Back to Journals » Cancer Management and Research » Volume 12
TMPO-AS1, a Novel E2F1-Regulated lncRNA, Contributes to the Proliferation of Lung Adenocarcinoma Cells via Modulating miR-326/SOX12 Axis
Authors Wei L, Liu Y, Zhang H ![]() , Ma Y
, Ma Y ![]() , Lu Z, Gu Z, Ding C
, Lu Z, Gu Z, Ding C
Received 24 June 2020
Accepted for publication 28 October 2020
Published 2 December 2020 Volume 2020:12 Pages 12403—12414
DOI https://doi.org/10.2147/CMAR.S269269
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Eileen O'Reilly
Lin Wei,1 Yugang Liu,1 Huijun Zhang,1 Yuefeng Ma,2 Zenghui Lu,1 Zhenning Gu,1 Chao Ding1
1Department of Thoracic Surgery, Xi’an Chest Hospital, Xi’an 710100, People’s Republic of China; 2Department of Thoracic Surgery, The Second Affiliated Hospital of Xi’an Jiao Tong University, Xi’an 710004, People’s Republic of China
Correspondence: Chao Ding
Department of Thoracic Surgery, Xi’an Chest Hospital, Xi’an, Shaanxi Province 710100, People’s Republic of China
Tel/Fax +86-29-62500053
Email [email protected]
Background: TMPO-AS1, an antisense lncRNA located at human chromosome 12p23.1, has been identified as an oncogene involved in cell proliferation in various cancers, including LUAD. In this study, we aimed to explore the novel molecular mechanism of TMPO-AS1 underlying LUAD growth.
Materials and Methods: The transcription levels of TMPO-AS1, miR-326, and SOX12 in LUAD tissues and cell lines were detected by quantitative real-time PCR (qRT-PCR). The cell proliferation ability was evaluatect 3d by cell counting kit-8 (CCK-8) assay. Cell cycle and apoptosis analysis was assessed by flow cytometry. The target relationship among TMPO-AS1, miR-326, and SOX12 and promoter activity of TMPO-AS1 was measured using dual-luciferase reporter assay. The protein levels of SOX12 in LUAD cells were determined by Western blot. ChIP-qPCR assay was performed to validate the direct binding between E2F1 and TMPO-AS1 promoter.
Results: TMPO-AS1 was up-regulated in LUAD tissues as well as cell lines. Boosted TMPO-AS1 expression was positively correlated with poor prognosis and pathological stage in LUAD. Down-regulation of TMPO-AS1 could restrain the proliferation of LUAD cells through arresting the cell cycle at G0/G1 phase and inducing apoptosis in vitro. Mechanically, we demonstrated that TMPO-AS1 could modulate the proliferation of LUAD cells through increasing SOX12 expression level via sponging miR-326 in accordance with bioinformatics analysis and experimental validation. Furthermore, we identified that TMPO-AS1 could be activated by E2F transcription factor 1 (E2F1) as a novel target gene.
Conclusion: TMPO-AS1 can modulate LUAD cell proliferation through E2F1/miR-326/SOX12 pathway.
Keywords: lung adenocarcinoma, TMPO-AS1, cell proliferation, miR-326, SOX12, E2F1
Introduction
Lung cancer is one of the most common malignant tumor types exhibiting a high incidence and mortality rates worldwide including China.1,2 As the predominant subtype, non-small cell lung cancer (NSCLC) accounts for approximately 85% of all lung cancer cases. Based on histopathological features, NSCLC can be further categorized into three main subtypes: lung adenocarcinoma, lung squamous cell carcinoma, and large cell lung carcinoma.3 Strikingly, lung adenocarcinoma (LUAD) is the leading sub‐class of NSCLC, accounting for about 40% of all lung cancers and a majority of cancer-related deaths.4 Despite the fact that great advancements have been made in diagnostic and therapeutic approaches involving early screening, surgical excision, radiotherapy, chemotherapy as well as immunotherapy, the prognosis of LUAD patients is still unfavorable owing to a high frequency of local/distant metastasis and recurrence.5 Thus, there is an urgent need to elucidate the molecular mechanisms underlying LUAD progression to develop novel promising diagnostic biomarkers and therapeutic targets for LUAD treatment or early detection.
Long non-coding RNAs (lncRNAs) represent a large class of RNA transcripts longer than 200 nucleotides but lacking overt protein-coding capacity. Increasing evidence indicates that lncRNAs, as key regulator, have been proven to play critical roles in progression of multiple cancer types, including lung cancer.6 Numerous lncRNAs have been demonstrated to be dysregulated in LUAD and contribute to tumor growth, invasion and metastasis through interacting with miRNA and mRNA to construct a competing endogenous RNA (ceRNA) network.7 For instance, a recent study demonstrated that LINC00460, an up-regulated lncRNA in LUAD, could promote cell growth by competitive binding with miR-302c-5p to elevate the expression of FOXA1, which is a modulator of cell proliferation in multiple cancer types, including LUAD.8
TMPO antisense RNA 1 (TMPO-AS1), an antisense lncRNA located in chromosome region 12p23.1, was transcribed in antisense orientation from the 5ʹ end region of thymopoietin (TMPO) that a coding gene. In recent years, many studies have demonstrated that TMPO-AS1, as an oncogene, was dramatically up-regulated in multiple cancer types and associated with unfavorable prognosis, as well as tumor progression.9–12 For example, in osteosarcoma, TMPO-AS1 depletion could suppress cell proliferation and facilitate cell apoptosis via modulating miR-199a-5p/WNT7B pathway;9 in prostate cancer, TMPO-AS1 was shown to be directly down-regulated by androgen receptor (AR) and its overexpression could accelerate cell proliferation as well as migration;10 in cervical cancer, silencing of TMPO-AS1 could restrain cell proliferation and migration and induce cell apoptosis via/miR-577/RAB14 regulatory axis. For LUAD, recent studies demonstrated that TMPO-AS1 was elevated in LUAD and correlated with unfavorable prognosis of patients, knockdown of TMPO-AS1 could significantly reduce abilities of cell proliferation, migration and invasion through up-regulating TMPO or down-regulating miR-383-5p.11,12
In the present study, we aimed to probe the roles of TMPO-AS1 on cell proliferation and its underlying regulating mechanism in LUAD. Our findings revealed that E2F1/TMPO‐AS1/miR-326/SOX12 pathway, as a novel regulatory axis, contributed to LUAD cell proliferation and might serve as a potential molecular target for LUAD therapy.
Materials and Methods
Tissue Samples
Twenty-five pairs of LUAD tissues and adjacent non-tumor tissues were collected from patients (aged from 33 to 74; 16 males and 9 females; 15 and 10 patients with or without lymph node metastasis) who received surgical resection at Xi’an Chest Hospital (Xi’an, Shaanxi Province, China). All samples were not subjected to radiotherapy or chemotherapy before collection and stored in liquid nitrogen. This study was approved by the Ethics Committee of Xi’an Chest Hospital and informed consent was provided by all patients.
Cell Culture
Human lung adenocarcinoma cells (HCC827, A549, H838, H1299, SK-LU-1 and H23) and a normal human lung epithelial cell line, BEAS-2B, were obtained from the American Type Culture Collection (ATCC, USA) and cultured with Dulbecco’s modified Eagle medium (Biological Industries, Cromwell, CT, USA) containing 10% FBS, 100 U/mL penicillin, 100 mg/mL streptomycin (Biological Industries, Cromwell, CT, USA) in humidified atmosphere with 5% CO2 under 37°C.
Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted using RNAiso Plus (TaKaRa, Dalian, China). cDNA for lncRNA/mRNA and microRNA (miRNA) detection was generated with the TIANScript Ⅱ RT Kit and miRcute Plus miRNA First-Strand cDNA Kit (TIANGEN Biotech, Beijing, China) respectively using total RNA. qRT-PCR reactions were carried out using SuperReal PreMix Plus and miRcute Plus miRNA qPCR Kit (TIANGEN Biotech, Beijing, China) to detect lncRNA/mRNA and miRNA, respectively. The 2−ΔΔCt method was performed to calculate the relative expression level of target genes. GAPDH and U6 were used as an endogenous control for lncRNA/mRNA and miRNA detection, respectively. Primers were as follows: TMPO-AS1, 5ʹ-CAGTTTAAAAGGCGCTGGGG-3ʹ(forward), 5ʹ- CGCACAAAAGCAGTACGACC-3ʹ (reverse); miR-326, 5ʹ- GGCGCCCAGATAATGCG-3ʹ (forward); SOX12, 5ʹ- GACATGCACAACGCCGAGATCT-3ʹ (forward), 5ʹ- GTAATCCGCCATGTGCTTGAGC-3ʹ (reverse); E2F1, 5ʹ-GGACCTGGAAACTGACCATCAG-3ʹ (forward), 5ʹ-CAGTGAGGTCTCATAGCGTGAC-3ʹ (reverse); GAPDH, 5ʹ- GTCTCCTCTGACTTCAACAGCG-3ʹ (forward), 5ʹ- ACCACCCTGTTGCTGTAGCCAA-3ʹ(reverse); U6, 5ʹ- CTCGCTTCGGCAGCACATATACT-3ʹ (forward), 5ʹ-ACGCTTCACGAATTTGCGTGTC-3ʹ (reverse).
siRNAs and Plasmid Transfection
Small interfering RNAs (siRNA) targeting TMPO-AS1, SOX12, E2F1 and negative control (NC) were synthesized by Sangon Biotech (Shanghai, China) using the following sequence: siTMPO-AS1, 5ʹ-GCUUGUCUGCAGGCACUCAUA-3ʹ; siSOX12, 5ʹ-GGGCCUUUCUUGCGCUCUAUU-3; siE2F1, 5ʹ-GUGGAUUCUUCAGAGACAUTT-3ʹ; siNC, 5ʹ-GCGUUGCUGUCGCUCUAUUCU-3ʹ. miR-326 mimics/inhibitors with or without biotin label and corresponding control were purchased from Sangon Biotech (Shanghai, China). The overexpression vector of TMPO-AS1 (pcDNA-TMPO-AS1), E2F1 (pcDNA-E2F1) and pcDNA3.1 empty vector (pcDNA3.1) were also purchased from Synbio Tech (Suzhou, China). X-tremeGENE siRNA Transfection Reagent (Roche, Basel, Switzerland) was utilized to transfect the nucleotides and plasmid into cells.
All nucleotides and plasmid mentioned previously were transfected into cells using X-tremeGENE siRNA Transfection Reagent (Roche, Basel, Switzerland).
Cell-Counting Kit-8 (CCK-8) Assay
CCK-8 assay was carried out to evaluate the cell proliferation activity. Briefly, cells were inoculated into 96-well plates at a density of 5×103 cells/well and cultured with CCK-8 reagent (MedChemExpress, Monmouth Junction, USA) for 2 hours under 37°C and the absorbance at 450 nm was measured to calculate the using microplate reader.
Cell Cycle and Apoptosis Analysis by Flow Cytometry
For cell cycle, the cells were harvested and fixed with 70% ethanol at 4°C overnight and stained with propidium iodide (PI) containing Ribonuclease A (Solarbio, Beijing, China) for 30 min at room temperature and the cell cycle distribution was then analyzed by flow cytometry (BD Biosciences, Franklin Lakes, NJ, USA). For apoptosis, the cells were harvested and detected using the Annexin V-FITC Apoptosis Detection Kit (Solarbio, Beijing, China) by flow cytometry (BD Biosciences, Franklin Lakes, NJ, USA) according to the manufacturer’s instructions. The data were analyzed using the FlowJo software.
Luciferase Reporter Assay
For miR-326 binding detection, TMPO-AS1 or SOX12 sequence containing wild-type (Wt) or Mutant (Mut) miR-326 binding sites was inserted into the pmirGLO reporter vector (Promega, Madison, WI, USA) by Synbio Tech (Suzhou, China), then the luciferase reporter assay was carried out after co-transfecting the reporter plasmid and miR-326 mimics or pcDNA-TMPO-AS1 vector in HEK293 cells using X-tremeGENE siRNA Transfection Reagent (Roche, Basel, Switzerland).
For promoter activity analysis, the promoter region (2000 bp upstream from transcription start site) of TMPO-AS1 was amplified by PCR from human genomic DNA and inserted into pGL3-Basic vector (Promega, Madison, WI, USA), then the luciferase reporter assay was carried out after co-transfecting the reporter plasmid and pcDNA-E2F1 vector in HEK293 cells using X-tremeGENE siRNA Transfection Reagent (Roche, Basel, Switzerland). Primers used to amplify the promoter of TMPO-AS1 was as follows: 5ʹ-GTTCTATAGAAAATTAGGTTTTTTAT-3ʹ (forward) and 5ʹ-TCCCGGGCGCCCCCTCGG-3ʹ (reverse).
After transfection, the luciferase activity was determined using the dual-luciferase reporter assay system (Promega, Madison, WI, USA) and was normalized to Renilla luciferase activity.
Biotin-Coupled miRNA Pull Down
The pull-down assay with biotinylated miR-326 was performed according to a previously published method.13 In brief, the biotinylated-has-miR-326 mimics and the negative control cel-miR-67 mimics (GenePharma, China) were transfected into H838 and SK-LU-1 cells for 24 hours. Then whole-cell lysates were prepared and incubated with Streptavidin Magnetic Beads (MedChemExpress, Monmouth Junction, USA) at 4°C overnight with rotation. The beads were then harvested and washed thoroughly. Next, the RNA-RNA complexes bound to the beads were extracted with RNeasy Mini Kit (Qiagen, Germany) and the expression of TMPO-AS1 and SOX12 was analyzed by qRT-PCR.
Western Blot
Total proteins were extracted by CelLytic™ MT Cell Lysis Reagent (Sigma-Aldrich, St Louis, USA) containing protease inhibitor (cOmplete™ Tablets, EDTA-free, Sigma-Aldrich). 10% SDS-PAGE was performed to separate cell lysate proteins which were then transferred onto 0.22 μm PVDF membrane. The membrane was blocked with 5% fat-free milk in TBST for 2 h at room temperature and incubated with the primary antibodies at 4°C overnight. Following the incubation with HRP labeled the secondary antibodies for 2 h at room temperature, signals were detected by exposure to films with the chemiluminescence system (SignalFire™ Plus ECL Reagent, CST, USA). GAPDH was used as an internal control. The antibodies used for Western blot were as follows: anti-SOX12 (ab54371, Abcam; 1:500), anti-Cyclin D1 (ab40754, Abcam; 1:2000), anti-CDK4 (ab68266, Abcam; 1:500), anti-CDK6 (ab124821, Abcam; 1:50,000) anti-GAPDH (ab8245, Abcam; 1:1000), HRP-conjugated Goat Anti-Mouse IgG (H+L) (ab205719, Abcam; 1:10,000), HRP-conjugated Goat Anti-Rabbit IgG (H+L) (ab205718, Abcam; 1:10,000).
Chromatin Immunoprecipitation
Chromatin immunoprecipitation (ChIP) experiment was performed using the SimpleChIP® Plus Enzymatic Chromatin IP Kit (#9003, CST, USA) according to the manufacturer’s instructions. The antibodies used for immunoprecipitating crosslinked protein-DNA complexes were as follows: rabbit anti-E2F1 (#3742, CST, USA), and normal rabbit IgG (#2729, CST, USA). The immunoprecipitated DNA was purified for qRT-PCR experiment and three pairs of was designed for detecting TMPO-AS1 promoter were as follows: Primer 1 (−122 to −244 bp), 5ʹ-AAGTCCCCGTCTAAACGCC-3ʹ (forward) and 5ʹ-CGCGTTCTTTGACGTGTGG-3ʹ (reverse); Primer 2 (−359 to −505 bp), 5ʹ-AAAGATGGCCTGTTGCTGGG-3ʹ (forward) and 5ʹ-TGGAGGGCAACTGATGACAC-3ʹ (reverse); Primer 3 (−911 to −1056 bp), 5ʹ-GCCGATTTGAGATTTAACCAGGA-3ʹ (forward) and 5ʹ-GTTTGGATTGCGTTTTCCCCA-3ʹ (reverse).
Bioinformatics Analysis
The TMPO-AS1 expression levels among lung adenocarcinoma (n=526) and normal tissues (n=59) were analyzed by ENCORI (http://starbase.sysu.edu.cn/panCancer.php) using TCGA data. The association between TMPO-AS1 expression and overall survival of patients was explored by GEPIA (http://gepia.cancer-pku.cn/) based on TCGA data, the Cutoff-High (%) was set as 70% and the Cutoff-Low (%) was set as 30%. The association between TMPO-AS1 expression and pathological stage of patients was also analyzed by GEPIA based on TCGA data and the major stag was adopted for plotting.
Statistical Analysis
Results were presented by means ± standard deviation (SD) and were analyzed with SPSS software using Student’s t-test, one-way ANOVA with Tukey’s post hoc test or Spearman correlation analysis. Differences were accepted as statistically significant at a level of p < 0.05.
Results
Up-Regulated TMPO-AS1 is Associated with Poor Prognosis in LUAD
To evaluate the expression level of TMPO-AS1 and its clinical significance in LUAD, we firstly examined the expression of TMPO-AS1 in LUAD based on TCGA data using online tool ENCORI (http://starbase.sysu.edu.cn/panCancer.php), and the results showed TMPO-AS1 was significantly up-regulated in LUAD (Figure 1A). The association between TMPO-AS1 expression and overall survival, as well as pathological stage were then investigated by GEPIA (http://gepia.cancer-pku.cn/), the results revealed that high expression level of TMPO-AS1 was closely associated with shorter overall survival in LUAD patients (Figure 1B) and positively correlated with advanced pathological grade (Figure 1C). Subsequently, up-regulation of TMPO-AS1 was validated in 25 pairs of LUAD and adjacent non-tumor tissues, as well as six LUAD cell lines by qRT-PCR. As expected, TMPO-AS1 expression was significantly up-regulated in LUAD tissue compared to adjacent normal tissue (Figure 1D). Consistently, we also found that TMPO-AS1 expression was elevated in all six detected LUAD cell lines, including HCC827, A549, H838, H1299, SK-LU-1, H23 cells, compared with BEAS-2B cells, a normal human lung epithelial cell line (Figure 1E).
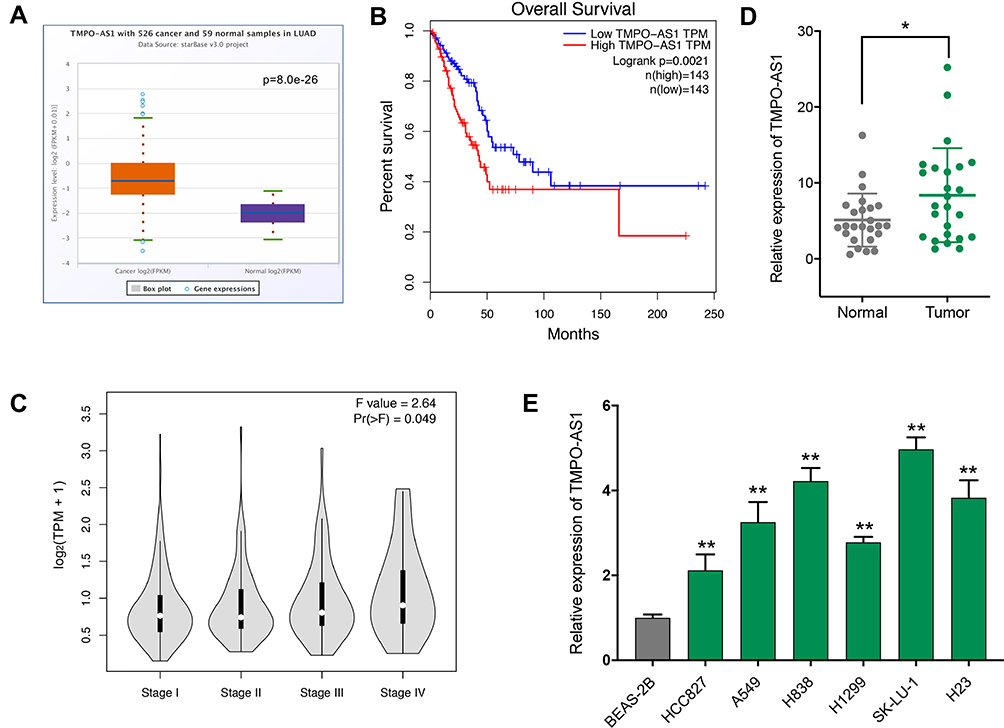 |
Figure 1 Up-regulated TMPO-AS1 is associated with poor prognosis in lung adenocarcinoma (LUAD). (A) TMPO-AS1 expression in LUAD (n=526) and non-cancerous (n=59) tissues was analyzed by ENCORI. (B) The relevance between TMPO-AS1 expression and pathological stage was assessed by GEPIA. (C) The relevance between TMPO-AS1 expression and overall survival was assessed by GEPIA. (D) The expression level of TMPO-AS1 in human LUAD tissues and paired adjacent normal tissues (n = 25) was determined by qRT-PCR. (E) The expression level of TMPO-AS1 in six LUAD cell lines and a normal human lung epithelial cell line (BEAS-2B) was determined by qRT-PCR. *P < 0.05, ** P < 0.01. |
Knockdown of TMPO-AS1 Inhibits Cell Proliferation Through Inducing Cell Cycle Arrest at G0/G1 Phase and Accelerates Apoptosis
To clarify the role of TMPO-AS1 in LUAD cells, we performed the loss-of-function experiment using siRNA in both H838 and SK-LU-1 cells which express higher level of TMPO-AS1 to observe the effects of TMPO-AS1 knockdown on cell proliferation, cell cycle and apoptosis in vitro. qRT-PCR results indicated that TMPO-AS1 expression level could be significantly silenced upon transfection of siTMPO-AS1 in both H838 and SK-LU-1 cells (Figure 2A). CCK-8 assay showed that TMPO-AS1 knockdown could suppress the proliferation ability of both H838 and SK-LU-1 cells (Figure 2B). To further determine whether the alteration of cell cycle progression and apoptosis contribute to the inhibition of cell proliferation caused by TMPO-AS1 knockdown, flow cytometric analysis was adopted to examine cell cycle and apoptosis after cells were stained with PI or/and Annexin V. We observed that TMPO-AS1 knockdown could arrest cell cycle at G0/G1 phase (Figure 2C) and promote the apoptosis (Figure 2D) in both H838 and SK-LU-1. In addition, we found cell cycle-related proteins Cyclin D1 and CDK4 were down-regulated in TMPO-AS1 knockdown LUAD cells (Figure 2E). These results suggested that knockdown of TMPO-AS1 could inhibit the proliferation of LUAD cells through inducing cell cycle arrest at G0/G1 phase and promote apoptosis.
 |
Figure 2 TMPO-AS1 knockdown inhibits LUAD cell proliferation. (A) The efficiency of TMPO-AS1 knockdown in H838 and SK-LU-1 cells was determined by qRT-PCR. (B) The effect of TMPO-AS1 knockdown on cell proliferation was evaluated by CCK-8 assay. (C) The effect of TMPO-AS1 knockdown on cell cycle distribution was detected by flow cytometry. (D) The effect of TMPO-AS1 knockdown on apoptosis was detected by flow cytometry. (E) The effect of TMPO-AS1 knockdown on the expression of Cyclin D1, CDK4 and CDK6 was detected by Western blot. siTMPO-AS1, knockdown group; siNC, negative control group; *P < 0.05, ** P < 0.01. |
TMPO-AS1 Serves as a Sponge for miR-326 in LUAD Cells
TMPO-AS1 has been demonstrated to contribute to tumor progression as a ceRNA to regulate the expression of target genes by sponging miRNA in several types of cancer.9,14 Thus, we speculated that TMPO-AS1 might modulate the cell proliferation of LUAD by ceRNA mechanism that regulates LUAD-related miRNAs and their target genes. Using online prediction tool for lncRNA-miRNA interaction (lncRNASNP2, http://bioinfo.life.hust.edu.cn/lncRNASNP), we found TMPO-AS1 could be targeted by many miRNAs (Supplementary Figure 1). Here, we particularly focused on miR-326, which has been demonstrated to be down-regulated in LUAD and functions as a tumor suppressor through directly interacting with lncRNA15 and circular RNA.16,17 The predicted binding site of miR-326 in the TMPO-AS1 sequence was illustrated in Figure 3A. As expected, we observed that miR-326 was down-regulated in LUAD tissues and cell lines (Figure 3B and C), and exhibited an inverse correlation with TMPO-AS1 expression in LUAD tissues (Figure 3D). Dual-luciferase reporter assays revealed that miR-326 mimics could significantly inhibit the reporter activities of TMPO-AS1-WT but not in the mutant type (Figure 3E). Biotin-labeled miRNA pull down assay further indicated that TMPO-AS1 was significantly enriched in the biotin-labeled miR-326-captured fraction compared to that in the negative control (biotinylated C.elegans miR-67) (Figure 3F). Moreover, we further found the miR-326 expression level could be enhanced by TMPO-AS1 knockdown (Figure 3G), while TMPO-AS1 expression could be down-regulated by miR-326 mimics (Figure 3H). All these results suggested that TMPO-AS1 could directly bind to miR-326 as an miRNA sponge in LUAD cells.
 |
Figure 3 TMPO-AS1 sponged miR-326 in LUAD cells. (A) Bioinformatics analysis showed the predicted binding sites between TMPO-AS1 wild type (TMPO-AS1-WT) and miR-326, TMPO-AS1-mutant (MUT) represented mutation of binding sites in TMPO-AS1. (B) The expression level of miR-326 in human LUAD tissues and paired non-tumor tissues (n=25). (C) The expression level of miR-326 in LUAD cell lines. (D) The relevance between the expression of TMPO-AS1 and miR-326 was evaluated with Spearman correlation analysis. (E) Dual luciferase reporter assay was used to confirm the combination between TMPO-AS1 and miR-326. (F) H383/SK-LU-1 cell lysates were incubated with biotin-labeled miR-326 or negative control cel-miR-67, and the relative enrichment of TMPO-AS1 was detected by qRT-PCR. (G) The effect of TMPO-AS1 knockdown on the expression level of miR-326 in LUAD cells. (H) The effect of overexpression of miR-326 on the expression level of TMPO-AS1 in LUAD cells. ** P < 0.01. |
TMPO-AS1 Modulates the Proliferation of LUAD Cells Through Up-Regulating SOX12 via Sponging miR-326
To further probe the molecular mechanism underlying the regulation of LUAD cell proliferation by TMPO-AS1 via sponging miR-326, we tried to identify the potential downstream target of miR-326 and found that SRY-box transcription factor 12 (SOX12) existed as a potential complementary binding site with miR-326 using TargetScan (http://www.targetscan.org) (Figure 4A). qRT-PCR results showed that SOX12 expression was significantly elevated in LUAD tissue and cell lines (Figure 4B and C), and there was a positive correlation between the expression of SOX12 and TMPO-AS1 in LUAD tissues (Figure 4D). Biotin-labeled miRNA pull down assay indicated that SOX12 mRNA was significantly enriched in the biotin-labeled miR-326-captured fraction compared to that in the negative control (Figure 4E). Dual-luciferase reporter assays further revealed that the luciferase activities of SOX12-WT reporter but not the mutant type were significantly suppressed by miR-326 mimics and could be recovered by overexpression of TMPO-AS1, indicating TMPO-AS1, miR-326 and SOX12 could form a ceRNA network (Figure 4F). Subsequently, we evaluated the regulatory effect of TMPO-AS1 and miR-326 on the expression of <italic>SOX12</italic> at mRNA level using qRT-PCR, the results showed that knockdown of TMPO-AS1 could down-regulate the SOX12 expression level in both H838 and SK-LU-1 cells and the down-regulation of SOX12 caused by TMPO-AS1 knockdown could be rescued by miR-326 inhibitor (Figure 4G); we then validated the regulatory effect of TMPO-AS1 and miR-326 on the expression of SOX12 at protein level in H838 cells using western blot and found the results were coincident with that of mRNA level (Figure 4H). In addition, we also revealed that miR-326 inhibition or SOX12 overexpression could abrogate the suppression of cell proliferation and the G0/G1 arrest caused by TMPO-AS1 knockdown in both H838 and SK-LU-1 cells (Figure 4I and J). Taken together, these findings indicate that TMPO-AS1 could modulate the proliferation of LUAD cells through up-regulating SOX12 via sponging miR-326.
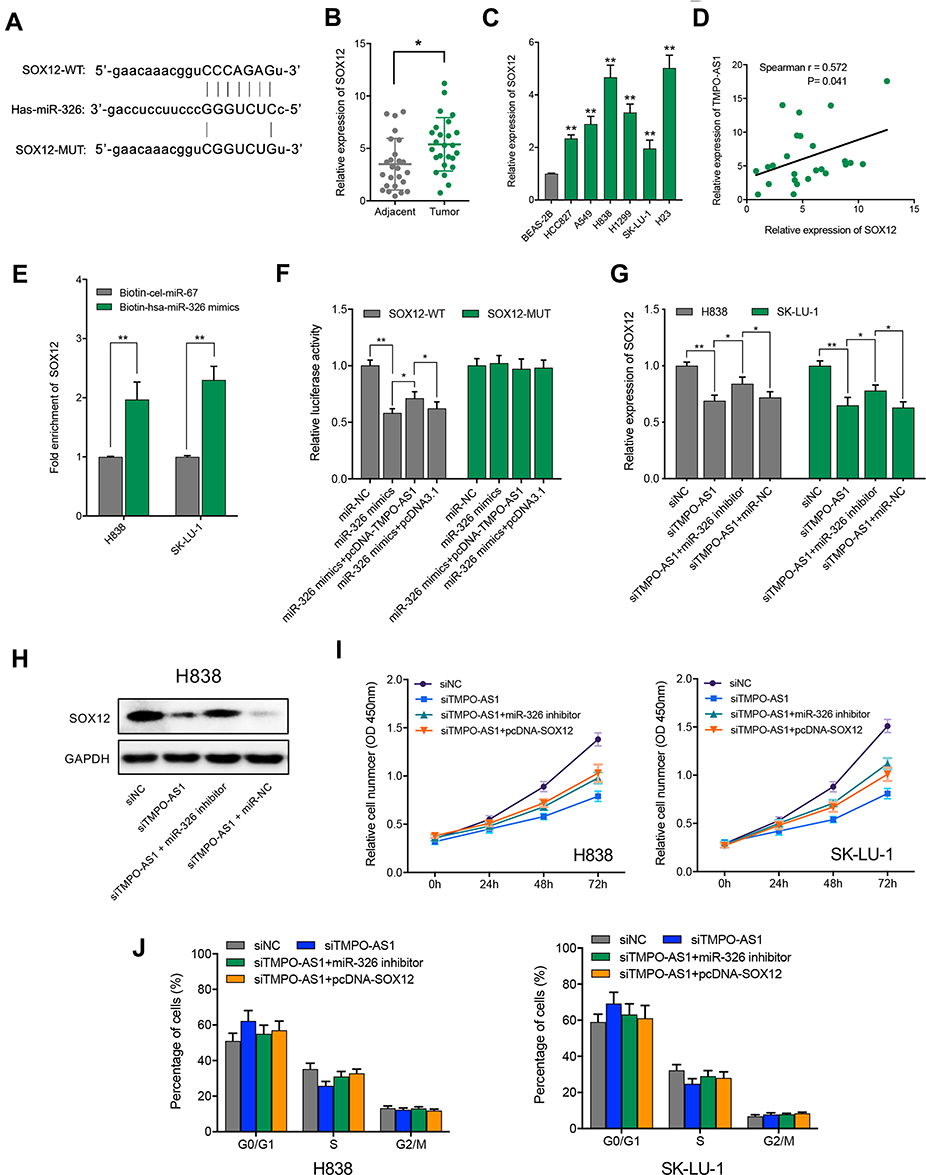 |
Figure 4 TMPO-AS1 modulates the proliferation of LUAD cells through regulation of miR-326/SOX12 axes. (A) Bioinformatics analysis showed the predicted binding sites between SOX12 wild type (SOX12-WT) and miR-326, SOX12-mutant (MUT) means mutation of binding sites in 3ʹ UTR of SOX12. (B) The expression level of SOX12 in human LUAD tissues and paired adjacent non-tumor tissues (n=25) was detected by qRT-PCR. (C) The expression level of SOX12 in LUAD cell lines and the normal human lung epithelial cell line (BEAS-2B) was detected by qRT-PCR. (D) The expression relevance between TMPO-AS1 and SOX12 was evaluated with Spearman correlation analysis. (E) H383/SK-LU-1 cell lysates were incubated with biotin-labeled miR-326 or negative control cel-miR-67, and the relative enrichment of SOX12 mRNA was detected by qRT-PCR. (F) Dual luciferase reporter assay was used to confirm the binding of SOX12 and miR-326 and observe the effect of overexpression of TMPO-AS1 on bindings. (G) SOX12 mRNA level could be regulated by TMPO-AS1 via miR-326 in H838 and SK-LU-1 cells. (H) SOX12 protein level could be regulated by TMPO-AS1 via miR-326 in H838 cells. (I) CCK-8 rescue assay indicated that TMPO-AS1 knockdown could suppress the LUAD cell proliferation through miR-326/SOX12 axis. (J) Cell cycle analysis showed that TMPO-AS1 knockdown could induce the G0/G1 phase arrest in LUAD cells via miR-326/SOX12 axis. *P < 0.05, ** P < 0.01. |
TMPO-AS1 is Activated by the Transcription Factor E2F1 in LUAD Cells
To survey the reason of TMPO-AS1 up-regulation in LUAD, we analyzed the TMPO-AS1 promoter region defined as 2000 bp up-steam of transcription start site (TSS) using bioinformatical tools of JASPAR (http://jaspar.genereg.netl) and 41 putative E2F1 binding sites were detected (Supplementary Table 1). Interestingly, E2F1 has been reported to be up-regulated in LUAD and promote LUAD cell proliferation as a critical regulator.18,19 Here, we found that the expression level of E2F1 was significantly up-regulated in LUAD tissues (Figure 5A), and positively correlated with TMPO-AS1 expression level (Figure 5B). Next, we evaluated the regulatory effect of E2F1 on the expression of TMPO-AS1 in LUAD cells through overexpressing or silencing E2F1, and qRT-PCR results indicated that overexpression of E2F1 could up-regulate the TMPO-AS1 expression level in both H838 and SK-LU-1 cells (Figure 5C), whereas E2F1 knockdown decreased TMPO-AS1 expression (Figure 5D). To confirm TMPO-AS1 was regulated by E2F1 at transcriptional level, TMPO-AS1 promoter was cloned into pGL3-basic vector and then co-transfected with E2F1 overexpression vector into HEK-293 cells, and we found luciferase activity could be facilitated by overexpression of E2F1 (Figure 5E). Furthermore, ChIP-qPCR assay was performed to validate the direct binding between E2F1 and TMPO-AS1 promoter, and the results showed a significant enrichment of E2F1 on TMPO-AS1 promoter in both H838 and SK-LU-1 cells (Figure 5F). These findings manifest that the up-regulation of TMPO-AS1 is mediated by E2F1 in LUAD.
 |
Figure 5 TMPO-AS1 is regulated by the transcription factor E2F1. (A) The expression level of E2F1 in human LUAD tissues and paired adjacent normal tissues (n = 25) was determined by qRT-PCR. (B) The expression relevance between TMPO-AS1 and E2F1 was assessed with Spearman correlation analysis. (C) Expression levels of TMPO-AS1 in H838 and SK-LU-1 cells after overexpression of E2F1. (D) Expression levels of TMPO-AS1 in H838 and SK-LU-1 cells after knockdown of E2F1. (E) Luciferase activities of TMPO-AS1 promoter upon overexpression of E2F1. (F) Direct binding between E2F1 and TMPO-AS1 promoter was validated in H838 and SK-LU-1 cells by ChIP-qPCR assay using three pairs of primers targeting different regions, 1% of genomic DNA was kept as input. *P < 0.05, ** P < 0.01. |
Discussion
Accumulating evidence demonstrated that lncRNAs represent a novel class of regulators for gene expression and play critical roles in progression and development of various cancers, including LUAD.20 To date, numerous dysregulated lncRNAs have been identified to be involved in LUAD as either tumor suppressor or oncogene, including some well-known lncRNAs such as HOXD-AS1,21 MALAT1,22 NEAT1,23 TUG1,24 and so on. In recent years, TMPO-AS1, a lncRNA located at 12p23.1 region, has been identified to be up-regulated in several cancer types and contribute to tumor progression, involving osteosarcoma,9 breast cancer,25 cervical cancer,14 prostate cancer,10 as well as lung adenocarcinoma.11 Yet, the underlying molecular mechanism of TMPO-AS1 in the progression of LUAD remains largely unknown, the present study was carried out to explore a novel insight into the underlying mechanism of TMPO-AS1 on cell proliferation.
Firstly, we revealed that TMPO-AS1 was strongly expressed in LUAD tissues and cell lines and the up-regulation of TMPO-AS1 was positively associated with poor prognosis and histologic grade in LUAD, suggesting that TMPO-AS1 might serve as a potential biomarker for early diagnosis and prognostic prediction of LUAD. Similar to our results, two recent studies in NSCLC also demonstrated that TMPO-AS1 was up-regulated in both clinical samples and cell lines of NSCLC and the high expression of TMPO-AS1 was significantly correlated with TNM stage, lymph node metastasis, and the increased risk of death.11,12
Secondly, we demonstrated that knockdown of TMPO-AS1 could suppress the proliferation of two LUAD cell lines (H838 and SK-LU-1) through inducing cell cycle arrest at G0/G1 phase and accelerate apoptosis, similar to that found in other LUAD cell lines. Qin et al revealed that the silencing of TMPO-AS1 could inhibit the proliferation ability of A549 and H1299 cells through blocking the cell cycle at G0/G1 phase and facilitating apoptosis. Another study revealed that TMPO-AS1 could also control the growth of H460 cells, another LUAD cell line.11
Thirdly, we uncovered a novel ceRNA pathway that contributes to regulation of cell proliferation by TMPO-AS1 in LUAD cells, that is, TMPO-AS1 could modulate the proliferation of LUAD cells through up-regulating SOX12 via sponging miR-326 (ie, miR-326/SOX12 axis). Several ceRNA pathways mediated by TMPO-AS1 have been identified by previous studies and demonstrated to be involved in cancer development through controlling cell growth and survival, such as miR‐199a‐5p/WNT7B in osteosarcoma,9 miR-577/RAB14 in cervical cancer.14 Here, we discovered that TMPO-AS1 could directly bind to miR-326 as a sponge in LUAD cells. A previous study demonstrated that miR-326 was down-regulated in LUAD cells/tissues and its low expression was significantly associated with worse prognosis in LUAD patients.26 Of note, miR-326 could inhibit LUAD cell proliferation by targeting several important oncogenes like PHOX2A,26 ADAM17,27 CCND1,28 and so on, indicating that miR-326 may serve as a tumor suppressor in LUAD. In addition, we also found SOX12 to be a novel target of miR-326 contributing to regulation of cell proliferation mediated by TMPO-AS1 through forming the ceRNA pathway in LUAD. The previous studies have demonstrated that SOX12, as an oncogene, play important roles in tumor progression, including LUAD. Wang et al found that SOX12 was up-regulated in lung cancer tissues and high SOX12 expression predicted reduced overall survival, and knockdown of SOX12 could impair lung cancer cell proliferation, migration and invasion.29 Interestingly, some recent studies revealed that SOX12 participated in tumor progression through forming lncRNA-miRNA-mRNA regulatory axis. For example, in tongue squamous cell carcinoma, lncRNA CASC9 was demonstrated to affect cell proliferation, migration, and invasion via regulating miR-423-5p/SOX12 axes;30 while in papillary thyroid carcinoma, it was found that lncRNA NR2F1-AS1 could regulate miR-423-5p/SOX12 to promote cell proliferation and invasion.31 In the current study, we discovered that SOX12 was up-regulated in LUAD tissues and cell lines and contributed to LUAD cell proliferation as downstream target of TMPO-AS1/miR-326 pathway.
Finally, we elucidated that transcription factor E2F1 contributed to the up-regulation of TMPO-AS1 in LUAD at transcriptional level. E2F1, a member of the E2F family of transcription factors known for regulating many pivotal cellular processes including cell proliferation and apoptosis,32,33 has been disclosed to be a key modulator responsible for controlling cell proliferation and apoptosis in LUAD through targeting several tumor-associated genes including lncRNA, such as lncRNA-PLACT1,34 LINC00461.18 In the present study, we identified TMPO-AS1 as novel transcriptional target gene of E2F1 in LUAD.
Conclusion
This paper revealed that TMPO-AS1, an E2F1-activated lncRNA, was up-regulated in LUAD and contributed to LUAD cell proliferation through modulating cell cycle and apoptosis via a novel ceRNA pathway of miR-326/SOX12 (Figure 6), suggesting that TMPO-AS1 might potentially act as an effective diagnostic marker and therapeutic target for LUAD.
 |
Figure 6 The proposed model for the regulation of LUAD growth via E2F1/TMPO‐AS1/miR-326/SOX12 pathway. |
Abbreviations
NSCLC, non-small cell lung cancer; LUAD, lung adenocarcinoma; TMPO-AS1, TMPO antisense RNA 1; LncRNA, long non-coding RNA; ceRNA, competing endogenous RNA; SOX12, SRY-box transcription factor 12.
Acknowledgment
This work was supported by the Key Research and Development Program of Shaanxi Province of China (2018SF-222).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Barta JA, Powell CA, Wisnivesky JP. Global epidemiology of lung cancer. Ann Glob Health. 2019;85(1):8.
2. Cao M, Chen W. Epidemiology of lung cancer in China. Thorac Cancer. 2019;10(1):3–7. doi:10.1111/1759-7714.12916
3. Inamura K. Lung cancer: understanding its molecular pathology and the 2015 WHO classification. Front Oncol. 2017;7:193. doi:10.3389/fonc.2017.00193
4. Herbst RS, Morgensztern D, Boshoff C. The biology and management of non-small cell lung cancer. Nature. 2018;553(7689):446–454. doi:10.1038/nature25183
5. Lemjabbar-Alaoui H, Hassan OU, Yang YW, Buchanan P. Lung cancer: biology and treatment options. Biochim Biophys Acta. 2015;1856(2):189–210.
6. Huang Y, Guo Q, Ding XP, Wang X. Mechanism of long noncoding RNAs as transcriptional regulators in cancer. RNA Biol. 2020;1–13.
7. Jiang L, Li Z, Wang R. Long noncoding RNAs in lung cancer: regulation patterns, biologic function and diagnosis implications (review). Int J Oncol. 2019;55(3):585–596.
8. Ye JJ, Cheng YL, Deng JJ, Tao WP, Wu L. LncRNA LINC00460 promotes tumor growth of human lung adenocarcinoma by targeting miR-302c-5p/FOXA1 axis. Gene. 2019;685:76–84. doi:10.1016/j.gene.2018.10.058
9. Cui H, Zhao J. LncRNA TMPO-AS1 serves as a ceRNA to promote osteosarcoma tumorigenesis by regulating miR-199a-5p/WNT7B axis. J Cell Biochem. 2020;121(3):2284–2293. doi:10.1002/jcb.29451
10. Huang W, Su X, Yan W, et al. Overexpression of AR-regulated lncRNA TMPO-AS1 correlates with tumor progression and poor prognosis in prostate cancer. Prostate. 2018;78(16):1248–1261. doi:10.1002/pros.23700
11. Qin Z, Zheng X, Fang Y. Long noncoding RNA TMPO-AS1 promotes progression of non-small cell lung cancer through regulating its natural antisense transcript TMPO. Biochem Biophys Res Commun. 2019;516(2):486–493. doi:10.1016/j.bbrc.2019.06.088
12. Mu X, Wu H, Liu J, et al. Long noncoding RNA TMPO-AS1 promotes lung adenocarcinoma progression and is negatively regulated by miR-383-5p. Biomed Pharmacother. 2020;125:109989. doi:10.1016/j.biopha.2020.109989
13. Orom UA, Lund AH. Isolation of microRNA targets using biotinylated synthetic microRNAs. Methods. 2007;43(2):162–165. doi:10.1016/j.ymeth.2007.04.007
14. Yang J, Liang B, Hou S. TMPO-AS1 promotes cervical cancer progression by upregulating RAB14 via sponging miR-577. J Gene Med. 2019;21(11):e3125. doi:10.1002/jgm.3125
15. Wu Y, Cheng K, Liang W, Wang X. lncRNA RPPH1 promotes non-small cell lung cancer progression through the miR-326/WNT2B axis. Oncol Lett. 2020;20(4):105. doi:10.3892/ol.2020.11966
16. Yu W, Jiang H, Zhang H, Li J. Hsa_circ_0003998 promotes cell proliferation and invasion by targeting miR-326 in non-small cell lung cancer. Onco Targets Ther. 2018;11:5569–5577. doi:10.2147/OTT.S174750
17. Chen J, Xu S, Chen S, et al. CircPUM1 promotes the malignant behavior of lung adenocarcinoma by regulating miR-326. Biochem Biophys Res Commun. 2019;508(3):844–849. doi:10.1016/j.bbrc.2018.11.176
18. Meng Q, Liu M, Cheng R. LINC00461/miR-4478/E2F1 feedback loop promotes non-small cell lung cancer cell proliferation and migration. Biosci Rep. 2020;40(2):BSR20191345. doi:10.1042/BSR20191345
19. Shi J, Li J, Yang S, et al. LncRNA SNHG3 is activated by E2F1 and promotes proliferation and migration of non-small-cell lung cancer cells through activating TGF-beta pathway and IL-6/JAK2/STAT3 pathway. J Cell Physiol. 2020;235(3):2891–2900.
20. Zhan Y, Zang H, Feng J, Lu J, Chen L, Fan S. Long non-coding RNAs associated with non-small cell lung cancer. Oncotarget. 2017;8(40):69174–69184. doi:10.18632/oncotarget.20088
21. Xia H, Jing H, Li Y, Lv X. Long noncoding RNA HOXD-AS1 promotes non-small cell lung cancer migration and invasion through regulating miR-133b/MMP9 axis. Biomed Pharmacother. 2018;106:156–162. doi:10.1016/j.biopha.2018.06.073
22. Li S, Mei Z, Hu HB, Zhang X. The lncRNA MALAT1 contributes to non-small cell lung cancer development via modulating miR-124/STAT3 axis. J Cell Physiol. 2018;233(9):6679–6688.
23. Yu PF, Wang Y, Lv W, et al. LncRNA NEAT1/miR-1224/KLF3 contributes to cell proliferation, apoptosis and invasion in lung cancer. Eur Rev Med Pharmacol Sci. 2019;23(19):8403–8410.
24. Zhang EB, Yin DD, Sun M, et al. P53-regulated long non-coding RNA TUG1 affects cell proliferation in human non-small cell lung cancer, partly through epigenetically regulating HOXB7 expression. Cell Death Dis. 2014;5(5):e1243. doi:10.1038/cddis.2014.201
25. Mitobe Y, Ikeda K, Suzuki T, et al. ESR1-stabilizing long noncoding RNA TMPO-AS1 promotes hormone-refractory breast cancer progression. Mol Cell Biol. 2019;39(23):e00261–19. doi:10.1128/MCB.00261-19
26. Wang R, Chen X, Xu T, et al. MiR-326 regulates cell proliferation and migration in lung cancer by targeting phox2a and is regulated by HOTAIR. Am J Cancer Res. 2016;6(2):173–186.
27. Cai M, Wang Z, Zhang J, et al. Adam17, a target of Mir-326, promotes emt-induced cells invasion in lung adenocarcinoma. Cell Physiol Biochem. 2015;36(3):1175–1185.
28. Sun C, Huang C, Li S, et al. Hsa-miR-326 targets CCND1 and inhibits non-small cell lung cancer development. Oncotarget. 2016;7(7):8341–8359. doi:10.18632/oncotarget.7071
29. Wang L, Hu F, Shen S, et al. Knockdown of SOX12 expression inhibits the proliferation and metastasis of lung cancer cells. Am J Transl Res. 2017;9(9):4003–4014.
30. Chen X, Xu H, Sun G, Zhang Y. LncRNA CASC9 affects cell proliferation, migration, and invasion of tongue squamous cell carcinoma via regulating miR-423-5p/SOX12 axes. Cancer Manag Res. 2020;12:277–287. doi:10.2147/CMAR.S220351
31. Yang C, Liu Z, Chang X, et al. NR2F1-AS1 regulated miR-423-5p/SOX12 to promote proliferation and invasion of papillary thyroid carcinoma. J Cell Biochem. 2020;121(2):2009–2018. doi:10.1002/jcb.29435
32. Wu L, Timmers C, Maiti B, et al. The E2F1-3 transcription factors are essential for cellular proliferation. Nature. 2001;414(6862):457–462. doi:10.1038/35106593
33. Lazzerini Denchi E, Helin K. E2F1 is crucial for E2F-dependent apoptosis. EMBO Rep. 2005;6(7):661–668. doi:10.1038/sj.embor.7400452
34. Ren X, Chen C, Luo Y, et al. lncRNA-PLACT1 sustains activation of NF-kappaB pathway through a positive feedback loop with IkappaBalpha/E2F1 axis in pancreatic cancer. Mol Cancer. 2020;19(1):35.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.