Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Tryptophan Indole Derivatives: Key Players in Type 2 Diabetes Mellitus
Authors Wu Y ![]() , Li T, Chen B
, Li T, Chen B ![]() , Sun Y, Song L
, Sun Y, Song L ![]() , Wang Y, Bian Y, Qiu Y, Yang Z
, Wang Y, Bian Y, Qiu Y, Yang Z ![]()
Received 18 December 2024
Accepted for publication 2 May 2025
Published 12 May 2025 Volume 2025:18 Pages 1563—1574
DOI https://doi.org/10.2147/DMSO.S511068
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Ernesto Maddaloni
Yingli Wu,1,2,* Tiangang Li,1,2,* Biaohua Chen,1,2,* Yi Sun,1,2 Liyun Song,1,2 Yinying Wang,1,2 Yao Bian,1,2 Yong Qiu,1,2 Zhongshan Yang1,2
1Yunnan Provincial Key Laboratory of Integrated Traditional Chinese and Western Medicine for Chronic Disease in Prevention and Treatment, Yunnan University of Chinese Medicine, Kunming, Yunnan, 650000, People’s Republic of China; 2Engineering Research Center of Classic Formula Regulate Immunity in Chronic Disease Prevention and Treatment, Kunming, Yunnan, 650000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yong Qiu; Yunnan University of Chinese Medicine, Kunming, 650000, People’s Republic of China, Email [email protected] Zhongshan Yang, Yunnan University of Chinese Medicine, Kunming, 650000, People’s Republic of China, Email [email protected]
Abstract: Type 2 diabetes mellitus (T2DM) is a complex clinical syndrome characterized by insulin resistance and associated with abnormal amino acid metabolism. Tryptophan is an aromatic dietary amino acid that affects T2DM by regulating glycolipid metabolism and insulin resistance. When tryptophan reaches the intestine, it is converted by gut microbiota and tryptophanase into indole derivatives such as indoleacetic acid, indolepropionic acid, and indolealdehyde. These indole derivatives may enhance insulin sensitivity, stimulate insulin secretion, and exert functions such as lowering blood glucose, regulating hepatic oxidative stress, reducing intestinal inflammation, and improving islet cell morphology by acting on the aryl hydrocarbon receptor (AHR) or Pregnane X receptor (PXR). In summary, this review aims to examine the interactions between tryptophan indole derivatives and T2DM thoroughly, elucidate potential therapeutic approaches, and pinpoint areas for further research.
Keywords: type 2 diabetes mellitus, tryptophan, indole derivatives, aryl hydrocarbon receptor, pregnane X receptor
Graphical Abstract:

Introduction
Relative deficiency in insulin secretion or reduced sensitivity of target organs to insulin and hyperglycemia, hyperlipidemia, and hyperinsulinemia resistance characterize Type 2 diabetes mellitus (T2DM).1 Statistics showed that there were 537 million patients with diabetes across the world by 2021, with projections suggesting an increase to 643 million by 2030 and reaching 783 million by 2045.2 Its development is associated with the functions of the pancreas, kidney, and liver, and recent pharmacological research has indicated the importance of gut microbiota in T2DM.3 A diet rich in fats and sugars frequently results in a deterioration in gut microbiome health, leading to an ecological imbalance within the human gut.4 This imbalance in gut microbiota can contribute to metabolic disorders associated with elevated insulin resistance (IR) levels and inflammation, ultimately triggering T2DM.5 Research indicates that restoring intestinal microbiota balance by regulating tryptophan metabolism can reduce insulin resistance and alleviate T2DM.6
The metabolism of tryptophan significantly influences the pathophysiology of T2DM via various biochemical pathways. Research has shown that patients with T2DM frequently exhibit altered tryptophan metabolism compared to non-diabetic individuals, indicating an increased rate of tryptophan catabolism in those with T2DM.7 Furthermore, dysregulated tryptophan metabolism in T2DM impacts glucose homeostasis and influences the levels of various metabolites, thereby affecting metabolic pathways related to insulin signaling and inflammation.8 This makes it a potential target for therapeutic interventions to improve metabolic health and manage diabetes.
During the metabolism of dietary tryptophan, intestinal microorganisms can convert it into indole, indole-3-acetic acid (IAA), and indole trialdehydes.9 Emerging evidence indicates that indole derivatives are crucial in metabolic regulation. As signaling molecules, they can activate the aryl hydrocarbon receptor(AHR) and Pregnane X receptor (PXR),10 with the activation regulating glycolipid metabolism, enhancing insulin sensitivity, enhancing the function of the intestinal epithelial barrier, hindering the entry of endotoxins, and stimulating the release of the intestinal hormone glucagon-like peptide 1 (GLP-1).11,12 It also has antioxidant effects, protecting the kidneys from damage caused by oxidative stress.13 Additionally, these indole derivatives can influence the microbial composition in the gut and improve the morphology of islet cells, affecting various physiological processes related to glucose metabolism.14 The intricate relationship between indole derivatives and T2DM has garnered significant attention in recent studies. Elucidating the precise mechanisms of tryptophan metabolism and exploring its therapeutic implications are anticipated to provide promising avenues for innovative interventions in T2DM treatment.
Tryptophan Metabolism and Its Impact on Type 2 Diabetes
Tryptophan, an aromatic amino acid, has a relatively high molecular weight. The human body cannot synthesize it independently; thus, it must be obtained through food sources. Tryptophan is a building block for creating various metabolites in the host and microorganisms.15 Around 4%-6% of tryptophan is taken up in the small intestine through the indole metabolic pathway, where it is later transformed into indole derivatives. The process also necessitates the involvement of specific enzymes: indole is produced through the action of tryptophanase; IAA is generated via tryptophan-2 monooxygenase and tryptophan decarboxylase; and 3-indole propionic acid (IPA) is formed through the action of aromatic amino acid transaminase. IPA represents the final product of tryptophan’s reductive metabolism.16 The onset of T2DM is strongly linked to the body’s immune response, the intestines’ inflammation, and the mucosal barrier’s preservation. A high-fat diet damages tight junction proteins, resulting in increased intestinal permeability. This process permits the entry of bacterial byproducts, like lipopolysaccharides, into the bloodstream, initiating systemic inflammation and leading to insulin resistance.17 Research shows that tryptophan is involved in the progression of T2DM. Indoleamine 2,3-dioxygenase (IDO), a marker of tryptophan and serotonin activity, is downregulated in the inflammatory state associated with insulin resistance in T2DM.18 AHR and PXR are critical receptors in the tryptophan metabolism pathway. Combining these two receptors can promote the synthesis of bioactive metabolites from symbiotic microbiota, activating immune responses, increasing the density of the intestinal epithelial barrier, and stimulating the secretion of intestinal hormones. Once secreted, these hormones exert anti-inflammatory and antioxidant effects, thereby regulating host homeostasis.19 Tryptophan derivatives may serve as promising therapeutic targets for treating T2DM by activating AHR and PXR.
AHR is a transcription factor that interacts with regulatory and signal transduction proteins, exhibiting ligand- and context-dependent activity.20 Under normal conditions, AHR resides in the cytoplasm in an inactive form, establishing a complex that includes dormant heat shock protein 90 (Hsp90), AHR interacting protein (AIP), p23, and c-Src protein kinase. When a ligand binds, AHR experiences a conformational shift that reveals the nuclear localization signal, leading to the release of HSP90, translocation of the receptor to the nucleus, and the interaction with AHR karyopherins to form heterodimers that collaboratively oversee the regulation of the expression of target genes downstream (Figure 1).21 Research has shown that the concentration of tryptophan in mice consuming a high-fat diet (HFD) is considerably reduced compared to levels observed under normal conditions. When the microbiota cannot metabolize tryptophan, there is a notable reduction in AHR ligands.22,23 Interestingly, when tryptophan metabolites act as ligands and bind to AHR, they promote cellular defense and regulate the intestinal microbiota-host-immune system. Thus, equilibrating the gut microbiome, decreasing mucosal inflammation, and suppressing the release of interleukin and pro-inflammatory cytokines may participate in the development of T2DM.24 Furthermore, they enhance the release of antimicrobial peptides from the cells lining the intestine, identifying and inhibiting harmful pathogens, boosting the development of beneficial bacteria, and aiding in the conversion of tryptophan into indole derivatives.25 Furthermore, AHR influences glucose and lipid metabolism. Xi Xu et al demonstrated that activation of AHR negatively regulates lipogenic enzymes, such as fatty acid synthase and the regulator of cholesterol metabolism, sterol regulatory element binding protein-1c,26,27 which play crucial roles in lipid metabolism. Moreover, AHR activators have the potential to lower blood glucose levels after fasting, enhance the metabolism of glucose and insulin abnormalities, and boost GLP-1 secretion in diabetic mice that are on a high-fat diet.22,28 Overall, the relationship between T2DM, tryptophan, and AHR is complex and multifaceted, involving the interaction between metabolic regulation and inflammation.
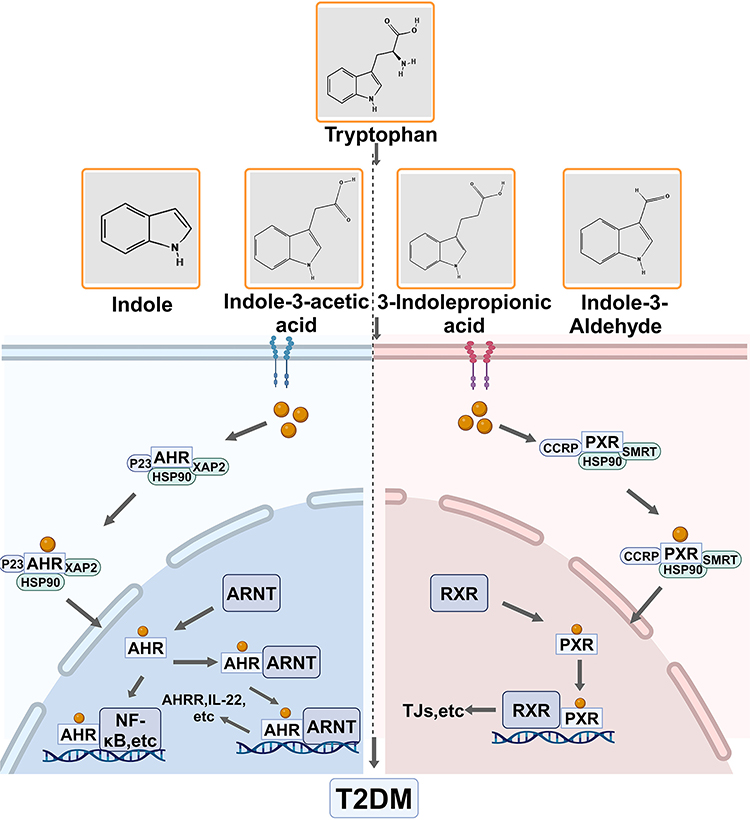 |
Figure 1 Tryptophan metabolic pathway.Tryptophan binds to AHR and PXR, resulting in conformational changes that promote the release of HSP90. This receptor is readily nucleated, facilitating the formation of heterodimers and co-regulating the expression of downstream target genes, which contributes to the development of T2DM. Created in BioRender. Wu, Y. (2024) https://BioRender.com/m65p141. Abbreviations: ARNT, Aryl Hydrocarbon Receptor Nuclear Translocator; AHRR, Aryl Hydrocarbon Receptor Repressor; RXR, Retinoid X receptor; TJs, tight junctions;HSP90,Heat shock protein 90. |
PXR belongs to the nuclear receptor superfamily and mainly acts as a transcription factor. It is activated by different ligands, forms dimers with RXR, and attaches to particular DNA response elements situated in the regulatory areas of target genes, thereby aiding in transcriptional regulation. PXR plays a crucial role in the liver and intestines, participating in various physiological functions such as maintaining energy balance, regulating immune responses, managing glucose metabolism, and overseeing lipid regulation (Figure 1).29 Research has demonstrated that metabolites resulting from the microbial breakdown of tryptophan can act as ligands for PXR, thereby influencing intestinal health and regulating immune responses. Indole derivatives have been shown to promote PXR activation, which inhibits inflammatory pathways, particularly the NF-κB signaling cascade.30,31 This underscores the potential of indole derivatives as therapeutic options for diseases characterized by inflammation. Moreover, T2DM is significantly associated with intestinal inflammation and damage.32 Thus, it is imperative to investigate further the structure-activity relationship of indole derivatives and their effects on PXR signaling to develop new treatments for T2DM.
Tryptophan Derivative Indole as a Regulator for Pancreatic Protection and Glucose Metabolism in T2DM
Tryptophan is a vital amino acid, while indole acts as a crucial signaling molecule with anti-inflammatory properties within the intestinal microbial metabolism pathway of tryptophan. Indole is produced by tryptophanase in response to both Gram-negative and Gram-positive bacteria, including Escherichia coli and Bacteroidetes. These bacteria can maintain elevated levels of indole in the intestine, contributing to the host’s overall health.33–35 The relationship between indole and T2DM is complex. Indole levels are significantly reduced in patients with T2DM, a phenomenon that involves mechanisms related to insulin resistance, gut microbiome interactions, and the management of glucose and lipid metabolism (Table 1).14,20 Thus, indole may play a pivotal role as an essential signaling factor in T2DM.
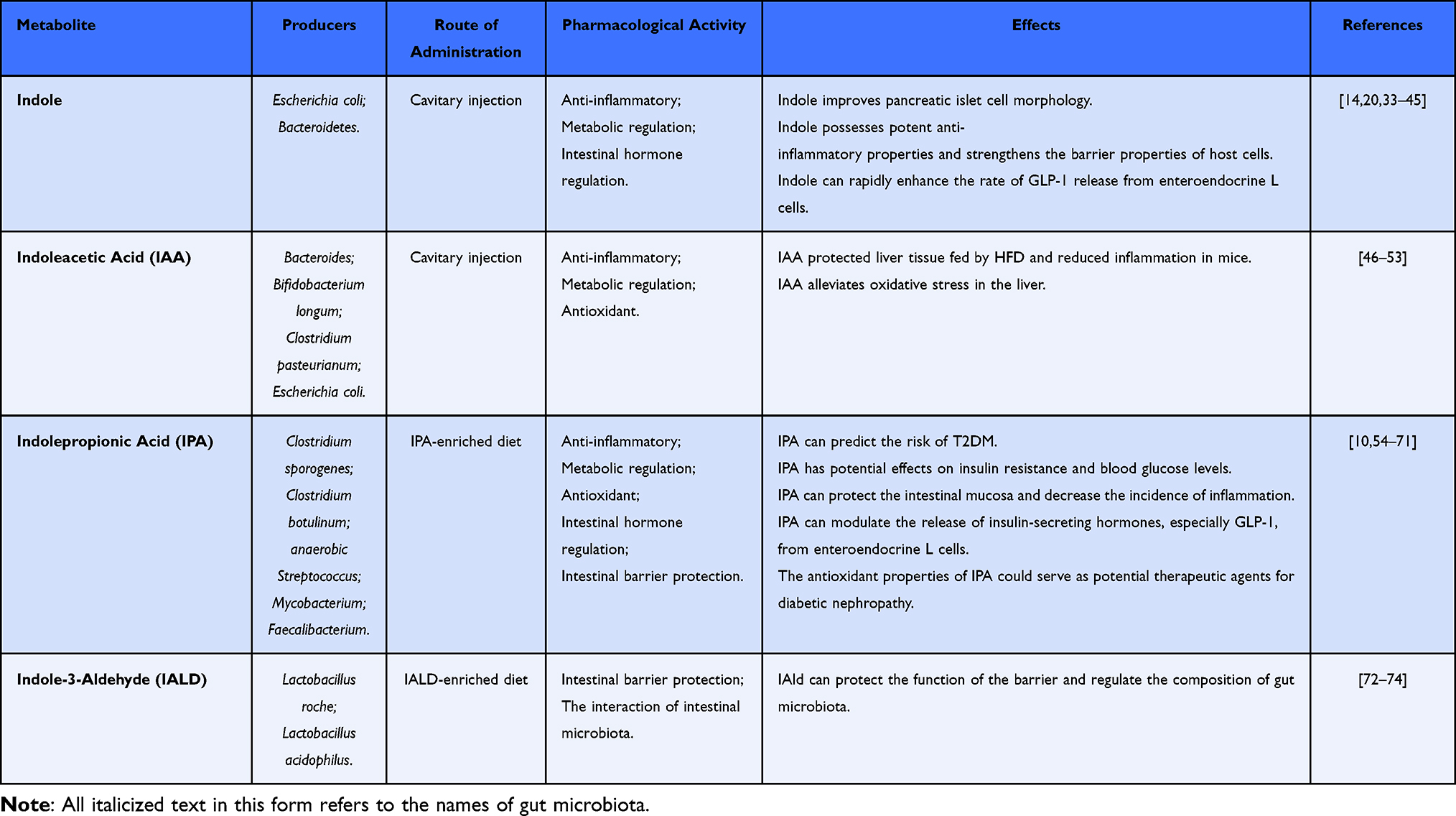 |
Table 1 Producers and Effects of Indole Derivatives |
The Spearman correlation analysis indicated a strong positive relationship between indole and various pancreatic β cell function indicators while showing a negative correlation between blood glucose levels and the length of diabetes.14 The shape and structure of pancreatic islet cells were regular, exhibiting precise edges. Under normal conditions, β cells are located at the center of the islet, while α cells are situated on the periphery.36,37 Fengying Yang sectioned the pancreas of db/db diabetic mice fed a high-fat diet and demonstrated that the morphological structure of the islets was significantly compromised, characterized by unclear boundaries, loosely arranged cells, and irregular shapes. The area ratio of α cells to β cells was unbalanced, with an increase in the percentage of α cells and a corresponding decrease in β cells. Notably, up to 60% of β cells lost their function, producing significantly elevated blood sugar levels. Following indole intervention, the morphological structure of the islets was partially restored; the borders of the islets improved, and the internal cells were arranged more tightly and orderly.14 Therefore, indole plays a significant role in the morphological changes of islets in diabetic mice.
T2DM is defined as a chronic condition marked by low-grade inflammation. Recent studies have demonstrated that indole possesses potent anti-inflammatory properties, mitigating inflammatory response symptoms by influencing immune cells and elevating IL-10 levels in mice subjected to a high-fat diet.25,38,39 The primary mechanism involves the recruitment of pro-inflammatory macrophages, predominantly of the M1 type, which are attracted by pancreatic β cells and infiltrate local islets. This infiltration enhances the expression of macrophage-related genes and increases the levels of pro-inflammatory cytokines in the circulatory system. Consequently, this process activates the NF-κB signaling pathway with the Janus kinase inflammatory cascade, exacerbating endoplasmic reticulum stress and jointly inhibiting glucose-stimulated insulin release in pancreatic β cells, increasing blood glucose levels.14,40 Conversely, when tryptophan is metabolized by intestinal flora into indole, it is recognized by intestinal epithelial cells, where it participates in regulating genes associated with tight junctions and forming the actin cytoskeleton. This action enhances the polarized transepithelial resistance of HCT-8 intestinal epithelial cells, inhibits the activation of NF-κB, reduces the adhesion ability of Escherichia coli, maintains resistance to pathogens, strengthens the barrier properties of host cells, and helps to sustain controlled inflammation.41
GLP-1 is a type of incretin produced by the L cells of the intestinal epithelium that enhances insulin release from pancreatic β cells while concurrently inhibiting glucagon secretion from pancreatic α cells. This dual action significantly influences glucose metabolism, reduces appetite, and delays gastric emptying.42 Research has demonstrated that indole can modulate GLP-1 secretion through various mechanisms.43,44 A key finding is that indole can rapidly enhance the rate of GLP-1 release from enteroendocrine L cells. It achieves this by expanding the action potential in these cells. This reduces the current flowing through voltage-gated potassium channels, thereby extending the activation period of channels regulated by calcium. This sequence of events increases intracellular calcium levels, thereby enhancing GLP-1 secretion.14,45 Consequently, indole serves as a significant therapeutic approach for the treatment of T2DM. It restores GLP-1 production in mice, protects pancreatic β cells and the intestinal barrier, reduces inflammation, and improves blood sugar levels and insulin sensitivity.
Tryptophan Derivative Indole-3-Acetic Acid Has the Potential to Reduce Liver Inflammation and Oxidative Stress in T2DM
IAA is a metabolite of tryptophan generated by Bacteroides thetaiotaomicron, Bifidobacterium longum, Clostridium pasteurianum, and Escherichia coli, with its synthesis catalyzed by tryptophan monooxygenase and indole-3-acetamide hydrolase.46 Research indicates that IAA possesses anti-inflammatory properties and is crucial in mitigating oxidative stress in the liver, thereby enhancing liver metabolism.47 In animal models of T2DM, significant infiltration of local inflammatory cells, along with degeneration and necrosis of hepatocytes, has been observed. Moreover, the liver functions as the central location for the generation of glucose and the metabolism of lipids, making it especially susceptible to oxidative stress. Elevated levels of reactive oxygen species (ROS) disrupt the insulin signaling pathway, which results in diminished insulin sensitivity and exacerbates hyperglycemia, a defining characteristic of T2DM (Table 1).48 Therefore, protecting liver tissue might be an effective strategy for managing T2DM.
A significant link exists between T2DM and non-alcoholic fatty liver disease (NAFLD). The inflammatory processes associated with T2DM may lead to liver damage.49 Yun Ji et al found that IAA has a significant relationship with the liver in the context of NAFLD. Following liver tissue damage, a pro-inflammatory response is triggered. This enhances the infiltration of macrophages into hepatocytes while also elevating the concentrations of inflammatory cytokines and monocyte chemoattractant protein-1 (MCP-1). However, IAA intervention significantly improved the inflammatory infiltration and morphology of hepatocytes, downregulating parameters related to inflammatory factors and participating in glucose, lipid, and cholesterol metabolism through its interaction with AHR. By suppressing the genes involved in lipogenesis, such as Scd1and PPARγ, IAA lowers fasting blood glucose concentrations, insulin resistance index, and alanine aminotransferase activity, thereby mitigating liver damage.47 Concurrently, it restores levels of oxidative stress-related indicators in liver tissue induced by high-fat diet, such as liver superoxide dismutase (SOD) activity.50 Therefore, investigating whether IAA can ameliorate diabetes symptoms through liver protection represents a promising therapeutic direction.
IAA is also involved in regulating gut microbiota, and alterations in the composition of gut microbiota can influence systemic inflammation and metabolic processes, including insulin sensitivity and glucose metabolism.51 The anti-inflammatory properties of IAA may offer potential benefits in diabetes management. The process of inflammation is crucial in the progression of T2DM, and substances that reduce the inflammatory reaction may assist in relieving specific symptoms linked to this condition.52,53 Overall, while research on IAA and its effects on diabetes is still developing, existing studies indicate that it could significantly influence metabolic health through immune modulation, interactions with gut microbiota, and direct effects on glucose metabolism.
Tryptophan Derivative Indole Propionic Acid as a Potential Therapeutic Agent for Intestinal Barrier and Antioxidant Effects in Type 2 Diabetes Mellitus
IPA is a potent oxidant generated through the deamination of dietary tryptophan, catalyzed by enzymes secreted by intestinal microorganisms. The levels of IPA are associated with fiber intake and can accumulate in serum.54 Gas-liquid chromatography-mass spectrometry analysis has confirmed that Clostridium sporogenes and Clostridium botulinum can metabolize tryptophan to IPA via the indole pathway.46 Dodd et al cultured 36 strains in a tryptophan-containing medium, revealing that anaerobic Streptococcus and Clostridium produced IPA, while Mycobacterium, Faecalibacterium, and Faecobacter prowazekii showed a positive correlation with IPA levels.75 Research indicates that IPA is a promising candidate for preventing and managing T2DM. Its ability to enhance intestinal health, modulate inflammatory responses, and improve metabolic parameters underscores its potential as a therapeutic agent 10 (Table 1).
IPA has been identified as a possible biomarker for predicting the risk of T2DM. By measuring IPA levels, it is possible to distinguish between different stages of T2DM and predict the associated risk.54 In research involving animals, a negative relationship was noted between serum IPA levels and the likelihood of developing T2DM in rats, observed both before the disease began and three months following its commencement.55,56 Data from the Finland Diabetes Prevention Study (DPS) further corroborate that higher levels of IPA are associated with a reduced likelihood of developing T2DM in individuals with impaired glucose tolerance, and these levels are inversely related to low-grade inflammation.57 The utilization of gut microbiota-derived metabolites holds significant promise for early screening and prevention strategies for T2DM.
IPA has emerged as a significant compound in the study of glucose metabolism, mainly due to its potential effects on insulin resistance and blood glucose levels. A study involving 1,018 women of middle age from the Twins UK cohort examined how circulating IPA levels, the composition of gut microbiota, and metabolic health are interrelated. The findings indicated that IPA levels are inversely associated with visceral fat content, insulin resistance, and fasting blood glucose levels.54 In a separate study, Anders Abildgaard et al administered isolated IPA extracts to rats to evaluate their effects on glucose metabolism and blood glucose concentrations. After six weeks of HFD enriched with IPA, the assessment of blood glucose levels in male rats with diabetes indicated that the intervention with IPA notably decreased fasting levels of blood glucose, plasma insulin, and the HOMA-IR index.58 One of the primary mechanisms through which IPA exerts its effects is by enhancing the function of glucose transporter, which is crucial for insulin-mediated glucose uptake.59 The improved translocation of GLUT to the plasma membrane facilitates glucose uptake from the bloodstream into muscle cells, reducing blood glucose levels and improving insulin sensitivity.60
T2DM is characterized by intestinal microbiota disturbances, intestinal barrier damage, heightened epithelial permeability, and decreased amounts of zonula occludens-1 (ZO-1) and occludin within the ileum.61 A primary mechanism involves the formation of a complex between lipopolysaccharide and glucose phosphate isomerase-anchored protein CD4, which is recognized by toll-like receptor 4 (TLR4) on the surface of monocytes and macrophages. This recognition activates NF-κB signaling, releasing inflammatory mediators contributing to the inflammatory response, insulin resistance, and eventually diabetes.62 Furthermore, research has shown that IPA influences various cellular activities, including proliferation, differentiation, and preserving barrier function in human intestinal epithelial cells. The introduction of spores from Clostridium that produces IPA into mice fed a high-fat diet has been demonstrated to enhance intestinal barrier function, modulate the composition of gut microbiota, prevent microbial imbalance, preserve the homeostasis of intestinal epithelial cells, decrease intestinal permeability, and increase the levels of ZO-1 as well as proteins such as occluding. IPA primarily stimulates cell proliferation through a mechanism involving the ERK2 pathway and regulates the expression of other proteins, such as c-Jun, which is essential for preserving the integrity of the intestinal barrier.63,64 Furthermore, Matthew Jennis et al found that IPA can improve the intestinal permeability of the T84 intestinal epithelial cell monolayer with compromised barrier function and mitigate the damaging effects of pro-inflammatory cytokines on T84 cells. Inflammatory conditions enhance the transcription of GLUT5 in intestinal epithelial cells, and IPA effectively reverses the increase in fructose transporter GLUT5 mRNA induced by pro-inflammatory interferon γ, thereby protecting the intestinal mucosa and decreasing the incidence of inflammation.59
The possible protective role of serum IPA in T2DM could be linked to its capacity to modulate the release of insulin-secreting hormones, especially GLP-1, from enteroendocrine L cells.44,65,66 Diabetic patients often exhibit a significant loss of GLP-1 functionality, which is crucial for enhancing glucose metabolism and overall metabolic health through the protection of β cells, reduction of apoptosis, and promotion of β cell proliferation and regeneration.67
As a potent oxidant, IPA protects β cells against damage induced by metabolic changes, oxidative stress, and amyloid accumulation.68 Diabetic nephropathy (DN), a complication of T2DM, has its core pathogenesis rooted in the increased production of reactive oxygen species (ROS) induced by hyperglycemia.69 The elevated ROS production further exacerbates oxidative stress, causing direct damage to podocytes, mesangial cells, and endothelial cells, leading to proteinuria and renal tubulointerstitial fibrosis.70 Studies have shown that Sorafenib and Lenvatinib are multi-target tyrosine kinase inhibitors in treating renal cell carcinoma. However, their use is often limited due to adverse effects related to oxidative stress.71 In a rat experiment where potassium bromate (KBrO3) induced renal tumors and lipid oxidation reactions, it was also observed that melatonin and IPA reduced lipid oxidative damage in the kidneys and serum of rats.13 In the case of IPA treatment alone, it was also found that IPA supplementation alleviated aflatoxin B1 (AFB1)-mediated redox imbalance in rat serum by upregulating antioxidant mediators and enzymes such as TSH, SOD, CAT, and GPx, thereby reducing the formation of reactive oxygen species, lipid peroxidation, and DNA adducts. Additionally, it can alleviate damage to liver and kidney tissues.13 Moreover, in a study involving participants with normal renal function, a diet rich in polyphenols (PR) was found to promote an increase in serum concentrations of IPA, which was negatively correlated with serum levels of C-reactive protein (CRP). However, no changes were observed in subjects with impaired renal function.76 These results collectively indicate the antioxidative effects of IPA on the kidneys, suggesting its potential as a therapeutic agent for diabetic nephropathy. These results indicate that IPA could be a potential alternative for treating metabolic disorders linked to insulin resistance.
Tryptophan Derivative IAld Has the Potential to Prevent Intestinal Inflammation and Alleviating Metabolic Parameters in T2DM
3-IAld is generated by the gut microbiota, and Lactobacillus Roche and Lactobacillus acidophilus metabolize tryptophan to 3-IAld,72 which exhibits anti-inflammatory properties that may aid in the management of diseases associated with chronic inflammation.73 Its mechanism of action seems to involve the AhR and the NF-κB signaling pathway. Research indicates that the therapeutic effects of 3-IAld are entirely diminished when AhR is knocked down, underscoring the essential role of AhR activation.72 In addition to alleviating inflammation, 3-IAld can promote a healthier intestinal environment, protect barrier function, regulate the composition of gut microbiota, support the growth of beneficial bacteria, and simultaneously inhibit pathogenic strains. Furthermore, inflammation and disruption of gut microbiota are essential factors in the progression of T2DM (Table 1).73 Thus, it is hypothesized that IAld could be crucial in managing T2DM.
Jiaqi Zhang et al found that IAld restored intestinal epithelial barrier function in mice fed an HFD. After administering FITC–dextran to mice orally, intestinal epithelium leakage was measured by serum fluorescence. Endotoxin leakage, decreased epithelial cell proliferation, and increased proinflammatory cytokine expression were observed in the intestinal epithelium of HFD-fed mice; however, the colonic tissue structure of the mice improved significantly, and inflammatory cell infiltration was alleviated in the IAld treatment group. After the 3-IAld intervention, the existing symptoms were alleviated considerably, NLRP3 and inflammatory cytokine levels decreased, and IL-lRa levels increased, indicating that 3-IAld could promote intestinal epithelial function and reduce inflammation in vivo.74 Additionally, HFD feeding affected metabolic parameters in the mice, and body weight and serum glucose levels measured in control and model mice showed that the HFD-fed mice exhibited significantly greater body weight and elevated blood glucose levels compared to the IAld control group after 8 weeks of treatment. The glucose tolerance test results showed that 3-IAld reduces blood glucose levels. Therefore, feeding an HFD for an extended period can trigger intestinal inflammation and alter glucose metabolism, which can be prevented and treated by treating it with IAld, which is a promising therapeutic target.73,74 Further research in the future may explore its clinical applications, potentially leading to new therapeutic strategies that utilize the benefits of this indole metabolite.
Discussion
Basic research findings show that the gut microbiota produces microbial metabolites that enhance insulin resistance in patients with T2DM through various pathways affecting host glucose metabolism.5 Among these, tryptophan metabolites have emerged as potential targets for T2DM treatment, primarily by acting as ligands for AHR and PXR involved in the progression of T2DM.77 Beyond mediating host-microbial interactions, these metabolites can enhance islet cell morphology, alleviate hepatic oxidative stress and inflammation, and improve glucose and lipid metabolism, paving the way for novel prevention and treatment strategies for T2DM.
In recent years, significant progress has been made in the research on applying tryptophan in clinical treatment, demonstrating its potential clinical value as a novel therapeutic agent in pathological processes such as metabolic syndrome, cardiovascular diseases, and urinary system disorders. Supplementing with a tryptophan-rich diet can effectively improve pancreatic islet function in T2DM. Studies have confirmed that providing young T2DM rats with an L-tryptophan-rich diet can effectively prevent the failure of β-cell function in their old age.78 It can also promote intestinal motility and treat cardiovascular diseases through anti-inflammatory mechanisms. Barbora Waclawiková et al demonstrated that 5-hydroxytryptophan, an intermediate metabolite of L-tryptophan in serotonin biosynthesis, stimulates intestinal motility and improves cardiovascular diseases by activating L-type calcium channels located on colonic smooth muscle cells.79 Interestingly, in patients with atherosclerotic cardiovascular disease and volunteers with coronary heart disease, supplementation of the tryptophan-derived metabolite IAA has been found to inhibit the TLR4/MyD88/NF-κB pathway in M1 macrophages, promote M2 macrophage polarization, restore the M1/M2 polarization balance, and ultimately alleviate aortic inflammation.80 The tryptophan metabolite kynurenine (KTR) possesses immunomodulatory and vasoactive properties, and the KTR to Trp ratio can predict the development of T2DM in individuals with coronary artery disease (CAD). By evaluating the plasma KTR and the KTR to Trp ratio in urine, it was found that plasma KTR showed no significant correlation with T2DM risk. In contrast, urinary KTR demonstrated a strong positive correlation. It has been confirmed that urinary KTR is a strong predictor of T2DM in CAD patients.81 Surprisingly, reducing the Kyn/Trp ratio protects the heart and has therapeutic effects on the kidneys. SempastianFilippas-Ntekouan et al proposed that after administering Dapagliflozin, changes in serum metabolites of T2DM patients were detected, showing a significant increase in tryptophan levels and a decrease in the Kyn/Trp ratio. This indicates that Dapagliflozin exerts protective effects on the kidneys and heart by increasing serum tryptophan levels and reducing IDO activity.82
However, tryptophan metabolites have shown significant bidirectional regulatory properties of the AHR pathway in the study of disease regulatory mechanisms. In metabolic diseases, they exert metabolic regulatory effects by activating the AHR signaling pathway, while in kidney diseases, they improve metabolic regulation by downregulating the expression levels of AHR. HuaMiao et al demonstrated that Lactobacillus improves membranous nephropathy by producing tryptophan indole derivatives that inhibit the aryl hydrocarbon receptor pathway. In patients with impaired renal function and BSA-induced MN rats, beneficial bacteria reduction and alterations in serum IAld and tryptophan indole derivatives are also observed. The primary mechanism is that the intrarenal AhR signaling pathway is activated by the decrease in indole derivatives acting as ligands. The model group exhibited increased intrarenal mRNA expression of AhR and its target genes CYP1A1, CYP1A2, and CYP1B1, accompanied by upregulation of nuclear AhR in renal tissues.83 Furthermore, in chronic renal failure, treatment with L. johnsonii significantly increased serum IAld levels in model rats by inhibiting the mRNA expression of AHR and its four downstream genes in renal tissues of the model group and suppressing intrarenal AHR nuclear translocation.84 This tissue-specific regulatory phenomenon indicates that the AHR signaling pathway has distinct pathophysiological functions in different disease microenvironments.
Meanwhile, the therapeutic effects of indole derivative-derived compounds on T2DM and its associated complications have been substantiated. Diindolylmethane (DIM), a compound naturally produced from indole-3-carbinol, enhances insulin sensitivity in diabetic models by inhibiting key signaling pathways, particularly protein kinase C (PKC) and transforming growth factor-beta (TGF-β1), which are associated with renal injury.85 Furthermore, the synthesis of novel triazole indole derivatives has been explored for their potential as alpha-glucosidase inhibitors, which may aid in managing postprandial hyperglycemia by delaying carbohydrate absorption.86 These properties have made indole derivatives a focal point in research for developing novel therapeutic strategies to manage T2DM and its associated complications effectively. As research delves deeper into the mechanisms of action of these derivatives, they may play a pivotal role in diabetes management programs. Although existing studies have preliminarily revealed the relationship between tryptophan-induced derivatives and T2DM, some limitations remain. Firstly, tryptophan metabolism is a complex process involving multiple pathways. Current research primarily focuses on the effects of these derivatives on AHR and PXR, overlooking other potential pathways. Secondly, the experimental models used in most studies are mainly based on mice or small human populations, which limits comprehensive understanding and the conclusions’ generalizability. Additionally, there is controversy regarding the effects of tryptophan-inducer derivatives, as they have both positive and negative impacts, and their clinical applicability remains to be validated. 3-Methylindole is one of the pulmonary toxicants present in cigarette smoke, and its inhalation may promote the development of lung cancer.87 Indoxyl sulfate can accelerate endothelial cell senescence, reduce NO production, and increase reactive oxygen species generation, thereby inducing endothelial dysfunction. In patients with chronic kidney disease, it exacerbates aortic calcification and vascular stiffness.88 Therefore, further research is necessary to evaluate its potential side effects, as ensuring safety is a key factor in advancing this field.
Acknowledgments
The graphical abstract was Created in BioRender. Wu, Y. (2024) https://BioRender.com/o95z752. Supported by open research fund program of Yunnan Key Laboratory of Integrated Traditional Chinese and Western Medicine for Chronic Disease in Prevention and Treatment.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by a grant from the National Natural Science Foundation of China (82360017). We also received a grant from Yunnan Provincial Science and Technology Department (202101AZ070001-012, 202201AS070084, 202301AZ070001-004, YNWR- QNBJ-2019-069, 202403AC100007), the central government guides local funds for scientific and technological development (202407AB110020).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Liu Y, Xu Z, Huang H, et al. Fucoidan ameliorates glucose metabolism by the improvement of intestinal barrier and inflammatory damage in type 2 diabetic rats. Int J Biol Macromol. 2022;201:616–629. doi:10.1016/j.ijbiomac.2022.01.102
2. Gupta S, Dominguez M, Golestaneh L. Diabetic kidney disease: an update. Med Clin North Am. 2023;107:689–705. doi:10.1016/j.mcna.2023.03.004
3. Li MY, Duan JQ, Wang XH, et al. Inulin inhibits the inflammatory response through modulating enteric glial cell function in type 2 diabetic mellitus mice by reshaping intestinal flora. ACS Omega. 2023;8:36729–36743. doi:10.1021/acsomega.3c03055
4. Christine PS, Saskia de P, Kenneth M, et al. The double burden of malnutrition—further perspective. The Lancet. 2020;396(10254):814–815. doi:10.1016/S0140-6736(20)31360-X
5. Gao R, Meng X, Xue Y, et al. Bile acids-gut microbiota crosstalk contributes to the improvement of type 2 diabetes mellitus. Front Pharmacol. 2022;13:1027212. doi:10.3389/fphar.2022.1027212
6. Calvani R, Rodriguez-Mañas L, Picca A, et al. Identification of a circulating amino acid signature in frail older persons with type 2 diabetes mellitus: results from the metabofrail study. Nutrients. 2020;12:199. doi:10.3390/nu12010199
7. Matsuoka K, Kato K, Takao T, et al. Concentrations of various tryptophan metabolites are higher in patients with diabetes mellitus than in healthy aged male adults. Diabetol Int. 2017;8:69–75. doi:10.1007/s13340-016-0282-y
8. Qi Q, Li J, Yu B, et al. Host and gut microbial tryptophan metabolism and type 2 diabetes: an integrative analysis of host genetics, diet, gut microbiome and circulating metabolites in cohort studies. Gut. 2022;71:1095–1105. doi:10.1136/gutjnl-2021-324053
9. Su X, Gao Y, Yang R. Gut microbiota-derived tryptophan metabolites maintain gut and systemic homeostasis. Cells. 2022;11:2296. doi:10.3390/cells11152296
10. Niu B, Pan T, Xiao Y, et al. The therapeutic potential of dietary intervention: based on the mechanism of a tryptophan derivative-indole propionic acid on metabolic disorders. Crit Rev Food Sci Nutr. 2024;1–20. doi:10.1080/10408398.2023.2299744
11. Hu W, Yan G, Ding Q, et al. Update of Indoles: promising molecules for ameliorating metabolic diseases. Biomed Pharmacother. 2022;150:112957. doi:10.1016/j.biopha.2022.112957
12. Zhao L, Zhang F, Ding X, et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science. 2018;359:1151–1156. doi:10.1126/science.aao5774
13. Owumi SE, Arunsi UO, Oyelere AK. The protective effect of 3-indolepropanoic acid on aflatoxin B1-induced systemic perturbation of the liver and kidney function in rats. Fundam Clin Pharmacol. 2023;37:369–384. doi:10.1111/fcp.12842
14. Yang FY. Impact of Gut Microbiota and Serum Metabolites on Pancreatic Islet Function in Type 2 Diabetes Mellitus.[Dissertation]. China: Huazhong University of Science & Technol; 2021.
15. Luo Z, Liu Y, Wang X, Fan F, Yang Z, Luo D. Exploring tryptophan metabolism: the transition from disturbed balance to diagnostic and therapeutic potential in metabolic diseases. Biochem Pharmacol. 2024;230:116554. doi:10.1016/j.bcp.2024.116554
16. Sári Z, Mikó E, Kovács T, et al. Indolepropionic acid, a metabolite of the microbiome, has cytostatic properties in breast cancer by activating AHR and PXR receptors and inducing oxidative stress. Cancers. 2020;12. doi:10.3390/cancers12092411
17. Li D, Zhong J, Zhang Q, Zhang J. Effects of anti-inflammatory therapies on glycemic control in type 2 diabetes mellitus. Front Immunol. 2023;14:1125116. doi:10.3389/fimmu.2023.1125116
18. Chen T, Zheng X, Ma X, et al. Tryptophan Predicts the Risk for Future Type 2 Diabetes. PLoS One. 2016;11:e0162192. doi:10.1371/journal.pone.0162192
19. Nieves KM, Hirota SA, Flannigan KL. Xenobiotic receptors and the regulation of intestinal homeostasis - harnessing the chemical output of the intestinal microbiota. Am J Physiol Gastrointest Liver Physiol. 2021;322:G268–G281. doi:10.1152/ajpgi.00160.2021
20. Stockinger B, Shah K, Wincent E. AHR in the intestinal microenvironment: safeguarding barrier function. Nat Rev Gastroenterol Hepatol. 2021;18:559–570. doi:10.1038/s41575-021-00430-8
21. Kudo I, Hosaka M, Haga A, et al. The regulation mechanisms of AhR by molecular chaperone complex. J Biochem. 2018;163:223–232. doi:10.1093/jb/mvx074
22. Roh E, Kwak SH, Jung HS, et al. Serum aryl hydrocarbon receptor ligand activity is associated with insulin resistance and resulting type 2 diabetes. Acta Diabetol. 2015;52:489–495. doi:10.1007/s00592-014-0674-z
23. Liu Y, Yang K, Jia Y, et al. Gut microbiome alterations in high-fat-diet-fed mice are associated with antibiotic tolerance. Nat Microbiol. 2021;6:874–884. doi:10.1038/s41564-021-00912-0
24. Lamas B, Richard ML, Leducq V, et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat Med. 2016;22:598–605. doi:10.1038/nm.4102
25. Liu D, Zhang S, Li S, et al. Indoleacrylic acid produced by Parabacteroides distasonis alleviates type 2 diabetes via activation of AhR to repair intestinal barrier. BMC Biol. 2023;21:90. doi:10.1186/s12915-023-01578-2
26. Lee JS, Cella M, McDonald KG, et al. AHR drives the development of gut ILC22 cells and postnatal lymphoid tissues via pathways dependent on and independent of Notch. Nat Immunol. 2011;13:144–151. doi:10.1038/ni.2187
27. Xu X, Sun S, Liang L, et al. Role of the aryl hydrocarbon receptor and gut microbiota-derived metabolites indole-3-acetic acid in sulforaphane alleviates hepatic steatosis in mice. Front Nutr. 2021;8:756565. doi:10.3389/fnut.2021.756565
28. Kalra S. Choosing appropriate glucagon-like peptide 1 receptor agonists: a patient-centered approach. Diabetes Ther. 2014;5:333–340. doi:10.1007/s13300-014-0058-y
29. Ma X, Chen J, Tian Y. Pregnane X receptor as the “sensor and effector” in regulating epigenome. J Cell Physiol. 2015;230:752–757. doi:10.1002/jcp.24838
30. Oladimeji PO, Lin W, Brewer CT, Chen T. Glucose-dependent regulation of pregnane X receptor is modulated by AMP-activated protein kinase. Sci Rep. 2017;7:46751. doi:10.1038/srep46751
31. Niu H, Zhou X, Gong P, et al. Effect of Lactobacillus rhamnosus MN-431 producing indole derivatives on complementary feeding-induced diarrhea rat pups through the enhancement of the intestinal barrier function. Mol Nutr Food Res. 2022;66:e2100619. doi:10.1002/mnfr.202100619
32. Li H, Illés P, Karunaratne CV, et al. Deciphering structural bases of intestinal and hepatic selectivity in targeting pregnane X receptor with indole-based microbial mimics. Bioorg Chem. 2021;109:104661. doi:10.1016/j.bioorg.2021.104661
33. Lee JH, Wood TK, Lee J. Roles of indole as an interspecies and interkingdom signaling molecule. Trends Microbiol. 2015;23:707–718. doi:10.1016/j.tim.2015.08.001
34. Niu H, Zhou X, Liu P, et al. Lactobacillus rhamnosus MN-431 metabolic tryptophan alleviates complementary food-induced diarrhea through PXR-NF-κB pathway and AHR-Th17 cell response pathways. Mol Nutr Food Res. 2023;67:e2200530. doi:10.1002/mnfr.202200530
35. Benech N, Rolhion N, Sokol H. Tryptophan metabolites get the gut moving. Cell Host Microbe. 2021;29:145–147. doi:10.1016/j.chom.2021.01.009
36. Ardestani A, Maedler K. MST1: a promising therapeutic target to restore functional beta cell mass in diabetes. Diabetologia. 2016;59:1843–1849. doi:10.1007/s00125-016-3892-9
37. Röder PV, Wu B, Liu Y, Han W. Pancreatic regulation of glucose homeostasis. Exp Mol Med. 2016;48:e219. doi:10.1038/emm.2016.6
38. Yelamanchi SD, Solanki HS, Radhakrishnan A, et al. Signaling network map of the aryl hydrocarbon receptor. J Cell Commun Signal. 2016;10:341–346. doi:10.1007/s12079-016-0341-y
39. Ma L, Li H, Hu J, et al. Indole alleviates diet-induced hepatic steatosis and inflammation in a manner involving myeloid Cell 6-Phosphofructo-2-kinase/fructose-2,6-biphosphatase 3. Hepatology. 2020;72:1191–1203. doi:10.1002/hep.31115
40. Hu X, Duan T, Wu Z, Tang C, Cao Z. Puerarin Inhibits the PERK-eIF2[Formula: see text]-ATF4-CHOP Pathway through Inactivating JAK2/STAT3 Signal in Pancreatic beta-Cells. Am J Chin Med. 2021;49:1723–1738. doi:10.1142/S0192415X21500816
41. Sun M, Ma N, He T, Johnston LJ, Ma X. Tryptophan (Trp) modulates gut homeostasis via aryl hydrocarbon receptor (AhR). Crit Rev Food Sci Nutr. 2020;60:1760–1768. doi:10.1080/10408398.2019.1598334
42. Drucker DJ. Mechanisms of action and therapeutic application of glucagon-like peptide-1. Cell Metab. 2018;27:740–756. doi:10.1016/j.cmet.2018.03.001
43. MacEachern SJ, Keenan CM, Papakonstantinou E, Sharkey KA, Patel BA. Alterations in melatonin and 5-HT signalling in the colonic mucosa of mice with dextran-sodium sulfate-induced colitis. Br J Pharmacol. 2018;175:1535–1547. doi:10.1111/bph.14163
44. Chimerel C, Emery E, Summers DK, Keyser U, Gribble FM, Reimann F. Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep. 2014;9:1202–1208. doi:10.1016/j.celrep.2014.10.032
45. Reimann F, Maziarz M, Flock G, Habib AM, Drucker DJ, Gribble FM. Characterization and functional role of voltage gated cation conductances in the glucagon-like peptide-1 secreting GLUTag cell line. J Physiol. 2005;563:161–175. doi:10.1113/jphysiol.2004.076414
46. Sinha AK, Laursen MF, Brinck JE, et al. Dietary fibre directs microbial tryptophan metabolism via metabolic interactions in the gut microbiota. Nat Microbiol. 2024;9:1964–1978. doi:10.1038/s41564-024-01737-3
47. Ji Y, Gao Y, Chen H, Yin Y, Zhang W. Indole-3-acetic acid alleviates nonalcoholic fatty liver disease in mice via attenuation of hepatic lipogenesis, and oxidative and inflammatory stress. Nutrients. 2019;11:2062. doi:10.3390/nu11092062
48. Choi Y, Kwon HK, Park S. Polygenic Variants Linked to oxidative stress and the antioxidant system are associated with type 2 diabetes risk and interact with lifestyle factors. Antioxidants. 2023;12:1280. doi:10.3390/antiox12061280
49. Targher G, Corey KE, Byrne CD, Roden M. The complex link between NAFLD and type 2 diabetes mellitus - mechanisms and treatments. Nat Rev Gastroenterol Hepatol. 2021;18:599–612. doi:10.1038/s41575-021-00448-y
50. Tan YQ, Wang YN, Feng HY, et al. Host/microbiota interactions-derived tryptophan metabolites modulate oxidative stress and inflammation via aryl hydrocarbon receptor signaling. Free Radic Biol Med. 2022;184:30–41. doi:10.1016/j.freeradbiomed.2022.03.025
51. Li M, Han X, Sun L, Liu X, Zhang W, Hao J. Indole-3-acetic acid alleviates DSS-induced colitis by promoting the production of R-equol from Bifidobacterium pseudolongum. Gut Microbes. 2024;16:2329147. doi:10.1080/19490976.2024.2329147
52. Yang W, Ren D, Shao H, et al. Theabrownin from fu brick tea improves ulcerative colitis by shaping the gut microbiota and modulating the tryptophan metabolism. J Agric Food Chem. 2023;71:2898–2913. doi:10.1021/acs.jafc.2c06821
53. Kiernan K, MacIver NJ. A novel mechanism for Th17 Inflammation in human type 2 diabetes mellitus. Trends in Endocrinology and Metabolism: TEM. 2020;31:1–2. doi:10.1016/j.tem.2019.11.002
54. Menni C, Hernandez MM, Vital M, Mohney RP, Spector TD, Valdes AM. Circulating levels of the anti-oxidant indoleproprionic acid are associated with higher gut microbiome diversity. Gut Microbes. 2019;10:688–695. doi:10.1080/19490976.2019.1586038
55. Zhao ZH, Xin FZ, Xue Y, et al. Indole-3-propionic acid inhibits gut dysbiosis and endotoxin leakage to attenuate steatohepatitis in rats. Exp Mol Med. 2019;51:1–14. doi:10.1038/s12276-019-0304-5
56. Tuomainen M, Lindström J, Lehtonen M, et al. Associations of serum indolepropionic acid, a gut microbiota metabolite, with type 2 diabetes and low-grade inflammation in high-risk individuals. Nutr Diabetes. 2018;8:35. doi:10.1038/s41387-018-0046-9
57. de Mello VD, Paananen J, Lindström J, et al. Indolepropionic acid and novel lipid metabolites are associated with a lower risk of type 2 diabetes in the Finnish diabetes prevention study. Sci Rep. 2017;7:46337. doi:10.1038/srep46337
58. Abildgaard A, Elfving B, Hokland M, Wegener G, Lund S. The microbial metabolite indole-3-propionic acid improves glucose metabolism in rats, but does not affect behaviour. Arch Physiol Biochem. 2018;124:306–312. doi:10.1080/13813455.2017.1398262
59. Jennis M, Cavanaugh CR, Leo GC, Mabus JR, Lenhard J, Hornby PJ. Microbiota-derived tryptophan indoles increase after gastric bypass surgery and reduce intestinal permeability in vitro and in vivo. Neurogastroenterol Motil. 2018;30. doi:10.1111/nmo.13178
60. Yu M, Wu S, Gong C, Chen L. Neuregulin-1β increases glucose uptake and promotes GLUT4 translocation in palmitate-treated C2C12 myotubes by activating PI3K/AKT signaling pathway. Front Pharmacol. 2022;13:1066279. doi:10.3389/fphar.2022.1066279
61. Siqueira F, Rodrigues FAP, Ribeiro SA, et al. Induced acute hyperglycemia modifies the barrier function of the intestinal epithelium by tissue inflammation and tight junction disruption resulting in hydroelectrolytic secretion in an animal model. Braz J Med Biol Res. 2024;57:e13309. doi:10.1590/1414-431x2024e13309
62. Nascimento JC, Matheus VA, Oliveira RB, Tada SFS, Collares-Buzato CB. High-fat diet induces disruption of the tight junction-mediated paracellular barrier in the proximal small intestine before the onset of type 2 diabetes and endotoxemia. Dig Dis Sci. 2021;66:3359–3374. doi:10.1007/s10620-020-06664-x
63. Kim Y, Lee Y, Heo G, et al, Modulation of intestinal epithelial permeability via protease-activated receptor-2-Induced Autophagy . Cells. 2022;11 (5):
64. Ismael S, Rodrigues C, Santos GM, et al. IPA and its precursors differently modulate the proliferation, differentiation, and integrity of intestinal epithelial cells. Nutr Res Pract. 2023;17:616–630. doi:10.4162/nrp.2023.17.4.616
65. Hinnen D. Glucagon-Like peptide 1 receptor agonists for type 2 diabetes. Diabetes Spectr. 2017;30:202–210. doi:10.2337/ds16-0026
66. Fernández-Millán E, Martín MA, Goya L, et al. Glucagon-like peptide-1 improves beta-cell antioxidant capacity via extracellular regulated kinases pathway and Nrf2 translocation. Free Radic Biol Med. 2016;95:16–26. doi:10.1016/j.freeradbiomed.2016.03.002
67. Himpe E, Cunha DA, Song I, et al. Phenylpropenoic acid glucoside from rooibos protects pancreatic beta cells against cell death induced by acute injury. PLoS One. 2016;11:e0157604. doi:10.1371/journal.pone.0157604
68. Lutz TA, Meyer U. Amylin at the interface between metabolic and neurodegenerative disorders. Front Neurosci. 2015;9:216. doi:10.3389/fnins.2015.00216
69. Hernandez LF, Eguchi N, Whaley D, Alexander M, Tantisattamo E, Ichii H. Anti-oxidative therapy in diabetic nephropathy. Front Biosci. 2022;14:14. doi:10.31083/j.fbs1402014
70. Samsu N. Diabetic nephropathy: challenges in pathogenesis, diagnosis, and treatment. Biomed Res Int. 2021;2021:1497449. doi:10.1155/2021/1497449
71. Stępniak J, Krawczyk-Lipiec J, Lewiński A, Karbownik-Lewińska M. Sorafenib versus lenvatinib causes stronger oxidative damage to membrane lipids in noncancerous tissues of the thyroid, liver, and kidney: effective protection by melatonin and indole-3-propionic acid. Biomedicines. 2022;10:2890. doi:10.3390/biomedicines10112890
72. Zhuang H, Li B, Xie T, et al. Indole-3-aldehyde alleviates chondrocytes inflammation through the AhR-NF-κB signalling pathway. Int Immunopharmacol. 2022;113:109314. doi:10.1016/j.intimp.2022.109314
73. Renga G, Nunzi E, Pariano M, et al. Optimizing therapeutic outcomes of immune checkpoint blockade by a microbial tryptophan metabolite. J Immunother Cancer. 2022;10:e003725. doi:10.1136/jitc-2021-003725
74. Zhang J, Zhang R, Chen Y, et al. Indole-3-aldehyde alleviates high-fat diet-induced gut barrier disruption by increasing intestinal stem cell expansion. J Agric Food Chem. 2024;72:18930–18941. doi:10.1021/acs.jafc.4c02381
75. Dodd D, Spitzer MH, Van Treuren W, et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature. 2017;551:648–652. doi:10.1038/nature24661
76. Langan D, Perkins DJ, Vogel SN, Moudgil KD. Microbiota-derived metabolites, indole-3-aldehyde and indole-3-acetic acid, differentially modulate innate cytokines and stromal remodeling processes associated with autoimmune arthritis. Int J Mol Sci. 2021;22:2017. doi:10.3390/ijms22042017
77. Alexeev EE, Lanis JM, Kao DJ, et al. Microbiota-derived indole metabolites promote human and murine intestinal homeostasis through regulation of interleukin-10 receptor. Am J Pathol. 2018;188:1183–1194. doi:10.1016/j.ajpath.2018.01.011
78. Inubushi T, Kamemura N, Oda M, et al. L-tryptophan suppresses rise in blood glucose and preserves insulin secretion in type-2 diabetes mellitus rats. J Nutr Sci Vitaminol. 2012;58:415–422. doi:10.3177/jnsv.58.415
79. Waclawiková B, Bullock A, Schwalbe M, et al. Gut bacteria-derived 5-hydroxyindole is a potent stimulant of intestinal motility via its action on L-type calcium channels. PLoS Biol. 2021;19:e3001070. doi:10.1371/journal.pbio.3001070
80. Liu W, Wang J, Yang H, et al. The metabolite indole-3-acetic acid of bacteroides ovatus improves atherosclerosis by restoring the polarisation balance of m1/m2 macrophages and inhibiting inflammation. Adv Sci. 2025;12:e2413010. doi:10.1002/advs.202413010
81. Rebnord EW, Strand E, Midttun Ø, et al. The kynurenine:tryptophan ratio as a predictor of incident type 2 diabetes mellitus in individuals with coronary artery disease. Diabetologia. 2017;60:1712–1721. doi:10.1007/s00125-017-4329-9
82. Filippas-Ntekouan S, Dimou A, Dafopoulos P, et al. Effect of dapagliflozin on the serum metabolome in patients with type 2 diabetes mellitus. J Diabetes Metab Disord. 2025;24:4. doi:10.1007/s40200-024-01508-1
83. Miao H, Wang YN, Yu XY, et al. Lactobacillus species ameliorate membranous nephropathy through inhibiting the aryl hydrocarbon receptor pathway via tryptophan-produced indole metabolites. Br J Pharmacol. 2024;181:162–179. doi:10.1111/bph.16219
84. Miao H, Liu F, Wang YN, et al. Targeting Lactobacillus johnsonii to reverse chronic kidney disease. Signal Transduct Target Ther. 2024;9:195. doi:10.1038/s41392-024-01913-1
85. Choi KM, Yoo HS. Amelioration of hyperglycemia-induced nephropathy by 3,3’-diindolylmethane in diabetic mice. Molecules. 2019;24:4474. doi:10.3390/molecules24244474
86. Wang J, Lu S, Sheng R, Fan J, Wu W, Guo R. Structure-activity relationships of natural and synthetic indole-derived scaffolds as α-glucosidase inhibitors: a mini-review. Mini Rev Med Chem. 2020;20:1791–1818. doi:10.2174/1389557520666200619121003
87. Weems JM, Cutler NS, Moore C, et al. 3-Methylindole is mutagenic and a possible pulmonary carcinogen. Toxicol Sci. 2009;112:59–67. doi:10.1093/toxsci/kfp201
88. Lekawanvijit S. Role of gut-derived protein-bound uremic toxins in cardiorenal syndrome and potential treatment modalities. Circ J. 2015;79:2088–2097. doi:10.1253/circj.CJ-15-0749
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.