Back to Journals » OncoTargets and Therapy » Volume 11
TRIM28 is overexpressed in glioma and associated with tumor progression
Received 19 March 2018
Accepted for publication 17 July 2018
Published 10 October 2018 Volume 2018:11 Pages 6447—6458
DOI https://doi.org/10.2147/OTT.S168630
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Faris Farassati
Chunhai Su,1,* Hui Li,2,* Wenbo Gao3
1Department of Neurosurgery, Jining No 1 People’s Hospital, Jining, China; 2School of Nursing, Jining Medical University, Jining, China; 3Department of Neurosurgery, Binzhou Medical University Hospital, Binzhou, China
*These authors contributed equally to this work
Background: Tripartite motif containing 28 (TRIM28) is a transcriptional co-factor targeting many genes with pleiotropic biological activities, but the study on the role of TRIM28 in glioma is rare.
Methods: To explore the function of TRIM28 in glioma, we first detected the expression levels of TRIM28 in glioma tissues and analyzed the correlations of TRIM28 expression with clinicopathological variables of patients in 85 cases of glioma. Meanwhile, we used shRNA to knockdown TRIM28 in glioma cell lines to detect the biological functions of TRIM28 in cell and animal experiments.
Results: We found that TRIM28 was expressed at significantly higher level in glioma tissues than in non-tumor brain, and TRIM28 expression correlated significantly with tumor malignancy. Furthermore, TRIM28 higher expression was also correlated with poor survival of glioma patients (P<0.01). Functionally, knockdown of TRIM28 could significantly inhibit cell proliferation and migration in glioma cells. Additionally, we found that TRIM28 could inhibit the expression of E-cadherin significantly by reducing its mRNA stability at the post-transcriptional level.
Conclusion: Our results suggest that TRIM28 overexpression is correlated with glioma malignant progression and patients’ poor survival, so targeting TRIM28 could be an efficacious strategy in glioma.
Keywords: TRIM28, glioma, progression, E-cadherin
Introduction
Malignant glioma is the most commonly occurring highly malignant primary brain tumor, with rapid proliferation and strong tendency for invasion and metastasis.1,2 Although the advanced diagnostic techniques and treatment for glioma have been applied over the past few decades, 3-year postoperative survival rate is still very low, and targeted therapy for improving glioma patients’ survival has not been well established.3 Therefore, exploiting the in-depth molecular mechanism and finding novel treatment strategies are crucial for glioma therapy.
Tripartite motif containing 28 (TRIM28) serves as a transcriptional co-factor by interacting with KRAB-containing zinc finger proteins (KRAB-ZFP).4,5 Meanwhile, more and more researchers find that TRIM28 has pleiotropic biological activities, and could regulate its target genes at both transcriptional and posttranscriptional levels.6 On the one hand, TRIM28 could affect the epigenetic modulation of chromatin structure, or the transcriptional activity of KRAB-ZFP-specific loci, and then regulate many genes expression.4 On the other hand, TRIM28 could serve as a small ubiquitin-related modifier (SUMO)/ubiquitin E3 ligase and regulate cell apoptosis.7 To date, many studies have indicated that TRIM28 plays an important role in promoting proliferation and migration of tumor cells, and higher levels of TRIM28 expression have been observed in breast, pancreatic, liver, gastric, lung, ovarian, and prostate cancers.7–19 Despite many efforts to elucidate the cellular functions and associated molecular mechanisms of TRIM28, the studies of relationship between TRIM28 and glioma are rare. To date, just two studies have detected TRIM28 expression in glioma, and showed a positive correlation between TRIM28 expression and glioma malignancy;20,21 however, these studies did not explore the molecular mechanism of TRIM28 in glioma. In this study, we investigated the expression level of TRIM28 in glioma tissues, and analyzed the correlation of TRIM28 expression with clinicopathological variables and survival of glioma patients. Meanwhile, in vivo and in vitro experiments were performed to detect the effect of TRIM28 on cell biological behavior of glioma cell lines. To explore the mechanism of TRIM28 in glioma, we also detected its effect on E-cadherin expression at both protein and mRNA levels.
Materials and methods
Antibodies and drugs
The primary antibodies for both Western blot and immunohistochemistry staining were listed as follows: rabbit anti-TRIM28 (Abcam, Cambridge, MA, USA), rabbit anti-E-cadherin (Abcam), rabbit anti-caspase3 (Abcam), rabbit anti-Ki67 (Abcam), and mouse anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Santa Cruz Biotechnology, Santa Cruz, CA, USA). The anti-rabbit secondary antibody (Zhongshan Goldbridge Biotechnology, Beijing, China) and anti-mouse secondary antibody (Santa Cruz Biotechnology) were purchased for Western blot. The secondary antibodies PV6001 was obtained from Zhongshan Goldbridge Biotechnology for immunohistochemistry staining.
Cycloheximide (CHX) and Actinomycin D (ActD) were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Patient samples
Eighty-five glioma samples were obtained from the Department of Pathology, Jining No 1 People’s Hospital, from 2007 to 2011. Final diagnosis of all tissue sections were confirmed by histopathological specialists, according to the World Health Organization criteria. All patients’ clinicopathological data collected are summarized in Table 1. This study complied with the Declaration of Helsinki and was approved by the Human Ethics and Research Ethics Committees of the hospital. Written informed consent was obtained from all patients.
 | Table 1 Correlations between TRIM28 expression and clinicopathological variables |
Immunohistochemistry staining and evaluation
Paraffin-embedded tissue sections were first deparaffinized with xylene and rehydrated with graded alcohol solutions. Then, 3% hydrogen peroxide was used to quench endogenous peroxidase, and tissues were boiled in 10 mM citrate buffer (pH 6.0) for 3 minutes to expose antigens. After cooling to room temperature, sections were incubated with primary antibodies for 18 hours at 4°C, and incubated with PV6001 for 30 minutes at 37°C, and then stained with DAB (Zhongshan Goldbridge Biotechnology, Beijing, China) for 1–2 minutes. Control sections were incubated with PBS instead of a primary antibody, and the washing buffer was PBS.
For scoring the stain of tissue sections, five high-power fields were chosen from each slice, and the mean percentage of stained cells was estimated. For TRIM28, the level ≤50% in tissues was defined as the lower expression group, and >50% as the higher expression group (Figure 1A).
 | Figure 1 TRIM28 expression in glioma and its associations with tumor progression. |
Cell culture and treatment
Human glioma cell lines (LN229 and U87) were purchased from the American Type Culture Collection (Rockville, MD, USA), and all cells were cultured in DMEM medium containing 10% of fetal bovine serum (Gibco; Invitrogen Life Technologies, Carlsbad, CA, USA), 100 U/mL of penicillin, and 100 μg/mL of streptomycin (Invitrogen). The cells were cultured at 37°C in a humidified incubator with 5% CO2. The dose 1 μM of actinomycin D (ActD) was used to detect the stability of E-cadherin mRNA, and 100 μM CHX was used to treat the cells.
Plasmid construction and cell infection
Small hairpin RNAs (shRNA) were constructed into pLKO.1 plasmids. The shRNA sequences were shTRIM28#1: CCTGGCTCTGTTCTCTGTCCT and shTRIM28#2: CTGAGACCAAACCTGTGCTTA, and synthesized by Shanghai Genechem Co., Ltd. (Shanghai, China). Full-length human TRIM28 cDNA was obtained by polymerase chain reaction (PCR) and was cloned into pCDH-puro-vector. The day before transfection, 1×105 cells were placed in 35 mm dishes in DMEM supplemented with 10% fetal bovine serum and without antibiotics. The transfected cells were cultured for 48 hours, and Western blot was utilized to detect TRIM28 expression protein level.
Western blot analysis
All agents were purchased from Santa Cruz Biotechnology. Cell lysates were prepared with cell lysis buffer (20 mmol Tris-HCl, pH 7.5, 150 mmol NaCl, 1% Triton, 2.5 mmol sodium pyrophosphate; 1 mmol/L b-glycerophosphate; 1 mmol/L Na3VO4; 1 mg/mL leupeptin; 1 mmol/L phenylmethylsulfonylfluoride). The protein concentration was measured by the Bradford method, and proteins were resolved on a denaturing 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis gel, subsequently transferred to polyvinylidene fluoride membranes via semidry transfer. The membrane was then blocked in 5% dried milk in Tris-buffered saline and Tween 20, incubated with primary antibodies for overnight at 4°C, and then with secondary antibodies for 1 hour at 37°C. GAPDH protein was used as a loading control.
RNA extraction and PCR
Total cellular RNA was extracted by using RNeasy® mini kit (Qiagen, Duesseldorf, Germany) according to the manufacturer’s instructions. RNA was quantified using the NanoDrop 1000 (Thermo Fisher Scientific, Waltham, MA, USA) and quality was assessed by gel electrophoresis. cDNA was synthesized using a Quantitect Reverse Transcription kit (Qiagen) according to the manufacturer’s instructions. The cDNA were used as templates for PCR, and the primers are as follows: TRIM28: forward 5′-GATCATGAAGGAGCTGAACAAGCG-3′ and reverse 5′-TGGATCTTAGTCATGGTCCAGTGC-3′; E-cadherin: forward 5′-GTCACTGACACCAACGATAATCCT-3′ and reverse 5′-TTTCAGTGTGGTGATTACGACGTTA-3′; and GAPDH: forward 5′-AAGGTGAAGGTCGGAGTCAA-3′ and reverse 5′-AATGAAGGGGTCATTGATGG-3′.
MTT assay for cell viability and proliferation
Cell viability and proliferation were analyzed via MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide, Sigma, St Louis, MO, USA) assay using a Roche Cell Proliferation Kit I according to its protocol. Untransfected or transfected cells were seeded into 96-well plates at a density of 3,000 cells/well, then incubated for 0, 24, 48, 72 and 96 hours. At the end of incubation, 20 μL of 5 mg/mL MTT were added to each well for 4 hours, following which 150 μL dimethyl sulfoxide was added. The absorbance was measured at 490 nm wavelength on a spectrophotometer.
Clonogenic survival assay
A total of 500 cells were seeded into 6-well plates and incubated for 14 days. Then the cells were fixed with methanol for 6 minutes at −20°C and stained with gentian violet. Counting the clones under the microscope, and colonies containing more than 50 cells were scored as surviving cells. The colony number and colony formation fraction were detected.
Soft agar colony formation assay
A total of 1×104 cells were plated in 0.4% agarose on top of a 1% agarose base supplemented with 2× complete medium in 6-well plates. Then cells were allowed to grow at 37°C for 4 weeks. Meanwhile, on day 0 and on day 14 of growth, 1 mL of 1× media without agarose was added again on top of the growth layer to prevent the agarose dry. The pictures were taken by microscope, and the number of colonies was counted by Quantity One software.
Wound-healing assay
Dishes measuring 3.5 cm were seeded with about 5×105 cells and grown until they reached 90% confluence. Cell monolayer was scratched with a 20 μL pipette tip, and washed with PBS 3 times. Then the incubation was continued in serum-free culture medium for 24 hours. The distances of cell migration were detected by microscopy.
Mice xenograft models
A total of 2×106 LN229 of shctrl and shTRIM28 cells were injected subcutaneously to the right of the dorsal midline in nu/nu mice to generate murine subcutaneous tumors. When the subcutaneous tumor size had reached the size of approximately 60–80 mm3, tumor volumes were calculated by the following formula: length×width×width/2, and the survival days were accounted. All animal studies were approved by the Jining Medical University in accordance with the principles and procedures outlined in the NIH Guide for the Care and Use of Laboratory Animals.
Statistical analysis
SPSS 16.0 was used to statistic the data. The chi-squared test was used to value the correlation of TRIM28 expression and clinical features. Kaplan–Meier analysis was used to analyze the patient univariate survival, and multivariate survival analysis of prognostic factors was analyzed by Cox’s proportional hazard regression model. Other data were analyzed by 2-tailed Student’s t-test or ANOVA least significant difference test. The significance level was defined as P<0.05. All experiments were performed in triplicate.
Results
TRIM28 is overexpressed in glioma and is associated with tumor progression
To investigate the role of TRIM28 in glioma, we first analyzed its mRNA expression level using The Cancer Genome Atlas database, and we found that TRIM28 mRNA was extensively expressed in glioma tissues than non-tumor brain (Figure 1A). Meanwhile, real-time PCR and Western blot were used to detect TRIM28 mRNA and protein expression levels in 5 fresh specimens of glioma tissues and adjacent normal tissues, and the results showed that both TRIM28 mRNA level (Figure 1B) and protein level (Figure 1C) were higher in glioma tissues. To further compare the expression level of TRIM28 in glioma specimens and adjacent non-tumor brain, 85 cases of human glioma were collected and immunohistochemistry was used to detect TRIM28 expression level. The results showed that TRIM28 was mainly localized to the cell nucleus, and the expression level of TRIM28 was significantly higher in glioma tissues (Figure 1D, P<0.01). Furthermore, the level of TRIM28 expression was also significantly higher in high-grade glioma (Figure 1E).
The associations of TRIM28 expression levels with clinicopathological variables of glioma, including age, sex, tumor diameter, KPS score, and survival status were also analyzed. For statistical analysis, the sections were scored as lower expression (positive stain cells ≤50%) and higher expression (positive stain cells >50%), and the results are summarized in Table 1. The results demonstrated that higher TRIM28 expression was significantly associated with tumor diameter (Chi-squared =4.06, P=0.04), KPS score (Chi-squared =11.10, P<0.01) and survival status (Chi-squared =26.25, P<0.01). The results suggested TRIM28 expression was associated with aggressive clinical features of glioma.
We have showed that TRIM28 was correlated with malignant progression of glioma, and cell proliferation is considered to play an important role in aggressive process. Ki67 always has been used as a proliferation marker in many cancer cells, including glioma. So, in this study, we investigated the Ki67 expression level in glioma and analyzed the correlation of TRIM28 and Ki67. We found that there was a significant positive correlation between TRIM28 and Ki67 expression, and the glioma tissues with TRIM28 higher expression always had Ki67 overexpression (Figure 1F).
TRIM28 expression is associated with poor survival of glioma patients
First, Kaplan–Meier survival analysis and COX analysis were carried out in these 85 cases to investigate the relationship between TRIM28 expression and patients’ survival. The results of Kaplan–Meier survival analysis showed that glioma patients with higher TRIM28 expression in tumor tissues not only always had poor overall survival (P<0.01, Figure 2A), but also had poor progression-free survival (P<0.01, Figure 2B). In addition, TRIM28 could be as a predictor of survival in multivariate analysis (hazard ratio =5.01, 95% CI=1.10–22.79, P=0.037), when entered into a model containing all clinicopathologic variables for COX analysis (Table 2). Furthermore, Kaplan–Meier analysis was also performed using the R2 genomics database (available online: http://hgserver1.amc.nl/cgi-bin/r2/main.cgi) to detect the effect of TRIM28 mRNA on glioma patients’ survival. As expected, the higher expression level of TRIM28 mRNA was also associated with the poor survival of patients (P<0.01, Figure 2C and D). All these results suggested that higher TRIM28 levels predicted a worse prognosis and TRIM28 could serve as a candidate biomarker for glioma.
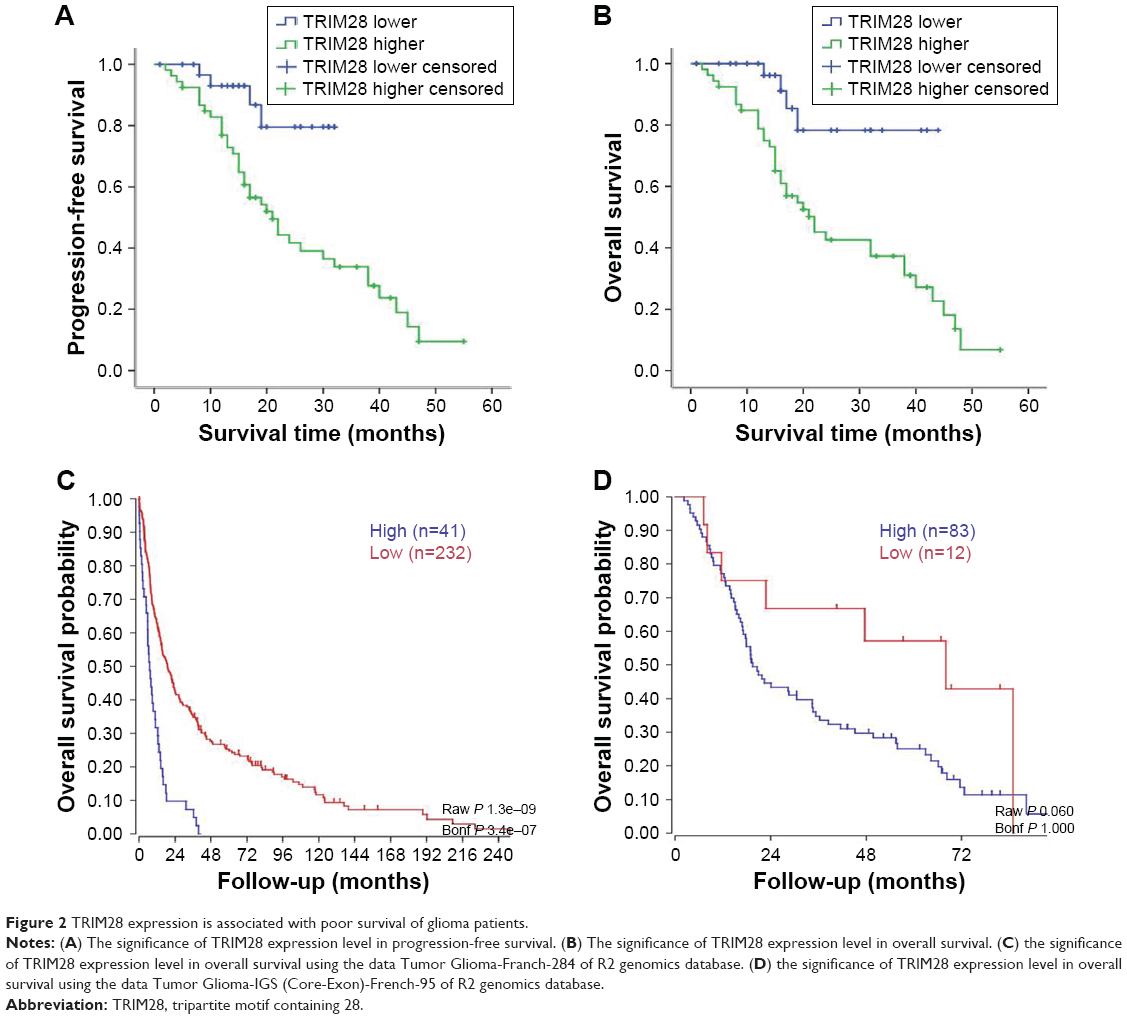 | Figure 2 TRIM28 expression is associated with poor survival of glioma patients. |
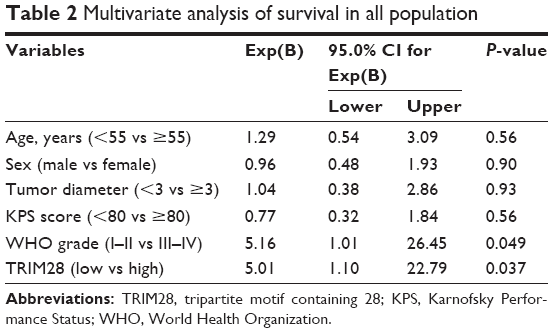 | Table 2 Multivariate analysis of survival in all population |
The effects of TRIM28 knockdown in tumor cell ability of proliferation and migration
Although it is clear that TRIM28 is overexpressed in glioma tissues, and TRIM28 is associated with tumor malignant progression and patient’s poor survival, the underlying mechanism is still not very clear. In this study, gene silencing of TRIM28 through lentiviral transduction system and shRNA was established in glioma cell lines LN229 and U87. Western blot analysis was first performed to detect the silencing efficiency of two distinct shRNAs targeting TRIM28 in LN229; the results found that TRIM28 was down regulated (Figure 3A). Meanwhile, PCR was performed to investigate the silencing efficiency of shTRIM28 in TRIM28 mRNA level. We found that the levels of TRIM28 mRNA were decreased after lentiviral transduction system was transducted into LN229 (Figure 3B). To investigate the role of TRIM28 in cell biology function, cell proliferation analysis, colony formation, and wound-healing analysis were performed in shTRIM28 and shctrl cells. The results showed that knockdown of TRIM28 could significantly affect the cell abilities of proliferation (Figure 3C), colony formation (Figure 3D and E) and migration (Figure 3F) in LN229. In addition, the same results were also found in U87 cell line; silencing TRIM28 (Figure 4A and B) not only repressed cell proliferation (Figure 4C) but also decreased the number of both two-dimensional colony formation (Figure 4D) and soft agar suspension colony formation (Figure 4E). Silencing TRIM28 could repress cell migration in MG63 cell line (Figure 4F). All these results suggested that higher TRIM28 levels promoted glioma cell malignancy in vitro.
 | Figure 3 TRIM28 silencing significantly affects cell proliferation and migration in LN229 cell line. |
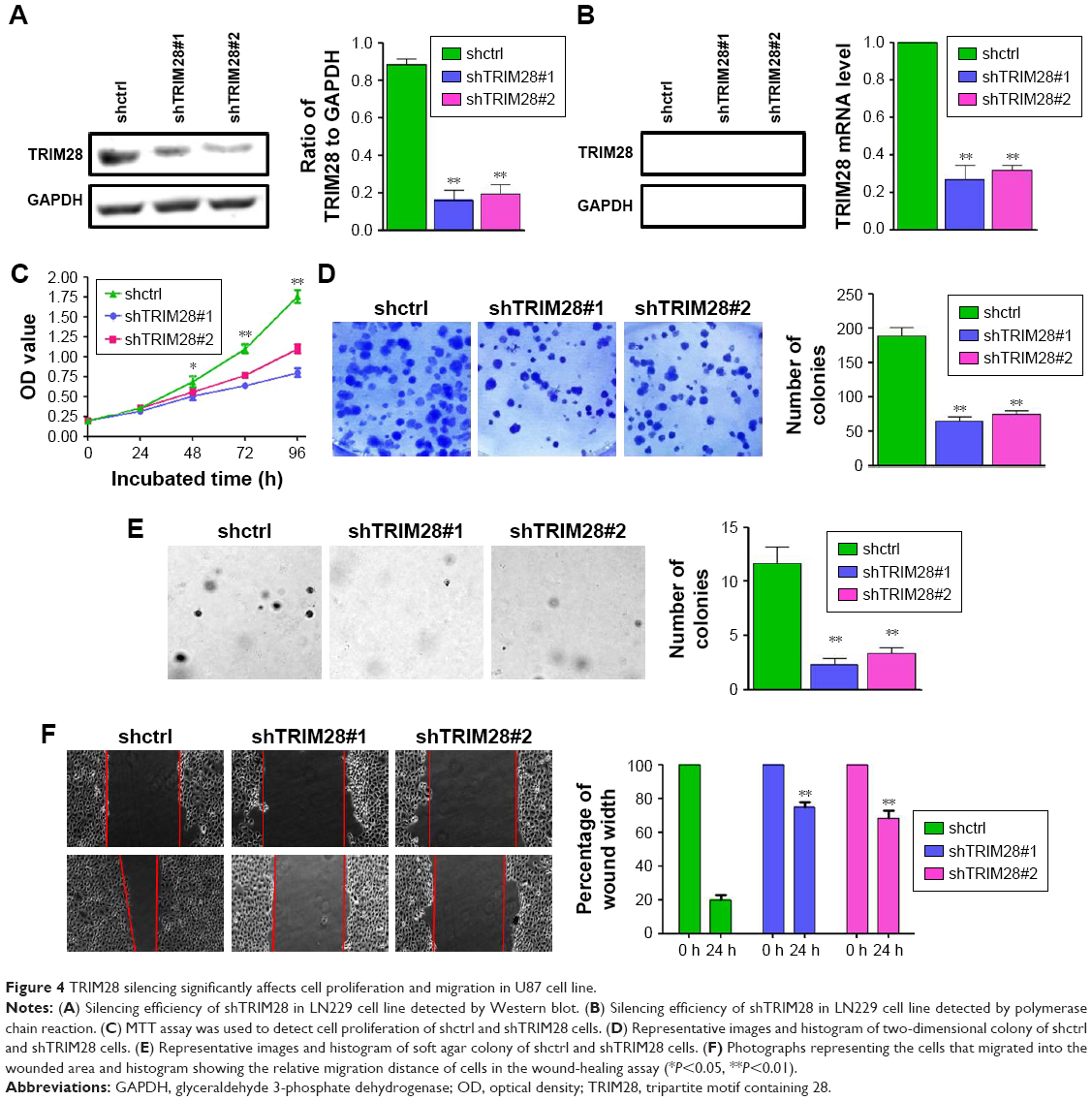 | Figure 4 TRIM28 silencing significantly affects cell proliferation and migration in U87 cell line. |
The effect of TRIM28 in glioma cells in vivo
To confirm the role of TRIM28, in vivo animal experiment was also performed, and shctrl and shTRIM28 LN229 cells were injected subcutaneously to dorsal midline in nu/nu mice to generate murine subcutaneous tumors. Then tumor volumes were measured, and the survival days of mice with tumor were accounted. We found that there was a marked difference in the tumor growth rate between the mice with shctrl and shTRIM28 cells, and the tumor growth rate (mean tumor volume) was much slower in shTRIM28 tumors (Figure 5B). The smallest tumor in the shctrl group was still bigger than the biggest one in the shTRIM28 group (Figure 5A). Meanwhile, the survival rate of shctrl mice decreased to 20% on day 30 after the injection of tumor cells, but the survival rate of shTRIM28 only decreased to 80% (Figure 5C). In addition, compared with the shctrl group, the area ratio of necrosis in LN229 xenografts was insignificantly increased in the shTRIM28 group (Figure 5D). The levels of caspase3 expression in tumor tissues of the shTRIM28 group were higher than that of the shctrl group (Figure 5E), which suggested that TRIM28 could inhibit cell apoptosis in vivo. As higher TRIM28 expression was significantly associated with tumor distant metastasis in clinical glioma, we investigated the relationship between TRIM28 expression and distant metastasis in mice. The results showed that the number of metastasis focus in both the lung and the liver was significantly increased in the shTRIM28 group (Figure 5F). Meanwhile, the expression levels of E-cadherin in the shTRIM28 group were significantly increased than that of E-cadherin in the shctrl group (Figure 5G).
 | Figure 5 The effect of TRIM28 in vivo of animal experiment for glioma cells. |
The effect of TRIM28 in the expression of E-cadherin
We have found that there was a close relationship between TRIM28 and E-cadherin expression in LN229 xenografts, and some studies have reported that TRIM28 contributed to the regulation of E-cadherin in breast cancer and lung cancer cells.22,23 To investigate if there is any relationship between TRIM28 and E-cadherin expression in glioma cell line in vitro, the expression level of E-cadherin was detected by Western blot and PCR in shTRIM28 and shctrl cells. Both the protein (Figure 6A) and mRNA (Figure 6B) levels of E-cadherin were increased in shTRIM28 cells. To explore the main mechanism of TRIM28 regulating E-cadherin, a transcription blocker, 1 μM ActD was used to detect the stability of E-cadherin mRNA. At different time points after ActD treatment, the cells were collected and analyzed for E-cadherin mRNA expression. We observed that the E-cadherin mRNA expression was significantly increased after treatment with ActD in shTRIM28 cells, compared with shctrl cells (Figure 6C). Our data suggested that TRIM28 regulated E-cadherin at the posttranscriptional level by affecting its mRNA stability. To further confirm the effect of E-cadherin in regulating E-cadherin expression, TRIM28 was overexpressed in LN229 using lentiviral transduction system. We found that the expression level of E-cadherin was significantly decreased in both protein and mRNA levels in TRIM28 overexpression cells (Figure 6D and E). Using the same method, we detected E-cadherin mRNA level in control and TRIM28 overexpression cells after treatment with ActD, and the results showed that E-cadherin mRNA expression was significantly decreased in cells overexpressing TRIM28 (Figure 6F). To exclude the effect of protein degradation in E-cadherin expression, 100 μM CHX was used to treat control and TRIM28 overexpression cells. The results demonstrated that the expression levels of E-cadherin were decreased after treatment with CHX, but there was no significant difference in E-cadherin expression between control and TRIM28 overexpression cells (Figure 6G), which suggested that the role of TRIM28 in regulating E-cadherin was not through the protein degradation system. Meanwhile, we found that there was a significantly negative correlation between TRIM28 and E-cadherin expression in glioma tissues, and the glioma tissues with TRIM28 higher expression always had lower E-cadherin expression (Figure 6H).
 | Figure 6 The effect of TRIM28 on the expression of E-cadherin. |
Discussion
In this study, we found that TRIM28 was overexpressed in glioma tissues, and its expression was correlated with malignant progression of glioma and poor survival of glioma patients. Silencing TRIM28 inhibited cell growth and migration ability in glioma cells. Meanwhile, TRIM28 played a crucial role in regulating E-cadherin expression in both cell experiment in vitro and animal experiment in vivo, and there was a significantly negative correlation between TRIM28 and E-cadherin expression in glioma tissues.
As we all know, glioma is the most lethal and aggressive type of brain tumor in adults.24 Extensive invasion and vigorous proliferation always make glioma extremely difficult to treat.25,26 Although the advanced diagnostic techniques and treatment for glioma have been applied over the past few decades, 3-year postoperative survival rate is still very low, because of its rapid proliferation and strong tendency for invasion and metastasis.27 Therefore, it is urgent to find new biomarkers that are able to predict the prognosis of glioma, exploit the in-depth molecular mechanism, and find novel treatment strategies that are crucial for glioma therapy.
Many studies have investigated the correlation between TRIM28 and tumor malignance and progression. Several studies demonstrated that TRIM28 overexpression was always associated with clinicopathological features, which gives prominence to the importance of TRIM28 in tumor progression. High levels of TRIM28 correlated with a significantly lower survival rate in patients with ovarian, gastric, and pancreatic cancers.9,15,28,29 Recently, it has been reported that TRIM28 could promote proliferation and metastatic progression in breast cancer cells.11 Meanwhile, TRIM28 was involved in EMT in lung cancer.19 Two studies have showed a positive correlation between TRIM28 expression and glioma malignancy,20,21 which suggested the possibility of TRIM28 as a treatment target for human glioma; however, these studies did not explore the molecular mechanism of TRIM28 in glioma. In this study, we found that TRIM28 was overexpressed in glioma tissues, and higher expression of TRIM28 led to a worse prognosis via promoting the cell proliferation ability and migration of glioma cells both in vitro and in vivo, which was agreement with previous studies.
Metastasis is one of the major obstacles for therapy of glioma, but little is known about the mechanism that mediates glioma metastasis. Loss of E-cadherin is thought to promote metastasis by disrupting intercellular contacts, and the significance of E-cadherin for metastasis has been shown in a variety of tumors.30,31 The correlation of TRIM28 and E-cadherin has been valued in some papers. TRIM28 decreased the expression level of E-cadherin and correlated with increased metastatic ability in pancreatic cancer.12 Chen et al found that TRIM28 could suppress the expression of E-cadherin in lung cancer via altering histone 3 modification of its promoter, which contributed to epithelial-mesenchymal transition (EMT) and lung cancer metastasis.21 In addition, TRIM28 promoted breast cancer metastasis by stabilizing TWIST1 protein, which indirectly affected E-cadherin expression.20
In this study, we also found that there was a significantly negative correlation between TRIM28 and E-cadherin expression in glioma tissues, and the glioma tissues with TRIM28 higher expression always had E-cadherin lower expression. Furthermore, we detected the level of E-cadherin in silencing TRIM28 cells and overexpression TRIM28 cells to explore the specific mechanism of TRIM28 in regulating E-cadherin. We found that TRIM28 could suppress the expression of E-cadherin, and the role of TRIM28 in regulating E-cadherin was not through affecting protein expression, but at the posttranscriptional level by reducing its mRNA stability. However, the identified molecular mechanism underlying the TRIM28 controlling E-cadherin mRNA stability needs further research.
Conclusion
In summary, we found that TRIM28 was overexpressed in glioma tissues, and its expression correlated with malignant progression of glioma and poor survival of glioma patients. Meanwhile, this study reveals a mechanism for transcriptional inactivation of E-cadherin by TRIM28, and this pathway may be a therapeutic target for glioma.
Acknowledgments
We appreciate all volunteers participating in this study sincerely. We also sincerely thank the Jining Medical University for their assistance in experiments.
Disclosure
The authors report no conflicts of interest in this work.
References
Qi XT, Zhan JS, Xiao LM, et al. The Unwanted Cell Migration in the Brain: Glioma Metastasis. Neurochem Res. 2017;42(6):1847–1863. | ||
Iwadate Y. Plasticity in Glioma Stem Cell Phenotype and Its Therapeutic Implication. Neurol Med Chir. 2018;58(2):61–70. | ||
Kline C, Felton E, Allen IE, Tahir P, Mueller S. Survival outcomes in pediatric recurrent high-grade glioma: results of a 20-year systematic review and meta-analysis. J Neurooncol. 2018;137(1):103–110. | ||
Bunch H, Calderwood SK. TRIM28 as a novel transcriptional elongation factor. BMC Mol Biol. 2015;16:14. | ||
Cheng CT, Kuo CY, Ann DK. KAPtain in charge of multiple missions: emerging roles of KAP1. World J Biol Chem. 2014;5(3):308–320. | ||
Iyengar S, Farnham PJ. KAP1 protein: an enigmatic master regulator of the genome. J Biol Chem. 2011;286(30):26267–26276. | ||
Neo SH, Itahana Y, Alagu J, et al. TRIM28 Is an E3 Ligase for ARF-Mediated NPM1/B23 SUMOylation That Represses Centrosome Amplification. Mol Cell Biol. 2015;35(16):2851–2863. | ||
van Tilborgh N, Spans L, Helsen C, et al. The transcription intermediary factor 1β coactivates the androgen receptor. J Endocrinol Invest. 2013;36(9):699–706. | ||
Cui Y, Yang S, Fu X, Feng J, Xu S, Ying G. High levels of KAP1 expression are associated with aggressive clinical features in ovarian cancer. Int J Mol Sci. 2014;16(1):363–377. | ||
Czerwińska P, Shah PK, Tomczak K, et al. TRIM28 multi-domain protein regulates cancer stem cell population in breast tumor development. Oncotarget. 2017;8(1):863–882. | ||
Addison JB, Koontz C, Fugett JH, et al. KAP1 promotes proliferation and metastatic progression of breast cancer cells. Cancer Res. 2015;75(2):344–355. | ||
Yu C, Zhan L, Jiang J, et al. KAP-1 is overexpressed and correlates with increased metastatic ability and tumorigenicity in pancreatic cancer. Med Oncol. 2014;31(7):25. | ||
Wei N, Shi Y, Truong LN, et al. Oxidative stress diverts tRNA synthetase to nucleus for protection against DNA damage. Mol Cell. 2014;56(2):323–332. | ||
Jin X, Pan Y, Wang L, et al. MAGE-TRIM28 complex promotes the Warburg effect and hepatocellular carcinoma progression by targeting FBP1 for degradation. Oncogenesis. 2017;6(4):e312. | ||
Yokoe T, Toiyama Y, Okugawa Y, et al. KAP1 is associated with peritoneal carcinomatosis in gastric cancer. Ann Surg Oncol. 2010;17(3):821–828. | ||
Wang YY, Li L, Zhao ZS, Wang HJ. Clinical utility of measuring expression levels of KAP1, TIMP1 and STC2 in peripheral blood of patients with gastric cancer. World J Surg Oncol. 2013;11:81. | ||
Herquel B, Ouararhni K, Khetchoumian K, et al. Transcription cofactors TRIM24, TRIM28, and TRIM33 associate to form regulatory complexes that suppress murine hepatocellular carcinoma. Proc Natl Acad Sci U S A. 2011;108(20):8212–8217. | ||
Kuo CY, Li X, Kong XQ, et al. An arginine-rich motif of ring finger protein 4 (RNF4) oversees the recruitment and degradation of the phosphorylated and SUMOylated Krüppel-associated box domain-associated protein 1 (KAP1)/TRIM28 protein during genotoxic stress. J Biol Chem. 2014;289(30):20757–20772. | ||
Liu L, Zhao E, Li C, et al. TRIM28, a new molecular marker predicting metastasis and survival in early-stage non-small cell lung cancer. Cancer Epidemiol. 2013;37(1):71–78. | ||
Wei C, Cheng J, Zhou B, et al. Tripartite motif containing 28 (TRIM28) promotes breast cancer metastasis by stabilizing TWIST1 protein. Sci Rep. 2016;6:29822. | ||
Chen L, Muñoz-Antonia T, Cress WD. Trim28 contributes to EMT via regulation of E-cadherin and N-cadherin in lung cancer cell lines. PLoS One. 2014;9(7):e101040. | ||
Ostrom QT, Gittleman H, Liao P, et al. CBTRUS Statistical Report: primary brain and other central nervous system tumors diagnosed in the United States in 2010–2014. Neuro Oncol. 2017;19(Suppl 5):v1–v88. | ||
Diaz RJ, Dios RR, Hattab EM, et al. Study of the biodistribution of fluorescein in glioma-infiltrated mouse brain and histopathological correlation of intraoperative findings in high-grade gliomas resected under fluorescein fluorescence guidance. J Neurosurg. 2015;122(6):1360–1369. | ||
Zetterling M, Roodakker KR, Berntsson SG, et al. Extension of diffuse low-grade gliomas beyond radiological borders as shown by the coregistration of histopathological and magnetic resonance imaging data. J Neurosurg. 2016;125(5):1155–1166. | ||
Westphal M, Lamszus K. The neurobiology of gliomas: from cell biology to the development of therapeutic approaches. Nat Rev Neurosci. 2011;12(9):495–508. | ||
Rauwel B, Jang SM, Cassano M, Kapopoulou A, Barde I, Trono D. Release of human cytomegalovirus from latency by a KAP1/TRIM28 phosphorylation switch. Elife. 2015;4:e06068. | ||
Hu M, Fu X, Cui Y, et al. Expression of KAP1 in epithelial ovarian cancer and its correlation with drug-resistance. Int J Clin Exp Med. 2015;8(10):17308–17320. | ||
Qi ZX, Cai JJ, Chen LC, et al. TRIM28 as an independent prognostic marker plays critical roles in glioma progression. J Neurooncol. 2016;126(1):19–26. | ||
Qi Z, Cai S, Cai J, et al. miR-491 regulates glioma cells proliferation by targeting TRIM28 in vitro. BMC Neurol. 2016;16(1):248. | ||
Petrova YI, Schecterson L, Gumbiner BM. Roles for E-cadherin cell surface regulation in cancer. Mol Biol Cell. 2016;27(21):3233–3244. | ||
Cercelaru L, Stepan AE, Mărgăritescu C, et al. E-cadherin, β-catenin and Snail immunoexpression in laryngeal squamous cell carcinoma. Rom J Morphol Embryol. 2017;58(3):761–766. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.