transport. They also play important roles in diseases, especially malignant tumors.4–6 However, there are only reports on individual member of TNFAIPs family until now, and lack of researches reveals the internal relationship between TNFAIPs' members. Understanding the basic characteristics and physiological functions of TNFAIPs is conducive to discovering the TNF-related mechanism of tumor development.
This paper summarizes the basic characteristics and physiological functions of TNFAIPs by NCBI, GeneCards, UniProt and STRING databases, and analysis the relationship between TNFAIPs and malignant tumors. The aim is to understand the characteristics and mechanisms of TNFAIPs in Homo sapiens and to elucidate the role of TNFAIPs in the development of malignant tumors.
Basic Characteristics and Physiological Functions of TNFAIPs
TNFAIP1
Basic Characteristics of TNFAIP1
TNFAIP1 is a highly conservative single-copy gene found in human umbilical vein endothelial cells. Protein encoded by this gene is identified as the first protein produced by TNF-α and is similar to potassium channel tetramerisation domain containing 10 (KCTD10) and potassium channel tetramerisation domain containing 13 (KCTD13) which belongs to a member of polymerase delta-interacting protein 1 (PDIP1) family.7
Physiological Functions and Mechanisms of TNFAIP1 Protein
In recent years, TNFAIP1 has been shown to participate in DNA synthesis, damage and repair, induce differentiation and apoptosis, and interfere signal transduction.7–9 In addition, the multiple upstream regulatory sequences and micro ribonucleic acid (miRNA) are revealed to be involved in transcriptional regulation of TNFAIP1.10–14 The physiological mechanism of TNFAIP1 is not yet clear. Previous studies have found that casein kinase 2 (CK2) could phosphorylation of TNFAIP1 in vivo and vitro, and the interaction between TNFAIP1 gene and proliferating cell nuclear antigen (PCNA) can be enhanced by interfering with the distribution of genes in nucleus, suggesting that the phosphorylated TNFAIP1 may have some physiological functions.9 However, other researchers have a different view and have published studies confirming that TNFAIP1 can interact directly with PCNA and DNA polymerase delta subgroup. Otherwise, TNFAIP1 may be associated with KCTD10 and enhanced the ubiquitination degradation of KCTD10 protein and inhibited the transcriptional activity of nuclear factor kappa B (NF-κB) receptor and activating protein-1 (AP-1) receptor.7
TNFAIP2
Basic Characteristics of TNFAIP2
TNFAIP2 locates in human chromosome 14q32.32 and codes a protein which has 654 amino acids and molecular weight of 73 kDa. TNFAIP2 belongs to the SEC6 family and is originally thought to be a novel gene induced by TNF-α in human endothelial cells.15 Previous studies have shown that TNFAIP2 is expressed in embryonic liver and kidney, male mature germ cells, hematopoietic and lymphoid tissues.16
Physiological Functions and Mechanisms of TNFAIP2
TNFAIP2 is involved in a variety of physiological processes during organ formation and development, including inflammation, angiogenesis, cell differentiation, bone marrow tissue production, sperm production and so on.16 In vitro experiments, TNFAIP2 used to be differentially expressed in blood vessels, and it maybe an inhibitor of NF-κB or an auto-inhibitor of TNF-α response.17 TNFAIP2 can also serve as a target gene for retinoic acid and play an important role in inducing apoptosis in acute promyelocytic leukemia and multiple tumors.18 Some studies have demonstrated that TNFAIP2 can regulate inflammation and angiogenesis, induce apoptosis, promote cell proliferation, adhesion, migration and invasion, and mediate the formation of membrane nanotubes by NF-κB signaling pathway, Retinoic acid signaling pathway and Kruppel-like factor 5 (KLF5) signaling pathway.19 In 2014, researchers confirmed that EBV oncoprotein and latent membrane protein 1 (LMP1) could transcriptionally induce TNFAIP2 expression by NF-κB signaling pathway, and up-regulation of TNFAIP2 could promote cell migration in nasopharyngeal carcinoma (NPC).20 In breast cancer, researchers discovered that TNFAIP2 interacted with two small GTPases Rac1 and Cdc42, thereby increasing their activities to change actin cytoskeleton and cell morphology. KLF5 had functions such as promotes cancer cell proliferation, migration and invasion through directly binding to the TNFAIP2 gene promoter and activated the transcription. Therefore, TNFAIP2 might be a direct KLF5 target gene.16 TNFAIP2 was also a microfold cells-associated gene which could be induced by adding cytokine receptor activator of NF-κB ligand to enteroid structure.21 Recently, Scholz et al identified potent translation-inhibitory upstream open reading frames (uORFs) in the transcript leader sequence (TLS) of TNFAIP2, and TNFAIP2 might be a novel target of uORF-mediated translational regulation.22 Moreover, TNFAIP2 expression could induce epithelial-to-mesenchymal transition (EMT) and confer platinum resistance in urothelial cancer cells, suggesting that there was a correlation between TNFAIP2 and EMT after cisplatin treatment, and the expression level of MTDH in tumor cells could affect TNFAIP2-derived EMT acquisition.23
TNFAIP3
Basic Characteristics of TNFAIP3
TNFAIP3 locates in chromosome 6q23.3 and had 12 exons. TNFAIP3 protein as a zinc finger protein and ubiquitin shearing enzyme can suppress activated NF-κB signal pathway and induce apoptosis of TNF by activating ubiquitin ligase and ubiquitinase.24 Some studies showed that TNFAIP3 could play an important role in immune and inflammatory responses mediated by cytokines.24–26
Physiological Functions and Mechanisms of TNFAIP3
TNFAIP3 is a ubiquitin modification enzyme and a negative regulator which can negatively regulate NF-κB signaling pathway through epigenetic silencing, deletion, somatic cell mutation and other inactivation. It is well known that NF-κB signaling pathway plays an important role in body immunity and inactivated NF-κB signaling pathway can lead to autoimmune or inflammatory diseases. In addition, TNFAIP3 re-expression can inhibit proliferation, decrease expression of target genes in NF-κB signaling pathway, increase cytotoxicity and induce apoptosis in cells lacking TNFAIP3.25 At present, there are few studies on the mechanism of TNFAIP3. Wenzl et al found that TNFAIP3 rs143002189 polymorphism was the substitution of isoleucine (Met) by isoleucine (Ile) at 788 sites of ZnF7 amino acid chain, which altered the structure and appearance of protein leading to tumors.27
TNFAIP4
Basic Characteristics of TNFAIP4
TNFAIP4, commonly known as Ephrin A1 (EFNA1), is located in chromosome 1q22 and has four exons and belongs to the receptor tyrosine kinase ligand family (Ephrins). Ephrins and receptors make up the largest subfamily of tyrosine kinase proteins and can play an important role in human growth, especially in regulating the nervous system and erythrocyte.28 According to structure and sequence, Ephrins is divided into Ephrin-A (EFNA) and Ephrin-B (EFNB). EFNA is attached to cell membrane by the glycosylphosphatidylinositol and can be combined with erythropoietin-producing hepatoma A2 (EphA2), EphA4, EphA5, EphA6 and EphA7 receptors. However, EFNB is a transmembrane protein and two transcriptional mutations encode different isomers.29
Physiological Functions and Mechanisms of TNFAIP4
TNFAIP4 is involved in cell migration, repulsion, adhesion, angiogenesis and dendritic spine morphogenesis. At present, Ephrin-eph signal theory is extremely complicated and the functional mechanism of TNFAIP4 is unclear.30 EFNA1, hypoxia and endothelial nitric oxide synthase (eNOS) were believed to be key roles in tumor angiogenesis several years ago. Hypoxia could increase EFNA1 in squamous cells, especially in the upper layer. Tumor angiogenesis induced by EFNA1 could increase the production of nitric oxide (NO) in human umbilical vein endothelial cells (HUVECs). The phosphorylation of protein kinase B (Akt) Ser473 and eNOS Ser1177 was increased in HUVECs by EFNA1 stimulation. LY294002 induced by EFNA1 was a specific inhibitor of phosphatidylinositol 3 (PI3K), and it could significantly reduce the expression of phosphorylated Akt Ser473 and eNOS Ser1177. Therefore, depending on activated eNOS and interaction between PI3K and Akt, EFNA1 could promote angiogenesis in the tumor microenvironment and may be a new mechanism of tumor angiogenesis.31
TNFAIP5
Basic Characteristics of TNFAIP5
TNFAIP5, also known as pentraxin 3 (PTX3) or tumor-specific glycoprotein 14 (TSG-14), is located in chromosome 3q25.32 and has 3 exons and a cyclic polymeric structure. TNFAIP5 expression is caused by inflammatory cytokines induced by inflammatory stimulation of mesenchymal cells and epithelial cells and can participate in acute inflammation and immune response.32
Physiological Functions and Mechanisms of TNFAIP5
PTX3 promotes the differentiation of fibroblasts, regulates inflammatory response and activates complement and participates in angiogenesis and tissue reconstruction.33 Currently, there are few studies on the mechanism of TNFAIP5 or PTX3, but the most common function of TNFAIP5 or PTX3 is to regulate inflammatory response. PTX3 is similar to C-reactive protein (CRP), which is caused by acute inflammation (eg, bacterial products, IL-1 and TNF). PTX3 is rapidly produced and released by macrophages, vascular endothelial cells, fibroblasts and dendritic cells, which are dominated by macrophages and vascular endothelial cells, and CRP is mainly produced by liver cells under the regulation of cytokine IL-6. Therefore, the level of PTX3 may be more directly responsive to the inflammatory state.32,34 Previous studies found that PTX3 participates in inflammatory, regulates the distribution of white blood cells and improves the permeability of blood vessels by binding to different ligands such as growth factors, extracellular matrix components, complement cascade molecules, pathogen recognition proteins, angiogenesis and adhesion molecules.35 In addition, it was reported that PTX3 can inhibit growth and invasion of tumor cells by blocking Akt and NF-κB signaling pathway, but the mechanism was still unclear.36
TNFAIP6
Basic Characteristics of TNFAIP6
TNFAIP6, also known as TSG-6, is located in chromosome 2q23.3 and has 6 exons, and it is composed of a CUB (complement subcomponents C1r/C1s, Uegf, BMP-1) domain and a hyaluronan-binding LINK domain.37 TNFAIP6 protein is a secreted protein which belongs to a member of the hyaluronic acid-binding protein family, and it may be related to the stability of extracellular matrix and cell migration.38,39
Physiological Functions and Mechanisms of TNFAIP6
TSG-6 is believed to be an inflammatory protein associated with accelerated wound healing and reduced tissue fibrosis, and it can regulate chemokines by inhibiting the interaction of chemokines and mucopolysaccharides.40,41 In rheumatoid arthritis and osteoarthritis, a study found that TSG-6 was high expression in patients’ joints, suggesting that TSG-6 might be a proinflammatory medium. Contrarily, TSG-6 could inhibit inflammatory models of arthritis and graft rejection, suggesting that TSG-6 also had anti-inflammatory property, and it was considered to be a major anti-inflammatory mediator of human mesenchymal stem cells in the models of myocardial infarction, corneal injury, peritonitis, traumatic brain injury, acute lung injury, trauma repair and diabetes.40 In vitro studies found that TSG-6 interacted with protein ligands and different mucopolysaccharides, and it could inhibit neutrophilic migration.39 In addition, TSG-6 could inhibit the transmigration of neutrophils stimulated by chemokines through direct interaction between TSG-6 and CXCL8 binding site. At the same time, TSG-6 might impair the binding of CXCL8 to cell surface mucopolysaccharide, as well as the transport of CXCL8 in endothelial cells.38 Another study found that TSG-6 interacted with chemokine CXCL11 and CCL5, suggesting that it might be similar to the encoded virus and had the function of specific chemokine binding proteins.40
TNFAIP8
Basic Characteristics of TNFAIP8
TNFAIP8 is an anti-apoptotic carcinogen induced by transcription factors and belongs to a member of TIPE family.42,43 In 1997, Patel et al discovered TNFAIP8 for the first time in human metastatic head and neck squamous cell carcinoma cells.44 Then, TNFAIP8 overexpression was found in various tumor cells such as breast cancer cells, non-small cell lung cancer (NSCLC) cells, gastric cancer cells and so on.45
Physiological Functions and Mechanisms of TNFAIP8
TNFAIP8 is similar to cytoplasmic scaffold protein and is mainly involved in tumor formation, immune response and inflammatory response. Studies confirmed that TNFAIP8 was a regulator of apoptosis and played an important role in proliferation, invasion, metastasis and apoptosis of tumor cells.42,43 In the basic experiment, TNFAIP8 induced by activated NF-κB transcription factors might regulate cellular function and NF-κB signaling pathway played a key role.46–48 Another study found that TNFAIP8 mRNA induced by TNF-α was irreversible under the action of IκBα which is a NF-κB inhibitor. Activation of NF-κB signaling pathway induced by TNF-α could increase TNFAIP8 expression, in contrast to inhibit cysteine protease 8 (caspase-8) and inhibit apoptosis, and it might lead to the formation and progression of tumors.49 In addition, TNFAIP8-like 2 (TIPE2) is also a member of TNFAIP8 family and could maintain immune homeostasis. In animal experiments, mice lacking TIPE2 were more likely to develop chronic inflammatory disease, and the abnormal expression of TIPE2 in humans was related to systemic autoimmune diseases, diabetic nephropathy and hepatitis B.47
TNFAIP9
Basic Characteristics of TNFAIP9
TNFAIP9 is located in chromosome 7q21.12 and belongs to prostate six-transmembrane epithelial antigen (STEAP) family. It is located in the golgi apparatus of cells and reduces Fe (3+) to Fe (2+) or reduces Cu (2+) to Cu (1+) with glutamate dehydrogenase (NAD+) receptor.
Physiological Functions and Mechanisms of TNFAIP9
Studies found that TNFAIP9 was involved in the development and metabolism of fat cells.50,51 In addition, it could affect the biological characteristics of normal prostate cells and lead to prostate cancer.47,52 At present, there were several findings about functional mechanism of TNFAIP9. Firstly, TNFAIP9 stimulates the production of fat by enhancing the expression of 1, 25-dihydroxy vitamin D3, CCAAT or enhancer-binding protein alpha (C/EBPα) and peroxisome proliferator-activated receptor gamma (PPAR-γ). Secondly, TNFAIP9 was involved in the transmission of nutrients, hormones and inflammatory signals in metabolic pathways. Fat-free diet, inflammatory signaling and fat factors regulated the expression of TNFAIP9 transmembrane protein. However, TNF-α, IL-6, IL-1β and leptin in fat cells regulated the metabolic activity of TNFAIP9, which is similar to insulin signaling pathway. Thirdly, TNFAIP9 increased the sensitivity of insulin by increasing tyrosine phosphorylation of insulin receptor and Akt kinase, translocating the glucose transporter GLUT4 to the cell membrane and uptaking glucose. Fourthly, Hepatitis B virus x (HBx) protein led to metabolic disorders, including insulin resistance and accumulation of abnormal lipid in liver. TNFAIP9 resisted HBx-related dysfunction of hepatocyte by reducing the stability of HBx protein.53–55
To sum up, genes and proteins of TNFAIPs have specific basic characteristics and physiological functions and may play an important role in the occurrence and development of various diseases such as malignant tumors, metabolic disorders and inflammation. Although mechanisms are still unclear, basic studies have been carried out. The basic characteristics of genes in TNFAIPs are summarized in Table 1, and the basic characteristics and physiological functions of TNFAIPs are pooled in Table 2.
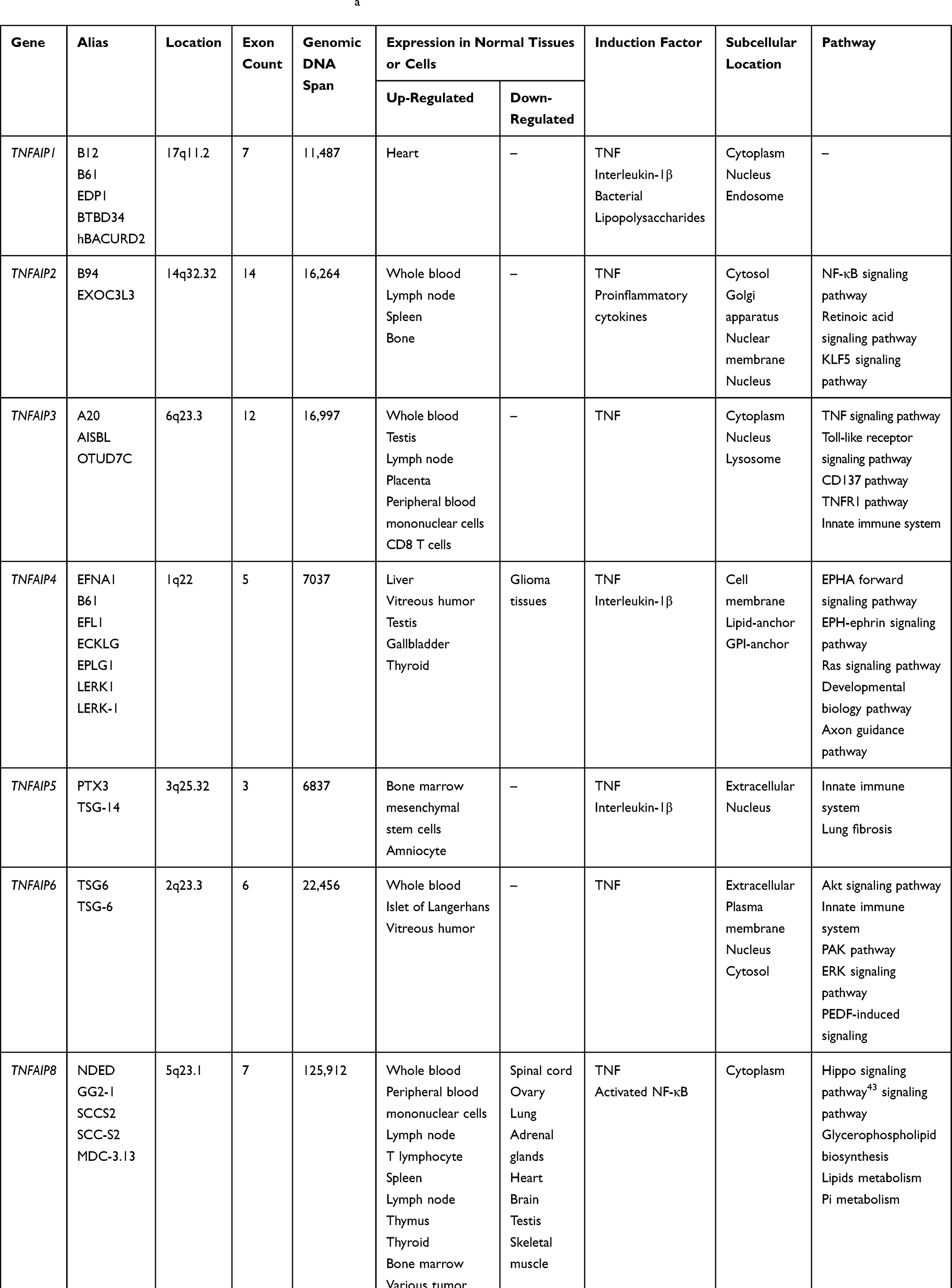 |
Table 1 Basic Characteristics of TNFAIPs Genesa |
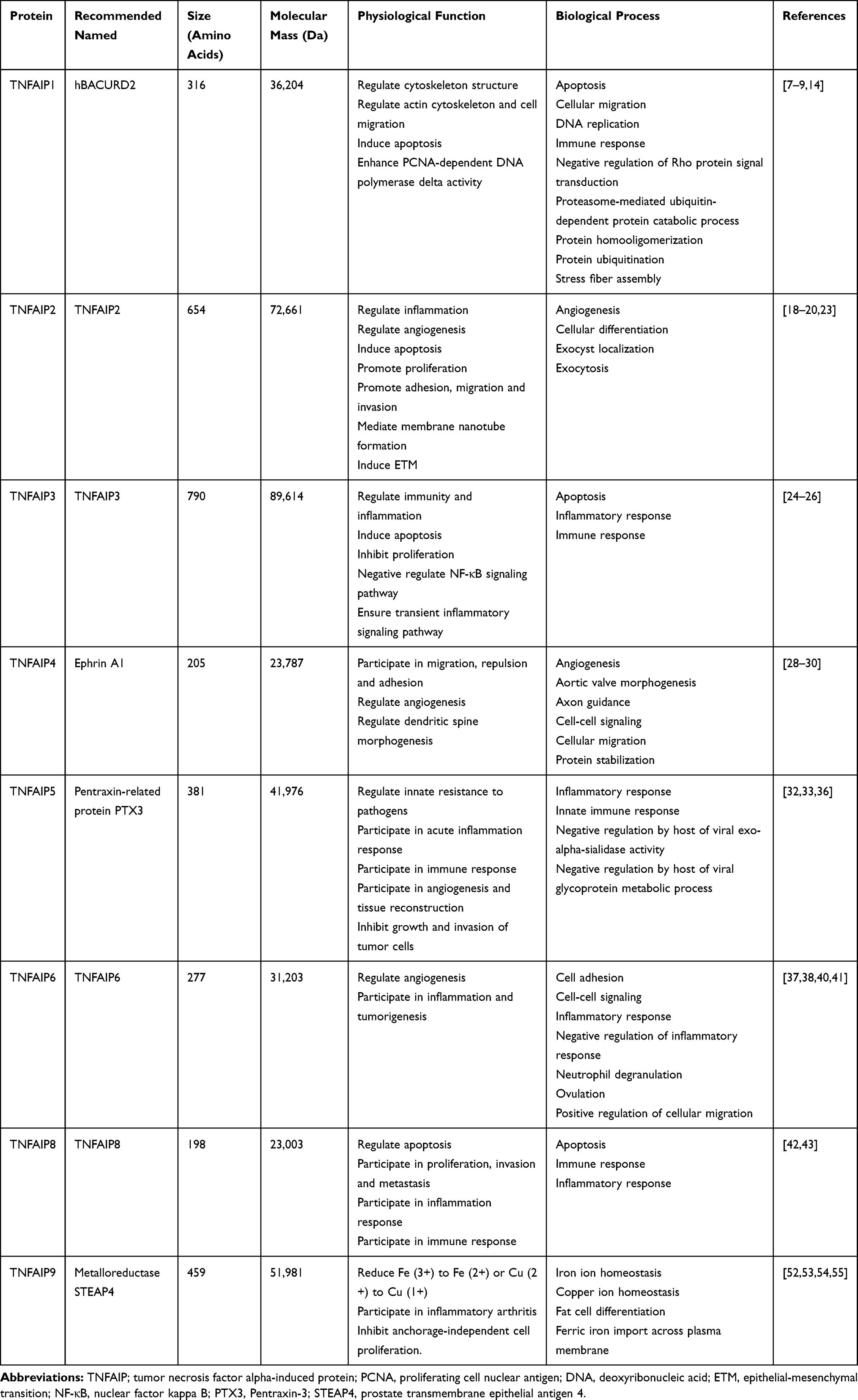 |
Table 2 Basic Characteristics and Physiological Functions of TNFAIPs |
TNFAIPs and Malignant Tumors
TNFAIP1 and Malignant Tumors
TNFAIP1 is caused by TNF-α and interleukin-6 (IL-6) in umbilical vein endothelial cells, which is high expression in normal cell lines and low expression in tumor cell lines.7,8 In 2013, two studies showed that miR-372 and miR-373 could negatively regulate TNFAIP1 expression by NF-κB signaling pathway to control proliferation and apoptosis in gastric cancer cells.10,11 A year later, Zhang et al found that TNFAIP1 was significantly overexpression in osteosarcoma. TNFAIP1 knockout could inhibit proliferation and induce apoptosis in osteosarcoma cells, and it up-regulated Caspase-3 and down-regulated NF-κB p65, PCNA and Matrix metalloproteinase-2 (MMP-2). This study confirmed that TNFAIP1 overexpression was also associated with distant metastasis and suggested that TNFAIP1 might be a potential therapeutic target for osteosarcoma.56 Similar to the above, Tian et al had verified that TNFAIP1 was a novel gene of miR-15a and could inhibit proliferation and invasion in osteosarcoma cells. Low expression of TNFAIP1 could regulate the level of miR-15a and might be a new therapeutic strategy for osteosarcoma.12 An American study showed that TNFAIP1 expression was negatively correlated with miR-224 in NSCLC. Both TNFAIP1 knockout and miR-224 overexpression could induce the same subtype and indicated that TNFAIP1 played a certain role in proliferation and invasion through miR-224 regulation.13 Another study discovered the interaction relationship between miR-181a and TNFAIP1 in the Chinese population. TNFAIP1 as a downstream target gene of miR-181a could regulate proliferation and metastasis in pancreatic cancer cells.14 In addition, some studies showed that TNFAIP1 was not only related to malignant tumors, but also played a role for inflammatory-related angiogenesis, natural immunity after HPV infection and paclitaxel resistance.57,58
TNFAIP2 and Malignant Tumors
TNFAIP2 expression was associated with malignant tumors and septic shock, especially for prediction and prognosis in NPC, esophageal squamous cell carcinoma (ESCC), gastric cancer, head and neck squamous cell carcinoma (SCCHN), lymphoma and breast cancer. NPC is a metastable epithelial carcinoma, which is closely related to Epstein–Barr virus (EBV) infection. Chen et al found that TNFAIP2 as cell transfer promoter protein was a correlation with invasion and metastasis, and it was overexpressed in NPC samples than adjacent normal tissues. TNFAIP2 overexpression might shorten the distant disease-free survival of NPC patients, so it might be an independent prognostic factor for NPC.59 In a cohort study of ESCC patients, Zhang et al discovered that functional TNFAIP2 rs8126 genetic polymorphism obviously increased the risk of ESCC in Chinese patients, at the same time to verify the functional genetic mutation in 3ʹuntranslated region (3ʹUTR) region may interfere with the miRNA mediated gene regulation.60 In gastric cancer, study showed that compared with TT+TC genotype, CC genotype of TNFAIP2 3ʹUTR rs8126 significantly increased the risk of gastric cancer, especially for drinking patients, and 3ʹUTR miRNA binding site of TNFAIP2 was confirmed as a sensitive predictive marker of gastric cancer.15 A study on SCCHN confirmed that TNFAIP2 3ʹUTR miR-184 binding site functional single nucleotide polymorphism (SNP) (rs8126 T>C) could regulate TNFAIP2 expression and was associated with the risk of SCCHN.61 In recent years, a study reported the relationship between TNFAIP2 and breast cancer. They found that both KLF5 and TNFAIP2 could promote proliferation, invasion and metastasis in breast cancer cells through Rac1 and Cdc42.16 In addition to the above tumors, TNFAIP2 is also associated with renal cancer, thymoma, sarcoma and so on, but these are all based on small sample basic studies, and large sample clinical studies are needed to verify the predictive and prognostic value of TNFAIP2.
TNFAIP3 and Malignant Tumors
TNFAIP3 gene polymorphism is closely related to autoimmune and inflammatory diseases, including rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), systemic sclerosis, psoriasis, allergic rhinitis and diabetes. Otherwise, inactivated TNFAIP3 may induce malignant tumors, especially lymphoma. A study confirmed that inactivated TNFAIP3 could promote the activation of uncontrolled NF-κB signaling pathway, which improved cell viability and led to lymphoma.25 Another study found that TNFAIP3, as a tumor suppressor gene, may be a key molecule in the transformation of chronic inflammation into cancer.62 In multiple myeloma cells, TNFAIP3 was found to be inactivated mainly because of deletions, not body mutations or promoter methylation. Inactivated TNFAIP3 could affect the activity of NF-κB signaling pathway and led to multiple myeloma, and it was suggested that TNFAIP3 might be a potential therapeutic target for multiple myeloma.27
TNFAIP4 and Malignant Tumors
EFNA1 was used as an alternative to TNFAIP4 in previous literature since 1990. In recent years, some studies have shown that EFNA1 and EphA2 expression was closely related to multiple tumors. In 2005, a study found that EFNA1 and EphA2 expression might affect the biological behavior of gastric cancer.63 Subsequently, Yuan et al confirmed that EphA2 and EFNA1 overexpression played an important role in gastric adenocarcinoma, and EphA2 is an independent factor for poor prognosis in postoperative patients.64 Further study found that EFNA1 3ʹUTR miR-200c binding site SNP (rs12904 G > A) regulated EFNA1 expression and was associated with susceptibility of gastric cancer.65 Another study found that compared with adjacent normal tissues, EFNA1 was overexpressed in colorectal cancer, and lack of EFNA1 reduced the risk of invasion and metastasis in colorectal cancer (CRC) cells. The results suggested that EFNA1 was involved in tumor angiogenesis and may be an independent prognostic factor for colorectal cancer.66,67 So far, no studies had reported on the relationship between TNFAIP4 and other diseases.
TNFAIP5 and Malignant Tumors
Previous studies showed that TNFAIP5 or PTX3 expression had important clinical value in tumor diagnosis and prognosis assessment. In 2013, Zhang et al found that PTX3 was significantly elevated in tissues and serum samples of patients with lung cancer, and had certain significance for diagnosis of lung cancer, early lung cancer and non-malignant lung disease.68 According to pathological classification, it was further confirmed that the up-regulation of PTX3 expression was associated with small cell lung cancer and lung adenocarcinoma.33,36 Recently, Infante et al found that serum PTX3 level in NSCLC was significantly higher than that in non-cancer smokers. Although preoperative serum PTX3 level was not correlated with the prognosis of patients, PTX3 overexpression in the interstitial mass of the removed tissue sample could shorten survival and might be an independent prognostic factor.69 In addition, Zhang et al found that increased PTX3 was related to the poor prognosis of patients with CRC, and confirmed that PTX3 was an independent prognostic factor for CRC.70 Carmo et al found that PTX3 polymorphism was associated with the incidence of hepatitis C. The polymorphism of PTX3 rs2305619 and plasma level were associated with liver cancer patients with Child grade B and C, while PTX3 seemed to be a risk factor for patients with liver cancer and chronic hepatitis C.71 In summary, TNFAIP5 or PTX3 overexpression in tumors might indicate the risk of disease recurrence and metastasis, and it could be a prediction and prognostic factor for patients with malignant tumors.
TNFAIP6 and Malignant Tumors
At present, the study on TNFAIP6 is still in the early stage. Only a few data from basic studies have been reported, and results on the relationship between TNFAIP6 and malignant tumors have not been seen.
TNFAIP8 and Malignant Tumors
In recent years, many studies confirmed that TNFAIP8 was associated with the development and prognosis of malignant tumors. As early as 2010, Dong et al found that TNFAIP8 overexpression was associated with lymph node metastasis and poor prognosis in patients with NSCLC.72 A new study confirmed that TNFAIP8 could promote proliferation and cisplatin chemoresistance of NSCLC by MDM2/p53 pathway.73 In a study on pancreatic cancer, compared with normal tissue, TNFAIP8 was strongly stained in tumor, and positive expression of TNFAIP8 was correlated with EGFR level.74 Another study on prostate cancer showed that TNFAIP8 expression was significantly associated with proliferation, invasion and metastasis and confirmed that TNFAIP8 might be an androgen-induced molecule and had the dual role of prediction and prognostic for the first time.48 In addition, TNFAIP8 gene polymorphism is closely related to the development of gynecological tumors. A study showed that TNFAIP8 rs11064 SNP might affect the affinity between miR-22 and 3ʹUTR domain, and it could cause cervical cancer by regulating the expression of TNFAIP8.75 Another study found that SNP rs11064 in TNFAIP8 increased endometrial cancer risk and significantly related with its protein expression in northern Chinese women.76 Recently, many studies on Chinese population suggested that TNFAIP8 expression was related to other tumors such as thyroid papillary carcinoma, epithelial ovarian cancer, gastric cancer and esophageal squamous cell carcinoma.42,45,47,77–80 A study showed that TNFAIP8 overexpression was not obviously related with the prognosis of gastric cancer patients, but it was an independent prognostic factor for disease-free survival (DFS) and overall survival (OS) in patients with intestinal type of gastric cancer.49 Recent studies related to TNFAIP8 and gastric cancer had focused on miRNAs, and it was well known that abnormal regulation of miRNAs could affect proliferation, apoptosis, invasion and metastasis in the development of tumors. TNFAIP8 was a negative target of miR-9 which was significantly down-regulated expression in gastric cancer. The results confirmed that miR-9-TNFAIP8 might be a promising predictive and prognostic biomarker for gastric cancer.45 A review on TNFAIP8 showed that overexpression of TNFAIP8 significantly improved vascular endothelial growth factor receptor 2 (VEGFR2), MMP-1, and MMP-9 levels in breast cancer cells, thereby promoting cancer cells to metastasize. It is suggested that high-risk breast cancer patients with TNFAIP8 overexpression possessed shorter OS and DFS.81 The latest research found that TNFAIP8 promoted VEGFR2-mediated angiogenesis by upregulating 3-phosphoinositide dependent kinase-1 (PDK1) expression and phosphorylation in colorectal cancer, and TNFAIP8 might be a target for antiangiogenic cancer therapy.82
TNFAIP9 and Malignant Tumors
Nowadays, studies found that TNFAIP9 was an androgen regulating gene and highly expressed in prostate cancer. The abnormal expression of TNFAIP9 led to the proliferation of prostate cancer cells. Further study found that TNFAIP9 obviously increased reactive oxygen species (ROS) and reduced the level of nicotinamide adenine dinucleotide phosphate (NADPH) in prostate cancer cells through the activity of iron reductase. Therefore, TNFAIP9 knockout could inhibit cellular proliferation, clone formation and adherent growth and significantly promoted cellular apoptosis.52 However, due to the limitations of experimental conditions and samples, some studies were needed to confirm the above results.
To sum up, protein expression and gene polymorphism have become the hot topics in recent years. Some studies have found that expression of TNFAIPs may be directly or indirectly associated with proliferation, invasion, metastasis, tumorigenesis and cisplatin chemoresistance of malignant tumors. The relationship between TNFAIPs and malignant tumors are summarized in Table 3 and the biological roles of TNFAIPs in malignant tumors are drawn in Figure 1.
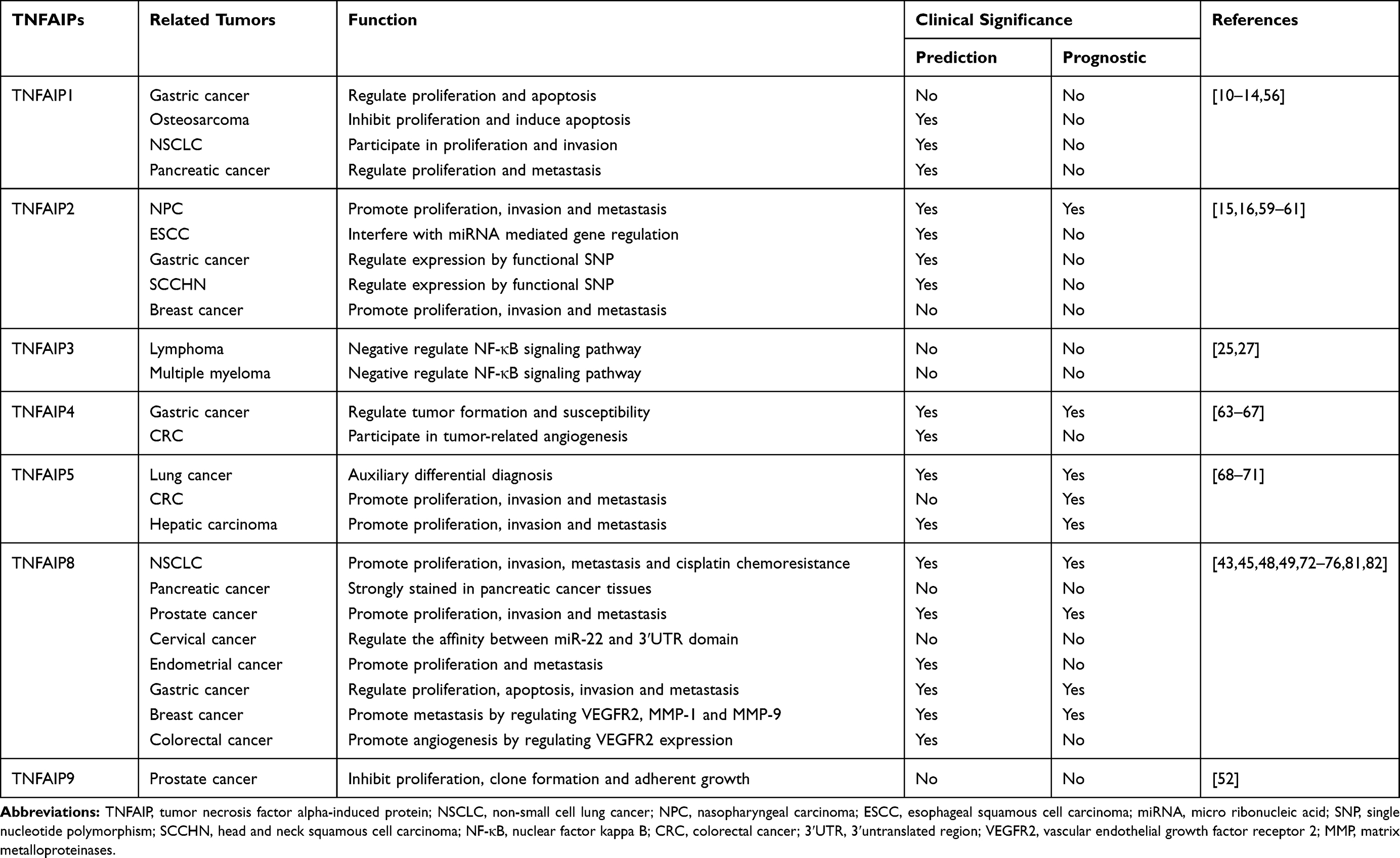 |
Table 3 Relationship Between TNFAIPs and Malignant Tumors |
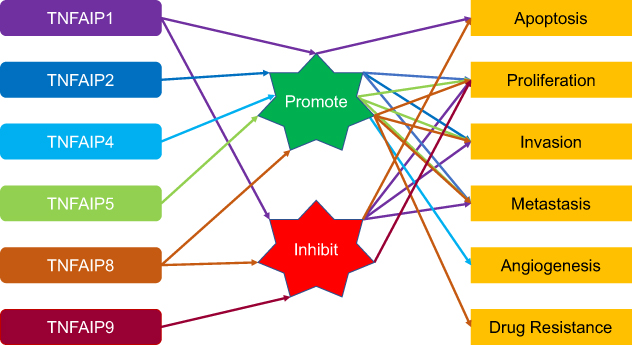 |
Figure 1 The biological roles of TNFAIPs in malignant tumors. TNFAIP1 promotes apoptosis and inhibits cellular proliferation, invasion and metastasis. TNFAIP2 promotes cellular proliferation, invasion and metastasis. TNFAIP4 promotes angiogenesis. TNFAIP5 promotes cellular proliferation, invasion and metastasis. TNFAIP8 inhibits apoptosis and promotes cellular proliferation, invasion, metastasis and drug resistance. TNFAIP9 inhibits cellular proliferation. The biological properties of TNFAIP3 and TNFAIP6 in malignant tumors have not been clarified. Abbreviation: TNFAIP, tumor necrosis factor alpha-induced protein. |
The Inner Relationship Among Members of TNFAIPs
We explore the inner relationship between members of TNFAIPs by bioinformatics databases on the internet. Firstly, we search for proteins that may be associated with each member of TNFAIPs through STRING database (Table 4). Secondly, we find that TNF is closely related to several members of TNFAIPs, and the protein–protein interaction diagram between TNFAIPs' members is drawn in Figure 2. We found that the positive expression of TNF directly activates TNFAIP1, TNFAIP4, TNFAIP5 and TNFAIP8. The unspecified expression of TNF affects the transcription of TNFAIP5. There is a correlation between TNF and TNFAIP6, but the exact relationship is unknown. The negative expression of TNFAIP3 inhibits TNF, and the unspecified expression of TNFAIP3 affects the catalysis of TNF. Binding structures can be formed between TNF and TNFAIP3 and between TNFAIP5 and TNFAIP6. There is a correlation between TNFAIP2 and TNFAIP3, but the exact relationship is unknown. TNFAIP4 has not yet found an internal link with other members of TNFAIPs. At present, no studies have shown other actual existing interactions between TNFAIPs' members.
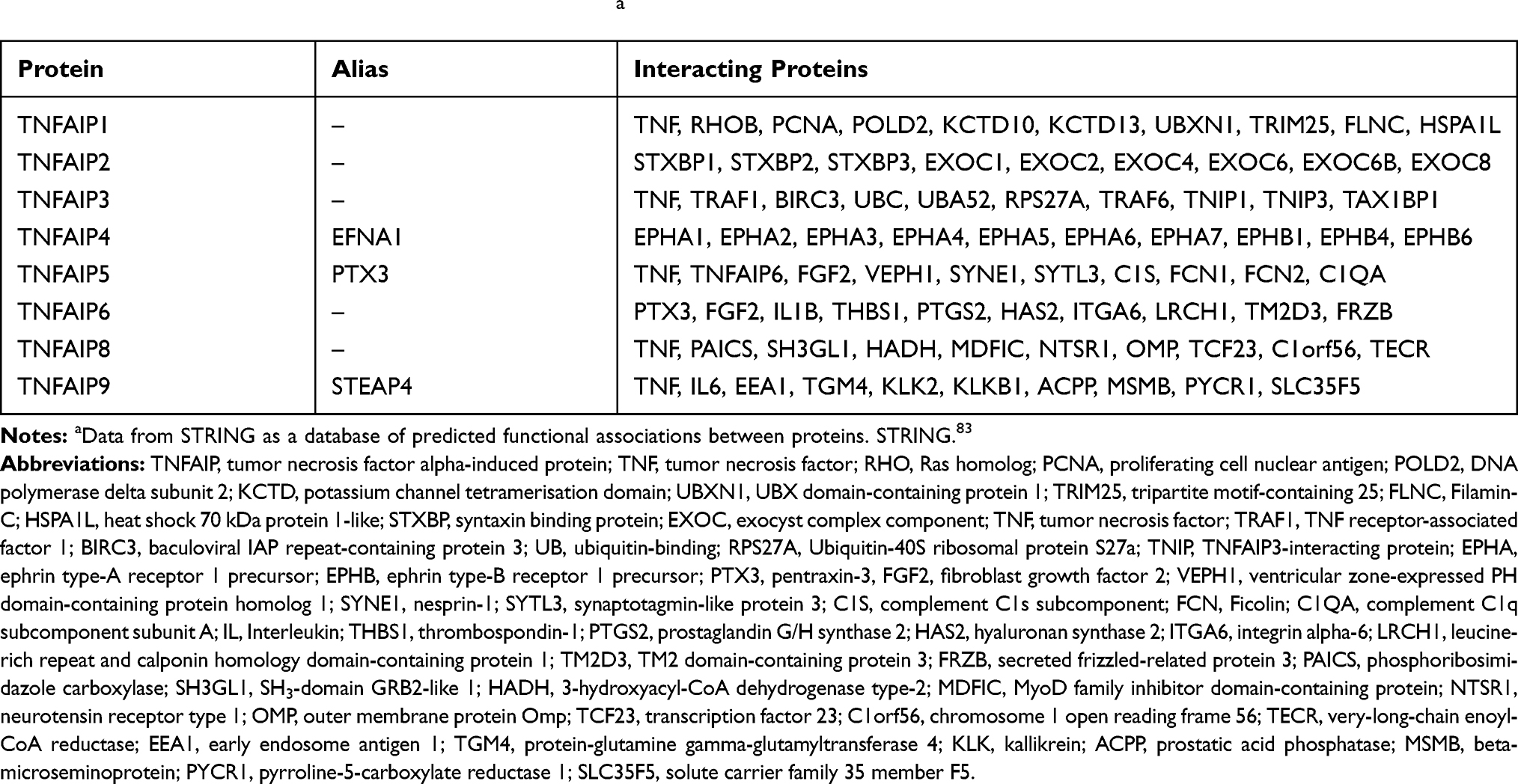 |
Table 4 Interacting Proteins in Members of TNFAIPsa |
 |
Figure 2 The protein–protein interaction diagram between TNFAIPs' members. The positive expression of TNF directly activates TNFAIP1, TNFAIP4, TNFAIP5 and TNFAIP8. The unspecified expression of TNF affects the transcription of TNFAIP5. There is a correlation between TNF and TNFAIP6, but the exact relationship is unknown. The negative expression of TNFAIP3 inhibits TNF and the unspecified expression of TNFAIP3 affects the catalysis of TNF. Binding structures can be formed between TNF and TNFAIP3 and between TNFAIP5 and TNFAIP6. There is a correlation between TNFAIP2 and TNFAIP3, but the exact relationship is unknown. TNFAIP4 has not yet found an internal link with other members of TNFAIPs. Abbreviations: TNF, tumor necrosis factor; TNFAIP, tumor necrosis factor alpha-induced protein. |
Summary and Prospects
This paper in the form of “Gene–Protein–Tumor correlation” systematically reviews the biological characteristics and physiological functions of the hitherto known eight members of TNFAIPs, and explores the relationship between TNFAIPs and malignant tumors. TNFAIPs is a protein family induced by TNF and protein expression of each member in the family almost all are associated with TNF. TNF is the first cytokine used in tumor biotherapy. However, due to its lack of highly functional and specific targets, it is currently only used for local treatment. If we can accurately screen the active targets of TNF, targeted drugs will be of great significance for clinical treatment, and studies on TNFAIPs may be a breakthrough.
As we all know, TNFAIPs participate in proliferation, invasion, metastasis and apoptosis of tumor cells and are closely related to the occurrence and development of various malignant tumors. For example, TNFAIP1 and TNFAIP8 are the most frequently reported proteins in TNFAIPs, and they play an important role in NSCLC and pancreatic cancer. TNFAIP1 regulates the proliferation and metastasis of NSCLC and pancreatic cancer by miRNA, but overexpression of TNFAIP8 is associated with lymph node metastasis and poor prognosis. In addition, TNFAIP1, TNFAIP2, TNFAIP4 and TNFAIP8 are all related to gastric cancer and negative regulation of TNFAIP1 control the proliferation and metastasis of gastric cancer cells. TNFAIP2 genetic polymorphism may interfere with miRNA mediated gene regulation, and overexpression of TNFAIP4 or TNFAIP8 may play an important role in prediction and prognostic of gastric cancer patients. Recently, both TNFAIP8 and TNFAIP9 have been reported in the studies on prostate cancer, and the former may be an androgen-induced molecule and is significantly associated with proliferation, invasion and metastasis, and knockout of the latter inhibit proliferation, clone formation and adherent growth, and can significantly promote apoptosis in prostate cancer cells. At present, the mechanism of TNFAIP1, TNFAIP2 and TNFAIP3 inducing tumors may be related to the NF-κB signaling pathway, but the mechanism of tumor induction by other members of TNFAIPs is not clear, and exploratory researches are urgently needed.
Nowadays, the understanding of TNFAIPs is very shallow. The studies on TNFAIPs are still at an early stage, mainly based on experimental studies. Some studies only focus on the characteristics, basic functions and protein expression of individual members, and there is lack of exploratory studies which reveal the relationship between genes and genes or between proteins and proteins. A few studies have reported that members of TNFAIPs are involved in the occurrence and development of tumors, but due to the limitations of sample size and detection methods, it is difficult to reveal the complex regulatory mechanism. In addition, some studies have explored the correlation between TNFAIPs and cancer risk, but few studies have mentioned relationship between TNFAIPs and prognosis in cancer patients due to the lack of clinical follow-up data.
In the future, we need to carry out in-depth research on relevant mechanisms of TNFAIPs by bioinformatics, molecular biology, genomics and other techniques and methods. The next step is to clarify the functional features and transduction pathways of TNFAIPs, which will play an important theoretical foundation for the translational research of “basic to clinical”.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
This study was partly supported by grants from the National Key R&D Program of China (Grant #2018YFC1311600).
Disclosure
The authors declare that there is no conflict of interest in this work.
References
1. Carswell EA, Old LJ, Kassel RL, et al. An endotoxin-induced serum factor that causes necrosis of tumors. Proc Natl Acad Sci U S A. 1975;72(9):3666–3670. doi:10.1073/pnas.72.9.3666
2. Wakefield PE, James WD, Samlaska CP, Meltzer MS. Tumor necrosis factor. J Am Acad Dermatol. 1991;24(5,pt 1):675–685. doi:10.1016/0190-9622(91)70102-8
3. Shalaby MR, Aggarwal BB, Rinderknecht E, Svedersky LP, Finkle BS, Palladino MA
4. Olmos G, Lladó J. Tumor necrosis factor alpha: a link between neuroinflammation and excitotoxicity. Mediators Inflamm. 2014;2014:861231. doi:10.1155/2014/861231
5. Balkwill F. Tumour necrosis factor and cancer. Nat Rev Cancer. 2009;9(5):361–371. doi:10.1038/nrc2628
6. Giat E, Ehrenfeld M, Shoenfeld Y. Cancer and autoimmune diseases. Autoimmun Rev. 2017;16(10):1049–1057. doi:10.1016/j.autrev.2017.07.022
7. Hu X, Yan F, Wang F, et al. TNFAIP1 interacts with KCTD10 to promote the degradation of KCTD10 proteins and inhibit the transcriptional activities of NF-κB and AP-1. Mol Biol Rep. 2012;39(11):9911–9919. doi:10.1007/s11033-012-1858-7
8. Liu N, Wei K, Xun Y, et al. Transcription factor cyclic adenosine monophosphate responsive element binding protein negatively regulates tumor necrosis factor alpha-induced protein 1 expression. Mol Med Rep. 2015;12(5):7763–7769. doi:10.3892/mmr.2015.4336
9. Yang L, Liu N, Hu X, et al. CK2 phosphorylates TNFAIP1 to affect its subcellular localization and interaction with PCNA. Mol Biol Rep. 2010;37(6):2967–2973. doi:10.1007/s11033-009-9863-1
10. Zhou C, Li X, Zhang X, et al. microRNA-372 maintains oncogene characteristics by targeting TNFAIP1 and affects NFκB signaling in human gastric carcinoma cells. Int J Oncol. 2013;42(2):635–642. doi:10.3892/ijo.2012.1737
11. Zhang X, Li X, Tan Z, et al. MicroRNA-373 is upregulated and targets TNFAIP1 in human gastric cancer, contributing to tumorigenesis. Oncol Lett. 2013;6(5):1427–1434. doi:10.3892/ol.2013.1534
12. Tian X, Zhang J, Yan L, Dong JM, Guo Q. MiRNA-15a inhibits proliferation, migration and invasion by targeting TNFAIP1 in human osteosarcoma cells. Int J Clin Exp Pathol. 2015;8(6):6442–6449.
13. Cui R, Meng W, Sun HL, et al. MicroRNA-224 promotes tumor progression in nonsmall cell lung cancer. Proc Natl Acad Sci U S A. 2015;112(31):E4288–E4297. doi:10.1073/pnas.1502068112
14. Zhang P, Guo Z, Hu R, He X, Jiao X, Zhu X. Interaction between microRNA-181a and TNFAIP1 regulates pancreatic cancer proliferation and migration. Tumour Biol. 2015;36(12):9693–9701. doi:10.1007/s13277-015-3704-8
15. Xu Y, Ma H, Yu H, et al. The miR-184 binding-site rs8126 T>C polymorphism in TNFAIP2 is associated with risk of gastric cancer. PLoS One. 2013;8(5):e64973. doi:10.1371/journal.pone.0064973
16. Jia L, Zhou Z, Liang H, et al. KLF5 promotes breast cancer proliferation, migration and invasion in part by upregulating the transcription of TNFAIP2. Oncogene. 2016;35(16):2040–2051. doi:10.1038/onc.2015.263
17. Thair SA, Topchiy E, Boyd JH, et al. TNFAIP2 inhibits early TNFα-Induced NF-x03BA;B signaling and decreases survival in septic shock patients. J Innate Immun. 2016;8(1):57–66. doi:10.1159/000437330
18. Rusiniak ME, Yu M, Ross DT, Tolhurst EC, Slack JL. Identification of B94 (TNFAIP2) as a potential retinoic acid target gene in acute promyelocytic leukemia. Cancer Res. 2000;60(7):1824–1829.
19. Jia L, Shi Y, Wen Y, Li W, Feng J, Chen C. The roles of TNFAIP2 in cancers and infectious diseases. J Cell Mol Med. 2018;22(11):5188–5195. doi:10.1111/jcmm.13822
20. Chen CC, Liu HP, Chao M, et al. NF-κB-mediated transcriptional upregulation of TNFAIP2 by the Epstein-Barr virus oncoprotein, LMP1, promotes cell motility in nasopharyngeal carcinoma. Oncogene. 2014;33(28):3648–3659. doi:10.1038/onc.2013.345
21. Wood MB, Rios D, Williams IR. TNF-α augments RANKL-dependent intestinal M cell differentiation in enteroid cultures. Am J Physiol Cell Physiol. 2016;311(3):C498–C507. doi:10.1152/ajpcell.00108.2016
22. Scholz A, Rappl P, Böffinger N, Mota AC, Brüne B, Schmid T. Translation of TNFAIP2 is tightly controlled by upstream open reading frames. Cell Mol Life Sci. 2019. doi:10.1007/s00018-019-03265-4
23. Niwa N, Tanaka N, Hongo H, et al. TNFAIP2 expression induces epithelial-to-mesenchymal transition and confers platinum resistance in urothelial cancer cells. Lab Invest. 2019;99(11):1702–1713. doi:10.1038/s41374-019-0285-y
24. Han JW, Wang Y, Li HB, et al. Single nucleotide polymorphisms of TNFAIP3 are associated with systemic lupus erythematosus in Han Chinese population. Int J Immunogenet. 2016;43(2):96–100. doi:10.1111/iji.12250
25. Wenzl K, Hofer S, Troppan K, et al. Higher incidence of the SNP Met 788 Ile in the coding region of A20 in diffuse large B cell lymphomas. Tumour Biol. 2016;37(4):4785–4789. doi:10.1007/s13277-015-4322-1
26. Zhu L, Zhou L, Wang L, et al. A20 SNP rs77191406 may be related to secondary cancer for rheumatoid arthritis and systemic lupus erythematosus patients. Asia Pac J Clin Oncol. 2016;12(4):409–414. doi:10.1111/ajco.12577
27. Troppan K, Hofer S, Wenzl K, et al. Frequent down regulation of the tumor suppressor gene a20 in multiple myeloma. PLoS One. 2015;10(4):e0123922. doi:10.1371/journal.pone.0123922
28. Makarov A, Ylivinkka I, Nyman TA, Hyytiäinen M, Keski-Oja J. Ephrin-As, Eph receptors and integrin α3 interact and colocalise at membrane protrusions of U251MG glioblastoma cells. Cell Biol Int. 2013;37(10):1080–1088. doi:10.1002/cbin.v37.10
29. Beauchamp A, Debinski W. Ephs and ephrins in cancer: ephrin-A1 signalling. Semin Cell Dev Biol. 2012;23(1):109–115. doi:10.1016/j.semcdb.2011.10.019
30. Ferluga S, Hantgan R, Goldgur Y, Himanen JP, Nikolov DB, Debinski W. Biological and structural characterization of glycosylation on ephrin-A1, a preferred ligand for EphA2 receptor tyrosine kinase. J Biol Chem. 2013;288(25):18448–18457. doi:10.1074/jbc.M113.464008
31. Song Y, Zhao XP, Song K, Shang ZJ. Ephrin-A1 is up-regulated by hypoxia in cancer cells and promotes angiogenesis of HUVECs through a coordinated cross-talk with eNOS. PLoS One. 2013;8(9):e74464. doi:10.1371/journal.pone.0074464
32. Zhang X, Ding W. Association of genetic variants in pentraxin 3 gene with ankylosing spondylitis. Med Sci Monit. 2016;22:2911–2916. doi:10.12659/MSM.896562
33. Liu C, Yao Y, Wang W. Pentraxin-3 as a prognostic marker in patients with small-cell lung cancer. Med Oncol. 2014;31(10):207. doi:10.1007/s12032-014-0207-1
34. Park KS, Kim JW, An JH, Woo JM. Elevated plasma pentraxin 3 and its association with retinal vein occlusion. Korean J Ophthalmol. 2014;28(6):460–465. doi:10.3341/kjo.2014.28.6.460
35. Fornai F, Carrizzo A, Ferrucci M, et al. Brain diseases and tumorigenesis: the good and bad cops of pentraxin3. Int J Biochem Cell Biol. 2015;69:70–74. doi:10.1016/j.biocel.2015.10.017
36. Hu FQ, Qiao T, Xie X, Hu R, Xiao HB. Knockdown of the inflammatory factor pentraxin-3 suppresses growth and invasion of lung adenocarcinoma through the AKT and NF-kappa B pathways. J Biol Regul Homeost Agents. 2014;28(4):649–657.
37. Maruotti N, Annese T, Cantatore FP, Ribatti D. Macrophages and angiogenesis in rheumatic diseases. Vasc Cell. 2013;5(1):11. doi:10.1186/2045-824X-5-11
38. Dyer DP, Thomson JM, Hermant A, et al. TSG-6 inhibits neutrophil migration via direct interaction with the chemokine CXCL8. J Immunol. 2014;192(5):2177–2185. doi:10.4049/jimmunol.1300194
39. Capp E, Milner CM, Williams J, et al. Modulation of tumor necrosis factor-stimulated gene-6 (TSG-6) expression in human endometrium. Arch Gynecol Obstet. 2014;289(4):893–901. doi:10.1007/s00404-013-3080-9
40. Dyer DP, Salanga CL, Johns SC, et al. The anti-inflammatory protein TSG-6 regulates chemokine function by inhibiting chemokine/glycosaminoglycan interactions. J Biol Chem. 2016;291(24):12627–12640. doi:10.1074/jbc.M116.720953
41. Qi Y, Jiang D, Sindrilaru A, et al. TSG-6 released from intradermally injected mesenchymal stem cells accelerates wound healing and reduces tissue fibrosis in murine full-thickness skin wounds. J Invest Dermatol. 2014;134(2):526–537. doi:10.1038/jid.2013.328
42. Yang M, Zhao Q, Wang X, et al. TNFAIP8 overexpression is associated with lymph node metastasis and poor prognosis in intestinal-type gastric adenocarcinoma. Histopathology. 2014;65(4):517–526. doi:10.1111/his.2014.65.issue-4
43. Niture S, Dong X, Arthur E, et al. Oncogenic role of tumor necrosis factor α-induced protein 8 (TNFAIP8). Cells. 2018;8(1). doi:10.3390/cells8010009.
44. Patel S, Wang FH, Whiteside TL, Kasid U. Identification of seven differentially displayed transcripts in human primary and matched metastatic head and neck squamous cell carcinoma cell lines: implications in metastasis and/or radiation response. Oral Oncol. 1997;33(3):197–203. doi:10.1016/S0964-1955(96)00065-6
45. Gao HY, Huo FC, Wang HY, Pei DS. MicroRNA-9 inhibits the gastric cancer cell proliferation by targeting TNFAIP8. Cell Prolif. 2017;50(2):e12331. doi:10.1111/cpr.12331
46. Kumar D, Gokhale P, Broustas C, Chakravarty D, Ahmad I, Kasid U. Expression of SCC-S2, an antiapoptotic molecule, correlates with enhanced proliferation and tumorigenicity of MDA-MB 435 cells. Oncogene. 2004;23(2):612–616. doi:10.1038/sj.onc.1207123
47. Zhang Y, Wang MY, He J, et al. Tumor necrosis factor-α induced protein 8 polymorphism and risk of non-Hodgkin’s lymphoma in a Chinese population: a case-control study. PLoS One. 2012;7(5):e37846. doi:10.1371/journal.pone.0037846
48. Zhang C, Kallakury BV, Ross JS, et al. The significance of TNFAIP8 in prostate cancer response to radiation and docetaxel and disease recurrence. Int J Cancer. 2013;133(1):31–42. doi:10.1002/ijc.v133.1
49. Li Y, Jing C, Chen Y, et al. Expression of tumor necrosis factor α-induced protein 8 is upregulated in human gastric cancer and regulates cell proliferation, invasion and migration. Mol Med Rep. 2015;12(2):2636–2642. doi:10.3892/mmr.2015.3690
50. Xu HM, Cui YZ, Wang WG, et al. Expression and clinical significance of obesity-associated gene STEAP4 in obese children. Genet Mol Res. 2016;15(4). doi:10.4238/gmr.15048705
51. Catalán V, Gómez-Ambrosi J, Rodríguez A, et al. Six-transmembrane epithelial antigen of prostate 4 and neutrophil gelatinase-associated lipocalin expression in visceral adipose tissue is related to iron status and inflammation in human obesity. Eur J Nutr. 2013;52(6):1587–1595. doi:10.1007/s00394-012-0464-8
52. Jin Y, Wang L, Qu S, et al. STAMP2 increases oxidative stress and is critical for prostate cancer. EMBO Mol Med. 2015;7(3):315–331. doi:10.15252/emmm.201404181
53. Gui LY, Hu YL, Hou ZL, Wang P. Expression and clinical significance of the obesity-related gene TNFAIP9 in obese children. Genet Mol Res. 2016;15(3). doi:10.4238/gmr.15037995
54. Qi Y, Yu Y, Wu Y, et al. Genetic variants in six-transmembrane epithelial antigen of prostate 4 increase risk of developing metabolic syndrome in a Han Chinese population. Genet Test Mol Biomarkers. 2015;19(12):666–672. doi:10.1089/gtmb.2015.0104
55. Yoo SK, Cheong J, Kim HY. STAMPing into mitochondria. Int J Biol Sci. 2014;10(3):321–326. doi:10.7150/ijbs.8456
56. Zhang CL, Wang C, Yan WJ, Gao R, Li YH, Zhou XH. Knockdown of TNFAIP1 inhibits growth and induces apoptosis in osteosarcoma cells through inhibition of the nuclear factor-κB pathway. Oncol Rep. 2014;32(3):1149–1155. doi:10.3892/or.2014.3291
57. Zhu Y, Yao Z, Wu Z, Mei Y, Wu M. Role of tumor necrosis factor alpha-induced protein 1 in paclitaxel resistance. Oncogene. 2014;33(25):3246–3255. doi:10.1038/onc.2013.299
58. Liu M, Sun Z, Zhou A, et al. Functional characterization of the promoter region of human TNFAIP1 gene. Mol Biol Rep. 2010;37(4):1699–1705. doi:10.1007/s11033-009-9588-1
59. Chen LC, Chen CC, Liang Y, Tsang NM, Chang YS, Hsueh C. A novel role for TNFAIP2: its correlation with invasion and metastasis in nasopharyngeal carcinoma. Mod Pathol. 2011;24(2):175–184. doi:10.1038/modpathol.2010.193
60. Zhang J, Yu H, Zhang Y, et al. A functional TNFAIP2 3ʹ-UTR rs8126 genetic polymorphism contributes to risk of esophageal squamous cell carcinoma. PLoS One. 2014;9(11):e109318. doi:10.1371/journal.pone.0109318
61. Liu Z, Wei S, Ma H, et al. A functional variant at the miR-184 binding site in TNFAIP2 and risk of squamous cell carcinoma of the head and neck. Carcinogenesis. 2011;32(11):1668–1674. doi:10.1093/carcin/bgr209
62. Etzel BM, Gerth M, Chen Y, et al. Mutation analysis of tumor necrosis factor alpha-induced protein 3 gene in Hodgkin lymphoma. Pathol Res Pract. 2017;213(3):256–260. doi:10.1016/j.prp.2016.11.001
63. Nakamura R, Kataoka H, Sato N, et al. EPHA2/EFNA1 expression in human gastric cancer. Cancer Sci. 2005;96(1):42–47. doi:10.1111/j.1349-7006.2005.00007.x
64. Yuan WJ, Ge J, Chen ZK, et al. Over-expression of EphA2 and Ephrin-A1 in human gastric adenocarcinoma and its prognostic value for postoperative patients. Dig Dis Sci. 2009;54(11):2410–2417. doi:10.1007/s10620-008-0649-4
65. Li Y, Nie Y, Cao J, et al. G-A variant in miR-200c binding site of EFNA1 alters susceptibility to gastric cancer. Mol Carcinog. 2014;53(3):219–229. doi:10.1002/mc.v53.3
66. Mao YY, Jing FY, Jin MJ, et al. rs12904 polymorphism in the 3ʹUTR of EFNA1 is associated with colorectal cancer susceptibility in a Chinese population. Asian Pac J Cancer Prev. 2013;14(9):5037–5041. doi:10.7314/APJCP.2013.14.9.5037
67. Yamamoto H, Tei M, Uemura M, et al. Ephrin-A1 mRNA is associated with poor prognosis of colorectal cancer. Int J Oncol. 2013;42(2):549–555. doi:10.3892/ijo.2012.1750
68. Zhang D, Ren WH, Gao Y, Wang NY, Wu WJ. Clinical significance and prognostic value of pentraxin-3 as serologic biomarker for lung cancer. Asian Pac J Cancer Prev. 2013;14(7):4215–4221. doi:10.7314/APJCP.2013.14.7.4215
69. Infante M, Allavena P, Garlanda C, et al. Prognostic and diagnostic potential of local and circulating levels of pentraxin 3 in lung cancer patients. Int J Cancer. 2016;138(4):983–991. doi:10.1002/ijc.29822
70. Zhang J, Wang TY, Niu XC. Increased plasma levels of pentraxin 3 are associated with poor prognosis of colorectal carcinoma patients. Tohoku J Exp Med. 2016;240(1):39–46. doi:10.1620/tjem.240.39
71. Carmo RF, Aroucha D, Vasconcelos LR, Pereira LM, Moura P, Cavalcanti MS. Genetic variation in PTX3 and plasma levels associated with hepatocellular carcinoma in patients with HCV. J Viral Hepat. 2016;23(2):116–122. doi:10.1111/jvh.12472
72. Dong QZ, Zhao Y, Liu Y, et al. Overexpression of SCC-S2 correlates with lymph node metastasis and poor prognosis in patients with non-small-cell lung cancer. Cancer Sci. 2010;101(6):1562–1569. doi:10.1111/j.1349-7006.2010.01557.x
73. Xing Y, Liu Y, Liu T, et al. TNFAIP8 promotes the proliferation and cisplatin chemoresistance of non-small cell lung cancer through MDM2/p53 pathway. Cell Commun Signal. 2018;16(1):43. doi:10.1186/s12964-018-0254-x
74. Liu K, Qin CK, Wang ZY, Liu SX, Cui XP, Zhang DY. Expression of tumor necrosis factor-alpha-induced protein 8 in pancreas tissues and its correlation with epithelial growth factor receptor levels. Asian Pac J Cancer Prev. 2012;13(3):847–850. doi:10.7314/APJCP.2012.13.3.847
75. Shi TY, Cheng X, Yu KD, et al. Functional variants in TNFAIP8 associated with cervical cancer susceptibility and clinical outcomes. Carcinogenesis. 2013;34(4):770–778. doi:10.1093/carcin/bgt001
76. Liu T, Jiang L, Yu L, Ge T, Wang J, Gao H. Association of TNFAIP8 gene polymorphisms with endometrial cancer in northern Chinese women. Cancer Cell Int. 2019;19:105. doi:10.1186/s12935-019-0827-9
77. Duan D, Zhu YQ, Guan LL, Wang J. Upregulation of SCC-S2 in immune cells and tumor tissues of papillary thyroid carcinoma. Tumour Biol. 2014;35(5):4331–4337. doi:10.1007/s13277-013-1568-3
78. Liu T, Xia B, Lu Y, Xu Y, Lou G. TNFAIP8 overexpression is associated with platinum resistance in epithelial ovarian cancers with optimal cytoreduction. Hum Pathol. 2014;45(6):1251–1257. doi:10.1016/j.humpath.2014.02.005
79. Liu T, Gao H, Yang M, Zhao T, Liu Y, Lou G. Correlation of TNFAIP8 overexpression with the proliferation, metastasis, and disease-free survival in endometrial cancer. Tumour Biol. 2014;35(6):5805–5814. doi:10.1007/s13277-014-1770-y
80. Sun Z, Liu X, Song JH, et al. TNFAIP8 overexpression: a potential predictor of lymphatic metastatic recurrence in pN0 esophageal squamous cell carcinoma after Ivor Lewis esophagectomy. Tumour Biol. 2016;37(8):10923–10934. doi:10.1007/s13277-016-4978-1
81. Zhang L, Liu R, Luan YY, Yao YM. Tumor necrosis factor-α induced protein 8: pathophysiology, clinical significance, and regulatory mechanism. Int J Biol Sci. 2018;14(4):398–405. doi:10.7150/ijbs.23268
82. Zhong M, Li N, Qiu X, et al. TIPE regulates VEGFR2 expression and promotes angiogenesis in colorectal cancer. Int J Biol Sci. 2020;16(2):272–283. doi:10.7150/ijbs.37906
83. STRING. Available from: https://string-db.org/cgi/network.pl?taskId=gAVM4i26EBt2.
84. National Centre for Biotechnology Information (NCBI). Available from: https://www.ncbi.nlm.nih.gov/.
85. GeneCards®: The Human Gene Database. Available from: https://www.genecards.org. Accessed March 18, 2020.
86. UniProt databases. Available from: https://www.uniprot.org/.
87. Stelzer G, Rosen R, Plaschkes I, et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analysis, Current Protocols in Bioinformatics (2016), 54:1.30.1 - 1.30.33. doi:10.1002/cpbi.5.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.