Back to Journals » Drug Design, Development and Therapy » Volume 19
Traditional Chinese Medicine Natural Products Targeting Shared Mechanisms of T2DM and AD: Potential Therapeutic Insights
Authors Song B, Yue D, Yan H ![]() , Feng L
, Feng L ![]() , Li M
, Li M
Received 2 April 2025
Accepted for publication 20 August 2025
Published 4 September 2025 Volume 2025:19 Pages 7681—7705
DOI https://doi.org/10.2147/DDDT.S531909
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Tamer Ibrahim
Bingbing Song,1,* DongJie Yue,2,* Huiying Yan,1 Lina Feng,1,3 Mingquan Li1
1Department of Neurology, the Third Affiliated Clinical Hospital of the Changchun University of Chinese Medicine, Changchun, Jilin Province, People’s Republic of China; 2Department of Thoracic Surgery, The 32183 Military Hospital of PLA, Baicheng, Jilin Province, People’s Republic of China; 3Shandong Key Laboratory of TCM Multi-Targets Intervention and Disease Control, the Second Affiliated Hospital of Shandong First Medical University, Taian, Shandong Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lina Feng, Boshuo Road, Department of Neurology, the Third Affiliated Clinical Hospital of the Changchun University of Chinese Medicine, Changchun, 130117, Jilin, People’s Republic of China, Tel +8615543120222, Email [email protected] Mingquan Li, Boshuo Road, Department of Neurology, the Third Affiliated Clinical Hospital of the Changchun University of Chinese Medicine, Changchun, 130117, Jilin, People’s Republic of China, Tel +8615948000577, Email [email protected]
Abstract: As highly prevalent chronic diseases globally, AD (Alzheimer’s disease) and T2DM (type 2 diabetes mellitus) not only severely affect the quality of life of patients but also impose a significant burden on their families. Numerous studies have gradually revealed that T2DM and AD are bidirectional risk factors, each capable of exacerbating the other. There is a notable correlation in their pathological mechanisms, primarily manifested in insulin resistance, OS (oxidative stress), and inflammatory responses. Currently, available drugs can only delay the progression of AD, such as Donepezil, Galantamine, and Carbamylcholine, making complete cures challenging to achieve. TCM (Traditional Chinese medicine) natural products exhibit characteristics such as multi-targeting, multi-pathway interactions, diverse biological activities, and relatively mild side effects, enabling synergistic intervention in both diseases. In recent years, TCM natural products have garnered increasing attention in research concentrated on the prevention and management of AD and T2DM. This article aims to comprehensively elucidate the collective pathogenic mechanism of AD and T2DM, and explore the progress of TCM natural products based on these mechanisms in the prevention and treatment of both diseases, thereby providing a theoretical foundation for the advancement of innovative treatment tactics.
Keywords: natural products, traditional Chinese medicine, type 2 diabetes mellitus, T2DM, Alzheimer’s disease, AD
Introduction
AD (Alzheimer’s disease) is a progressive, persistent, and degenerative disorder of the CNS (central nervous system).1 Its main features include a gradual decline in memory, cognitive deficits, alterations in personality, language issues, and various other neuropsychiatric symptoms. In 2018, the Alzheimer’s Disease International Organization approximated that around 50 million people worldwide were affected by dementia, with forecasts suggesting that this number could triple by the year 2050.2 According to its etiopathology, AD is typically classified into fAD (familial AD) and sAD (sporadic AD), with sAD comprising the largest proportion. This form is often associated with complications such as stroke, obesity, cardiovascular diseases, and T2DM (type 2 diabetes mellitus), which can exacerbate the progression of sAD.3 Among these complications, diabetes is one of the most common diseases affecting human beings. In 2019, the worldwide occurrence of diabetes was estimated to be 9.3%, affecting 463 million people. This figure is anticipated to increase to 10.2% (578 million individuals) by 2030 and reach 10.9% (700 million individuals) by 2045.4 Diabetes is a metabolic disorder characterized by elevated blood sugar levels, with T2DM accounting for approximately 90% of all diabetes cases. Many studies have shown a intense association between AD and T2DM, with the risk of AD being nearly twice as high in diabetic patients compared to those without diabetes.5,6 Notably, T2DM is closely linked to an increased risk of developing AD,7,8 and the incidence of AD rises with age.9,10 AD can be considered a central metabolic disorder characterized by impaired glucose metabolism, which is associated with compromised insulin and IGF (insulin-like growth factor) signaling pathways in the brain.11 Additionally, both AD and T2DM are characterized by neuronal and β-cell loss.12 Consequently, AD is often referred to as type 3 diabetes.13 Current research identifies extracellular amyloid protein deposition and intracellular tau protein phosphorylation as the two primary pathological markers of AD.14 Furthermore, IR (insulin resistance) in the brain is recognized as a potential etiological factor in AD15 and serves as a crucial link between AD and T2DM.16 Similarly, significant genetic risk elements for AD, including the ApoE (apolipoprotein E) gene, could intensify the impact of diabetes on cognitive abilities.17
T2DM and AD exhibit overlapping molecular pathways, which encompass IR, OS (oxidative stress), Aβ (amyloid β-protein) deposition, inflammatory, tauhyperphosphorylation, mitochondrial dysfunction, the presence of the ApoE4 allele, and AGEs (advanced glycation end products).11 Peripheral IR can disrupt insulin signaling pathways in the CNS, while IR of CNS may lead to increased Aβ levels, tau hyperphosphorylation, OS, and neuroinflammation.9 Research has found that individuals with diabetes had a 65% higher risk of developing AD than those without diabetes.18,19 Richard et al20 discovered that participants diagnosed with diabetes encountered a more rapid deterioration in cognitive function over the following decade. In studies involving diabetic monkeys, peripheral IR was found to exacerbate AD-related pathology, suggesting that T2DM has the potential to serve as a risk factor for AD.21 An autopsy case revealed the presence of aggregated Aβ and p-tau (phosphorylated tau) protein in the pancreatic islet cells of T2DM patients.22 Juliette et al23 reported that the likelihood of T2DM and impaired fasting blood glucose levels increased among AD patients, with autopsy findings indicating a higher frequency of pancreatic islet amyloidosis in this population. A prospective cohort study indicated that T2DM contributes to Aβ protein deposition.24 In the frontal cortex of T2DM patients undergoing autopsy, an increase in tau phosphorylation was observed, further suggesting that T2DM may elevate the risk of AD.25 Harold et al26 found that abnormal insulin levels in the brain may lead to reduced cognitive function suggests a potential link between IR and AD. Neuroimaging studies have indicated that elderly individuals with IR and T2DM are more likely to exhibit structural and functional brain changes, such as brain atrophy.27 Recent research has indicated that damage to gray matter is closely associated with cognitive impairment,28,29 and cortical atrophy may serve as an independent predictor of cognitive decline,30,31 underscoring the strong relationship between T2DM and AD.
Despite the unremitting efforts of medical researchers, current drugs provide only limited relief of symptoms and do not prevent or reverse T2DM and AD. Metformin has been shown to reduce the prevalence of AD,32 but it may also lead to the overphosphorylation of p-tau and affect Aβ protein aggregation.33,34 The insulin sensitizer rosiglitazone can improve IR in T2DM and protect synapses;35 however, it is not utilized in cases of T2DM combined with AD due to its inability to cross the BBB (blood-brain barrier) and its serious side effects.36 Tacrine has demonstrated efficacy in resisting AD and enhancing cognitive function in db/db mice,37 but it poses significant risks to cardiac and hepatic health.38 Rivastigmine can inhibit neuroinflammation and enhance memory capabilities in T2DM-AD mice, yet it does not modify the insulin abnormalities or amyloid pathology observed in these models.39 Hai et al, investigated the potential of identifying anti-AD medicines within hypoglycemic traditional Chinese medicinal formulas, presenting an alternative approach for screening AD medications. They found that Huangqi-Gegen is a promising treatment for AD, with its mechanism possibly involving the suppression of protein production of acetylcholinesterase (AChE) and cyclooxygenase-2 (PTGS2), reduction of serotonin (5-HT) intake, and subsequent decrease in Aβ deposition.40 Huang-Lian-Jie-Du decoction (HLJDD) is a traditional Chinese formula known for its efficacy in treating diabetes mellitus, Alzheimer’s disease, and diabetic encephalopathy. Tian et al, demonstrated that HLJDD ameliorates cognitive dysfunction in rats with T2DM. The neuroprotective effects are mediated through the modulation of glucose and lipid metabolism, the upregulation of autophagy, and the inhibition of the NLRP3 inflammasome signaling pathway.41 The findings indicate that research on TCM concerning AD and T2DM is still in its nascent stages and requires urgent exploration. Consequently, scholars both domestically and internationally are continuously investigating innovative therapeutic approaches. Over the past few years, TCM (traditional Chinese medicine) has garnered more popularity and acclaim among the general populace. It is regarded as economical, safe, and associated with minimal side effects, while also possessing multi-target functions, which has attracted the attention of a growing number of researchers. Notably, natural products derived from TCM and their active components have shown considerable potential in inhibiting Aβ deposition, reducing p-tau, and improving IR, thus demonstrating significant therapeutic effects on both AD and T2DM. Research has demonstrated that Berberine and Geniposide significantly mitigates hippocampal neuronal injury, reduces Aβ plaque deposition, alleviates abnormal tau protein phosphorylation, and suppresses neuronal apoptosis in diabetic rat model.42,43
This article compiles recent studies on the use of natural products from TCM in the treatment of AD complicated by T2DM, aiming to provide a theoretical and experimental basis for both diseases.
Search Strategy and Selection Criteria
We conducted a comprehensive literature search utilizing Google Scholar, PubMed, the Chinese National Knowledge Infrastructure database, the full-text database of the Chinese Medical Journal (yiigle.com), and the Excerpta Medica (EMBASE) database. The search was guided by the following keywords: “Alzheimer’s disease”, “AD”, “T2DM”, “Type 2 Diabetes Mellitus”, “Cognitive Dysfunction”, “Cognitive Impairment”, “Natural Products”, and “Traditional Chinese Medicine”. This search specifically targeted articles published between 2015 and 2025 and was executed by the author LF.
Shared Pathogenesis of T2DM and AD
Compared to healthy individuals, diabetic patients exhibit a reduction in gray matter volume in the bilateral frontal cortex, parahippocampal gyrus, cerebellar cortex, vermis, brainstem, and bilateral cerebellar tonsil regions.44 Analyses from the cross-sectional study suggested that the decrease in gray matter volume may play a mediating role in the connection between diabetes and cognitive capabilities.45 Currently, the primary pathogenesis of diabetes complicated by cognitive dysfunction includes IR, inflammation, OS, microbiota-gut-brain axis disorder, and cerebral microvascular dysfunction. Among these, IR is a vital feature of T2DM, encompassing both peripheral and central IR. In diabetic patients, IR diminishes the expression of AChE (acetylcholinesterase), reduces ACh (acetylcholine) synthesis, and consequently affects learning and memory functions.
The main pathological features of AD include Aβ and tau protein hyperphosphorylation.46 Aβ is produced through the hydrolysis of APP (amyloid precursor protein) by β and γ-secretases.47 IR has the capacity to affect signaling pathways involving the IRS (insulin receptor substrate) and Akt (protein kinase B), stimulate GSK (glycogen synthase kinase)-3, and facilitate both extracellular deposition of Aβ and hyperphosphorylation of intracellular tau protein,48,49 ultimately leading to NFTs (neurofibrillary tangles).50 The inflammation occurring in the CNS is regarded as one of the underlying pathological mechanisms found in individuals with AD. Research has shown that this inflammatory response is similarly evident in patients with diabetes, characterized by the activation of microglia and astrocytes alongside the infiltration of neuroimmune cells. When astrocytes are activated, they produce a range of pro-inflammatory substances, such as cytokines, chemokines, complement proteins, and ROS (reactive oxygen species), all of which are essential features of CNS inflammation.
Additionally, brain tissue possesses physiological characteristics such as a high oxygen consumption rate, a rich lipid content, and relatively fewer antioxidant enzymes, making it susceptible to oxidative damage. Research findings demonstrate that hyperglycemia can elevate the production of ROS by decreasing the activities of SOD (superoxide dismutase) and GPx (glutathione peroxidase), leading to neuronal apoptosis and necrosis.50 Furthermore, OS is involved in inflammatory responses, by activating microglia and astrocytes and releasing pro-inflammatory cytokines, it contributes to neuroinflammation and neurodegenerative changes.The intestinal microbiota primarily exerts effects on the host through the bacteria themselves and their metabolites. GLP-1 (Glucagon-like peptide-1) and other substances are significantly influenced by the intestinal microbiota and its metabolites, playing roles in the CNS and cognitive function. Another study found that patients suffering from DCD (diabetic cognitive dysfunction) exhibit thickening of the vascular basement membrane, dysfunction in the vascular endothelium, and heightened permeability of the BBB within their brain microvascular system.51 Vascular endothelial dysfunction primarily relates to the accumulation of AGEs, toxic lipids, and protein clumps within small blood vessels. Such deposition can enhance the production of ROS, increase vascular permeability, and damage brain neurons.52
Substantial epidemiological findings indicate a correlation between AD and T2DM. To investigate whether Aβ and p-tau are present in T2DM, pancreatic tissues from 21 autopsy cases (including 10 with T2DM and 11 control subjects) were examined. The analysis reveals that the deposition of Aβ, the hyperphosphorylation of tau, along with immunoreactivities for ubiquitin and ApoE, are distinctive traits associated with T2DM.22 Similar to AD, the deposition of amyloid peptides in the pancreas is a characteristic of T2DM. Aβ and IAPP (islet amyloid polypeptide, also known as amylin) exhibit similar folding patterns and conformations, with approximately 90% overlap in their structural characteristics.23 IAPP is a peptide secreted by islet β cells and induces toxicity in these cells in a manner analogous to Aβ’s effect on neurons.53 Comprising 37 amino acids, IAPP has been shown to exist in the brain either as independent plaques or co-precipitating with Aβ to form composite plaques.54 Under pathological conditions, Aβ and its oligomers exhibit neurotoxicity towards neurons, leading to OS and inflammatory damage, which in turn results in further Aβ deposition and IR (Figure 1).55 Notably, Aβ binds to the insulin receptor, and following specific knockout of this receptor, the activity of GSK-3β increases, leading to neuroinflammation, excessive tau phosphorylation, and the induction of Aβ generation.56–58
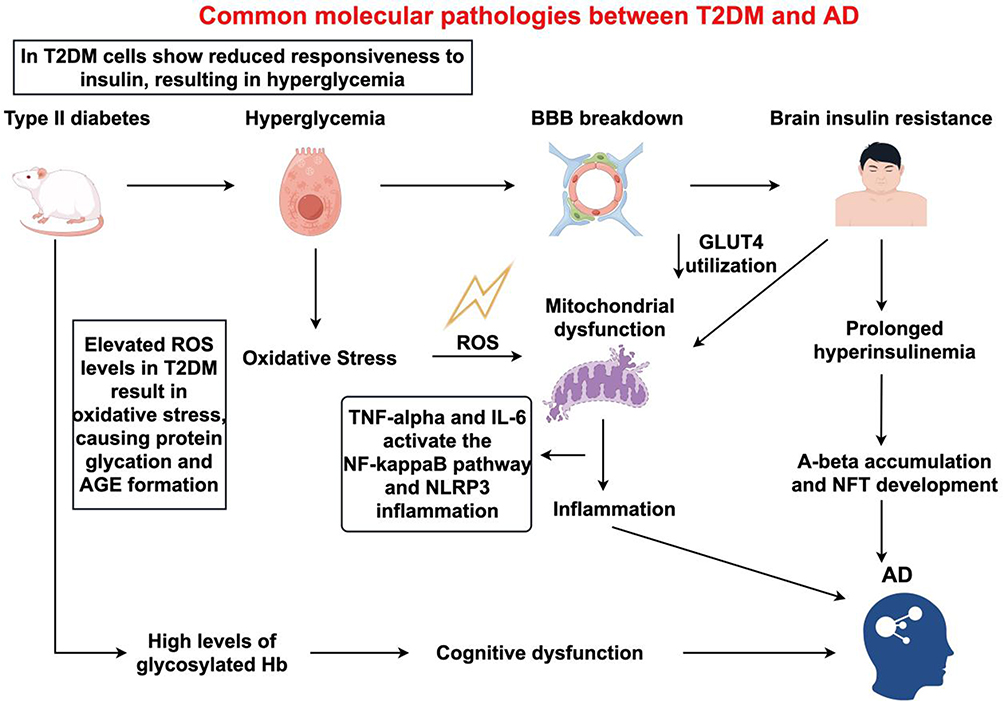 |
Figure 1 The pathogenesis of Type 2 Diabetes Mellitus and Alzheimer’s disease. In T2DM, the sensitivity of cells to insulin decreases, leading to hyperglycemia. The hyperglycemic state results in elevated levels of reactive oxygen species (ROS), which produce oxidative stress (OS), subsequently causing protein glycation and the formation of advanced glycation end products (AGEs). Additionally, hyperglycemia increases the level of glycated hemoglobin (Hb), which can lead to cognitive dysfunction. OS Cascade: The ROS generated by OS can cause mitochondrial dysfunction. Mitochondrial dysfunction further promotes inflammation, during which cytokines such as TNF-α and IL-6 activate the NF-κB signaling pathway and the NLRP3 inflammasome. Blood-Brain Barrier (BBB) and Brain Insulin Resistance: Hyperglycemia can also lead to the disruption of the BBB, resulting in insulin resistance in the brain. Brain insulin resistance impedes the utilization of glucose transporter 4 (GLUT4), leading to a prolonged state of hyperinsulinemia. |
TCM Natural Products Target Mechanisms of T2DM and AD
Insulin Resistance
The insulin signaling pathway predominantly comprises the PI3K (phosphatidylinositol 3-kinase)/Akt signaling pathway and the MAPK (mitogen-activated protein kinase) signaling pathway.59 Abnormalities in this pathway can result in IR, which refers to a diminished sensitivity of the body’s tissues and organs to insulin.60 Given that insulin can traverse the BBB, IR may manifest in both central and peripheral systems,61 contributing to the development of diabetes and leading to the deposition of Aβ and p- tau.55 Under normal conditions, insulin and IGF receptors are abundantly present in the brain, with their degradation primarily mediated by IDE (insulin-degrading enzyme). Both types of receptors are classified as RTKs (receptor tyrosine kinases). Upon binding with their respective ligands, these receptors undergo self-phosphorylation, which subsequently promotes the phosphorylation of various intracellular receptor substrates, including IRS molecules, Shc (SH2-containing proteins), and PI3K, thereby activating distinct pathways and fulfilling physiological functions. In T2DM, impaired insulin signaling can lead to excessive tau phosphorylation, resulting in the formation of NFTs and ultimately causing hippocampal atrophy.62
Research has shown that under normal conditions, the activation of the PI3K/AKT signaling pathway by insulin or IGF primarily influences the synthesis of Aβ40 and Aβ42 by inhibiting the activity of GSK-3α and regulating γ-secretase. A deficiency of insulin or IGF can disrupt Aβ formation and hinder its extracellular transport. Concurrently, elevated levels of Aβ can enhance the clearance of insulin receptors from the cell surface, leading to IR, thereby creating a vicious cycle.63 The promoter region of the human BACE1 (beta-site amyloid precursor protein cleaving enzyme 1) features two active binding sites for NF-κB (nuclear factor-κB). Both in vitro and in vivo studies have indicated that the activation of GSK-3β plays a role in the nuclear translocation of NF-κB/p65 and its interaction with the BACE1 promoter regions, leading to enhanced levels of BACE1 protein and the processing of APP mediated by BACE1, consequently resulting in the production of Aβ.64 The cholinergic system effectively regulates inflammation and immunity by releasing ACh, which acts on the nAChRs (α7-nicotinic acetylcholine receptor) and the M-AchRM1 (muscarinic acetylcholine receptor M1).65 There is an interaction between the signaling pathways downstream of the insulin receptor and the transduction of signals from inflammatory factors. Inflammatory factors produced during non-specific inflammation disrupt the IRS/PI3K signaling pathway, which is the primary molecular mechanism that leads to IR.62
IDE is a protease that degrades Aβ, insulin, IGF-1, and IAPP.61,66 A deficiency in IDE leads to IR. IDE is a neutral thiol endopeptidase with a relative molecular weight of 110,000. Research have indicated that knocking out the IDE gene in mice significantly increases the levels of Aβ40 and Aβ42 in the mouse brain.67 Leissring et al68 transfected neurons to overexpress IDE, resulting in a significant reduction in the level of Aβ in the brain. This overexpression could also slow down and block the formation of amyloid plaques and related cellular pathological changes. Jia et al69 found in their research that the PS-1 V97L mutation may inhibit the expression of IDE by upregulating the transcription factor GATA-3, thereby affecting the content of Aβ42 and participating in the formation of AD. Research indicates that IDE can not only degrade extracellular monomeric Aβ and disrupt the functional domain of APP but also regulate the degradation and clearance of Aβ in the brain, thereby reducing the toxic effects of Aβ.70 Insulin signaling plays a direct role in regulating IDE. The suppression of insulin signaling through PI3K leads to a decrease in IDE protein levels. Strategies aimed at enhancing insulin signaling may represent a novel treatment option for AD. IDE serves as a key regulator of Aβ levels in both microglial and neuronal cells. It was observed that a reduction in PI3K activity resulted in increased Aβ deposition, causing a gradual accumulation of Aβ plaques due to the reduced function of IDE linked to PI3K inhibition and a corresponding decline in insulin signaling. IDE is pivotal in the metabolism of Aβ and its buildup in the brain.71 The increased levels of IDE lead to the down-regulation of IGF-1, which subsequently enhances the activity of IL (interleukin)-1β. An excess of interleukins generates OS within the brain. Various research efforts suggest that IL-1β interacts with A2M (alpha-2-microglobulin), influencing ApoE function, which is involved in the production of APP and, consequently, affects the pathology related to AD.72
Kim et al73 investigated the beneficial impacts of ginsenoside Re on IR induced by a HFD (high-fat diet) in C57BL/6 mice to evaluate its physiological role. Their findings suggest that ginsenoside may improve HFD-induced IR by alleviating hyperglycemia through the protection of cholinergic and antioxidant systems in the brains of the mice. Kang et al74 investigated the EFAA (effects of Artemisia argyi) and 4,5-diCQA (4,5-dicaffeoylquinic acid) on cognitive dysfunction induced by a HFD. Both EFAA and 4,5-diCQA demonstrated efficacy in enhancing cognitive function in the context of HFD-induced cognitive impairment. In conclusion, it is suggested that EFAA may contribute to the amelioration of cognitive deficits associated with HFD, an effect that appears to be closely linked to the physiological activity of 4,5-diCQA. Liu et al75 revealed that injection with STZ (streptozotocin) can effectively inhibit IDE expression and diminish the degradation of Aβ peptides, leading to increased levels of Aβ peptides and the promotion of various pathological features associated with AD. Furthermore, they proposed that treatment with geniposide ameliorated this phenomenon in STZ-induced rats. Zhang et al42 found that administering geniposide for a period of four weeks led to a notable reduction in the phosphorylated levels of tau, as well as a decrease in the acceleration of GSK-3β phosphorylation, in the brains of APP/PS1 (APPswe/PSEN1dE9) transgenic mice affected by insulin deficiency. These results establish a mechanistic and possibly fundamental relationship between diabetes and AD, supporting the idea that geniposide might be crucial in promoting tau protein phosphorylation through the enhancement of insulin signaling, potentially conveying therapeutic benefits in AD. Yan et al76 explored the treatment of AD from two perspectives. On one hand, they demonstrated that a HFD can exacerbate the progression of AD. On the other hand, they found that Okra polysaccharides can ameliorate cognitive disorders in AD model mice. Furthermore, their study confirmed that Okra polysaccharides can reverse cognitive impairment and metabolic disorders induced by HFD through the insulin signaling pathway. Schisandrin A, a lignan derived from the dried fruit of Schisandra chinensis, exhibits a variety of pharmacological effects, including the modulation of immune system control, suppression of apoptosis, and properties of antioxidant and anti-inflammation. Guo et al77 demonstrated that Schisandrin A can improve the deficiencies in spatial learning and memory associated with diabetes, presumably by reducing inflammatory responses and regulating the insulin signaling pathway. BBR (Berberine), derived from TCM, has been utilized for the treatment of diabetes and diabetic cognitive impairment, although its underlying mechanisms remain largely unexplored. In the study conducted by Zhang et al,77 a STZ-induced diabetic rat model fed a HFD was employed to evaluate the effects of BBR in comparison to metformin. The results indicated a decrease in hippocampal neuronal damage, Aβ deposition, tau protein abnormal phosphorylation, and neuronal apoptosis, which are considered to be related mechanisms of action. Another study78 demonstrated that BBR has the potential to enhance memory function, reorganize misaligned nerve cells, and lessen neuronal injury. Additionally, it increased the count of TUNEL-positive cells and lowered high levels of fasting blood glucose, triglycerides, total cholesterol, and glycosylated serum protein in diabetic rats suffering from AD. Furthermore, The treatment with BBR reduced both the transcription levels of mRNAs and the protein expression related to ER (endoplasmic reticulum) stress. Chen et al79 also demonstrated that BBR inhibited the activation of inflammatory pathways and IR in the mPFC (medial prefrontal cortex) of diabetic rats. Furthermore, it improved cognitive impairment in these diabetic rats. Finally, it alleviated cognitive impairment Wu et al79 demonstrated that monomeric BBR treatment improved IR, potentially through the modulation of proteins associated with the dysfunctional insulin signaling pathway. Furthermore, BBR was found to inhibit the production of Aβ42 and mitigate axonal damage in neurons. Ultimately, these findings suggest that BBR may alleviate cognitive impairments induced by diabetes mellitus. Gypenoside LXXV, a novel natural Peroxisome PPAR-γ (proliferator-activated receptor-γ) agonist isolated from Gynostemma pentaphyllum, has been shown to ameliorate cognitive deficits in db/db mice. In the study conducted by Meng et al80 it was demonstrated that Gypenoside LXXV enhanced the cognitive function of APP/PS1 and APP/PS1/db/db mice. This improvement was achieved by promoting glucose uptake, triggered by the activation of the PPARγ/Akt/Glucose transporter (GLUT) 4 signaling pathways. Parida et al81 demonstrated that 1-Deoxynojirimycin seemed to reduce the rise in p-tau and Aβ1-42 levels driven by IR by facilitating the insulin-induced phosphorylation of GSK-3β (a principal tau kinase) and boosting the mRNA expression of the IDE, which is crucial for the clearance of insulin and Aβ. Wang et al82 discovered that Calycosin improved cognitive deficits caused by diabetes in rats by decreasing OS through the PI3K/Akt/GSK-3β signaling pathway.
IR, as a “common soil” for T2DM and AD, is transitioning its research focus from single organ pathology to a systemic metabolic network. Future efforts may involve the design and development of dual-site modulators (such as peripheral IR inhibition combined with brain IR activation), providing a new paradigm for addressing the global health challenge posed by diabetes coexisting with AD.
Neuroinflammation
Glial cells in the CNS primarily consist of microglia, astrocytes, and oligodendrocytes.83 Research has demonstrated that activated microglia aggregate near Aβ. Initially, they clear extracellular Aβ by engulfing, degrading, or releasing lysosomes; however, if the concentration of Aβ secreted by neurons increases to a critical level, disrupting the balance between Aβ clearance and accumulation, long-term chronic inflammation may ensue.84,85 Soluble Aβ oligomers and Aβ fibrils can bind to various receptors on the surface of microglia (such as CD14, CD36, CD47, and TLRs (Toll-like receptors), thereby triggering the release of inflammatory cytokines and chemokines (including IL-1, IL-6, IL-12, IL-23, TNF (tumor necrosis factor)-α, and other inflammatory markers). This cascade leads to chronic inflammation, mediating neuroinflammation and neurotoxicity, and contributes to the accumulation of Aβ and p-tau.86–90 Astrocytes, the most common and largest type of glial cells, support and separate nerve cells while participating in the formation of the BBB.91 Microglia inhibit the destructive phenotype of astrocytes induced by NF-κB, chemokines, and vascular endothelial growth factor by releasing IL-17, sphingolipids, neurotrophin receptors, and cytokine signaling pathways. This inhibitory effect leads to the loss of astrocyte’s ability to promote neuronal survival, growth, synaptogenesis, and phagocytosis, ultimately resulting in the death of neurons and oligodendrocytes.92 In the brain, inflammation caused by IR intensifies neuroinflammatory processes.93 IR leads to persistently high blood sugar levels and an unhealthy imbalance of lipids in the bloodstream, factors that can harm and increase the permeability of the BBB.94 When the BBB is compromised, free fatty acids and elevated blood glucose levels activate microglia, prompting them to adopt a pro-inflammatory phenotype. This activation can result in the release of cytokines, triggering neuroinflammatory cascades and reactive astrogliosis.94,95 Consequently, neuroinflammation exacerbates further damage to the BBB, resulting in even greater permeability.96 This increased permeability raises the already high levels of glucose in the brain, contributing to a detrimental cycle of neuroinflammation driven by hyperglycemia.97,98
Liu et al99 demonstrated that Cucurbitacin B treatment resulted in an increased density of viable neurons in the cortex and hippocampus of rats subjected to STZ- ICV (intracerebroventricular) neurotoxicity. This treatment was associated with enhanced antioxidant activity, a reduction in inflammatory markers, decreased cell death biomarkers, and lower levels of AChE and glutamate. The majority of the Elettaria cardamomum compounds identified were terpenoids. Administration of the Elettaria cardamomum extract over a period of 8 weeks mitigated AD-like alterations through the activation of the damaged insulin signaling pathway in the brain.100 This intervention also reduced associated OS and neuroinflammation, evidenced by a decrease in hippocampal TNF-α and IL-1β levels, while IL-6 levels remained unchanged. Additionally, there was a reduction in GSK-3β levels in the brains of T2DM rats.100 Resveratrol, a type of polyphenol mainly located in grapes and red wine, demonstrates properties that protect against OS, reduce inflammation, and neuroprotective properties. Tian et al101 reported that cognitive performance in the diabetic group was significantly impaired, which was accompanied by notable alterations in OS and inflammatory parameters, as well as reduced expression of SYN (synapse) and GAP-43 (growth-associated protein-43) in the hippocampus. In contrast, chronic administration of resveratrol improved cognitive function and reduced neuronal damage in diabetic rats by alleviating OS and inflammation, while also preventing synaptic degeneration. Another study demonstrated that Resveratrol notably enhanced the expression of sirtuin1 and effectively suppressed memory impairment and mitigated the elevated levels of AChE, MDA (malondialdehyde), IL-1β, and IL-6, while also restoring the decreased levels of ChAT (choline acetyltransferase), SOD, and GSH (glutathione) in this rat model of diabetes and concomitant AD.102 Hyperoside, a flavanone glycoside, is recognized for its wide-ranging pharmacological advantages, such as antioxidant and anti-inflammatory characteristics. Chen et al103 reported that hyperoside mitigated neuroinflammation, cognitive impairment, and OS by suppressing the TNF-α/NF-κB/caspase-3 signaling pathway in rats with T2DM. The findings of Park et al104 demonstrated that Agrimonia pilosa Ledeb, Lonicera japonica Thunb, and Cinnamomum cassia Blume are effective in preventing cognitive dysfunction as well as the disturbance in energy and glucose homeostasis resulting from Aβ accumulation. This effect was mediated through the reduction of neuroinflammation and the enhancement of insulin signaling, which occurred via the activation of the AKT/GSK-3β signaling pathway, alongside an increase in iNOS (inducible nitric oxide synthase) and TNF-α levels. Dioscin exhibits a range of pharmacological activities against various diseases; however, its underlying mechanisms related to DCD remain unclear. The P2X7R (P2X7 receptor), consisting of 595 amino acids, serves as a crucial plasma membrane receptor.105 It promotes the release of pro-inflammatory cytokines, including IL-6, IL-10, and TGF-β (transforming growth factor-β), in microglial cells, fibroblasts, and mast cells.106–108 Additionally, P2X7R is a potent activator of NLRP3 (NOD-like receptor protein 3) inflammasome signaling and facilitates the cleavage of Caspase-1, leading to the release of IL-1β.109,110 Lu et al111 performed a series of pharmacological assessments involving the P2X7R in conjunction with dioscin. Their findings revealed that both P2X7R and NLRP3 inflammasome pathways were initiated in DCD through both in vivo and in vitro analyses. Furthermore, it was established that dioscin’s anti-DCD effects were associated with the suppression of the P2X7R/NLRP3 pathway.111 TMP (Tetramethylpyrazine) is recognized for its anti-diabetic and neuroprotective properties. In the research carried out by Dhaliwal et al112 it was observed that the treatment of diabetic rats with TMP resulted in improved insulin sensitivity, as well as a reduction in hyperglycemia and dyslipidemia. Additionally, the treatment led to increased levels of BDNF (brain-derived neurotrophic factor), p-Akt, and p-CREB (phosphorylated cAMP response element-binding protein), normalization of cholinergic dysfunction, and suppression of OS, inflammation, and apoptotic markers in the hippocampus. Lu et al113 reported that a 2-week administration of TMP reduced impairments in both short-term and long-term memory caused by the ICV injection of STZ in an established rat model of AD. Furthermore, TMP was found to inhibit GSK-3β activity, an essential kinase involved in hippocampal synaptic dysfunction and memory impairments linked to DM. Ultimately, the researchers discovered that treatment with TMP led to the restoration of cholinergic neuron functionality.113 In recent years, extensive research has demonstrated that Rhodiola exhibits a broad spectrum of pharmacological effects on cardiovascular diseases, primarily through its ability to eliminate superoxide anions and hydroxyl radicals in various chemical reactions. The findings of one study indicated that salidroside effectively mitigated cognitive impairment, lowered serum glucose levels, and promoted body weight gain.114 Additionally, salidroside decreased the levels of fasting blood glucose and lipids while inhibiting OS, inflammation, and neuronal apoptosis in a diabetic rat model.114 These findings indicate that salidroside protects against diabetes-induced cognitive impairment by modulating the Rho/ROCK (Rho-associated coiled-coil kinase) / SIRT1 (sirtuin 1)/NF-κB signaling pathway.114
Future research can focus on the penetration of anti-inflammatory components of TCM across the BBB. This can be achieved by utilizing AI-assisted design of brain-targeting carriers, combined with organoid models to predict the distribution and anti-inflammatory activity of effective components from natural products and formulas of TCM within the brain.
Mitochondrial Homeostasis and Oxidative Stress
Mitochondria serve as the primary energy factories of eukaryotic cells, and the adult human brain stands out as one of the body’s most dynamically metabolizing organs, consuming 20% of the total energy supply. In neurons, mitochondria synthesize ATP (adenosine triphosphate) via the ETC (electron transport chain) to support physiological activities, including neurotransmitter transmission and calcium homeostasis.115 Mitochondrial homeostasis is closely linked to mitochondrial biogenesis, dynamics (including fission and fusion), and the regulatory processes of mitophagy. An imbalance in mitochondrial homeostasis, characterized by a reduction in mitochondria, impaired fusion and fission, and defective mitophagy, is associated with the onset and progression of AD and T2DM.116,117 OS is defined as the imbalance between the processes of oxidation and antioxidation that occur within the body, which can diminish peripheral insulin sensitivity by causing dysfunction in pancreatic β-cells and mitochondria, promoting inflammatory responses, downregulating GLUT 4, and impairing normal insulin signaling pathways. This cascade ultimately leads to IR and the onset of diabetes.118 Concurrently, OS is a critical factor in neuronal damage and serves as a significant trigger for the core pathological changes related to the occurrence and advancement of AD.119–123 The brain exhibits a high oxygen consumption rate, and its cell membranes are abundant in polyunsaturated fatty acids, rendering it vulnerable to OS, which can result in neuronal damage and synaptic loss.124 In animal models of IR, an imbalance in redox reactions—characterized by excessive mitochondrial fusion and diminished ATP production—has been observed, demonstrating a correlation between IR and OS. Xu et al also reported the generation of OS in T2DM rats induced by a HFD.125 Research has indicated that OS can induce the serine phosphorylation of IRS-1, which impairs insulin signal transduction and concurrently boosts the expression of phosphorylated GSK-3β, and upregulating the expression of the BACE1 gene, which further exacerbates the production of Aβ and induces neuronal apoptosis.125–127 A substantial accumulation of ROS gives rise to the generation of AGEs and stimulates the expression of the pro-inflammatory transcription factor NF-κB.128,129 It can be inferred that IR, OS, and related factors are closely associated with T2DM and AD, contributing to damage in pancreatic β cells and neurons, which ultimately exacerbates both T2DM and AD.130,131
Ermis et al132 found that diabetic rats exhibited elevated levels of Aβ, tau protein, and TOC (total oxidant capacity), while TAC (total antioxidant capacity) levels were reduced. Furthermore, in rats with active diabetes that received curcumin treatment, the levels of Aβ protein were decreased both in the serum and the brain. Additionally, serum TAC levels were higher in curcumin-treated rats. These results suggest that curcumin may ameliorate the complications of diabetes by enhancing antioxidant activity and mitigating OS associated with the condition. Centella asiatica has been reported to exhibit CNS activity, including the enhancement of cognitive functions such as intelligence. Another study demonstrated that due to its neuroprotective properties, Centella asiatica effectively reduced cognitive decline and alleviated OS caused by disrupted brain glucose metabolism in rats treated with ICV-STZ.133 Crocin improved spatial learning ability and mitigated memory impairments in diabetic rats by lessening oxidative damage in the cerebral cortex.134 This effect might be credited to its antidiabetic and antioxidant properties, suggesting potential clinical applications for treating cognitive dysfunction in diabetic patients.134 Xia et al135 demonstrated that an appropriate dose of quercetin could attenuate OS through the PI3K/AKT/ Nrf 2 (nuclear factor erythroid-2-related factor 2) signaling pathway, thereby enhancing hippocampus-dependent cognition in mice with T2DM. Quercetin exerts beneficial impacts by decreasing protein glycation, OS, and inflammation. It achieves this through upregulating Glo-1 (Glyoxalase 1), a mechanism that has the potential to alleviate diabetic encephalopathy. Glo-1 is a widely distributed cellular enzyme involved in the detoxification process of MG (methylglyoxal), a harmful byproduct of glycolysis that causes protein modification, specifically AGEs, OS, and inflammation. These findings provide a novel strategy for the treatment of AD in conjunction with T2DM.136 Parvin et al137 demonstrated that both simultaneous and separate administration of artemisinin significantly ameliorated the adverse effects associated with AD by significantly decreasing the formation of Aβ plaques in the hippocampus. Additionally, artemisinin reduced levels of glucose, MDA, ROS, and TNF-α, while increasing TAC levels.137 These research results indicate that artemisinin possesses the capacity to ameliorate the pathological manifestations of AD in a rat model that simulates both AD and diabetes. Consequently, artemisinin has the potential to act as an auxiliary therapeutic option and a safeguard for individuals suffering from AD.137 Khan et al138 conducted a study aiming to evaluate the effectiveness and dose-response relationship of nuciferine and the crude extract derived from Nelumbo nucifera in treating T2DM and AD. The study concluded that both nuciferine and the crude extract of Nelumbo nucifera remarkably enhance memory and can potentially be regarded as an effective phytomedicine for T2DM, AD, and OS.138 Diosgenin, a steroidal saponin, is noted for its anti-diabetic effects and multiple protective properties. Mahmoudi et al139 demonstrated that diosgenin can improve cognitive deficits in animals with STZ-induced diabetes. This enhancement is partly due to its ability to reduce OS, inflammation, and astrogliosis, while also potentially improving cholinergic function and exhibiting neuroprotective properties. Paeoniflorin, a natural glycoside derived from the Paeonia lactiflora Pall, is recognized for its properties that include anti-diabetic, antioxidant, anti-inflammatory, and neuroprotective effects.140 Wang et al140 demonstrated the protective effects of Paeoniflorin on neurobehavior, OS, brain insulin signaling, and synaptic alterations in mice with cognitive impairment induced by ICV-STZ. Overall, the findings of their research indicate that Paeoniflorin may possess therapeutic potential for alleviating the cognitive impairments induced by ICV-STZ. Furthermore, another study demonstrated that Paeoniflorin has beneficial effects in alleviating cognitive deficits associated with diabetes by modulating the SOCS2 (suppressor of cytokine signaling 2)/IRS-1 pathway.141 This suggests that Paeoniflorin may offer a viable therapeutic approach for managing cognitive dysfunction related to diabetes.141 Matrine, an active component derived from the root extracts of Sophora flavescens Ait, exhibits a wide range of pharmacological activities. Zhang et al142 demonstrated that Matrine ameliorates fasting blood glucose levels and attenuates the spatial learning and memory dysfunctions caused by diabetes. The PK2/PKRs (prokineticin 2/prokineticin receptors) signaling pathway plays a role in the neuroprotective actions of Matrine. Furthermore, Matrine alleviated the expression of ER stress-related proteins in diabetic mice. Oxymatrine serves as the major quinolizidine alkaloid that is extracted from the root of Sophora flavescens Ait. Huang et al143 demonstrated that Oxymatrine alleviates the cognitive decline associated with diabetes, OS, and apoptosis through the inhibition of NOX 2 (nicotinamide adenine dinucleotide phosphate hydrogen oxidase) and NOX 4. However, the aforementioned research indicates that the investigation of TCM natural products in regulating mitochondrial homeostasis in the context of T2DM combined with AD remains in its early stages, presenting significant potential for further exploration (Figure 2).
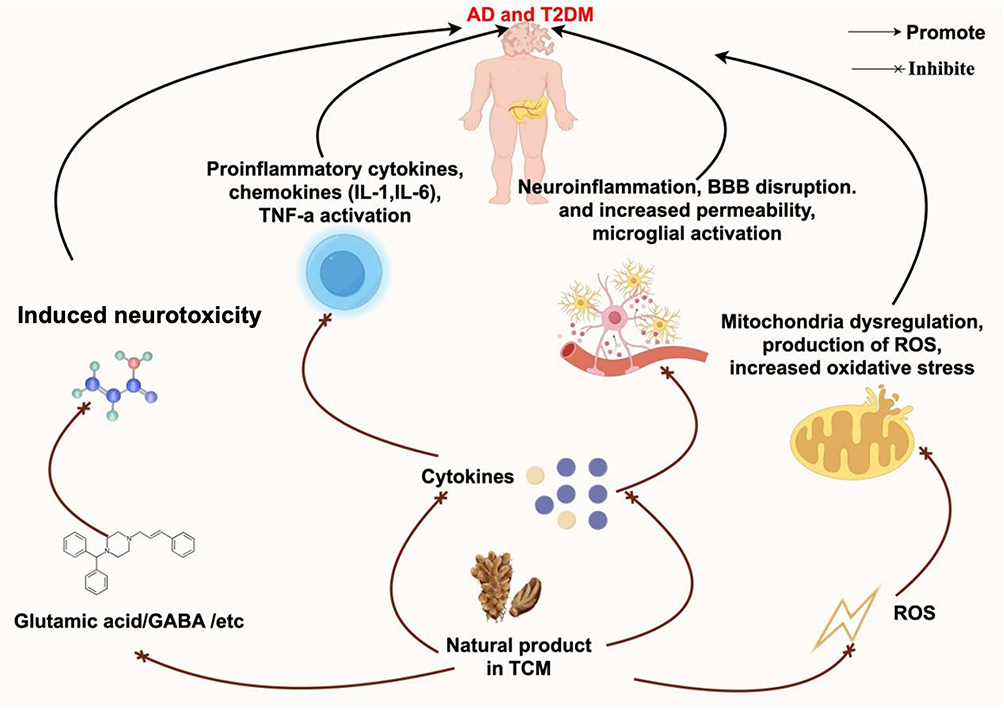 |
Figure 2 The potential mechanisms of action of Natural products in traditional Chinese medicine: on Type 2 Diabetes Mellitus and Alzheimer’s disease. Natural products in Traditional Chinese Medicine (TCM) exhibit mechanisms of action as follows: Regulation of cytokines: Natural products in TCM act on cytokines, inhibiting the activation of pro-inflammatory cytokines and chemokines (such as IL-1, IL-6) and TNF-α, thereby reducing neuroinflammation, blood-brain barrier (BBB) disruption, increased permeability, and microglial activation, which in turn affects Alzheimer’s disease (AD) and type 2 diabetes mellitus (T2DM). Inhibition of neurotoxicity: These products can inhibit neurotoxicity induced by glutamic acid and γ-aminobutyric acid (GABA), thereby mitigating their adverse effects on AD and T2DM. Improvement of mitochondrial function: By inhibiting mitochondrial dysregulation, they reduce the production of reactive oxygen species (ROS) and oxidative stress, thus playing a role in AD and T2DM. |
Future research could further explore the dual-edged effects of OS regulation, such as designing responsive release of antioxidants that break through the limitations of traditional antioxidant therapies through ‘smart responses’, upgrading “passive antioxidant” strategies to “active precision interventions”. Additionally, cryo-electron microscopy could be employed to elucidate the binding sites between TCM components and mitochondrial proteins, analyzing the interactions between components and targets.
AGEs
AGEs are irreversible end products formed through a series of reactions between the aldehyde or ketone groups of glucose or other reducing sugars in the body and the free amino groups of macromolecules, such as proteins, nucleic acids, and lipids. These products trigger a cascade of oxidative reactions within the organism, subsequently leading to tissue damage.144,145 The primary mechanism mechanism by which AGEs induce IR and diabetic complications involves their binding to the RAGE (receptor for advanced glycation end) products, which subsequently initiates a series of pathological effects.146 The extracellular domain of AGEs binds to RAGE and activates its cytoplasmic domain, generating multiple signaling pathways, including MAPK, JNK (c-Jun N-terminal kinase)/ SAPK (stress-activated protein kinase), ERK (extracellular signal-regulated kinases), and JAK-STAT (Janus kinase-signal transducer and activator of transcription). This activation results in the sustained activation of intracellular NF-κB and STAT 3, ultimately upregulating various inflammatory cytokines and activating reduced nicotinamide adenine dinucleotide oxidase to generate ROS.147 In recent years, AGEs have been recognized as significant risk factors for cognitive dysfunction. The serum of diabetic patients contains six distinct AGE structures, referred to as AGEs-1 through AGEs-6. Among these, AGE-2 demonstrates biological activity towards vascular wall cells and cortical neurons, playing a crucial role in the pathogenesis of diabetic complications and AD. Studies have indicated that the accumulation of AGEs occurs in the brains of diabetic rats, where they can promote the formation of NFT and Aβ, enhance the cytotoxicity of Aβ, and participate in the pathological processes associated with AD.148 In transgenic AD mice, RAGE in microglia increases the release of pro-inflammatory cytokines such as IL-1β and TNF-α, which impairs neuronal function and directly influences Aβ accumulation.149 Furthermore, glycosylation of AGEs at the N-terminal of tau protein, the microtubule binding site, results in microtubule instability.150 This instability promotes the entanglement of neurofilaments and the aggregation of amyloid oligomers within brain tissue.151 Moreover, animal models have confirmed that AGEs precursors can induce symptoms of AD, contributing to the onset and progression of cognitive dysfunction in diabetes.152,153 In vivo studies have shown that, compared to patients with AD alone, the postmortem brains of patients with both AD and diabetes exhibit increased levels of RAGE and elevated AGE levels in immunohistochemical markers. This suggests that the OS mechanism promoted by AGEs may underlie the development of AD pathology in diabetic patients.52,154 A cross-sectional study involving 167 patients with T2DM revealed that elevated serum AGE levels and reduced RAGE concentrations are significantly associated with an increased prevalence of mild cognitive impairment.155 Schnaider Beeri et al156 included 684 non-demented elderly individuals in a cohort study with an average follow-up of three years to explore the association between dietary AGEs and cognitive decline in the elderly. The study found that higher baseline dietary AGEs were linked to a faster decline in overall cognitive function, indicating a close relationship between AGEs and cognitive dysfunction in diabetes.
Lychee seed is a TCM known for its numerous beneficial effects, including the regulation of blood glucose and lipid levels, antioxidant effects, antiviral properties, and antitumor activities.157 Research has shown that the possible mechanisms linked to lychee seed could be related to enhanced IR and a reduction in the production of tau protein, Aβ, and AGEs within the hippocampus of rats suffering from T2DM.157 Astragalus membranaceus, commonly referred to as “Huangqi”, is a herb widely utilized in TCM and is also employed in the formulation of various health supplements. The herb’s antioxidant,158 anti-diabetic,159 and neuroprotective160 properties have contributed to its extensive clinical applications. Calycosin, a bioactive flavonoid compound isolated from Astragalus membranaceus roots, exhibits diverse pharmacological properties, holds considerable importance in its therapeutic applications. Studies indicated that calycosin successfully suppresses GSK-3β activity, leading to a significant reduction in p-tau.160 Additionally, it inhibited the expression of RAGE and BACE-1 proteins, leading to attenuated Aβ generation. Furthermore, calycosin regulated the PGC-1α/TFAM (peroxisome proliferator-activated receptor-γ coactivator-1α/mitochondrial transcription factor A) signaling pathway to repair mitochondrial dysfunction.161 Therefore, it can be concluded that calycosin may have considerable therapeutic benefits that can be utilized in managing AD, particularly in cases related to diabetes mellitus.161 Scutellaria baicalensis Georgi is a renowned herbal remedy in TCM, incorporated into a range of clinical formulations. Modern pharmacological studies have demonstrated that the ethanol extract from S. baicalensis has the ability to improve memory in mice subjected to chronic injections of D-galactose and sodium nitrate.162 This restorative effect is attributed to its antioxidant, anti-inflammatory, and anti-apoptotic properties.163 Zheng et al164 demonstrated that baicalin could reinstate cognitive and spatial memory in diabetic GK (Goto-Kakizaki) rats. Moreover, it lessened the loss of neurons and degraded the expression levels of Aβ and p-tau within these rats.164 At the cellular level, baicalin exhibited significant antioxidant and anti-inflammatory effects in response to high-glucose stimulation.164 The primary molecular mechanism underlying baicalin neuroprotective action against diabetes-induced cognitive impairment is the upregulation of Nrf2 signaling. AGE is a recognized risk factor and emerging biomarker for the chronic complications of diabetes. Numerous studies have confirmed the correlation between AGE and these complications. However, therapeutic interventions targeting AGE are still in their infancy and require further clinical trials to assess therapeutic potential and safety. The current challenges primarily include the lack of standardized detection methods for AGEs. In the future, multi-center clinical data calibration detection methods could be employed to reduce inter-laboratory variability. Additionally, targeted traditional Chinese medicine formulations for AGEs could be developed; targeted nanoenzymes for AGEs could also be created. Furthermore, network pharmacology combined with organoid models can be utilized to screen for effective components. With advancements in technology and the accumulation of clinical research experience, AGE is poised to become a significant predictor and intervention target for the chronic complications of diabetes. For more details about Natural Products on T2DM and AD, please refer to Table 1.
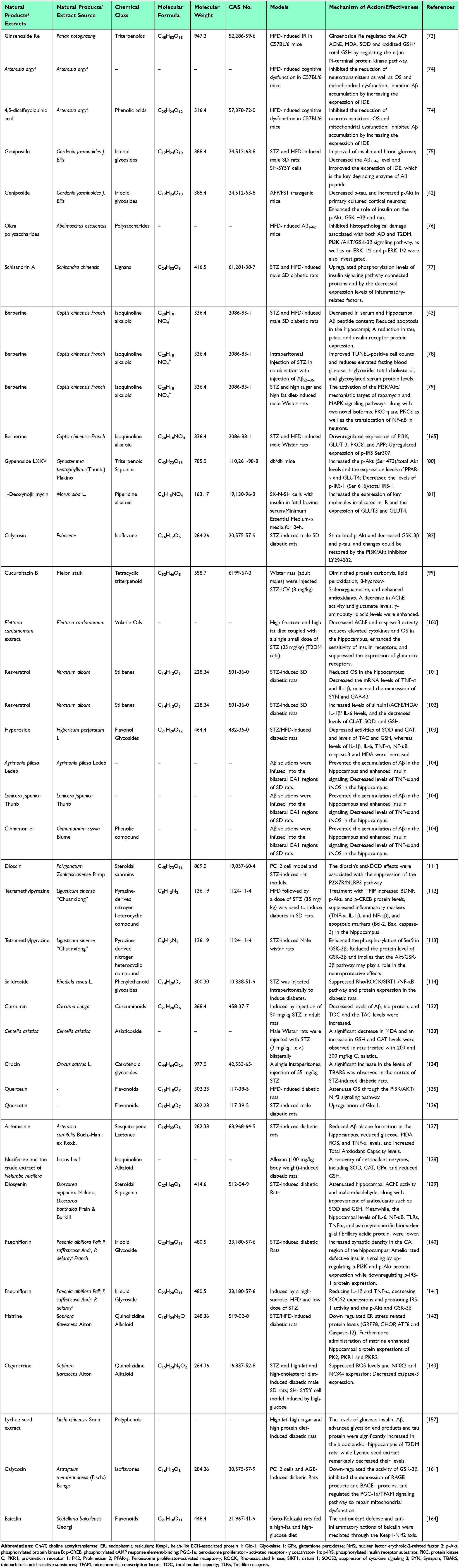 |
Table 1 The Potential Mechanisms of Natural Products on Type 2 Diabetes Mellitus and Alzheimer’s Disease |
Discussion
In 1950, Russell first proposed the concept of diabetic encephalopathy, describing the central neuropathy associated with diabetes and incorporating cognitive impairment as a complication of the disease. In 1965, pathological changes observed in the brains of 16 adolescents who died from diabetic vascular complications further substantiated the existence of diabetic encephalopathy.166 Moheet et al167 discovered that patients with T2DM experienced a notable decline in cognitive abilities in comparison to the control group. Previous cross-sectional studies have indicated that the prevalence of MCI (Mild Cognitive Impairment) among patients with T2DM ranges from approximately 20% to 30%, while the incidence of dementia is about 17.3%.168 A domestic meta-analysis established that patients with T2DM exhibit significantly elevated risks of developing cognitive dysfunction, when contrasted with the general populace, revealing a risk for developing AD that is 1.46 times greater than that of non-diabetic individuals.169 The cognitive dysfunction associated with diabetes has an insidious onset, primarily affecting middle-aged and elderly individuals. Following the onset of the disease, the decline in cognitive function among patients occurs approximately 50% faster than that observed in the naturally aging population.168 Advances in medical technology and the extension of life expectancy have drawn increasing attention to chronic diseases in the elderly. Diabetes, as a prominent example of chronic metabolic diseases, leads to cognitive dysfunction that significantly impairs the quality of life among the aging population and reduces family happiness indices. Consequently, there is an urgent need for improved diagnostic and therapeutic methods for intervention. The treatment of T2DM in conjunction with AD using natural products from TCM has attracted growing interest from scholars due to its stable efficacy, fewer adverse reactions, and multi-target characteristics. These treatments can improve IR, inhibit OS, mitochondrial dysfunction, and neuroinflammation, thereby delaying the onset and progression of diabetic complications to some extent.
Research on the application of natural products derived from TCM in the treatment of AD in conjunction with T2DM presents unique advantages across multiple dimensions. In terms of mechanisms of action, natural TCM products typically exhibit multi-component and multi-target properties. They not only regulate the core pathological processes of T2DM but also intervene in the neurodegenerative changes associated with AD by inhibiting Aβ, improving excessive p-tau, and reducing neuroinflammation, thereby achieving a synergistic regulation of both diseases. Moreover, many natural TCM products have demonstrated a high level of safety in long-term clinical applications, which can minimize the risk of adverse reactions often associated with single-target medications. This characteristic makes them particularly suitable for patients with chronic and complex conditions requiring prolonged treatment. Additionally, TCM adheres to the principle of “holistic regulation”, which can simultaneously enhance the overall metabolic status and cognitive function of patients. This approach offers more comprehensive benefits in alleviating systemic symptoms related to comorbidities, thus providing a treatment strategy that better aligns with the overall health needs of individuals suffering from the complex interplay of AD and T2DM.
Despite the significant associations in clinical comorbidity and pathological features between T2DM and AD, the direct mechanisms of action between the two remain under exploration. Current research primarily relies on epidemiological correlations and indirect evidence, such as the markedly increased risk of cognitive dysfunction in diabetic patients and the manifestation of insulin resistance in both diseases. However, there is still a lack of direct elucidation regarding how diabetes drives the pathological processes of AD, or how AD-related lesions affect the molecular mechanisms of glucose metabolic homeostasis. Currently, existing studies mainly focus on the mechanisms of single herbs or herb pairs, or predict potential targets of formulas through network pharmacology and molecular docking techniques, yet often do not integrate multi-omics approaches. Additionally, the number of targets investigated remains limited, and the mechanisms explored are frequently overly simplistic. Future research should consider exploring more potential mechanisms of action using methods such as spatial transcriptomics and single-cell sequencing. There is a pressing need to strengthen interdisciplinary collaboration, incorporating organoids and gene chips as research subjects to better simulate the in vivo micro-environment. Simultaneously, comprehensive studies on the compatibility of formulas specifically for AD combined with T2DM are scarce. Future efforts should involve high-quality clinical studies, such as multicenter, large-sample trials and collaborative analyses of TCM and Western medicine mechanisms, to further explore the potential of these formulas and provide optimized treatment plans for complex comorbidities. Furthermore, it is essential to enhance preclinical and clinical trial research, optimize the extraction, separation, and modification techniques of natural products, and develop efficient and safe natural drugs. Besides, the lack of standardization has made it difficult for TCM to gain widespread recognition internationally, thereby limiting its process of internationalization. Low bioavailability may lead to the insufficient efficacy of medications, affecting clinical treatment outcomes and hindering the translation process from laboratory research to clinical application. Standardization encompasses multiple dimensions, including the quality of medicinal materials, processing techniques, formulation specifications, and efficacy evaluation. For instance, significant differences in the content of active ingredients in medicinal materials from different origins and harvest seasons can lead to unstable drug quality due to the absence of unified standards. Moreover, many active components of TCM exhibit poor water solubility, difficult intestinal absorption, or are easily metabolized by the first-pass effect, which affects their clinical efficacy. In the future, a comprehensive quality traceability system for medicinal materials can be established, along with strict processing and formulation specifications, and the introduction of modern scientific efficacy evaluation indicators. To address bioavailability issues, methods such as nanotechnology, liposome encapsulation, and structural modification can be employed to improve the physicochemical properties of TCM components and enhance their absorption efficiency. These efforts will provide new insights and methodologies for the prevention and treatment of AD and T2DM, thereby promoting the widespread application of TCM in addressing these conditions.
Abbreviations
Aβ, amyloid β-protein; ACh, acetylcholine; AChE, acetylcholinesterase; AD, Alzheimer’s disease; AGEs, advanced glycation end products; Akt, protein kinase B; ApoE, apolipoprotein E; APP, amyloid precursor protein; APP/PS1, APPswe/PSEN1dE9; BACE1, beta-site amyloid precursor protein cleaving enzyme 1; BBB, blood-brain barrier; BBR, Berberine; BDNF, brain-derived neurotrophic factor; CAT, catalase; CNS, central nervous system; DCD, diabetic cognitive dysfunction; EFAA, effects of Artemisia argyi; ERK, extracellular signal-regulated kinases; GLUT, Glucose transporter; GSH, glutathione; GSK, glycogen synthase kinase; HFD, high-fat diet; IAPP, islet amyloid polypeptide; ICV, intracerebroventricular; IDE, insulin-degrading enzyme; IGF, insulin-like growth factor; IL, interleukin; iNOS, inducible nitric oxide synthase; IR, insulin resistance; IRS, insulin receptor substrate; MAPK, mitogen-activated protein kinase; MDA, malondialdehyde; NF-κB, nuclear factor-κB; NFTs, neurofibrillary tangles; NLRP3, NOD-like receptor protein 3; NOX, nicotinamide adenine dinucleotide phosphate hydrogen oxidase; Nrf2, nuclear factor erythroid-2-related factor 2; OS, oxidative stress; PI3K, phosphatidylinositol 3-kinase; PKRs, prokineticin receptors; p-tau, phosphorylated tau; P2X7R, P2X7 receptor; RAGE, receptor for advanced glycation end-products; ROS, reactive oxygen species; sAD, sporadic AD; SD, Sprague-Dawley; SOD, superoxide dismutase; STZ, streptozotocin; TAC, total antioxidant capacity; TCM, traditional Chinese medicine; TMP, Tetramethylpyrazine; TNF, Tumor Necrosis Factor; T2DM, type 2 diabetes mellitus; 4,5-diCQA, 4,5-dicaffeoylquinic acid.
Data Sharing Statement
All of the data used to support the findings of this study are included within the article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Natural Science Foundation of Shandong (ZR2023QH159) and the Shandong Province Medical Health Science and Technology Development Plan Project (202313011384).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
1. Masters CL, Bateman R, Blennow K, et al. Alzheimer’s disease. Nat Rev Dis Primers. 2015;1:15056. doi:10.1038/nrdp.2015.56
2. Scheltens P, De Strooper B, Kivipelto M, et al. Alzheimer’s disease. Lancet. 2021;397(10284):1577–1590. doi:10.1016/S0140-6736(20)32205-4
3. Burillo J, Marques P, Jimenez B, et al. Insulin resistance and diabetes mellitus in alzheimer’s disease. Cells. 2021;10(5). doi:10.3390/cells10051236.
4. Saeedi P, Petersohn I, Salpea P, et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: results from the international diabetes federation diabetes atlas, 9(th) edition. Diabet Res Clin Pract. 2019;157:107843. doi:10.1016/j.diabres.2019.107843
5. de la Monte SM. Insulin resistance and alzheimer’s disease. BMB Rep. 2009;42(8):475–481. doi:10.5483/BMBRep.2009.42.8.475
6. Sims-Robinson C, Kim B, Rosko A, Feldman EL. How does diabetes accelerate Alzheimer disease pathology?. Nat Rev Neurol. 2010;6(10):551–559. doi:10.1038/nrneurol.2010.130
7. Saczynski JS, Jonsdottir MK, Garcia ME, et al. Cognitive impairment: an increasingly important complication of type 2 diabetes: the age, gene/environment susceptibility--Reykjavik study. Am J Epidemiol. 2008;168(10):1132–1139. doi:10.1093/aje/kwn228
8. Tolppanen A, Lavikainen P, Solomon A, et al. History of medically treated diabetes and risk of Alzheimer disease in a nationwide case-control study. Diabetes Care. 2013;36(7):2015–2019.
9. Nguyen TT, Ta Q, Nguyen T, Nguyen T, Giau VV. Type 3 diabetes and its role implications in alzheimer’s disease. Int J Mol Sci. 2020;21(9):3165. doi:10.3390/ijms21093165
10. Sridhar GR, Lakshmi G, Nagamani G. Emerging links between type 2 diabetes and Alzheimer’s disease. World J Diabetes. 2015;6(5):744–751. doi:10.4239/wjd.v6.i5.744
11. Hamze R, Delangre E, Tolu S, et al. Type 2 diabetes mellitus and alzheimer’s disease: shared molecular mechanisms and potential common therapeutic targets. Int J Mol Sci. 2022;23(23):15287. doi:10.3390/ijms232315287
12. Li L, Holscher C. Common pathological processes in alzheimer disease and type 2 diabetes: a review. Brain Res Rev. 2007;56(2):384–402. doi:10.1016/j.brainresrev.2007.09.001
13. Liu P, Wang ZH, Kang SS, et al. High-fat diet-induced diabetes couples to Alzheimer’s disease through inflammation-activated C/EBPbeta/AEP pathway. Mol Psychiatry. 2022;27(8):3396–3409. doi:10.1038/s41380-022-01600-z
14. DeTure MA, Dickson DW. The neuropathological diagnosis of Alzheimer’s disease. Mol Neurodegener. 2019;14(1):32. doi:10.1186/s13024-019-0333-5
15. Talbot K, Wang H, Kazi H, et al. Demonstrated brain insulin resistance in Alzheimer’s disease patients is associated with IGF-1 resistance, IRS-1 dysregulation, and cognitive decline. J Clin Invest. 2012;122(4):1316–1338. doi:10.1172/JCI59903
16. Haan MN. Therapy Insight: type 2 diabetes mellitus and the risk of late-onset Alzheimer’s disease. Nat Clin Pract Neurol. 2006;2(3):159–166. doi:10.1038/ncpneuro0124
17. McFall GP, Wiebe SA, Vergote D, Anstey KJ, Dixon RA. Alzheimer’s genetic risk intensifies neurocognitive slowing associated with diabetes in non-demented older adults. Alzheimers Dement. 2015;1(4):395–402. doi:10.1016/j.dadm.2015.08.002
18. Arvanitakis Z, Wilson RS, Bienias JL, Evans DA, Bennett DA. Diabetes mellitus and risk of Alzheimer disease and decline in cognitive function. Arch Neurol. 2004;61(5):661–666. doi:10.1001/archneur.61.5.661
19. Alsharif AA, Wei L, Ma T, et al. Prevalence and incidence of dementia in people with diabetes mellitus. J Alzheimers Dis. 2020;75(2):607–615. doi:10.3233/JAD-191115
20. Tuligenga RH, Dugravot A, Tabak AG, et al. Midlife type 2 diabetes and poor glycaemic control as risk factors for cognitive decline in early old age: a post-hoc analysis of the Whitehall II cohort study. Lancet Diabetes Endocrinol. 2014;2(3):228–235. doi:10.1016/S2213-8587(13)70192-X
21. Krizan I, Solingapuram SK, Damuka N, et al. Exploratory Dual PET imaging of [(18)F] fluorodeoxyglucose and [(11)C] acetoacetate in type 2 diabetic nonhuman primates. Bioorg Med Chem Lett. 2024;111:129906. doi:10.1016/j.bmcl.2024.129906
22. Miklossy J, Qing H, Radenovic A, et al. Beta amyloid and hyperphosphorylated tau deposits in the pancreas in type 2 diabetes. Neurobiol Aging. 2010;31(9):1503–1515. doi:10.1016/j.neurobiolaging.2008.08.019
23. Janson J, Laedtke T, Parisi JE, et al. Increased risk of type 2 diabetes in Alzheimer disease. Diabetes. 2004;53(2):474–481. doi:10.2337/diabetes.53.2.474
24. van Arendonk J, Neitzel J, Steketee R, et al. Diabetes and hypertension are related to amyloid-beta burden in the population-based Rotterdam Study. Brain. 2023;146(1):337–348. doi:10.1093/brain/awac354
25. Liu Y, Liu F, Grundke-Iqbal I, Iqbal K, Gong CX. Brain glucose transporters, O-GlcNAcylation and phosphorylation of tau in diabetes and Alzheimer’s disease. J Neurochem. 2009;111(1):242–249. doi:10.1111/j.1471-4159.2009.06320.x
26. Cholerton B, Baker LD, Craft S. Insulin resistance and pathological brain ageing. Diabet Med. 2011;28(12):1463–1475. doi:10.1111/j.1464-5491.2011.03464.x
27. Brundel M, Kappelle LJ, Biessels GJ. Brain imaging in type 2 diabetes. Eur Neuropsychopharmacol. 2014;24(12):1967–1981. doi:10.1016/j.euroneuro.2014.01.023
28. Eijlers AJC, van Geest Q, Dekker I, et al. Predicting cognitive decline in multiple sclerosis: a 5-year follow-up study. Brain. 2018;141(9):2605–2618. doi:10.1093/brain/awy202
29. Stellmann J, Wanke N, Maarouf A, et al. Cognitive performance shows domain specific associations with regional cortical thickness in multiple sclerosis. Neuroimage Clin. 2021;30:102606. doi:10.1016/j.nicl.2021.102606
30. Benedict RHB, Amato MP, DeLuca J, Geurts JJG. Cognitive impairment in multiple sclerosis: clinical management, MRI, and therapeutic avenues. Lancet Neurol. 2020;19(10):860–871. doi:10.1016/S1474-4422(20)30277-5
31. Sastre-Garriga J, Pareto D, Battaglini M, et al. MAGNIMS consensus recommendations on the use of brain and spinal cord atrophy measures in clinical practice. Nat Rev Neurol. 2020;16(3):171–182. doi:10.1038/s41582-020-0314-x
32. Zheng J, Xu M, Walker V, et al. Evaluating the efficacy and mechanism of metformin targets on reducing Alzheimer’s disease risk in the general population: a Mendelian randomisation study. Diabetologia. 2022;65(10):1664–1675. doi:10.1007/s00125-022-05743-0
33. Al-Kuraishy HM, Al-Gareeb AI, Saad HM, Batiha GE. Benzodiazepines in Alzheimer’s disease: beneficial or detrimental effects. Inflammopharmacology. 2023;31(1):221–230. doi:10.1007/s10787-022-01099-4
34. Chen Y, Zhou K, Wang R, et al. Antidiabetic drug metformin (GlucophageR) increases biogenesis of Alzheimer’s amyloid peptides via up-regulating BACE1 transcription. Proc Natl Acad Sci U S A. 2009;106(10):3907–3912. doi:10.1073/pnas.0807991106
35. De Felice FG, Vieira MN, Bomfim TR, et al. Protection of synapses against Alzheimer’s-linked toxins: insulin signaling prevents the pathogenic binding of Abeta oligomers. Proc Natl Acad Sci U S A. 2009;106(6):1971–1976. doi:10.1073/pnas.0809158106
36. Ma L, Hu W. Progress on the relationship of Alzheimer’s disease with diabetes and their treatment. Chin J Geriatr. 2015;34(9):1037–1041.
37. Zhao Q, Matsumoto K, Tsuneyama K, et al. Diabetes-induced central cholinergic neuronal loss and cognitive deficit are attenuated by tacrine and a Chinese herbal prescription, kangen-karyu: elucidation in type 2 diabetes db/db mice. J Pharmacol Sci. 2011;117(4):230–242. doi:10.1254/jphs.11115FP
38. Peng Y, Yao S, Chen Q, et al. True or false? Alzheimer’s disease is type 3 diabetes: evidences from bench to bedside. Ageing Res Rev. 2024;99:102383. doi:10.1016/j.arr.2024.102383
39. Matsuda T, Hisatsune T. Cholinergic modification of neurogenesis and gliosis improves the memory of abetappswe/psen1de9 alzheimer’s disease model mice fed a high-fat diet. J Alzheimers Dis. 2017;56(1):1–23. doi:10.3233/JAD-160761
40. Hai Y, Ren K, Hou W, et al. Hypoglycemic TCM formulas (Huangqi-Gegen drug pair) have the potential as an Alzheimer’s disease. Phytomedicine. 2024;130:155723. doi:10.1016/j.phymed.2024.155723
41. Tian R, Liu X, Jing L, et al. Huang-Lian-Jie-Du decoction attenuates cognitive dysfunction of rats with type 2 diabetes by regulating autophagy and NLRP3 inflammasome activation. J Ethnopharmacol. 2022;292:115196. doi:10.1016/j.jep.2022.115196
42. Zhang Y, Yin F, Liu J, Liu Z. Geniposide attenuates the phosphorylation of tau protein in cellular and insulin-deficient APP/PS1 transgenic mouse model of alzheimer’s disease. Chem Biol Drug Des. 2016;87(3):409–418. doi:10.1111/cbdd.12673
43. Zhang J, Song J, Zhang J, et al. Effects of berberine on diabetes and cognitive impairment in an animal model: the mechanisms of action. Am J Chin Med. 2021;49(6):1399–1415. doi:10.1142/S0192415X21500658
44. Roy B, Ehlert L, Mullur R, et al. Regional brain gray matter changes in patients with type 2 diabetes mellitus. Sci Rep. 2020;10(1):9925. doi:10.1038/s41598-020-67022-5
45. Callisaya ML, Beare R, Moran C, et al. Type 2 diabetes mellitus, brain atrophy and cognitive decline in older people: a longitudinal study. Diabetologia. 2019;62(3):448–458. doi:10.1007/s00125-018-4778-9
46. Andrew RJ, Kellett KA, Thinakaran G, Hooper NM. A Greek tragedy: the growing complexity of alzheimer amyloid precursor protein proteolysis. J Biol Chem. 2016;291(37):19235–19244. doi:10.1074/jbc.R116.746032
47. Gao Y, Tan L, Yu JT, Tan L. Tau in alzheimer’s disease: mechanisms and therapeutic strategies. Curr Alzheimer Res. 2018;15(3):283–300. doi:10.2174/1567205014666170417111859
48. Goedert M, Spillantini MG. A century of Alzheimer’s disease. Science. 2006;314(5800):777–781. doi:10.1126/science.1132814
49. Avila J, Diaz-Nido J. Tangling with hypothermia. Nat Med. 2004;10(5):460–461. doi:10.1038/nm0504-460
50. Zhao H, Wu C, Zhang X, et al. Insulin resistance is a risk factor for mild cognitive impairment in elderly adults with T2DM. Open Life Sci. 2019;14(1):255–261. doi:10.1515/biol-2019-0029
51. Yin Q, Ma J, Han X, et al. Spatiotemporal variations of vascular endothelial growth factor in the brain of diabetic cognitive impairment. Pharmacol Res. 2021;163:105234. doi:10.1016/j.phrs.2020.105234
52. Ehtewish H, Arredouani A, El-Agnaf O. Diagnostic, prognostic, and mechanistic biomarkers of diabetes mellitus-associated cognitive decline. Int J Mol Sci. 2022;23(11):6144. doi:10.3390/ijms23116144
53. Sandhir R, Gupta S. Molecular and biochemical trajectories from diabetes to Alzheimer’s disease: a critical appraisal. World J Diabetes. 2015;6(12):1223–1242. doi:10.4239/wjd.v6.i12.1223
54. Jackson K, Barisone GA, Diaz E, et al. Amylin deposition in the brain: a second amyloid in Alzheimer disease?. Ann Neurol. 2013;74(4):517–526. doi:10.1002/ana.23956
55. Kellar D, Craft S. Brain insulin resistance in Alzheimer’s disease and related disorders: mechanisms and therapeutic approaches. Lancet Neurol. 2020;19(9):758–766. doi:10.1016/S1474-4422(20)30231-3
56. Najem D, Bamji-Mirza M, Chang N, Liu QY, Zhang W. Insulin resistance, neuroinflammation, and Alzheimer’s disease. Rev Neurosci. 2014;25(4):509–525. doi:10.1515/revneuro-2013-0050
57. Avila J, Leon-Espinosa G, Garcia E, et al. Tau phosphorylation by GSK3 in different conditions. Int J Alzheimers Dis. 2012;2012:578373. doi:10.1155/2012/578373
58. Schubert M, Gautam D, Surjo D, et al. Role for neuronal insulin resistance in neurodegenerative diseases. Proc Natl Acad Sci U S A. 2004;101(9):3100–3105. doi:10.1073/pnas.0308724101
59. Sedzikowska A, Szablewski L. Insulin and Insulin Resistance in Alzheimer’s Disease. Int J Mol Sci. 2021;22(18):9987. doi:10.3390/ijms22189987
60. Mielke JG, Taghibiglou C, Liu L, et al. A biochemical and functional characterization of diet-induced brain insulin resistance. J Neurochem. 2005;93(6):1568–1578. doi:10.1111/j.1471-4159.2005.03155.x
61. Tian Y, Jing G, Zhang M. Insulin-degrading enzyme: roles and pathways in ameliorating cognitive impairment associated with Alzheimer’s disease and diabetes. Ageing Res Rev. 2023;90:101999. doi:10.1016/j.arr.2023.101999
62. Nguyen TT, Ta Q, Nguyen T, Le TT, Vo VG. Role of insulin resistance in the alzheimer’s disease progression. Neurochem Res. 2020;45(7):1481–1491. doi:10.1007/s11064-020-03031-0
63. Dineley KT, Jahrling JB, Denner L. Insulin resistance in Alzheimer’s disease. Neurobiol Dis. 2014;72 Pt A:92–103. doi:10.1016/j.nbd.2014.09.001
64. Lauretti E, Dincer O, Pratico D. Glycogen synthase kinase-3 signaling in Alzheimer’s disease. Biochim Biophys Acta Mol Cell Res. 2020;1867(5):118664. doi:10.1016/j.bbamcr.2020.118664
65. Nasoohi S, Parveen K, Ishrat T. Metabolic syndrome, brain insulin resistance, and alzheimer’s disease: thioredoxin interacting protein (TXNIP) and inflammasome as core amplifiers. J Alzheimers Dis. 2018;66(3):857–885. doi:10.3233/JAD-180735
66. Kurochkin IV, Guarnera E, Berezovsky IN. Insulin-degrading enzyme in the fight against alzheimer’s disease. Trends Pharmacol Sci. 2018;39(1):49–58. doi:10.1016/j.tips.2017.10.008
67. Blomqvist ME, Chalmers K, Andreasen N, et al. Sequence variants of IDE are associated with the extent of beta-amyloid deposition in the Alzheimer’s disease brain. Neurobiol Aging. 2005;26(6):795–802. doi:10.1016/j.neurobiolaging.2004.07.011
68. Leissring MA, Farris W, Chang AY, et al. Enhanced proteolysis of beta-amyloid in APP transgenic mice prevents plaque formation, secondary pathology, and premature death. Neuron. 2003;40(6):1087–1093. doi:10.1016/S0896-6273(03)00787-6
69. Qin W, Zhou A, Zuo X, et al. The study on the role of transcription factor GATA binding protein 3 in familial Alzheimer’s disease pathogenesis. Chin J Neuro. 2011;44(3):159–162.
70. Luchsinger JA, Tang MX, Shea S, Mayeux R. Hyperinsulinemia and risk of Alzheimer disease. Neurology. 2004;63(7):1187–1192. doi:10.1212/01.WNL.0000140292.04932.87
71. Birajdar SV, Mazahir F, Alam MI, Kumar A, Yadav AK. Repurposing and clinical attributes of antidiabetic drugs for the treatment of neurodegenerative disorders. Eur J Pharmacol. 2023;961:176117. doi:10.1016/j.ejphar.2023.176117
72. Mittal K, Mani RJ, Katare DP. Type 3 diabetes: cross talk between differentially regulated proteins of type 2 diabetes mellitus and alzheimer’s disease. Sci Rep. 2016;6:25589. doi:10.1038/srep25589
73. Kim JM, Park CH, Park SK, et al. Ginsenoside re ameliorates brain insulin resistance and cognitive dysfunction in high fat diet-induced C57BL/6 mice. J Agric Food Chem. 2017;65(13):2719–2729. doi:10.1021/acs.jafc.7b00297
74. Kang JY, Park SK, Kim JM, et al. 4,5-dicaffeyolquinic acid improves high-fat diet-induced cognitive dysfunction through the regulation of insulin degrading enzyme. J Food Biochem. 2019;43(7):e12855. doi:10.1111/jfbc.12855
75. Liu J, Zhang Y, Deng X, Yin F. Geniposide decreases the level of Abeta1-42 in the hippocampus of streptozotocin-induced diabetic rats. Acta Biochim Biophys Sin. 2013;45(9):787–791. doi:10.1093/abbs/gmt069
76. Yan T, Nian T, Wu B, et al. Okra polysaccharides can reverse the metabolic disorder induced by high-fat diet and cognitive function injury in Abeta(1-42) mice. Exp Gerontol. 2020;130:110802. doi:10.1016/j.exger.2019.110802
77. Guo X, Lei M, Ma G, et al. Schisandrin A alleviates spatial learning and memory impairment in diabetic rats by inhibiting inflammatory response and through modulation of the PI3K/AKT pathway. Mol Neurobiol. 2024;61(5):2514–2529. doi:10.1007/s12035-023-03725-w
78. Xuan W, Wang H, Zhou P, et al. Berberine ameliorates rats model of combined Alzheimer’s disease and type 2 diabetes mellitus via the suppression of endoplasmic reticulum stress. 3 Biotech. 2020;10(8):359. doi:10.1007/s13205-020-02354-7
79. Chen Q, Mo R, Wu N, et al. Berberine ameliorates diabetes-associated cognitive decline through modulation of aberrant inflammation response and insulin signaling pathway in DM rats. Front Pharmacol. 2017;8:334. doi:10.3389/fphar.2017.00334
80. Meng X, Zhang Y, Li Z, et al. Increasing brain glucose uptake by Gypenoside LXXV ameliorates cognitive deficits in a mouse model of diabetic Alzheimer’s disease. Phytother Res. 2023;37(2):611–626. doi:10.1002/ptr.7639
81. Parida IS, Takasu S, Ito J, Eitsuka T, Nakagawa K. 1-Deoxynojirimycin attenuates pathological markers of Alzheimer’s disease in the in vitro model of neuronal insulin resistance. FASEB J. 2024;38(13):e23800. doi:10.1096/fj.202302600R
82. Wang X, Zhao L. Calycosin ameliorates diabetes-induced cognitive impairments in rats by reducing oxidative stress via the PI3K/Akt/GSK-3beta signaling pathway. Biochem Biophys Res Commun. 2016;473(2):428–434. doi:10.1016/j.bbrc.2016.03.024
83. Keren-Shaul H, Spinrad A, Weiner A, et al. A unique microglia type associated with restricting development of alzheimer’s disease. Cell. 2017;169(7):1276–1290.e17. doi:10.1016/j.cell.2017.05.018
84. Tarasoff-Conway JM, Carare RO, Osorio RS, et al. Clearance systems in the brain-implications for Alzheimer disease. Nat Rev Neurol. 2015;11(8):457–470. doi:10.1038/nrneurol.2015.119
85. De Strooper B, Karran E. The cellular phase of alzheimer’s disease. Cell. 2016;164(4):603–615. doi:10.1016/j.cell.2015.12.056
86. Dani M, Wood M, Mizoguchi R, et al. Microglial activation correlates in vivo with both tau and amyloid in Alzheimer’s disease. Brain. 2018;141(9):2740–2754. doi:10.1093/brain/awy188
87. Morales I, Jimenez JM, Mancilla M, Maccioni RB. Tau oligomers and fibrils induce activation of microglial cells. J Alzheimers Dis. 2013;37(4):849–856. doi:10.3233/JAD-131843
88. Gorlovoy P, Larionov S, Pham TT, Neumann H. Accumulation of tau induced in neurites by microglial proinflammatory mediators. FASEB J. 2009;23(8):2502–2513. doi:10.1096/fj.08-123877
89. Leng F, Edison P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here?. Nat Rev Neurol. 2021;17(3):157–172. doi:10.1038/s41582-020-00435-y
90. Hansen DV, Hanson JE, Sheng M. Microglia in Alzheimer’s disease. J Cell Biol. 2018;217(2):459–472. doi:10.1083/jcb.201709069
91. Walker CD, Risher WC, Risher M. Regulation of synaptic development by astrocyte signaling factors and their emerging roles in substance abuse. Cells. 2020;9(2):297. doi:10.3390/cells9020297
92. Liddelow SA, Guttenplan KA, Clarke LE, et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017;541(7638):481–487. doi:10.1038/nature21029
93. Ferreira ST, Clarke JR, Bomfim TR, De Felice FG. Inflammation, defective insulin signaling, and neuronal dysfunction in Alzheimer’s disease. Alzheimers Dement. 2014;10(1 Suppl):S76–83. doi:10.1016/j.jalz.2013.12.010
94. Ferreira LSS, Fernandes CS, Vieira MNN, De Felice FG. Insulin resistance in alzheimer’s disease. Front Neurosci. 2018;12:830. doi:10.3389/fnins.2018.00830
95. Michailidis M, Moraitou D, Tata DA, et al. Alzheimer’s disease as type 3 diabetes: common pathophysiological mechanisms between alzheimer’s disease and type 2 diabetes. Int J Mol Sci. 2022;23(5):2687. doi:10.3390/ijms23052687
96. Van Dyken P, Lacoste B. Impact of metabolic syndrome on neuroinflammation and the blood-brain barrier. Front Neurosci. 2018;12:930. doi:10.3389/fnins.2018.00930
97. Carranza-Naval MJ, Vargas-Soria M, Hierro-Bujalance C, et al. Alzheimer’s disease and diabetes: role of diet, microbiota and inflammation in preclinical models. Biomolecules. 2021;11(2):262. doi:10.3390/biom11020262
98. Shah K, Desilva S, Abbruscato T. The role of glucose transporters in brain disease: diabetes and Alzheimer’s Disease. Int J Mol Sci. 2012;13(10):12629–12655. doi:10.3390/ijms131012629
99. Liu Z, Kumar M, Kabra A. Cucurbitacin B exerts neuroprotection in a murine Alzheimer’s disease model by modulating oxidative stress, inflammation, and neurotransmitter levels. Front Biosci. 2022;27(2):71. doi:10.31083/j.fbl2702071
100. Gomaa AA, Makboul RM, El-Mokhtar MA, et al. Terpenoid-rich Elettaria cardamomum extract prevents Alzheimer-like alterations induced in diabetic rats via inhibition of GSK3beta activity, oxidative stress and pro-inflammatory cytokines. Cytokine. 2019;113:405–416. doi:10.1016/j.cyto.2018.10.017
101. Tian X, Liu Y, Ren G, et al. Resveratrol limits diabetes-associated cognitive decline in rats by preventing oxidative stress and inflammation and modulating hippocampal structural synaptic plasticity. Brain Res. 2016;1650:1–9. doi:10.1016/j.brainres.2016.08.032
102. Ma X, Sun Z, Han X, et al. Neuroprotective effect of resveratrol via activation of sirt1 signaling in a rat model of combined diabetes and alzheimer’s disease. Front Neurosci. 2019;13:1400. doi:10.3389/fnins.2019.01400
103. Chen X, Famurewa AC, Tang J, Olatunde OO, Olatunji OJ. Hyperoside attenuates neuroinflammation, cognitive impairment and oxidative stress via suppressing TNF-alpha/NF-kappaB/caspase-3 signaling in type 2 diabetes rats. Nutr Neurosci. 2022;25(8):1774–1784. doi:10.1080/1028415X.2021.1901047
104. Park S, Kang S, Kim DS, Moon BR. Agrimonia pilosa Ledeb. Cinnamomum cassia Blume, and Lonicera japonica Thunb. protect against cognitive dysfunction and energy and glucose dysregulation by reducing neuroinflammation and hippocampal insulin resistance in beta-amyloid-infused rats. Nutr Neurosci. 2017;20(2):77–88. doi:10.1080/1028415X.2015.1135572
105. Di Virgilio F, Schmalzing G, Markwardt F. The elusive P2X7 macropore. Trends Cell Biol. 2018;28(5):392–404. doi:10.1016/j.tcb.2018.01.005
106. Munoz FM, Patel PA, Gao X, et al. Reactive oxygen species play a role in P2X7 receptor-mediated IL-6 production in spinal astrocytes. Purinergic Sig. 2020;16(1):97–107. doi:10.1007/s11302-020-09691-5
107. Yang C, Shi S, Su Y, Tong J, Li L. P2X7R promotes angiogenesis and tumour-associated macrophage recruitment by regulating the NF-kappaB signalling pathway in colorectal cancer cells. J Cell Mol Med. 2020;24(18):10830–10841. doi:10.1111/jcmm.15708
108. Gentile D, Lazzerini PE, Gamberucci A, et al. Searching novel therapeutic targets for scleroderma: P2X7-receptor is up-regulated and promotes a fibrogenic phenotype in systemic sclerosis fibroblasts. Front Pharmacol. 2017;8:638. doi:10.3389/fphar.2017.00638
109. Zhang Y, Xu Y, Sun Q, et al. Activation of P2X(7)R- NLRP3 pathway in retinal microglia contribute to retinal ganglion cells death in chronic ocular hypertension (COH). Exp Eye Res. 2019;188:107771. doi:10.1016/j.exer.2019.107771
110. Wu Y, Zhang Y, Zhang J, et al. Cathelicidin aggravates myocardial ischemia/reperfusion injury via activating TLR4 signaling and P2X(7)R/NLRP3 inflammasome. J Mol Cell Cardiol. 2020;139:75–86. doi:10.1016/j.yjmcc.2019.12.011
111. Lu Z, Yao Y, Wang J, Peng J. Dioscin ameliorates diabetes cognitive dysfunction via adjusting P2X7R/NLRP3 signal. Int Immunopharmacol. 2021;101(Pt B):108314. doi:10.1016/j.intimp.2021.108314
112. Dhaliwal J, Dhaliwal N, Akhtar A, Kuhad A, Chopra K. Tetramethylpyrazine attenuates cognitive impairment via suppressing oxidative stress, neuroinflammation, and apoptosis in type 2 diabetic rats. Neurochem Res. 2022;47(8):2431–2444. doi:10.1007/s11064-022-03640-x
113. Lu F, Li X, Li W, et al. Tetramethylpyrazine reverses intracerebroventricular streptozotocin-induced memory deficits by inhibiting GSK-3beta. Acta Biochim Biophys Sin. 2017;49(8):722–728. doi:10.1093/abbs/gmx059
114. Hao X, Yuan J, Dong H. Salidroside prevents diabetes‑induced cognitive impairment via regulating the Rho pathway. Mol Med Rep. 2019;19(1):678–684. doi:10.3892/mmr.2018.9621
115. Bhatia V, Sharma S. Role of mitochondrial dysfunction, oxidative stress and autophagy in progression of Alzheimer’s disease. J Neurol Sci. 2021;421:117253. doi:10.1016/j.jns.2020.117253
116. Lin MT, Beal MF. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature. 2006;443(7113):787–795. doi:10.1038/nature05292
117. Moreira PI, Santos MS, Selca R, Oliveira CR. Brain mitochondrial dysfunction as a link between Alzheimer’s!disease and diabetes. J Neurol Sci. 2007;257(1–2):206–214. doi:10.1016/j.jns.2007.01.017
118. Yaribeygi H, Sathyapalan T, Atkin SL, Sahebkar A. Molecular mechanisms linking oxidative stress and diabetes mellitus. Oxid Med Cell Longev. 2020;2020:8609213. doi:10.1155/2020/8609213
119. Ceriello A, Motz E. Is oxidative stress the pathogenic mechanism underlying insulin resistance, diabetes, and cardiovascular disease? The common soil hypothesis revisited. Arterioscler Thromb Vasc Biol. 2004;24(5):816–823. doi:10.1161/01.ATV.0000122852.22604.78
120. Du H, Meng X, Yao Y, Xu J. The mechanism and efficacy of GLP-1 receptor agonists in the treatment of Alzheimer’s disease. Front Endocrinol. 2022;13:1033479. doi:10.3389/fendo.2022.1033479
121. Carvalho C, Moreira PI. Metabolic defects shared by Alzheimer’s disease and diabetes: a focus on mitochondria. Curr Opin Neurobiol. 2023;79:102694. doi:10.1016/j.conb.2023.102694
122. Akter K, Lanza EA, Martin SA, et al. Diabetes mellitus and Alzheimer’s disease: shared pathology and treatment?. Br J Clin Pharmacol. 2011;71(3):365–376. doi:10.1111/j.1365-2125.2010.03830.x
123. Kroner Z. The relationship between Alzheimer’s disease and diabetes: type 3 diabetes?. Altern Med Rev. 2009;14(4):373–379.
124. Ott A, Stolk RP, van Harskamp F, et al. Diabetes mellitus and the risk of dementia: the Rotterdam Study. Neurology. 1999;53(9):1937–1942. doi:10.1212/WNL.53.9.1937
125. Xu J, Gao H, Zhang L, et al. Melatonin alleviates cognition impairment by antagonizing brain insulin resistance in aged rats fed a high-fat diet. J Pineal Res. 2019;67(2):e12584. doi:10.1111/jpi.12584
126. Tamagno E, Guglielmotto M, Vasciaveo V, Tabaton M. Oxidative stress and beta amyloid in alzheimer’s disease. which comes first: the chicken or the egg?. Antioxidants. 2021;10(9). doi:10.3390/antiox10091479
127. Hernandez-Rodriguez M, Arciniega-Martinez IM, Garcia-Marin ID, Correa-Basurto J, Rosales-Hernandez MC. Chronic administration of scopolamine increased GSK3betaP9, beta secretase, amyloid beta, and oxidative stress in the hippocampus of wistar rats. Mol Neurobiol. 2020;57(9):3979–3988. doi:10.1007/s12035-020-02009-x
128. Yang H, Jin X, Kei LC, Yan SK. Oxidative stress and diabetes mellitus. Clin Chem Lab Med. 2011;49(11):1773–1782. doi:10.1515/cclm.2011.250
129. Pratico D, Sung S. Lipid peroxidation and oxidative imbalance: early functional events in Alzheimer’s disease. J Alzheimers Dis. 2004;6(2):171–175. doi:10.3233/JAD-2004-6209
130. Moreira PI, Santos MS, Moreno AM, Seica R, Oliveira CR. Increased vulnerability of brain mitochondria in diabetic (Goto-Kakizaki) rats with aging and amyloid-beta exposure. Diabetes. 2003;52(6):1449–1456. doi:10.2337/diabetes.52.6.1449
131. Dewanjee S, Chakraborty P, Bhattacharya H, et al. Altered glucose metabolism in Alzheimer’s disease: role of mitochondrial dysfunction and oxidative stress. Free Radic Biol Med. 2022;193(Pt 1):134–157. doi:10.1016/j.freeradbiomed.2022.09.032
132. Ermis M, Ciftci G. Role of curcumin on beta-amyloid protein, tau protein, and biochemical and oxidative changes in streptozotocin-induced diabetic rats. Naunyn Schmiedebergs Arch Pharmacol. 2024;397(12):9833–9844. doi:10.1007/s00210-024-03231-3
133. Veerendra Kumar MH, Gupta YK. Effect of Centella asiatica on cognition and oxidative stress in an intracerebroventricular streptozotocin model of Alzheimer’s disease in rats. Clin Exp Pharmacol Physiol. 2003;30(5–6):336–342. doi:10.1046/j.1440-1681.2003.03842.x
134. Ahmadi M, Rajaei Z, Hadjzadeh MA, Nemati H, Hosseini M. Crocin improves spatial learning and memory deficits in the Morris water maze via attenuating cortical oxidative damage in diabetic rats. Neurosci Lett. 2017;642:1–6. doi:10.1016/j.neulet.2017.01.049
135. Xia S, Xie Z, Qiao Y, et al. Differential effects of quercetin on hippocampus-dependent learning and memory in mice fed with different diets related with oxidative stress. Physiol Behav. 2015;138:325–331. doi:10.1016/j.physbeh.2014.09.008
136. Zhu X, Cheng Y, Lu Q, et al. Enhancement of glyoxalase 1, a polyfunctional defense enzyme, by quercetin in the brain in streptozotocin-induced diabetic rats. Naunyn Schmiedebergs Arch Pharmacol. 2018;391(11):1237–1245. doi:10.1007/s00210-018-1543-z
137. Poorgholam P, Yaghmaei P, Noureddini M, Hajebrahimi Z. Effects of artemisinin and TSP-1-human endometrial-derived stem cells on a streptozocin-induced model of Alzheimer’s disease and diabetes in Wistar rats. Acta Neurobiol Exp. 2021;81(2):141–150. doi:10.21307/ane-2021-013
138. Khan S, Khan HU, Khan FA, et al. Anti-alzheimer and antioxidant effects of nelumbo nucifera l. alkaloids, nuciferine and norcoclaurine in alloxan-induced diabetic albino rats. Pharmaceuticals. 2022;15(10):1205. doi:10.3390/ph15101205
139. Mahmoudi N, Kiasalari Z, Rahmani T, et al. Diosgenin attenuates cognitive impairment in streptozotocin-induced diabetic rats: underlying mechanisms. Neuropsychobiology. 2021;80(1):25–35. doi:10.1159/000507398
140. Wang D, Liu L, Li S, Wang C. Effects of paeoniflorin on neurobehavior, oxidative stress, brain insulin signaling, and synaptic alterations in intracerebroventricular streptozotocin-induced cognitive impairment in mice. Physiol Behav. 2018;191:12–20. doi:10.1016/j.physbeh.2018.03.016
141. Sun X, Li S, Xu L, et al. Paeoniflorin ameliorates cognitive dysfunction via regulating SOCS2/IRS-1 pathway in diabetic rats. Physiol Behav. 2017;174:162–169. doi:10.1016/j.physbeh.2017.03.020
142. Zhang R, Liao W, Wu K, et al. Matrine alleviates spatial learning and memory impairment in diabetic mice by inhibiting endoplasmic reticulum stress and through modulation of PK2/PKRs pathway. Neurochem Int. 2022;154:105289. doi:10.1016/j.neuint.2022.105289
143. Huang Y, Li X, Zhang X, Tang J. Oxymatrine ameliorates memory impairment in diabetic rats by regulating oxidative stress and apoptosis: involvement of NOX2/NOX4. Oxid Med Cell Longev. 2020;2020:3912173. doi:10.1155/2020/3912173
144. Schroter D, Hohn A. Role of advanced glycation end products in carcinogenesis and their therapeutic implications. Curr Pharm Des. 2018;24(44):5245–5251. doi:10.2174/1381612825666190130145549
145. Stirban A, Gawlowski T, Roden M. Vascular effects of advanced glycation endproducts: clinical effects and molecular mechanisms. Mol Metab. 2014;3(2):94–108. doi:10.1016/j.molmet.2013.11.006
146. Sato T, Shimogaito N, Wu X, et al. Toxic advanced glycation end products (TAGE) theory in Alzheimer’s disease. Am J Alzheimers Dis Other Demen. 2006;21(3):197–208. doi:10.1177/1533317506289277
147. Khalid M, Petroianu G, Adem A. Advanced glycation end products and diabetes mellitus: mechanisms and perspectives. Biomolecules. 2022;12(4):542. doi:10.3390/biom12040542
148. Pugazhenthi S, Qin L, Reddy PH. Common neurodegenerative pathways in obesity, diabetes, and Alzheimer’s disease. Biochim Biophys Acta Mol Basis Dis. 2017;1863(5):1037–1045. doi:10.1016/j.bbadis.2016.04.017
149. Fang F, Lue L, Yan S, et al. RAGE-dependent signaling in microglia contributes to neuroinflammation, Abeta accumulation, and impaired learning/memory in a mouse model of Alzheimer’s disease. FASEB J. 2010;24(4):1043–1055. doi:10.1096/fj.09-139634
150. Ramasubbu K, Devi RV. Impairment of insulin signaling pathway PI3K/Akt/mTOR and insulin resistance induced AGEs on diabetes mellitus and neurodegenerative diseases: a perspective review. Mol Cell Biochem. 2023;478(6):1307–1324. doi:10.1007/s11010-022-04587-x
151. Woltjer RL, Maezawa I, Ou JJ, Montine KS, Montine TJ. Advanced glycation endproduct precursor alters intracellular amyloid-beta/A beta PP carboxy-terminal fragment aggregation and cytotoxicity. J Alzheimers Dis. 2003;5(6):467–476. doi:10.3233/JAD-2003-5607
152. Takeuchi M, Yamagishi S. Alternative routes for the formation of glyceraldehyde-derived AGEs (TAGE) in vivo. Med Hypotheses. 2004;63(3):453–455. doi:10.1016/j.mehy.2004.03.005
153. Li WY, Lee CY, Lee KM, et al. Advanced glycation end-product precursor methylglyoxal may lead to development of alzheimer’s disease. Diabetes Metab Syndr Obes. 2022;15:3153–3166. doi:10.2147/DMSO.S382927
154. Gill V, Kumar V, Singh K, Kumar A, Kim JJ. Advanced glycation end products (AGES) may be a striking link between modern diet and health. Biomolecules. 2019;9(12):888. doi:10.3390/biom9120888
155. Wang P, Huang R, Lu S, et al. RAGE and AGEs in mild cognitive impairment of diabetic patients: a cross-sectional study. PLoS One. 2016;11(1):e0145521. doi:10.1371/journal.pone.0145521
156. Schnaider Beeri M, Lotan R, Uribarri J, et al. Higher dietary intake of advanced glycation end products is associated with faster cognitive decline in community-dwelling older adults. Nutrients. 2022;14(7):1468. doi:10.3390/nu14071468
157. Tang Y, Yu C, Wu J, et al. Lychee seed extract protects against neuronal injury and improves cognitive function in rats with type II diabetes mellitus with cognitive impairment. Int J Mol Med. 2018;41(1):251–263. doi:10.3892/ijmm.2017.3245
158. Shahzad M, Shabbir A, Wojcikowski K, Wohlmuth H, Gobe GC. The antioxidant effects of radix Astragali (Astragalus membranaceus and Related Species) in protecting tissues from injury and disease. Curr Drug Targets. 2016;17(12):1331–1340. doi:10.2174/1389450116666150907104742
159. Guo M, Dai Y, Gao J, Chen P. Uncovering the mechanism of astragalus membranaceus in the treatment of diabetic nephropathy based on network pharmacology. J Diabetes Res. 2020;2020:5947304. doi:10.1155/2020/5947304
160. Cao J, Chen Z, Zhu Y, et al. Huangqi-Honghua combination and its main components ameliorate cerebral infarction with Qi deficiency and blood stasis syndrome by antioxidant action in rats. J Ethnopharmacol. 2014;155(2):1053–1060. doi:10.1016/j.jep.2014.05.061
161. An F, Zhao R, Xuan X, et al. Calycosin ameliorates advanced glycation end product-induced neurodegenerative changes in cellular and rat models of diabetes-related Alzheimer’s disease. Chem Biol Interact. 2022;368:110206. doi:10.1016/j.cbi.2022.110206
162. Song J, Long J, Xie L, et al. Applications, phytochemistry, pharmacological effects, pharmacokinetics, toxicity of Scutellaria baicalensis Georgi. and its probably potential therapeutic effects on COVID-19: a review. Chin Med. 2020;15:102. doi:10.1186/s13020-020-00384-0
163. Jeong K, Shin Y, Park S, et al. Ethanol extract of Scutellaria baicalensis Georgi prevents oxidative damage and neuroinflammation and memorial impairments in artificial senescense mice. J Biomed Sci. 2011;18(1):14. doi:10.1186/1423-0127-18-14
164. Zheng Y, Wang C, Liu W, et al. Upregulation of Nrf2 signaling: a key molecular mechanism of Baicalin’s neuroprotective action against diabetes-induced cognitive impairment. Biomed Pharmacother. 2024;174:116579. doi:10.1016/j.biopha.2024.116579
165. Wu N, Liu W, Wang J, et al. Berberine ameliorates neuronal AD-like change via activating Pi3k/PGCepsilon pathway. Biofactors. 2021;47(4):587–599. doi:10.1002/biof.1725
166. Mijnhout GS, Scheltens P, Diamant M, et al. Diabetic encephalopathy: a concept in need of a definition. Diabetologia. 2006;49(6):1447–1448. doi:10.1007/s00125-006-0221-8
167. Moheet A, Mangia S, Seaquist ER. Impact of diabetes on cognitive function and brain structure. Ann N Y Acad Sci. 2015;1353:60–71. doi:10.1111/nyas.12807
168. Chinese Society of Endocrinology. Expert consensus on diabetic cognitive dysfunction. Chin J Diabetes Mellitus. 2021;13(7):678–694.
169. Cheng G, Huang C, Deng H, Wang H. Diabetes as a risk factor for dementia and mild cognitive impairment: a meta-analysis of longitudinal studies. Intern Med J. 2012;42(5):484–491. doi:10.1111/j.1445-5994.2012.02758.x
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.