Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Topical Application of miR-200b-3p by Poloxamer 407-Based Hydrogel Accelerates Diabetic Wound Healing
Authors Lo WY, Sin CH, Wang HJ ![]()
Received 27 May 2025
Accepted for publication 30 September 2025
Published 16 October 2025 Volume 2025:18 Pages 3845—3864
DOI https://doi.org/10.2147/DMSO.S538346
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Rebecca Baqiyyah Conway
Wan-Yu Lo,1 Cian-Huei Sin,1 Huang-Joe Wang2,3
1Department of Food Science and Technology, Hungkuang University, Taichung, Taiwan; 2Department of Medicine, Division of Cardiovascular Medicine, China Medical University Hospital, Taichung, Taiwan; 3School of Medicine, China Medical University, Taichung, Taiwan
Correspondence: Huang-Joe Wang, Department of Medicine, Division of Cardiovascular Medicine, China Medical University Hospital, Taiwan, Email [email protected]
Background: Diabetic foot ulcers (DFUs) are refractory to healing owing to excessive oxidative stress, increased proinflammatory cytokines, accumulation of senescent cells, and impaired angiogenesis. Local injections of miR-146a-5p and miR-200b-3p have pro-healing properties in diabetic wounds but are unrealistic in clinical practice. We investigated whether the topical application of miR-146a-5p and miR-200b-3p using a poloxamer 407-based hydrogel accelerated diabetic wound healing in db/db diabetic mice. In addition, we compared these two miRNAs to select a better microRNA for further application in patients with DFUs.
Methods: The wound was created using an 8-mm punch biopsy. One hundred microliters of poloxamer P407 hydrogel formulations (gel, gel+miR-negative control, gel+miR-146a-5p, and gel+miR-200b-3p) were topically applied to each wound, and the hydrogel was changed every two days. Skin samples were sent for real-time PCR, H&E staining, and immunohistochemical staining.
Results: On the 14th day, the gel+miR-200b-3p group demonstrated the best wound healing performance compared with the other three groups, as supported by the reduced wound size and enhanced granulation tissue thickness. Mechanistic insights showed that gel+miR-200b-3p caused significant upregulation of Col1α 2 gene expression and downregulation of Nox1, Nox4, HO-1, IL-6, IL-1β, OGT, p21, and p53 gene expression. The CD68 protein levels were significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups. Furthermore, CD31 proteins showed a significant increase in the gel+miR-200b-3p group compared with the other three groups. These data demonstrate that the gel+miR-200b-3p group had better pro-healing efficacy than the gel+miR-146a-5p group through its effects on anti-oxidative stress, anti-inflammation, anti-senescence, and pro-angiogenesis.
Conclusion: Topical application of miR-200b-3p by poloxamer 407 hydrogel showed better results in accelerating diabetic wound healing than miR-146a-5p. These data suggest that topical miR-200b-3p formulation is an effective treatment approach for DFUs.Graphical Abstract
Keywords: poloxamer 407 hydrogel, microRNA-146a-5p, microRNA-200b-3p, oxidative stress, inflammation, senescence, angiogenesis
Graphical Abstract:

Introduction
Diabetic foot ulcer (DFU) is a devastating complication, with a lifetime risk of amputation in approximately 20% of patients.1 As a common cause of chronic wounds, only 30–40% of DFU patients achieve complete wound healing by the end of 12 weeks.2 The pathological mechanisms underlying chronic diabetic wounds are complex and include excessive reactive oxidative stress (ROS), increased proinflammatory cytokines, accumulation of senescent cells, and impaired angiogenesis.3
In diabetes, hyperglycemia and the accumulation of advanced glycation end-products (AGE) cause an increase in cellular ROS.4 One of the most important ROS-generating systems is the nicotinamide adenine dinucleotide phosphate oxidase (Nox).5 Among the different subtypes, Nox1, Nox2, and Nox5 produced superoxide, while Nox4 mainly produced hydrogen peroxide.6 Although excessive ROS production in diabetes leads to impaired wound healing, its level is tightly controlled by nuclear factor-erythroid 2 p45-related factor 2 (Nrf2).7 Nrf2 is a transcription factor that is involved in maintaining redox homeostasis by reducing ROS damage, tissue inflammation, and cellular senescence.8 One of the most important regulators of the Nrf2 signaling pathway depends on its repressor Kelch-like ECH-associated protein-1 (Keap-1). ROS disrupts the Nrf2 interaction between Keap-1, and the released Nrf2 is translocated into the nucleus to upregulate multiple antioxidant genes, including NAD(P)H quinone oxidoreductase-1, glutathione peroxidase, superoxide dismutase, catalase, and heme oxygenase-1 (HO-1).7
During wound healing, tissue macrophages first induce intensive inflammation, which is followed by resolution of inflammation to remove tissue debris. M1 macrophages express a pro-inflammatory phenotype by secreting tumor necrosis factor-alpha, interleukin-6 (IL-6), and interleukin-1 beta (IL-1β). Instead, M2 macrophages control the resolution of inflammation by secreting resolutory cytokines interleukin-4 and interlukin-10.9–11 The transition from M1 to M2 macrophages is impaired in diabetic wounds, and sustained inflammasome activity in tissue macrophages impairs diabetic wound healing.12
Cellular senescence is characterized by irreversible cell cycle arrest due to various stressors, including oncogenic signaling, genomic instability, telomere shortening, mitochondrial dysfunction, and excessive ROS.13,14 Hyperglycemia can induce premature senescence in endothelial cells, macrophages, and epithelial cells.15–17 Moreover, receptors for AGEs can mediate endoplasmic reticulum stress, inducing premature senescence in tubular epithelial cells.18 Upon cellular damage, cells activate a DNA damage response to initiate premature cellular senescence and increase the expression of cyclin-dependent kinase inhibitors (CDKI) such as CDKN2B (p15), CDKN2A (p16), and CDKN1A (p21).19 The senescent cells expressed multiple senescence-associated secretory phenotype (SASP) factors, such as proinflammatory cytokines, chemokines, protease, shed cell surface molecules, and matrix metalloproteinases.20,21 When the senescent cells are accumulated in diabetic wounds, the SASP factors drive the persistent low-grade inflammation in skins and impair the wound healing by an increase of inflammatory mediators, a decrease of cell proliferation and migration, and an increase of extracellular matrix proteases.22,23
MicroRNAs (miRs) are important post-transcriptional regulators of diabetes.24 Among variable miRs, the miR-146a-5p acts as an anti-inflammatory miR by targeting IL-1 receptor-associated kinase 1 and TNF receptor-associated factor 6 proteins,25,26 Nox4,27 and IL-6.28 By utilizing its anti-inflammatory properties, the local injection of miR-146a has been shown to improve diabetic wound healing.29 In contrast, the miR-200 family (miR-141, miR-200a, miR-200b, miR-200c, and miR-429) acts as a tumor suppressor in most cancer types to regulate invasiveness, epithelial-mesenchymal transition, and metastasis of cancer cells.30,31 Recently, our laboratory reported that a single local injection of miR-146a-5p and miR-200b-3p could accelerate wound healing in db/db mice.32
The major limitation of local therapy in DFUs is the difficulty in maintaining the treatment drugs for a prolonged period in wound tissues. Patients with DFUs have wound areas far larger than those in the diabetic mouse wound model. Indeed, the local injection method requires multiple needle punctures to deliver sufficient drug to the wound, which can be traumatic, painful, and unrealistic in clinical practice for DFUs. Thus, a suitable topical formulation is required. Poloxamer 407 (trademark Pluronic® F-127)-based hydrogels were initially developed as artificial skin for burns.33 The FDA-approved poloxamer 407 is a nonionic triblock polymer composed of hydrophobic polyoxypropylene and hydrophilic polyoxyethylene. A unique feature of the poloxamer 407 polymer is its thermoresponsive property. A thermoresponsive phase transition (solution to gelation, sol-to-gel) occurs when the temperature exceeds the lower critical solution temperature.34 The in-situ gelling property makes the poloxamer 407 hydrogel a suitable dressing material for topical application to cutaneous wounds by delivering the desired bioactive compounds for a prolonged time.
Currently, no known microRNA formulations have been approved by the FDA for clinical applications in patients with DFUs. In this study, we aimed to investigate the research gap in this study. We hypothesized that topical formulations of miR-146a-5p and miR-200b-3p by poloxamer 407 hydrogel share similar pro-healing properties with the local injection administration method in diabetic wounds. We also compared miR-146a-5p and miR-200b-3p hydrogel formulations to select a better miRNA formulation for further clinical applications.
Materials And Methods
Animal Model
All experiments were approved according to the guidelines of the Institutional Animal Care and Use Committee of Hungkuang University (Approved Number: HK-P-11114). In 2023, Couturier et al published a meta-analysis to examine the different mouse models of diabetes-related ulcers and they reported that the db/db mouse model was consistently associated with the largest delay in wound healing.35 We randomly assigned five db/db mice to different groups in this study because the difference of wound improvement modified by the miR-200b-3p formulation was easily identified with small animal numbers.29,36,37 Twelve-week-old male db/db mice (National Laboratory Animal Center, Taipei, Taiwan) were wounded with an 8-mm biopsy punch (Miltex Inc., York, PA) on their left and right back on the 0th day, as previously described.32 Four groups were investigated: hydrogel alone (gel), hydrogel+miR-negative control (gel+miR NC), hydrogel+miR-146a-5p mimic (gel+miR-146a-5p), and hydrogel+miR-200b-3p mimic (gel+miR-200b-3p). The miR-NC was purchased from the manufacturer, and it was a scrambled control. For effective transfection, miR-NC, miR-146a-5p mimic and miR-200b-3p mimic were mixed with lipofectamineTM 3000 (Thermo Fisher Scientific) immediately before the wounding surgery. Briefly, 1 μL different miRNAs including 200 ng miR-NC, 200 ng miR-200b-3p mimic and 200 ng miR-146a-5p mimic were mixed evenly with an equal volume of lipofectamine to prepare three lipofectamine-miRNA complexes. To improve the solubility of the three lipofectamine-miRNA complexes in sticky poloxamer 407 solutions, we first added 18 μL normal saline to make a total of 20 μL lipofectamine-miRNA complexes diluted solution. Then, the lipofectamine-miRNA complexes were added to poloxamer 407 at a ratio of 1:9 to create different hydrogel formulations in the final concentration of 17% poloxamer 407 hydrogel. Immediately after surgery, 100 ng of miRNA prepared in 100 µL poloxamer 407 hydrogel formulation was topically applied to each wound using a pipette (i.e., 200 μL per mouse in the back of right and left wounds). The wounds were subjected to hydrogel change every two days until the end of the study. All mice were sacrificed on the 14th day by CO2 narcosis and full-thickness skin samples were obtained from the healing tissues for real-time PCR, hematoxylin and eosin (H&E) staining, and immunohistochemical examination. The wound area was calculated using the ImageJ software (NIH, Bethesda, MD, USA), and the wound area (% of wound area on the 0th day) was expressed as a function of time. The granulation tissue thickness was determined using the following equation: granulation tissue thickness (mm) = granulation tissue area / wound width.38
Preparation of Poloxamer 407 Hydrogel
Poloxamer 407 was purchased from Sigma-Aldrich, and the stock solution was prepared using the cold stirring method.33 Briefly, a weighed amount of poloxamer 407 powder was added to ice-cold ddH2O and stirred by using a magnetic bar. The solution was then cooled to 4。C overnight. After complete liquefaction, the stock solution was stored at 4。C until use.
Tilting Method
The gelation time of poloxamer 407 solutions at concentrations of 14, 17, and 20% (w/w) was determined using the tilting method. Briefly, 1 mL of each solution was placed at 37。C on a glass slide tilted at 45°. The solution then flowed slowly on the surface of the glass slide until it completely stopped owing to the gelation.39 The gelation time of individual hydrogel formulations was recorded. The gelation time at different concentrations was also measured after the addition of 1% glycerol. Besides, the effects of lipofectamine and lipofectamine plus miR-200b-3p on the gelation time were also examined.
Real-Time Quantitative PCR
Wound tissues on the 14th day were homogenized in TRIzol (Invitrogen), and total cellular RNAs were isolated and purified following the manufacturer’s protocol. The first-strand cDNA was synthesized by SuperScript® III First-Strand Synthesis SuperMix (Invitrogen, Carlsbad, CA, USA). The mRNA expression levels of the 11 genes of interest in the mouse wound tissues were analyzed using a StepOnePlus Real-Time PCR instrument (Applied Biosystems), as previously described.32 The individual primer sequences and Universal ProbeLibrary probes (Roche) for β-actin, collagen type I alpha 2 (Col1α2), collagen type III alpha 1 (Col3α1), Nox-1, Nox-4, HO-1, IL-6, IL-1β, O-linked N-acetylglucosamine transferase (OGT), CDKN1A (p21) and p53 were provided in Supplemental Data 1. β-actin was used as an internal control.
Immunohistochemistry (IHC) Staining
Wound samples were fixed with 10% formaldehyde and prepared for H&E and IHC staining of paraffin sections. Each group consisted of five db/db mice. IHC staining was performed on 5 μm section using anti‐CD31 primary antibody (Abcam, 1:2000) and anti‐CD68 primary antibody (Abcam, 1:500), as previously described.32
Statistical Analysis
All data are expressed as mean ± standard error of the mean (SEM) of five animals. Data were analyzed using a paired t-test when pairwise comparisons were performed within the same group. Data were analyzed using an unpaired t-test when pairwise comparisons were performed between different groups. Statistical significance was set at p < 0.05.
Results
Effects of Different Additives on Gelation Time of Poloxamer 407 Hydrogel
The phase transition (sol-to-gel) of the poloxamer 407 hydrogel occurred at a physiological temperature of 37。C (Figure 1A). The three different concentrations resulted in gelation time of 90.6, 44.6 and 16.4 seconds at a gel concentration of 14%, 17%, and 20%, respectively. Three different concentrations with 1% glycerol additive caused the gelation time of 101.8, 54.4 and 14.2 seconds at a gel concentration of 14%, 17%, and 20%, respectively. Our data showed that the addition of 1% glycerol did not significantly change the gelation time of the hydrogel at a concentration of 17%. However, the addition of lipofectamineTM 3000 to the gel/glycerol formulation resulted in an increase of the gelation time to 123.0, 71.4 and 36.6 seconds at a gel concentration of 14%, 17%, and 20%, respectively. Further addition of miR-200b-3p to the gel/glycerol/lipofectamineTM 3000 formulation resulted in a minimal change of the gelation time of 123.4, 69.4 and 37.2 seconds at a gel concentration of 14%, 17%, and 20%, respectively. The therapeutic formulation of poloxamer 407 hydrogel at a concentration of 17% was selected for all animal experiments (Figure 1B).
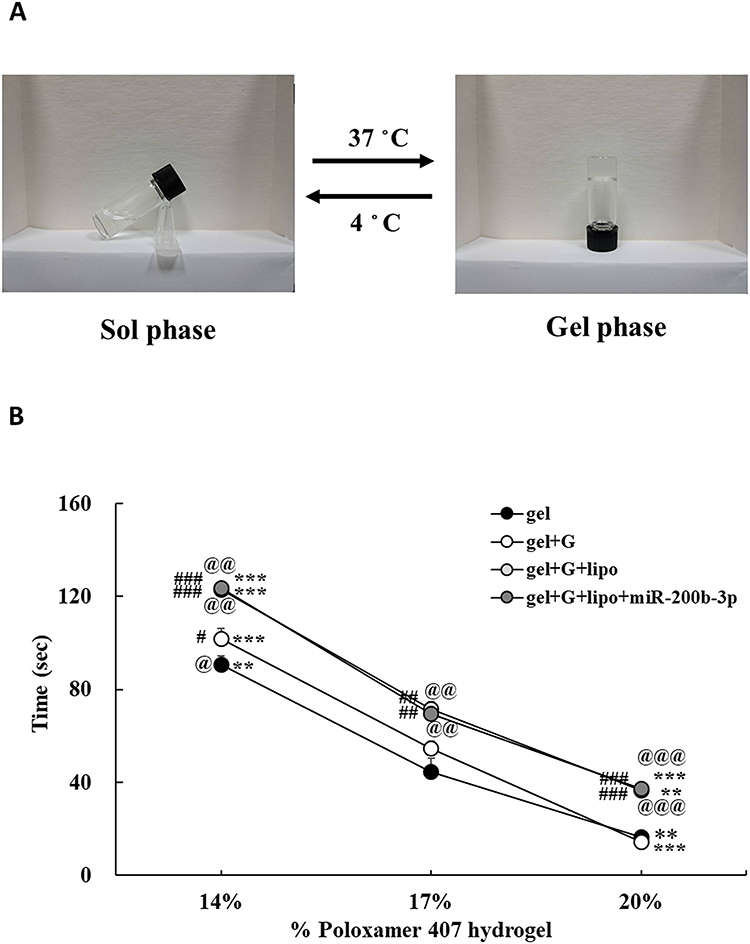 |
Figure 1 Effects of hydrogel concentrations and different additives on gelation time. (A). The thermoresponsive phase transition (sol-to-gel) occurred at a physiological temperature of 37 ◦C. (B). Different concentrations of hydrogel changed the gelation time at 37◦C. The addition of 1% glycerol did not significantly change the gelation time of hydrogel at a concentration of 17% (w/w). The addition of lipofectamine to the gel/glycerol formulation caused a 17.0 second increase of the gelation time at a gel concentration of 17%, as compared with the gel/glycerol formulation. However, further addition of miR-200b-3p to the gel/glycerol/lipofectamine formulation had no discernible effect on the gelation time at different gel concentrations. Data represent mean ± SEM. n=5. **P < 0.01 and ***P < 0.001 vs 17% of their respective groups. #P < 0.05, ##P<0.01, ###P< 0.001 vs gel groups. @P < 0.05, @@P < 0.01, @@@P < 0.001 vs gel+glycerol group. Abbreviations: gel, hydrogel; G, glycerol; lipo, lipofectamine. |
Effects of gel+miR-200b-3p on Body Weight Loss
The relative body weight (%) of the mice in the four treatment groups gradually decreased from the 0th day to 14th day. Although the relative body weight loss between groups on the 14th day was not statistically significant, the relative body weight loss between the 0th day and 14th day was the greatest in the gel group (−10.17%) compared to that in the gel+miR-NC (−8.48%), gel+miR-146a-5p (−8.05%), and gel+miR-200b-3p groups (−6.47%) (Figure 2).
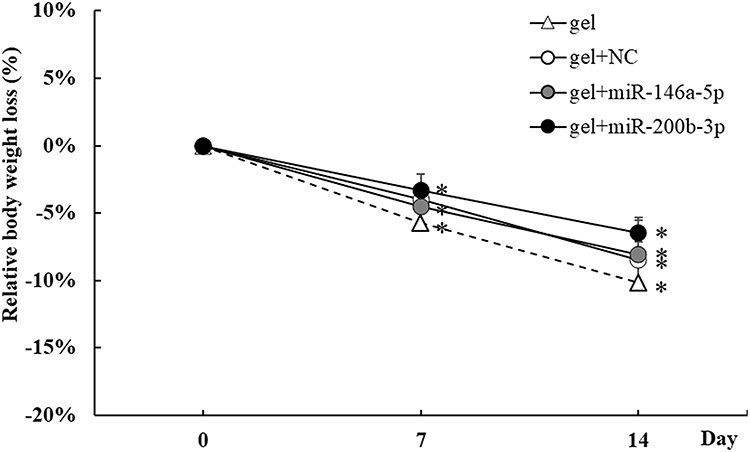 |
Figure 2 Effects of gel+miR-200b-3p on relative body weight. The four treatment groups represented gel, gel+miR-negative control (NC), gel+miR-146a-5p, and gel+miR-200b-3p. The relative body weight loss (% of 0th day) of mice in the gel, gel+miR-146a-5p, and gel+miR-200b-3p groups decreased significantly on day 7 post-wounding compared with that on the 0th day. The relative body weight loss (% of 0th day) of mice in the gel, gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups decreased significantly on the 14th day post-wounding compared with that on the 0th day. The relative body weight loss between groups on the 14th day was not statistically significant. Data are presented as the mean ± SEM. n=5. *P < 0.05 vs 0th day. |
Effects of gel+miR-200b-3p on Wound Size and Granulation Tissue Thickness
Representative wound photographs of the four treatment groups are shown in Figure 3A. The percentages of wound size related to the 0th day are shown in Figure 3B. The relative wound size between the 0th day and 14th day was 67.2%, 51.5%, 32.1%, and 28.5% in the gel, gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively. The wounds healed significantly on the 13th, 8th, 8th, and 6th day in the gel, gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively, compared with that on the 0th day. The statistically significant reduction of relative wound size between gel (67.2%) and gel+miR-200b-3p (28.5%) groups on the 14th day confirmed the efficacy of gel+miR-200b-3p formulation. In addition, the wound size of the gel+miR-200b-3p group consistently reached a significant reduction compared with that of the gel group since the 4th day. The granulation tissue thickness of the gel+miR-200b-3p group significantly increased to 1.267 times in gel+miR-200b-3p group compared with that in the gel group (Figure 3C).
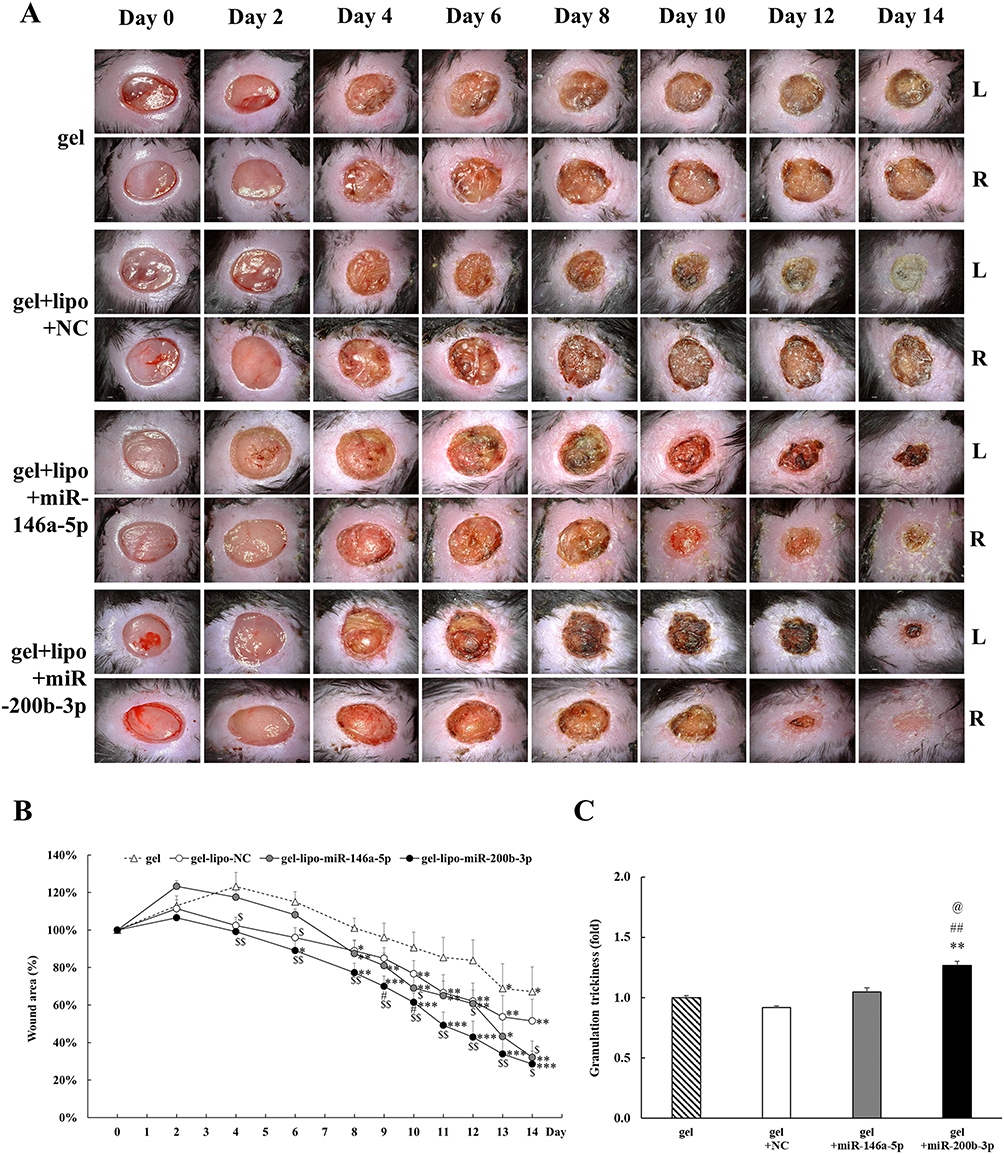 |
Figure 3 Effects of gel+miR-200b-3p on wound healing and granulation tissue thickness. (A). Representative photographs of wounds from four treatment groups were shown. L: left back, R: right back. Scale bar, 2.0 mm. (B). Percentage of wound areas from the four treatment groups compared to that on the 0th day. Gel+miR-200b-5p group caused the most significant wound healing improvement compared to that of the other three groups. Data represent mean ± SEM. n=5. *P<0.05, **P < 0.01, and ***P < 0.001 vs 0th day. $P<0.05, $$P<0.01, and $$$P<0.001 vs gel group. #P<0.05 vs gel+miR-NC group. (C). The granulation tissue thickness of the gel+miR-200b-3p group significantly increased to 1.267 times in gel+miR-200b-3p group compared with that in the gel group on the 14th day. Data represent mean ± SEM. **P<0.01 vs gel group. ##P<0.01 vs gel+miR-NC group. @P<0.05 vs gel+miR-146a-5p group. |
Effects of gel+miR-200b-3p on Col1α2 and Col3α1 Gene Expression
On the 14th day, Col1α2 gene expression increased by 1.319, 1.327 and 1.553 times in the gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively, compared to that in the gel group. In addition, Col1α2 gene expression was significantly increased in the gel+miR-200b-3p group compared with that of the gel group (Figure 4A). Col3α1 gene expression in wound tissues increased by 1.089, 1.077, and 1.223 times in the gel+miR-NC, gel+miR-146a-5p, and miR-miR-200b-3p groups, respectively, compared to that in the gel group. However, no significant differences in Col3α1 expression levels were observed between the groups (Figure 4B).
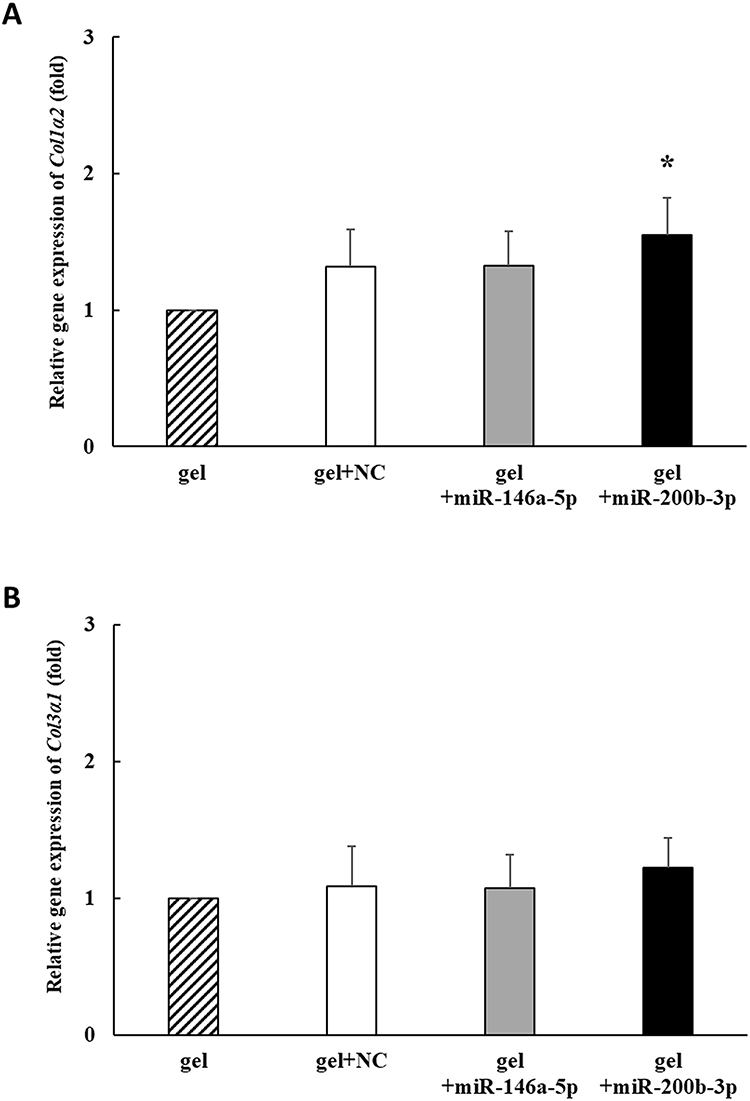 |
Figure 4 Effects of gel+miR-200b-3p on gene expression of Col1α2 and Col3α1. (A). On the 14th day, the gene expression of Col1α2 significantly increased to 1.553 times in gel+miR-200b-3p group compared with that in the gel group. Data represent mean ± SEM. n=5. *P<0.05 vs gel group. (B). On the 14th day, the gene expression of Col3α1 non-significantly increased to 1.223 times in miR-200b-3p group compared with that in the gel group. Data represent mean ± SEM. n=5. |
Effects of gel+miR-200b-3p on Nox-1, Nox-4 and HO-1 Gene Expression
On the 14th day, Nox-1 gene expression decreased by 0.813, 0.516 and 0.283 times in the gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively, compared to that in the gel group. Nox-1 gene expression was significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups compared with that in the gel group. In addition, Nox-1 gene expression was significantly decreased in the gel+miR-200b-3p group compared to that in the gel+miR-NC group (Figure 5A). Nox-4 expression decreased by 0.643 and 0.606 times in the gel+miR-146a-5p and gel+miR-200b-3p groups, respectively, compared to that in the gel group. In addition, Nox-4 gene expression was significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups compared with that in the gel group (Figure 5B). HO-1 gene expression decreased by 0.970, 0.768 and 0.582 times in the gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively, compared to that in the gel group. The HO-1 gene expression was significantly lower in the gel+miR-200b-3p group than the gel group (Figure 5C).
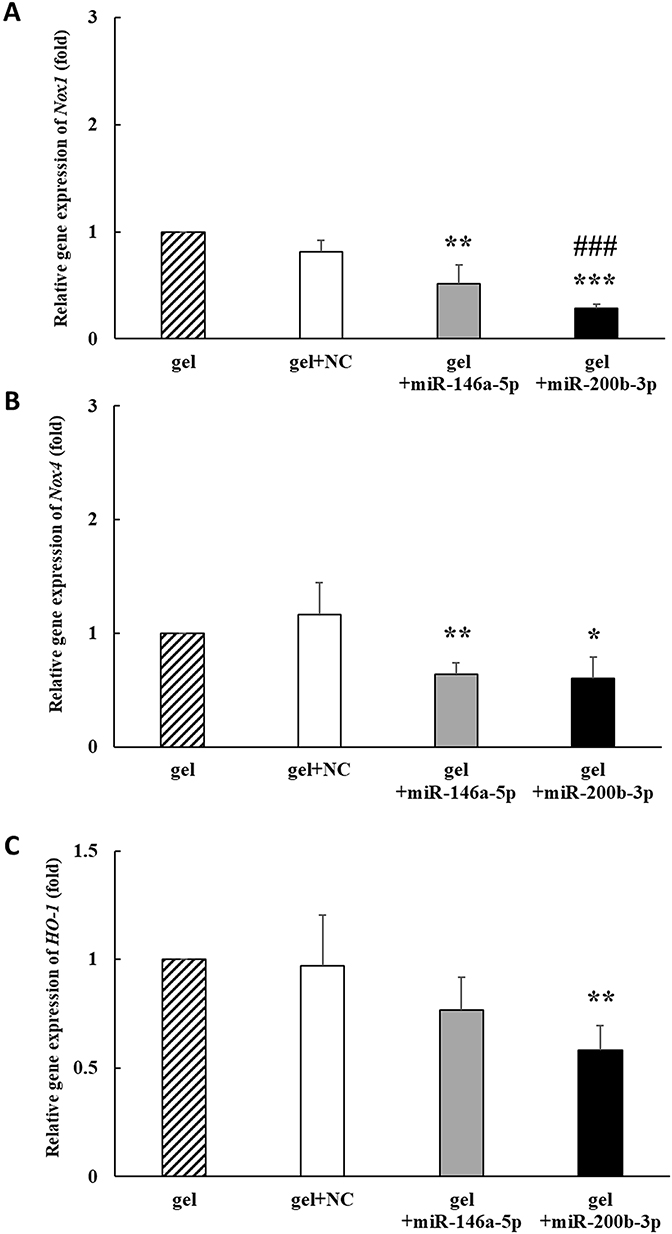 |
Figure 5 Effects of gel+miR-200b-3p on gene expression of Nox1, Nox4, and HO-1. (A). On the 14th day, the Nox1 gene expression in the gel+miR-146a-5p and gel+miR-200b-3p treatment groups significantly decreased to 0.516 and 0.283 times, respectively, compared with that in the gel group. Besides, the Nox1 gene expression in the gel+miR-200b-3 group significantly decreased, as compared with that in the gel+miR-NC group. Data represent mean ± SEM. n=5. **P < 0.01 and ***P < 0.001 vs gel group. ###P < 0.001 vs gel+miR-NC group. (B). On the 14th day, the Nox4 gene expression in the gel+miR-146a-5p and gel+miR-200b-3p treatment groups significantly decreased to 0.643 and 0.606 times compared with that in the gel group. Data represent mean ± SEM. n=5. *P < 0.05 and **P < 0.01 vs gel group. (C). On the 14th day, the HO-1 gene expression in the gel+miR-200b-3p treatment groups significantly decreased to 0.582 times compared with that in the gel group. Data represent mean ± SEM. n=5. **P < 0.01 vs gel group. |
Effects of gel+miR-200b-3p on IL-6, IL-1β and OGT Gene Expression
On the 14th day, IL-6 gene expression decreased by 0.950, 0.402 and 0.255 times in the gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively, compared with that in the gel group. IL-6 gene expression was significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups compared with that in the gel group. In addition, IL-6 gene expression was significantly decreased in the gel+miR-200b-3p group compared to that in the gel+miR-NC group (Figure 6A). IL-1β gene expression decreased by 0.717, 0.500 and 0.142 times in the gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively, compared with that in the gel group. IL-1β gene expression was significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups compared with that in the gel group. In addition, IL-1β gene expression was significantly decreased in the gel+miR-200b-3p group compared to that in the gel+miR-NC and gel+miR-146a-5p groups (Figure 6B). OGT gene expression decreased by 0.725 times in the gel+miR-200b-3p group compared with that in the gel group. In addition, OGT gene expression was significantly lower in the gel+miR-200b-3p group than the other three groups (Figure 6C).
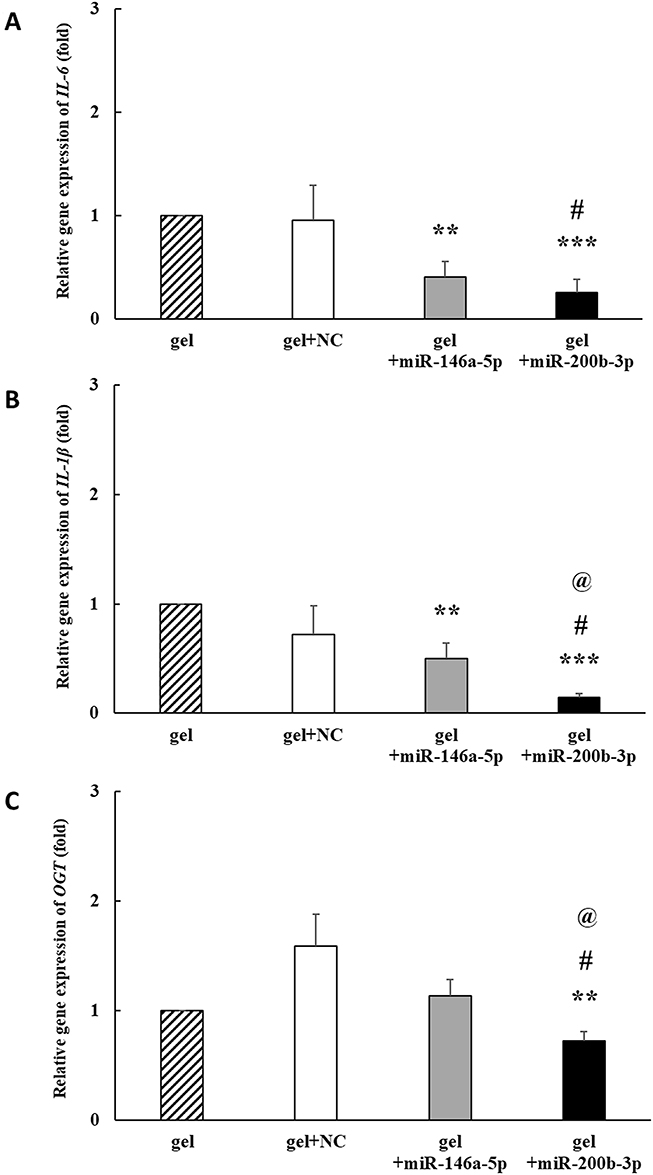 |
Figure 6 Effects of gel+miR-200b-3p on gene expression of IL-6, IL-1β, and OGT. (A). On the 14th day, the IL-6 gene expression in the gel+miR-146a-5p and gel+miR-200b-3p treatment groups significantly decreased to 0.402 and 0.255 times compared with that in the gel group. Besides, the IL-6 gene expression in the gel+miR-200b-3p group significantly decreased, as compared with that in the gel+miR-NC group. Data represent mean ± SEM. n=5. **P< 0.01 and ***P<0.001, vs gel group. #P<0.05 vs gel+miR-NC group. (B) On the 14th day, the IL-1β gene expression in the gel+miR-146a-5p and gel+miR-200b-3p treatment groups significantly decreased to 0.500 and 0.142 times compared with that in the gel group. Besides, the IL-6 gene expression in the gel+miR-200b-3p group significantly decreased, as compared with that in the gel+miR-NC and gel+miR-146a-5p groups. Data represent mean ± SEM. n=5. **P<0.01 and ***P < 0.001 vs gel group. #P<0.05 vs gel+miR-NC group. @P<0.05 vs gel+miR-146a-5p group. (C). On the 14th day, the OGT gene expression in the gel+miR-200b-3p treatment group significantly decreased to 0.725 times compared with that in the gel group. Besides, the OGT gene expression in the gel+miR-200b-3p group significantly decreased, as compared with that in the gel+miR-NC and gel+miR-146a-5p groups. Data represent mean ± SEM. n=5. **P < 0.01 vs gel group. #P < 0.05 vs gel+miR-NC group. @P<0.05 vs gel+miR-146a-5p group. |
Effects of gel+miR-200b-3p on p21 and p53 Gene Expression
The cyclin-dependent kinase inhibitor p21 is a key regulator of cellular senescence in DFUs.40 In addition, the transcription factor p53 is a central tumor suppressor that regulates p21 expression.41 On the 14th day, p21 expression decreased by 0.927-, 0.500 and 0.364 times in the gel+miR-NC, gel+miR-146a-5p, and gel+miR-200b-3p groups, respectively, compared to that in the gel group. The p21 gene expression was significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups, as compared with that in the gel and gel+miR-NC groups. In addition, p21 gene expression was significantly decreased in the gel+miR-200b-3p group compared to that in the gel+miR-146a-5p group (Figure 7A). The p53 gene expression decreased by 0.885 and 0.643 times in the gel+miR-146a-5p and gel+miR-200b-3p groups, respectively, compared with that in the gel group. In addition, p53 gene expression was significantly decreased in the gel+miR-200b-3p group compared to that in the gel and gel+miR-NC groups (Figure 7B).
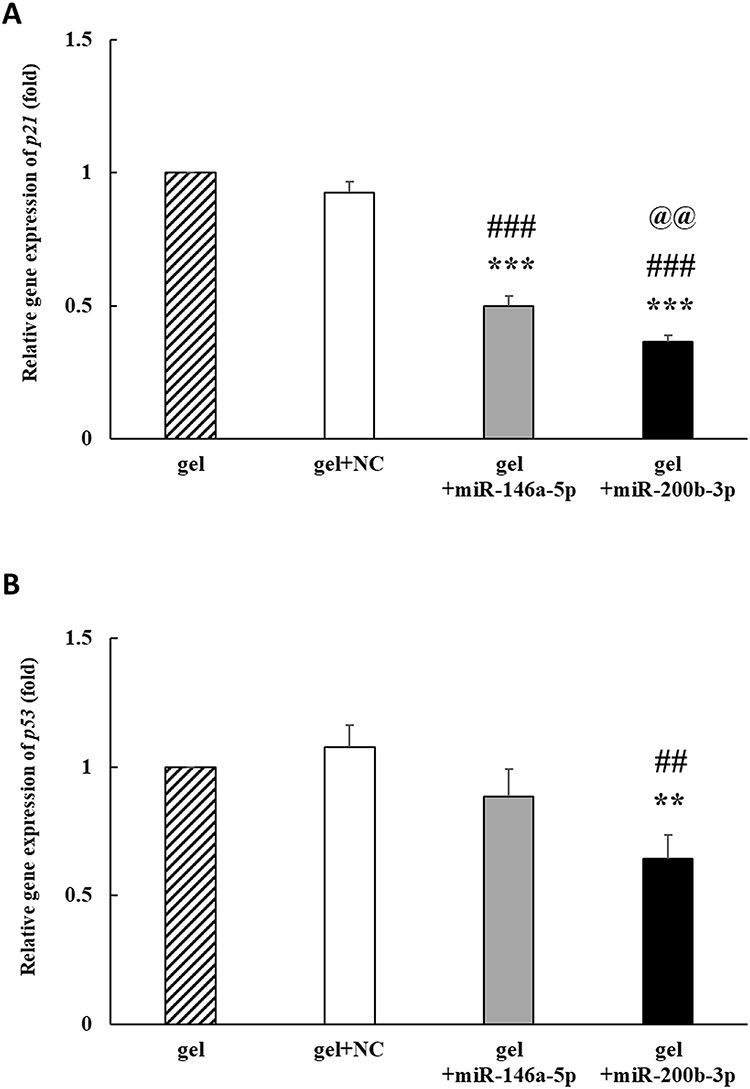 |
Figure 7 Effects of gel+miR-200b-3p on gene expression of p21 and p53. (A). On the 14th day, the p21 gene expression in the gel+miR-146a-5p and gel+miR-200b-3p treatment groups significantly decreased to 0.500 and 0.364 times compared with that in the gel group. Besides, the p21 gene expression in the gel+miR-200b-3p group significantly decreased compared with that in the gel+miR-NC and gel+miR-146a-5p groups. Data represent mean ± SEM. n=5. ***P<0.001 vs gel group. ###P<0.001 vs gel+miR-NC group. @@P<0.01 vs gel+miR-146a-5p group. (B) On the 14th day, the p53 gene expression in the gel+miR-200b-3p treatment groups significantly decreased to 0.643 times compared with that in the gel group. Besides, the p53 gene expression in the gel+miR-200b-3p group significantly decreased, as compared with that in the gel+miR-NC group. Data represent mean ± SEM. n=5. **P<0.01 vs gel group. ##P<0.01 vs gel+miR-NC group. |
Effects of gel+miR-200b-5p on CD68 Protein Expression
IHC staining images of CD68 (tissue macrophage marker) are shown in Figure 8A (brown). On the 14th day, CD68 immunoreactivity decreased by 0.759 and 0.646 times in the gel+miR-146a-5p and gel+miR-200b-3p groups, compared with that in the gel group. In addition, CD68 immunoreactivity was significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups compared to that in the gel and gel+miR-NC groups (Figure 8B).
 |
Figure 8 Effects of gel+miR-200b-3p on protein expression of CD68. (A). On the 14th day, CD68 immunoreactivity (brown) in the four different treatment groups. Magnification 400×. Scale bar, 100 μm. (B). Quantification of CD68 immunoreactivity showed a significant decrease in the miR-146a-5p and miR-200b-3p groups, as compared with that of the gel and gel+miR-NC groups. Data represent mean ± SEM. n=5. **P < 0.01 and ***P<0.001 vs gel group. ###P<0.001 vs gel+miR-NC group. |
Effects of gel+miR-200b-5p on CD31 Protein Expression
IHC staining images of CD31 (endothelial cell marker) are shown in Figure 9A (brown). On the 14th day, CD31 immunoreactivity increased by 1.164 and 1.993 times in the gel+miR-146a-5p and gel+miR-200b-3p groups, respectively, compared with that in the gel group. In addition, CD31 immunoreactivity was significantly increased in the gel+miR-146a-5p and gel+miR-200b-3p groups compared to that in the gel+miR-NC group (Figure 9B). Importantly, miR-200b-3p caused a significant increase in CD31 protein levels compared with the other three groups.
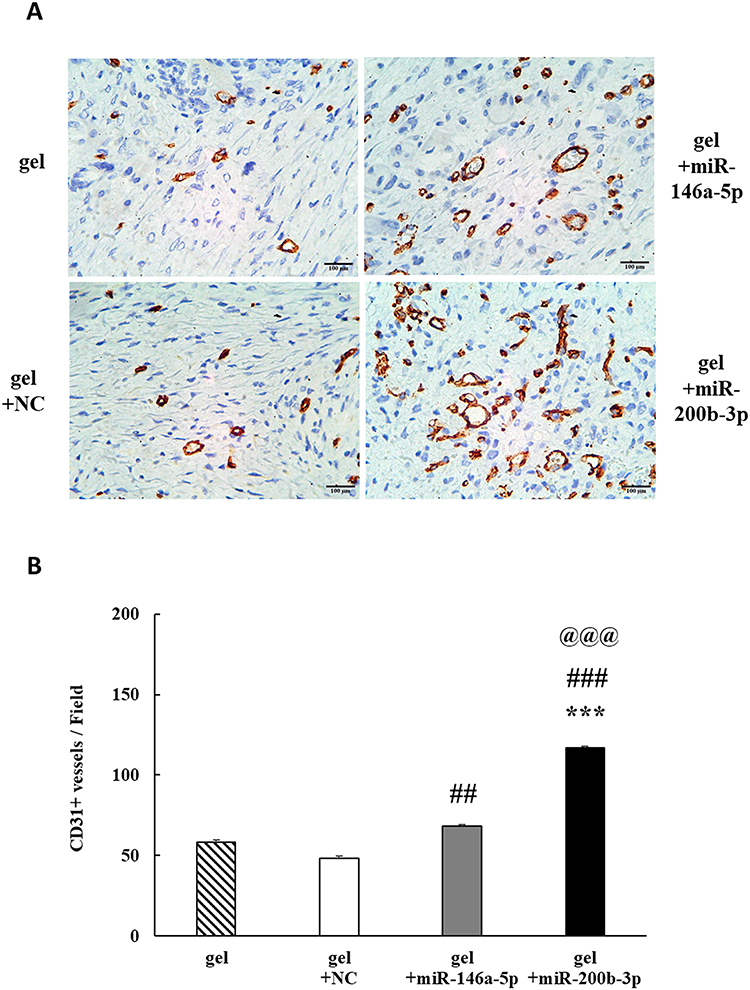 |
Figure 9 Effects of gel+miR-200b-3p on protein expression of CD31. (A). On the 14th day, CD31 immunoreactivity (brown) in the four different treatment groups. Magnification 400×. Scale bar, 100 μm. (B). Quantification of CD31 immunoreactivity showed a significant increase in the miR-146a-5p and miR-200b-3p groups, as compared with that of the gel+miR-NC group. Besides, the CD31 immunoreactivity in the gel+miR-200b-3p group significantly increased, as compared with that in the other three groups. Data represent mean ± SEM. n=5. ***P<0.001 vs gel group. ##P < 0.01 and ###P < 0.001 vs gel+miR-NC group. @@@P < 0.001 vs gel+miR-146a-5p group. |
Discussion
The present study demonstrates that both miR-146a-5p and miR-200b-3p poloxamer P407-based hydrogel formulations can accelerate diabetic wound healing, but the miR-200b-3p hydrogel showed better pro-healing properties than miR-146a-5p hydrogel. The miR-200b-3p hydrogel decreases Nox-1, Nox-4, HO-1, IL-6, IL-1β, OGT, p21 and p53 gene expression in diabetic wounds, findings that represent anti-oxidative stress, anti-inflammation, and anti-senescence as important mechanisms for improving wound healing by miR-200b-3p hydrogel. The miR-200b-3p hydrogel also increases CD31 protein expression in diabetic wound tissues, representing pro-angiogenesis also as a pro-healing characteristic of miR-200b-3p hydrogel. Our animal experiments demonstrate that the topical application of miR-200b-3p hydrogel formulation is a feasible and effective therapeutic adjuvant for treating diabetic wounds.
The concentrations of the poloxamer 407 hydrogel and additives changed the gelation time of the hydrogel. Previous studies have reported that poloxamer 407 hydrogel at a concentration of 15–20% (w/w) is a useful drug carrier in physiological temperature ranges.39,42 In this study, three different concentrations were examined to determine the gelation time. In addition, 1% glycerol was added to the hydrogel to maintain wound moisture and act as a membrane-stabilizing agent.42 To study the effects of transfection reagent lipofectamine and miR-200b-3p on the gelation time, the final therapeutic formulations were also examined. Our data showed that lipofectamine addition to the gel/glycerol formulation caused an increase of the gelation time of 21.2, 17.0 and 11.8 seconds at a gel concentration of 14%, 17%, and 20%, respectively, as compared with the gel/glycerol formulation. Further addition of miR-200b-3p to the gel/glycerol/lipofectamine formulation did not change the gelation time. The final formulation consisted of glycerol, lipofectamine, miR-200b-3p and 14% concentration of poloxamer 407 hydrogel had a gelation time up to 123.4 seconds, a time that was too long to stably keep the microRNA formulation on the wound surface. In contrast, the final formulation of 20% concentration of poloxamer 407 hydrogel had a gelation time of 37.2 seconds, which were relatively too short to adequately handle animal experiments. At a middle 17% concentration of the final formulation of poloxamer 407 hydrogel, the gelation time was 69.4 seconds, an appropriate time that was selected for further animal experiments. Importantly, in vivo the topical administration of poloxamer 407 hydrogel is safe for skin applications,39 and multiple poloxamer 407 hydrogel formulations with various active compounds have been developed for mucosal applications.34
In diabetes, progressive loss of lean body weight is a common problem, and body weight loss worsens further in the presence of wounding. Wound healing is a hypercatabolic process because the inflammatory and proliferative phases of wound healing are highly energy dependent.43 A previous study also showed that poorly healed wounds were associated with significant weight loss in diabetic mice.44 Using a single local injection method in a diabetic wound mouse model, we previously showed that relative body weight loss (% of baseline) on the 14th days was −7.90% and −8.93% in the miR-146a-5p and mR-200b-3p groups, respectively.32 In this study, the relative body weight loss on the 14th day was −8.05% and −6.47% in gel+146a-5p and gel+miR-200b-3p, respectively, suggesting that gel+miR-200b-5p achieved a better protein-energy balance toward an anabolic state than gel+miR-146a-5p.
The cellular uptake of miRNAs is poor for their therapeutic applications. In addition, abundant nucleases in wound tissues further compromise the efficacy of miRNA. The commercial transfection reagent lipofectamine is a catatonic lipid-based reagent by encapsulating miRNAs within the lipid bilayer for efficient cellular uptake.45 Our previous studies, both in vitro and in vivo, also demonstrated the efficacy of miR-146a-5p and miR-200b-3p cellular transfection by lipofectamine.26,27,32,46,47 The release of miRNA from hydrogels follows two phases. The initial rapid phase is caused by hydrogel swelling due to water penetration into the hydrogel. The following slower, sustained phase is caused by the gradual hydrogel degradation.48 Dai et al loaded two antagomiR-467a-3p and antagomiR-874-5p into extracellular vesicles. They reported that 30% poloxamer 407 hydrogel formulation only released around 50% cumulative DiO-labelled extracellular vesicle in 12 days.49 In our study, the miRNAs were pre-complexed with lipofectamine to ensure the miR-146a-5p and miR-200b-3p cellular uptake. The miRNA/lipofectamine complexes were then mixed with 17% poloxamer 407 hydrogel scaffolds to prevent rapid clearance and degradation from diabetic wounds. The wound healing efficacy of this simple formulation preparation is quite impressive.
In a proof-of-concept study, we initiated a single injection of miR-146a-5p and miR-200b-3p around the wound edges on the 0th day. In that study, the wound area was significantly reduction on the 14th day, 13th and 12th in the miR-NC, miR-146a-5p, and miR-200b-3p groups, respectively, compared with that on the 0th day.32 In this study, the treatment frequency was increased to the 0th, 2nd, 4th, 6th, 8th, 10th, and 12th days by the topical application of poloxamer 407 hydrogel. This approach appeared to maintain the integrity and stability of miRNAs in wound tissues, as shown by the wound healing acceleration in the gel+miR-146a-5p and gel+miR-200b-3p groups on the 8th and 6th day, respectively, compared to that on the 0th day. Interestingly, the topical application of gel alone only reached statistically significant healing on the 13th day compared to that on the 0th day, suggesting that gel alone as a wound dressing material is not effective for wound healing improvement and that active compounds such as miR-146a-5p or miR-200b-3p are needed. The early improvement and sustainability of wound healing acceleration in the gel+miR-200b-5p group were further supported by its consistently reduced wound area compared to the gel group after the 4th day, when only two treatments of gel+miR-200b-3p were administered. Compared to the single injection study with a wound area reduction of 68.4% (% of 0th day) in the miR-200b-3p group on the 14th day,32 the benefit of treatment frequency increase was further supported by the reduction of wound area to 28.5% (% of 0th day) in the gel+miR-200b-3p group on the same 14th day in this study. In addition, granulation tissue thickness of the gel+miR-200b-3p group was also significantly increased, as compared with that of the gel+miR-146a-5p group on the 14th day. These data suggest that the topical application of gel+miR-200b-3p achieves better wound healing performance than gel+miR-146a-5p in diabetic wounds, and the treatment frequency is an important factor in promoting wound healing.
Collagen types I and III are the main types of collagens present in the extracellular matrix. Collagen type I is a stiff fibrillar protein that provides tensile strength to the skin, whereas collagen type III produces elastic strength.50 After the inflammation phase wanes out, fibroblasts produce a large amount of collagen type III in the proliferative phase, and the nascent collagen type III is replaced by collagen type I in the remodeling phase to increase wound strength.51 In this study, Col1α2 gene expression on the 14th day was significantly increased by 1.553 times in the gel+miR-200b-3p group compared to the gel group. These data suggested that the remodeling phase of wound healing was more effective in the gel+miR-200b-3p group than the other three groups.
The excessive Nox1 and Nox4 expressions have been shown to play a detrimental role in diabetic complications in several literatures. Zhu et al reported that Nox1-driven ROS activated protein kinase C by modulating the p38/p27 signaling pathway and causing deterioration of diabetic nephropathy.52 Harb et al reported that alagebrium (i.e., an AGE-crosslink breaker) treatment reduced Nox1 expression and accelerated diabetic wound healing.53 Sangartit et al reported that high-fat diet-induced type 2 diabetic mice exhibited an increase in Nox4 expression and tetrahydrocurcumin (i.e., a major metabolite of curcumin) treatment suppressed Nox4 expression to attenuate the severity of diabetic nephropathy.54 We also reported that high-glucose/thrombin-stimulated human aortic endothelial cells (HAEC) exhibited a significant increase in Nox4 expression, and miR-146a-5p attenuated the high glucose/thrombin-induced upregulation of Nox4 expression, ROS generation, and inflammatory phenotypes.27 In this study, the gene expression of Nox-1 and Nox-4 in wound tissues was significantly decreased in the gel+miR-146a-5p and gel+miR-200b-3p groups. However, we observed a greater reduction in the gene expression of Nox-1 and Nox-4 in the miR-200b-5p group than that in the miR-146a-5p group. The reduced Nox-1 and Nox-4 driven ROS production in the miR-200b-3p group was further supported by lower antioxidant HO-1 gene production to prevent excessive antioxidant activity. These data suggest that the efficacy of gel+miR-200b-5p in wound healing is partially attributable to the stronger anti-ROS ability of gel+miR-200b-3p than that of gel+miR-146a-5p.
IL-6 and IL-1β are important inflammatory mediators that are involved in acute wound healing.11 In this study, the gel+miR-200b-3p group showed a more significant downregulation of IL-6 and IL-1β gene expression than the gel+miR-146a-5p group. Although both the gel+miR-146a-5p and gel+miR-200b-3p groups showed a significant decrease in CD68 proteins compared to the gel and gel+miR-NC groups, the relative reduction in CD68 proteins was more obvious in the gel+miR-200b-3p group (64.6% of gel group) than the miR-146a-5p group (75.9% of gel group). These data demonstrated the advantage of gel+miR-200b-3p over gel+miR-146a-5p in decreasing tissue macrophages and pro-inflammatory cytokines in diabetic wound tissues.
O-GlcNAcylation is an important post-translational modification process that regulates the activity of cellular protein activity.55 The tissue levels of O-GlcNAcylation modifications are tightly controlled by OGT56 and an increase in cellular O-GlcNAcylation levels has been implicated in the development of cardiovascular complications in diabetes.55,57 We previously reported that both miR-146a-5p and miR-200b-3p groups act as important post-transcriptional regulators of OGT to exert the anti-inflammatory effect on endothelial cells of high-glucose-stimulated HAECs and db/db mice.46,47 The current study showed a consistently larger reduction in IL-6, IL-1β, and OGT gene expression in the gel+miR-200b-3p group than the gel+miR-146a-5p group, suggesting that the anti-inflammatory effect of gel+miR-200b-3p is partially attributed to a significant downregulation of OGT expression in diabetic wound tissues.
When transiently present, early cellular senescence is crucial for normal wound healing response by regulating tissue inflammation, fibrosis, extracellular matrix deposition, and epithelization for optimal wound healing.58,59 However, persistent chronic inflammation increases premature cellular senescence in diabetes, and SASP factors further drive inflammation aggravation to impair wound healing.22,23 In addition, the paracrine effect of SASP factors can cause a small number of primary senescent cells in diabetes to induce cellular senescence in anatomically distant sites and cause an increase in the number of senescent cells, which further contributes to wound healing impairment.60 Wilkinson et al reported that an increase in senescent cells was associated with p21 upregulation in diabetic mice compared to young wild-type mice in an acute wound healing model.61 Recently, Yu et al reported that skin biopsies of DFUs increased the expression of CDKN1A (p21), CXCL8, IGFBP2, IL1A, MMP10, SERPINE1, and TGFA compared with that of uninvolved diabetic foot skin biopsies and they concluded that p21 is a key senescent regulator in DFUs.40 These findings suggest that p21 may be a potential therapeutic target in diabetic wounds. In this study, both gel+miR-200b-3p and gel+miR-146a-5p significantly downregulated the gene expression of p21 compared to gel and gel+miR-NC groups. Notably, the present study demonstrated the superiority of gel+miR-200b-3p over gel+miR-146a-5p in decreasing the expression of senescent marker p21 in diabetic wound tissues.
Both retinoblastoma protein (RB) and transcription factor p53 are key tumor suppressors. Following the activation of p53 by a DNA damage response, p21 expression was upregulated by p53 activation. High levels of p21 can cause an increase in RB-E2F transcription factor complex formation, resulting in the transcriptional repression of many cell cycle genes to arrest the cell cycle.41 In this study, p53 expression was significantly lower in the gel+miR-200b-3p group than that in the gel and gel+miR-NC groups. These data suggest that miR-200b-3p possesses a more potent anti-senescent effect than miR-146a-5p and this anti-senescent effect is p53-dependent.
For the wound to heal, adequately controlled angiogenesis is necessary to provide new blood vessels within the granulation tissues. However, angiogenesis is impaired in diabetic wounds owing to endothelial dysfunction.62 In addition, recruitment of endothelial progenitor cells from bone marrow into the diabetic wounds are impaired.10,63 In a study using micro-computed tomography imaging technology, the vessel surface area, branch junction number, total vessel length, and total branch number of diabetic mice were significantly lower than those of wild-type mice.64 Type I collagen has been shown to stimulate angiogenesis by ligation of endothelial cell surface α1β2/α2β1 integrin receptors by its GFPGER (502–507) sequence in the type I collagen.65 In this study, the superiority of the proangiogenic effect of the miR-200b-3p group was consistent with its greater increase in Col1α2 gene expression than that of the other groups. Similar to our previous study with a single local injection treatment,32 the current study demonstrated that multiple topical applications of gel+miR-146a-5p and gel+miR-200b-3p formulations increased angiogenesis in diabetic wounds. Crucially, the pro-angiogenic effect of gel+miR-200b-3p was superior to that of gel+miR-146a-3p. The active component of the FDA-approved RegranexTM gel in 1997 was composed of 0.01% human recombinant platelet-derived growth factor-BB (PDGF-BB), which promotes angiogenesis as a pro-healing mechanism.66 However, the pharmacological concentration of RegranexTM (15 g/tube) is at least > 1000‑times the physiological PDGF‑BB level, which once caused a warning label of an increased risk of cancer mortality in patients who require treatment with ≥ 3 tubes of RegranexTM.67 In contrast to the potential tumorigenic potential of RegranexTM, the role of miR-200b-3p as a tumor suppressor is clear.31 The multilevel efficacy of miR-200b-3p against impaired diabetic wound healing suggests that it is an ideal adjuvant therapy for patients with DFUs.
Recent developments have focused on integrating various miRNAs into different gel dressings for wound healing application. Sener et al conjugated miR-146a with cerium oxide nanoparticles (CNP) and incorporated the miR-146a/CNPs into the zwitterionic cryogel. Their gel formulation promoted diabetic wound healing and reduced wound inflammation in db/db mice.68 Wei et al prepared small miR-17-5p-engineered extracellular vesicles (sEVs) and incorporated the sEVs into the gelatin methacryloyl hydrogel. Their hydrogel formulation accelerated diabetic wound healing and promoted collagen deposition and angiogenesis by targeting p21 and phosphatase and tension homolog (PTEN) in db/db mice.69 Like our study by utilizing poloxamer 407 hydrogel as a carrier dressing, other investigators utilized it as a microRNA carrier for promoting wound healing. Li et al reported that liposome-formulated miR-132 mimics mixed with Pluronic® F-127 hydrogel (i.e., trade name of poloxamer 407) improved re-epithelialization of human ex vivo skin wounds.70 Lou et al reported that miR-155-5p antagomir mixed with Pluronic® F-127 hydrogel promoted diabetic wound healing and angiogenesis by enhancing growth differentiation factor 6 in db/db mice.71 In this study, we provided the first evidence that miR-200b-3p hydrogel formulation can accelerate diabetic wound healing through multiple mechanisms. The ease of preparation favors this kind of topical therapy as an adjuvant strategy for patients with DFUs.
Current standard therapies of DFUs include surgical debridement, traditional inert dressings, wound decompression, management of peripheral arterial vascular diseases, intensive glycemic control and infection treatments.72 Traditional dressing such as dry gauze, cotton pads and bandages are effective to prevent wound contamination and absorb exudate. However, they do not accelerate wound healing and tend to cause secondary damage to the healing granulation tissues when the dressing is replaced.73 Our poloxamer 407-based hydrogel is a thermoresponsive hydrogel. The hydrogel in the solution phase can completely cover the uneven surface of diabetic ulcers, even in a deep lesion cracking into the tendon or capsule. The hydrogel in the gelation phase can create a physiological barrier to provide excellent insulation ability. Besides, the addition of 1% glycerol helps to increase moisturizing ability. Most importantly, the addition of miR-200b-3p as an active component of composite hydrogel dressing demonstrates its superiority in promoting wound healing. When the dressing is replaced, the cold saline washing can help poloxamer 407 hydrogel removal to prevent secondary damage. Based on the current study, we recommend that our miR-200b-3p composite hydrogel dressing is suitable in the management for DFU patients with Wagner classification of grade 1 and 2.74 The incorporation of different microRNAs into composite dressing is in the early stage. However, the excellent potential in accelerating diabetic wound healing in a relatively short time by our miR-200b-3p formulation points to an important implication in clinical practice. Specifically, the topical application of easily prepared miR-200b-3p formulation in patients with DFUs can transform unhealed chronic wounds into healed wounds and lessen the further amputation risks and associated high medical costs. Further clinical investigations are necessary to validate miR-200b-3p-based therapeutic applications.
In conclusion, the topical application of the miR-200b-3p poloxamer 407-based hydrogel shows a more promising result in accelerating diabetic wound healing than the miR-146a-5p hydrogel. These therapeutic differences are attributed to multiple pro-healing properties, including anti-oxidative stress, anti-inflammation, anti-senescence, and pro-angiogenesis. Some of major limitations of the current study deserved mention. First, the db/db mouse is a small animal model of type 2 diabetic mouse and it’s not clear whether the same miR-200b-3p hydrogel formulation is effective in the type 1 diabetes. Second, we do not evaluate our hydrogel formulation in the infected wounds. The small db/db mouse wounds are relatively clean, which are completely different to many wounds of DFUs, a clinical scenario that is frequently infected with multiple bacterial pathogens. Therefore, adequate debridement before application of miR-200b-3p hydrogel formulation is necessary to avoid the detrimental effects of infections. Third, the off-target effects of microRNA therapeutics potentially exist. Although we did not examine the inner organs, Li et al reported that local injections with miR-132 mimics resulted in a significant elevation of miR-132 mimics in the skin wounds but no discernible elevation in the inner organs such as liver, spleen, lungs and kidneys in db/db mice.70 The more body weight gain and the better pro-healing properties of the miR-200b-3p group compared to the gel group suggested a low systemic toxicity of the current hydrogel formulation. Our data suggest that the topical application of the miR-200b-3p formulation with poloxamer 407 hydrogel is a feasible and effective treatment approach for patients with DFUs.
Acknowledgments
Funding was provided by China Medical University Hospital (DMR-114-076) and the National Science and Technology Council (NSTC 114-2314-B-039-072).
Disclosure
The authors report no conflicts of interest in this work.
References
1. McDermott K, Fang M, Boulton AJM, Selvin E, Hicks CW. Etiology, epidemiology, and disparities in the burden of diabetic foot ulcers. Diabetes Care. 2023;46(1):209–221. doi:10.2337/dci22-0043
2. Armstrong DG, Tan TW, Boulton AJM, Bus SA. Diabetic foot ulcers: a review. JAMA. 2023;330(1):62–75. doi:10.1001/jama.2023.10578
3. Frykberg RG, Banks J. Challenges in the treatment of chronic wounds. Adv Wound Care. 2015;4(9):560–582. doi:10.1089/wound.2015.0635
4. Paneni F, Beckman JA, Creager MA, Cosentino F. Diabetes and vascular disease: pathophysiology, clinical consequences, and medical therapy: part I. Eur Heart J. 2013;34(31):2436–2443. doi:10.1093/eurheartj/eht149
5. Sena CM, Pereira AM, Seica R. Endothelial dysfunction - a major mediator of diabetic vascular disease. Biochim Biophys Acta. 2013;1832(12):2216–2231. doi:10.1016/j.bbadis.2013.08.006
6. Sahoo S, Meijles DN, Pagano PJ. NADPH oxidases: key modulators in aging and age-related cardiovascular diseases? Clin Sci. 2016;130(5):317–335. doi:10.1042/CS20150087
7. Suntar I, Cetinkaya S, Panieri E, et al. Regulatory role of Nrf2 signaling pathway in wound healing process. Molecules. 2021;26(9):2424. doi:10.3390/molecules26092424
8. O’Connell MA, Hayes JD. The Keap1/Nrf2 pathway in health and disease: from the bench to the clinic. Biochem Soc Trans. 2015;43(4):687–689. doi:10.1042/BST20150069
9. Qing C. The molecular biology in wound healing & non-healing wound. Chin J Traumatol. 2017;20(4):189–193. doi:10.1016/j.cjtee.2017.06.001
10. Eming SA, Martin P, Tomic-Canic M. Wound repair and regeneration: mechanisms, signaling, and translation. Sci Transl Med. 2014;6(265):265sr6. doi:10.1126/scitranslmed.3009337
11. Wilkinson HN, Hardman MJ. Wound healing: cellular mechanisms and pathological outcomes. Open Biol. 2020;10(9):200223. doi:10.1098/rsob.200223
12. Mirza RE, Fang MM, Weinheimer-Haus EM, Ennis WJ, Koh TJ. Sustained inflammasome activity in macrophages impairs wound healing in type 2 diabetic humans and mice. Diabetes. 2014;63(3):1103–1114. doi:10.2337/db13-0927
13. Kuehnemann C, Wiley CD. Senescent cells at the crossroads of aging, disease, and tissue homeostasis. Aging Cell. 2024;23(1):e13988. doi:10.1111/acel.13988
14. Noor SNM, Musa M, Azlina A, Gan SH, Thirumulu KP. Polyphenols in bee products and prevention of cell senescence. Biomedicine. 2024;14(3):1–11. doi:10.37796/2211-8039.1458
15. Wan Y, Liu Z, Wu A, et al. Hyperglycemia promotes endothelial cell senescence through AQR/PLAU signaling axis. Int J Mol Sci. 2022;23(5):2879. doi:10.3390/ijms23052879
16. Prattichizzo F, De Nigris V, Mancuso E, et al. Short-term sustained hyperglycaemia fosters an archetypal senescence-associated secretory phenotype in endothelial cells and macrophages. Redox Biol. 2018;15:170–181. doi:10.1016/j.redox.2017.12.001
17. Kitada K, Nakano D, Ohsaki H, et al. Hyperglycemia causes cellular senescence via a SGLT2- and p21-dependent pathway in proximal tubules in the early stage of diabetic nephropathy. J Diabetes Complications. 2014;28(5):604–611. doi:10.1016/j.jdiacomp.2014.05.010
18. Liu J, Huang K, Cai GY, et al. Receptor for advanced glycation end-products promotes premature senescence of proximal tubular epithelial cells via activation of endoplasmic reticulum stress-dependent p21 signaling. Cell Signal. 2014;26(1):110–121. doi:10.1016/j.cellsig.2013.10.002
19. Ogrodnik M, Acosta JC, Adams PD, et al. Guidelines for minimal information on cellular senescence experimentation in vivo. Cell. 2024;187(16):4150–4175. doi:10.1016/j.cell.2024.05.059
20. Tavenier J, Nehlin JO, Houlind MB, et al. Fisetin as a senotherapeutic agent: evidence and perspectives for age-related diseases. Mech Ageing Dev. 2024;222:111995. doi:10.1016/j.mad.2024.111995
21. Coppe JP, Patil CK, Rodier F, et al. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008;6(12):2853–2868. doi:10.1371/journal.pbio.0060301
22. Wei X, Li M, Zheng Z, et al. Senescence in chronic wounds and potential targeted therapies. Burns Trauma. 2022;10:tkab045. doi:10.1093/burnst/tkab045
23. Pilkington SM, Bulfone-Paus S, Griffiths CEM, Watson REB. Inflammaging and the Skin. J Invest Dermatol. 2021;141(4S):1087–1095. doi:10.1016/j.jid.2020.11.006
24. Shantikumar S, Caporali A, Emanueli C. Role of microRNAs in diabetes and its cardiovascular complications. Cardiovasc Res. 2012;93(4):583–593. doi:10.1093/cvr/cvr300
25. Taganov KD, Boldin MP, Chang KJ, Baltimore D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A. 2006;103(33):12481–12486. doi:10.1073/pnas.0605298103
26. Lo WY, Peng CT, Wang HJ. MicroRNA-146a-5p mediates high glucose-induced endothelial inflammation via targeting interleukin-1 receptor-associated kinase 1 expression. Front Physiol. 2017;8:551. doi:10.3389/fphys.2017.00551
27. Wang HJ, Huang YL, Shih YY, Wu HY, Peng CT, Lo WY. MicroRNA-146a decreases high glucose/thrombin-induced endothelial inflammation by inhibiting NAPDH oxidase 4 expression. Mediators Inflamm. 2014;2014:379537. doi:10.1155/2014/379537
28. Wang HJ, Lo WY, Lin LJ. Angiotensin-(1-7) decreases glycated albumin-induced endothelial interleukin-6 expression via modulation of miR-146a. Biochem Biophys Res Commun. 2013;430(3):1157–1163. doi:10.1016/j.bbrc.2012.12.018
29. Zgheib C, Hilton SA, Dewberry LC, et al. Use of cerium oxide nanoparticles conjugated with MicroRNA-146a to correct the diabetic wound healing impairment. J Am Coll Surg. 2019;228(1):107–115. doi:10.1016/j.jamcollsurg.2018.09.017
30. Sundararajan V, Burk UC, Bajdak-Rusinek K. Revisiting the miR-200 family: a clan of five siblings with essential roles in development and disease. Biomolecules. 2022;12(6):781. doi:10.3390/biom12060781
31. Klicka K, Grzywa TM, Mielniczuk A, Klinke A, Wlodarski PK. The role of miR-200 family in the regulation of hallmarks of cancer. Front Oncol. 2022;12:965231. doi:10.3389/fonc.2022.965231
32. Wang HJ, Sin CH, Yang SH, Hsueh HM, Lo WY. miR-200b-3p accelerates diabetic wound healing through anti-inflammatory and pro-angiogenic effects. Biochem Biophys Res Commun. 2024;731:150388.
33. Schmolka IR. Artificial skin. I. Preparation and properties of pluronic F-127 gels for treatment of burns. J Biomed Mater Res. 1972;6(6):571–582. doi:10.1002/jbm.820060609
34. Giuliano E, Paolino D, Fresta M, Cosco D. Mucosal applications of poloxamer 407-based hydrogels: an overview. Pharmaceutics. 2018;10(3):159.
35. Couturier A, Calissi C, Cracowski JL, Sigaudo-Roussel D, Khouri C, Roustit M. Mouse models of diabetes-related ulcers: a systematic review and network meta-analysis. EBioMedicine. 2023;98:104856. doi:10.1016/j.ebiom.2023.104856
36. Vagesjo E, Ohnstedt E, Mortier A, et al. Accelerated wound healing in mice by on-site production and delivery of CXCL12 by transformed lactic acid bacteria. Proc Natl Acad Sci U S A. 2018;115(8):1895–1900. doi:10.1073/pnas.1716580115
37. Wei Q, Wang Y, Ma K, et al. Extracellular vesicles from human umbilical cord mesenchymal stem cells facilitate diabetic wound healing through MiR-17-5p-mediated enhancement of angiogenesis. Stem Cell Rev Rep. 2022;18(3):1025–1040. doi:10.1007/s12015-021-10176-0
38. Gallant-Behm CL, Piper J, Dickinson BA, Dalby CM, Pestano LA, Jackson AL. A synthetic microRNA-92a inhibitor (MRG-110) accelerates angiogenesis and wound healing in diabetic and nondiabetic wounds. Wound Repair Regen. 2018;26(4):311–323.
39. Yogev S, Shabtay-Orbach A, Nyska A, Mizrahi B. Local toxicity of topically administrated thermoresponsive systems: in vitro studies with in vivo correlation. Toxicol Pathol. 2019;47(3):426–432. doi:10.1177/0192623318810199
40. Yu GT, Monie DD, Khosla S, Tchkonia T, Kirkland JL, Wyles SP. Mapping cellular senescence networks in human diabetic foot ulcers. Geroscience. 2024;46(1):1071–1082. doi:10.1007/s11357-023-00854-x
41. Engeland K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022;29(5):946–960. doi:10.1038/s41418-022-00988-z
42. Khattak SF, Bhatia SR, Roberts SC. Pluronic F127 as a cell encapsulation material: utilization of membrane-stabilizing agents. Tissue Eng. 2005;11(5–6):974–983. doi:10.1089/ten.2005.11.974
43. Collins N. Protein-energy malnutrition and involuntary weight loss: nutritional and pharmacological strategies to enhance wound healing. Expert Opin Pharmacother. 2003;4(7):1121–1140. doi:10.1517/14656566.4.7.1121
44. Park SA, Teixeira LB, Raghunathan VK, et al. Full-thickness splinted skin wound healing models in db/db and heterozygous mice: implications for wound healing impairment. Wound Repair Regen. 2014;22(3):368–380. doi:10.1111/wrr.12172
45. Wang T, Larcher LM, Ma L, Veedu RN. Systematic screening of commonly used commercial transfection reagents towards efficient transfection of single-stranded oligonucleotides. Molecules. 2018;23(10):2564.
46. Lo WY, Yang WK, Peng CT, Pai WY, Wang HJ. MicroRNA-200a/200b modulate high glucose-induced endothelial inflammation by targeting O-linked N-acetylglucosamine transferase expression. Front Physiol. 2018;9:355. doi:10.3389/fphys.2018.00355
47. Lo WY, Wang SJ, Wang HJ. Non-canonical interaction between O-linked N-acetylglucosamine transferase and miR-146a-5p aggravates high glucose-induced endothelial inflammation. Front Physiol. 2020;11:1091. doi:10.3389/fphys.2020.01091
48. Makada H, Singh M. Hydrogels as suitable miRNA delivery systems: a review. Polymers. 2025;17(7):915. doi:10.3390/polym17070915
49. Dai H, Luo J, Deng L, et al. Hierarchically injectable hydrogel sequentially delivers AntagomiR-467a-3p-loaded and AntagomiR-874-5p-loaded satellite-cell-targeting bioengineered extracellular vesicles attenuating sarcopenia. Adv Healthc Mater. 2023;12(17):e2203056. doi:10.1002/adhm.202203056
50. Singh D, Rai V, Agrawal DK. Regulation of collagen I and collagen III in tissue injury and regeneration. Cardiol Cardiovasc Med. 2023;7(1):5–16. doi:10.26502/fccm.92920302
51. Diegelmann RF, Evans MC. Wound healing: an overview of acute, fibrotic and delayed healing. Front Biosci. 2004;9:283–289. doi:10.2741/1184
52. Zhu K, Kakehi T, Matsumoto M, et al. NADPH oxidase NOX1 is involved in activation of protein kinase C and premature senescence in early stage diabetic kidney. Free Radic Biol Med. 2015;83:21–30. doi:10.1016/j.freeradbiomed.2015.02.009
53. Harb A, Elbatreek MH, Elshahat A, El-Akabawy N, Barakat W, Elkomy NM. Repurposing alagebrium for diabetic foot ulcer healing: impact on AGEs/NFkappaB/NOX1 signaling. Eur J Pharmacol. 2023;959:176083. doi:10.1016/j.ejphar.2023.176083
54. Sangartit W, Ha KB, Lee ES, et al. Tetrahydrocurcumin ameliorates kidney injury and high systolic blood pressure in high-fat diet-induced type 2 diabetic mice. Endocrinol Metab. 2021;36(4):810–822. doi:10.3803/EnM.2021.988
55. Issad T, Masson E, Pagesy P. O-GlcNAc modification, insulin signaling and diabetic complications. Diabetes Metab. 2010;36(6 Pt 1):423–435. doi:10.1016/j.diabet.2010.09.001
56. Bond MR, Hanover JA. A little sugar goes a long way: the cell biology of O-GlcNAc. J Cell Biol. 2015;208(7):869–880. doi:10.1083/jcb.201501101
57. Lima VV, Spitler K, Choi H, Webb RC, Tostes RC. O-GlcNAcylation and oxidation of proteins: is signalling in the cardiovascular system becoming sweeter? Clin Sci. 2012;123(8):473–486. doi:10.1042/CS20110638
58. Shvedova M, Thanapaul RJRS, Thompson EL, Niedernhofer LJ, Roh DS. Cellular Senescence in Aging, Tissue Repair, and Regeneration. Plast Reconstr Surg. 2022;150:4S–11S. doi:10.1097/PRS.0000000000009667
59. Demaria M, Desprez PY, Campisi J, Velarde MC. Cell autonomous and non-autonomous effects of senescent cells in the skin. J Invest Dermatol. 2015;135(7):1722–1726. doi:10.1038/jid.2015.108
60. Admasu TD, Rae M, Stolzing A. Dissecting primary and secondary senescence to enable new senotherapeutic strategies. Ageing Res Rev. 2021;70:101412. doi:10.1016/j.arr.2021.101412
61. Wilkinson HN, Clowes C, Banyard KL, Matteuci P, Mace KA, Hardman MJ. Elevated local senescence in diabetic wound healing is linked to pathological repair via CXCR2. J Invest Dermatol. 2019;139(5):1171–1181e6. doi:10.1016/j.jid.2019.01.005
62. Huang K, Mi B, Xiong Y, et al. Angiogenesis during diabetic wound repair: from mechanism to therapy opportunity. Burns Trauma. 2025;13:tkae052. doi:10.1093/burnst/tkae052
63. Short WD, Steen E, Kaul A, et al. IL-10 promotes endothelial progenitor cell infiltration and wound healing via STAT3. FASEB J. 2022;36(7):e22298. doi:10.1096/fj.201901024RR
64. Okonkwo UA, Chen L, Ma D, et al. Compromised angiogenesis and vascular Integrity in impaired diabetic wound healing. PLoS One. 2020;15(4):e0231962. doi:10.1371/journal.pone.0231962
65. Twardowski T, Fertala A, Orgel JP, Antonio JDS. Type I collagen and collagen mimetics as angiogenesis promoting superpolymers. Curr Pharm Des. 2007;13(35):3608–3621. doi:10.2174/138161207782794176
66. Tang L, Cai S, Lu X, et al. Platelet-derived growth factor nanocapsules with tunable controlled release for chronic wound healing. Small. 2024;20(27):e2310743. doi:10.1002/smll.202310743
67. Chen M, Chang C, Levian B, Woodley DT, Li W. Why are there so few FDA-approved therapeutics for wound healing? Int J Mol Sci. 2023;24(20):15109.
68. Sener G, Hilton SA, Osmond MJ, et al. Injectable, self-healable zwitterionic cryogels with sustained microRNA - cerium oxide nanoparticle release promote accelerated wound healing. Acta Biomater. 2020;101:262–272. doi:10.1016/j.actbio.2019.11.014
69. Wei Q, Su J, Meng S, et al. MiR-17-5p-engineered sEVs encapsulated in GelMA hydrogel facilitated diabetic wound healing by targeting PTEN and p21. Adv Sci. 2024;11(13):e2307761. doi:10.1002/advs.202307761
70. Li X, Li D, Wang A, et al. MicroRNA-132 with therapeutic potential in chronic wounds. J Invest Dermatol. 2017;137(12):2630–2638. doi:10.1016/j.jid.2017.08.003
71. Lou R, Chen J, Zhou F, et al. Exosomal miRNA-155-5p from M1-polarized macrophages suppresses angiogenesis by targeting GDF6 to interrupt diabetic wound healing, Molecular therapy. Nucleic Acids. 2023;34:102074. doi:10.1016/j.omtn.2023.102074
72. Everett E, Mathioudakis N. Update on management of diabetic foot ulcers. Ann N Y Acad Sci. 2018;1411(1):153–165. doi:10.1111/nyas.13569
73. Jiang P, Li Q, Luo Y, et al. Current status and progress in research on dressing management for diabetic foot ulcer. Front Endocrinol. 2023;14:1221705. doi:10.3389/fendo.2023.1221705
74. Monteiro-Soares M, Boyko EJ, Jeffcoate W, et al. Diabetic foot ulcer classifications: a critical review. Diabetes Metab Res Rev. 2020;36(Suppl 1):e3272. doi:10.1002/dmrr.3272
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.