Back to Journals » Neuropsychiatric Disease and Treatment » Volume 15
Thymosin β4 prevents oxygen-glucose deprivation/reperfusion-induced injury in rat cortical neurons
Authors Zhang Z, Liu S, Huang S
Received 13 March 2019
Accepted for publication 23 July 2019
Published 22 August 2019 Volume 2019:15 Pages 2385—2393
DOI https://doi.org/10.2147/NDT.S208600
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Yu-Ping Ning
Zhongsheng Zhang, Shuangfeng Liu, Sichun Huang
Department of Neurology, The 6th Affiliated Hospital of Guangzhou Medical University, Qingyuan People’s Hospital, Qingyuan 511518, People’s Republic of China
Correspondence: Zhongsheng Zhang
Department of Neurology, The 6th Affiliated Hospital of Guangzhou Medical University, Qingyuan People’s Hospital, Qingyuan 511518, People’s Republic of China
Tel +86 135 3952 3445
Email [email protected]
Purpose: This study investigated whether thymosin (T) β4 protects against oxygen-glucose deprivation/reperfusion (OGD/R) injury in rat cortical neurons, as well as the underlying mechanisms.
Methods: Primary rat cortical neurons were transfected with Tβ4 overexpression plasmid; the transfection efficiency was confirmed by detecting Tβ4 expression by fluorescence quantitative PCR and Western blotting. The OGD/R model was established and apoptotic cells were quantified by flow cytometry and terminal deoxynucleotidyl transferase dUTP nick end labeling. Structural changes in the endoplasmic reticulum were visualized by transmission electron microscopy. The expression levels of 78-kDa glucose-regulated protein (GRP) 78, C/EBP-homologous protein (CHOP), B-cell lymphoma (Bcl)-2, and Bcl-2-associated X protein (Bax) were determined by Western blotting. The effect of Tβ4 on OGD/R injury was evaluated by adding exogenous Tβ4 to neuronal cultures.
Results: Cortical neurons were identified by the expression of neuron-specific enolase. In OGD/R cells, the rate of apoptosis was increased and GRP78, CHOP, and Bax were upregulated whereas Bcl-2 was downregulated relative to the control group. These effects were reversed by Tβ4 overexpression. Endoplasmic reticulum (ER) stress was observed in the OGD/R group, but this was abolished in neurons overexpressing Tβ4. The protective effect of Tβ4 against OGD/R injury was also demonstrated in cells treated with exogenous Tβ4 (10 ng/mL), which blocked OGD/R-induced apoptosis by inhibiting ER stress-related and pro-apoptotic protein expression.
Conclusion: Tβ4 prevents OGD/R-induced ER stress-dependent apoptosis in cortical neurons, and is a potential treatment for cerebral ischemia-reperfusion injury.
Keywords: Tβ4, cortical neuron culture, OGD/R, endoplasmic reticulum stress
Introduction
Cerebral ischemia injury encompasses neurological deficits resulting from focal or whole cerebral ischemia and hypoxia due to the disruption of cerebrovascular circulation. Clinical manifestations include sudden restriction of limb movement, language and sensory impairment (e.g., vision and hearing), vertigo, tinnitus, and dysphagia. Additionally, hemiplegia, unconsciousness, and loss of self-care ability can result in cases where infarction involves a large area or occurs in key brain areas. Ischemia leads to tissue damage and organ dysfunction, with the duration and severity determining the physiological response and probability of survival.1
Mitochondria are the first organelles to respond to cerebral ischemia and hypoxia, which force the change from aerobic to anaerobic respiration. The latter produces only a small amount of ATP but a large amount of lactic acid, leading to acidosis and causing damage to brain tissue.2 At the same time, excitatory neurotransmitters released from nerve terminals activate their cognate postsynaptic receptors, contributing to osmotic swelling and neuronal damage. ATP depletion also disrupts the normal metabolism of neurons and glia, causing the accumulation of free radicals that destroy the membrane of organelles including the endoplasmic reticulum (ER).3 Thus, cerebral ischemic injury can induce ER stress in neurons.
Thymosin (T) β4 is a polypeptide composed of 43 amino acids that is widely distributed in the body. Tβ4 is purified from the thymus4 and was shown to enhance angiogenesis and wound healing by promoting the differentiation and migration of vascular endothelial cells;5 treatment with Tβ4 for 4 days potently stimulated angiogenesis in diabetic rats.6 Tβ4 also increased myocardial microvessel density and the expression of vascular endothelial growth factor in patients with myocardial infarction.5 The protective effect of Tβ4 has also been reported in animal models of stroke.7,8 However, it remains unclear how Tβ4 prevents ischemic injury. This was investigated in the present study in an oxygen-glucose deprivation/reperfusion (OGD/R)-induced neuronal injury model established in primary cultures of rat cortical neurons.
Materials and methods
Cortical neuron cultures
Neonatal rats were sacrificed within 48 h and after sterilization with 75% alcohol, the skull was opened with ophthalmic forceps. The brain tissue was dissected and placed in pre-cooled Hank’s equilibrium salt solution and cut into pieces about 1 mm3 in size and digested in 0.125% trypsin at 37 °C. Dissociated cells were cultured in neurobasal medium supplemented with fetal bovine serum (FBS; Hyclone, Logan, UT, USA), B27, and β-nerve growth factor. After filtration through a 70-mesh sieve, the cells were centrifuged and resuspended in fresh Dulbecco’s Modified Eagle’s Medium containing 10% FBS. All animals were treated in accordance with the Guide for the Care and Use of Laboratory Animals, and all experiments were approved and performed according to the guidelines of the Ethics Committee of Ethics Committee of Qingyuan People’s Hospital (Qingyuan, China).
Immunofluorescence analysis
Cells cultured in dishes were washed three times with phosphate-buffered saline (PBS) for 3 min each and fixed in 4% paraformaldehyde (PFA) for 15 min. After washing three times with PBS, the cells were incubated with 0.5% Triton X-100 in PBS for 20 min at room temperature and blocked in 5% bovine serum albumin at 37 °C for 30 min. The cells were incubated overnight at 4 °C with primary antibody against neuron-specific enolase (NSE) (1:200, Abcam, Cambridge, UK). After three washes for 15 min each in PBS, the samples were incubated with fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit IgG (Life Technologies, Carlsbad, CA, USA) for 2 h at room temperature. Cell nuclei were stained with 3,3ʹ-diaminobenzidine for 3 min at room temperature, and images were acquired on a confocal laser scanning microscope (FV1000; Olympus, Tokyo, Japan).
Cell transfection
When cells reached 70% confluence, the culture medium was replaced with serum-free medium and 125 μl of Opti-MEM was added along with 5 μl of Lipofectamine 3000 and 2.5 μg of plasmid harboring the Tmsb4x gene for Tβ4 overexpression or empty plasmid as a control. The mixture was incubated at room temperature for 5 min and divided into two equal parts that were added dropwise into appropriate wells of a 12-well plate. The cells were incubated at 37 °C for 4 h, and the medium was replaced with complete medium containing 20% FBS.
Fluorescence quantitative PCR
Total RNA was extracted using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA), and cDNA was synthesized with a reverse transcription kit (Promega, Madison, WI, USA) and used as the template for fluorescence quantitative PCR. The expression level of Tβ4 was calculated relative to that of β-actin (internal reference). The reaction contained RNase-free dH2O (9.5 μl), cDNA (1 μl), primers (2 μl), and 2× ULtraSYBR mixture (12.5 μl), and the reaction conditions were as follows: 95 °C for 10 min, followed by 40 cycles of 95 °C for 10 s, 58.5 °C for 30 s, and 72 °C for 30 s. The forward and reverse primer sequences were as follows: Tmsb4x, 5ʹ-GACAAACCCGATATGGCTGA-3ʹ and 5ʹ-GTTTCTTTTGAAGGCAGAGGAT-3ʹ; and β-actin, 5ʹ-AGGGAAATCGTGCGTGAC-3ʹ and 5ʹ-ATACCCAGGAAGGAAGGCT-3ʹ.
OGD/R model
The OGD/R model was established as previously described.9 Briefly, cells grown to 70% confluence were deprived of oxygen and glucose for 6 h and then reoxygenated for 12 h. The cells were divided into six groups: control (untransfected cells), Tβ4 overexpression, empty vector, OGD/R, OGD/R+empty vector, and OGD/R+Tβ4 overexpression.
To investigate the effect of exogenously applied Tβ4, cells were divided into control, model (OGD for 6 h, reoxygenation for 12 h), and Tβ4 (10 ng/mL for 12 h, OGD for 6 h, reoxygenation for 12 h) groups. Cell viability was evaluated with Cell Counting Kit (CCK)-8, and apoptosis was detected by flow cytometry and the terminal dexynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay. The expression of 78-kDa glucose-regulated protein (GRP)78, C/EBP-homologous protein (CHOP), B-cell lymphoma (Bcl)-2, and Bcl-2-associated X protein (Bax) was evaluated by Western blotting.
Western blotting
Western blotting was carried out as previously described.11 Briefly, protein was extracted from cells using the ReadyPrep protein isolation kit (GE Healthcare, Little Chalfont, UK) and the concentration was determined using a bicinchoninic assay kit (Thermo Fisher Scientific); 20 μg of protein were loaded into each lane of a 12% polyacrylamide gel and separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis, then transferred to a nitrocellulose membrane. After blocking in 5% skim milk for 2 h at room temperature, the membrane was incubated overnight at 4 °C with the following primary antibodies: rabbit polyclonal anti-Tβ4 (Affinity, Cincinnati, USA; cat. no. DF12334, 1:1000), rabbit polyclonal anti-GRP78 (ImmunoWay, Plano, TX, USA; cat. no. YT5858, 1:1000), rabbit polyclonal anti-CHOP (Abcam; cat. no. ab179823, 1:3000), rabbit polyclonal anti-Bax (Abcam; cat. no. ab32503; 1:5000), and mouse monoclonal anti-Bcl-2 (Bioss Antibodies, Woburn, MA, USA; bsm-33047M, 1:500). The membrane was washed three times and incubated for 2 h at 4 °C with horseradish peroxidase-labeled goat anti-rabbit IgG secondary antibody (Thermo Fisher Scientific; cat. no. A16104SAMPLE, 1:10,000). Protein expression was visualized using an enhanced chemiluminescence kit (Thermo Fisher Scientific) and the blots were scanned using a ChemiDoc XRS imager (Bio-Rad, Hercules, CA, USA). Densitometric analysis was performed with ImageJ v.7.0 software (National Institutes of Health, Bethesda, MD, USA).
CCK-8
After treatment with exogenous Tβ4 for 48 h, cell viability was evaluated with the CCK-8 assay (Gibco, Grand Island, NY, USA) as previously described.10 The formazan crystals were dissolved in dimethyl sulfoxide and absorbance was measured with a microplate reader (Thermo Fisher Scientific) at a wavelength of 450 nm.
Flow cytometry
Cells were collected after digestion with trypsin (Gibco) and incubated with Annexin V-FITC and propidium iodide (Beyotime, Ningbo, China; C1062) for 30 min in the dark. Apoptotic cells were detected by flow cytometry (BD Bioscience, Franklin Lakes, NJ, USA) and data were analyzed with FlowJo 10 software (Tree Star, Ashland, OR, USA).
Tunel
Cells cultured in dishes were washed three times with PBS for 3 min each and fixed in 4% PFA for 15 min. After three washes with PBS, the cells were incubated with 0.5% Triton X-100 in PBS for 20 min at room temperature, and apoptotic cells were detected with the TUNEL assay as previously described.11,12
Transmission electron microscopy (TEM)
After treatment, cells were fixed with 2.5% glutaraldehyde and then dehydrated, embedded, and cut into 70-nm sections that were stained with 3% uranyl acetate and lead citrate and imaged by TEM at 80 kV (JEM-1230; JEOL, Tokyo, Japan).
Statistical analysis
Data are expressed as mean ± SD and were statistically analyzed using SPSS 19 (IBM, Armonk, NY, USA). One-way analysis of variance followed by the Newman–Keuls post-hoc test was used to evaluate statistical significance, which was defined as P<0.05.
Results
Tβ4 overexpression prevents OGD/R-induced apoptosis in rat cortical neurons
The purity of the cortical neuron culture was confirmed by immunofluorescence detection of NSE (Figure 1). The transfection efficiency of the Tmsb4x overexpression plasmid was confirmed by fluorescence quantitative PCR and Western blotting (Figure 2). Compared with the control group, Tmsb4x mRNA (Figure 2A) and protein (Figure 2B) levels were higher in Tmsb4x-overexpressing cells (P<0.05). Meanwhile, apoptosis was increased by OGD/R (P<0.05), as determined by flow cytometry (Figure 3A) and the TUNEL assay (Figure 3B). However, the number of apoptotic cells was decreased in the Tβ4 group (P<0.05).
 |
Figure 1 Isolation and identification of rat cortical neurons. The expression of NSE in cortical neurons was evaluated by quantitative PCR and Western blotting. Scale bar: 100 μm. |
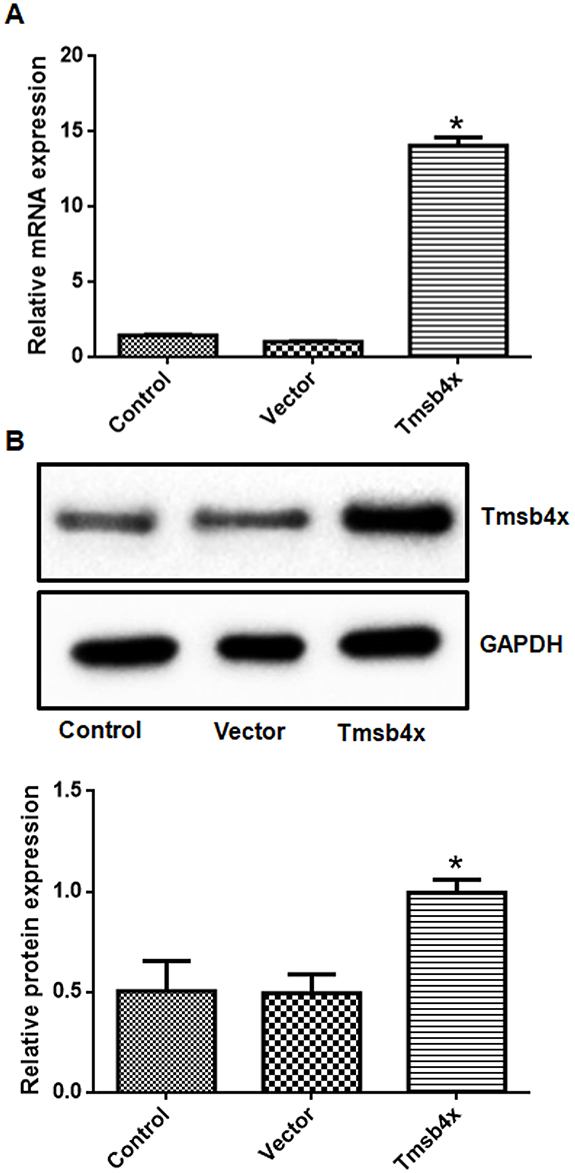 |
Figure 2 Evaluation of transfection efficiency. (A, B) Transfection of the Tmsb4x overexpression plasmid increased Tβ4 mRNA (A) and protein (B) levels in cortical neurons. Data shown as mean ± SD. *P<0.05 vs control. |
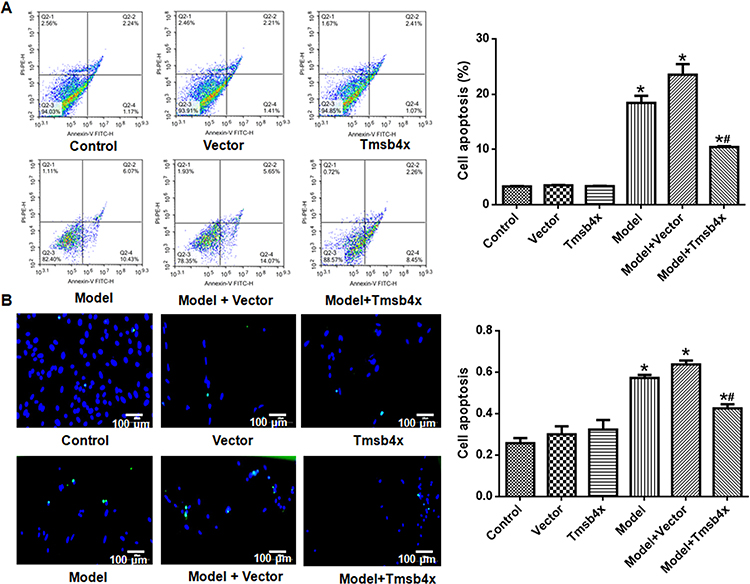 |
Figure 3 Tβ4 overexpression prevents OGD/R-induced apoptosis in rat cortical neurons. (A) Detection of apoptotic cells by flow cytometry. Left: representative flow cytometry plots; right: quantitative analysis. (B) Detection of apoptotic cells with the TUNEL assay. Left: representative images (apoptotic cells are green and cell nuclei are blue); right: quantitative analysis. Data shown as mean ± SD. *P<0.05 vs control; #P<0.05 vs OGD/R. |
GRP78, CHOP, and Bax expression was upregulated whereas the anti-apoptotic factor Bcl-2 was downregulated by OGD/R relative to the control group (Figure 4). Tβ4 overexpression reversed these changes (P<0.05).
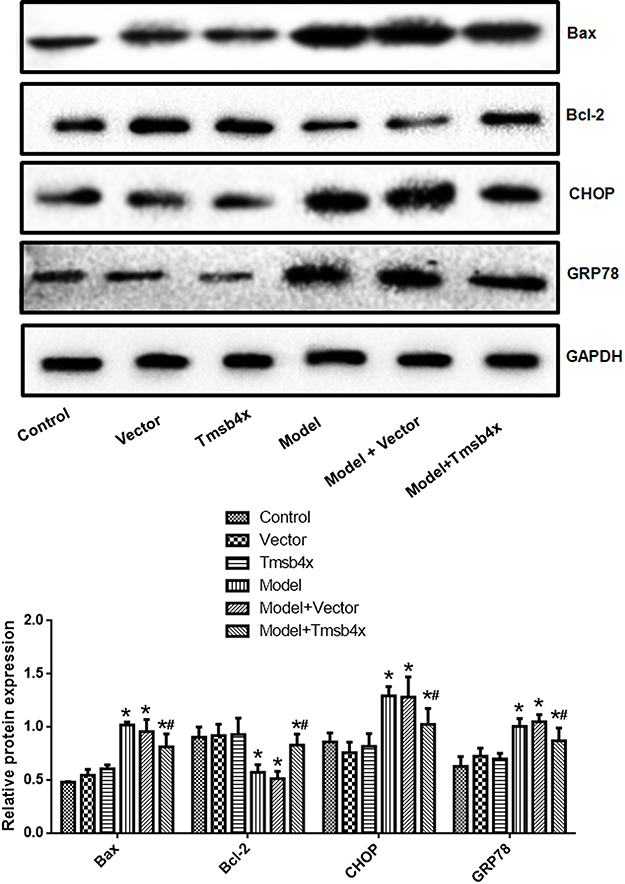 |
Figure 4 Tβ4 overexpression reverses OGD/R-induced changes in ER and apoptosis-related protein expression. Upper panel: representative immunoblots; lower panel: quantitative analysis. Data shown as mean ± SD. *P<0.05 vs control; #P<0.05 vs OGD/R. |
Tβ4 overexpression improves OGD/R-induced ER stress
Transmission electron micrographs of cortical neurons in control, empty vector, Tβ4 overexpression, and OGD/R+Tβ4 overexpression groups showed a regular arrangement of ER cisternae throughout the cytoplasm. In contrast, ER cisternae were fragmented and highly disorganized in the OGD/R and OGD/R+empty vector groups (Figure 5).
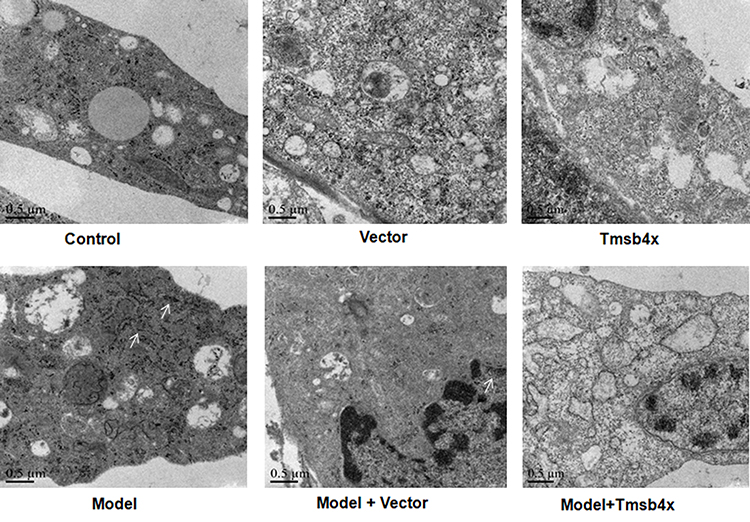 |
Figure 5 Tβ4 overexpression alleviates OGD/R-induced ER stress. ER structure was evaluated by TEM. Arrows indicate ER with abnormal morphology. |
Exogenous Tβ4 prevents OGD/R-induced apoptosis
Cell viability was decreased (Figure 6A) whereas apoptosis was increased (Figure 6B and C) by OGD/R (P<0.05), but these effects were reversed by exogenous application of Tβ4 (10 ng/mL) (P<0.05). Similarly, increases in the levels of GRP78, CHOP, and Bax and decrease in that of Bcl-2 induced by OGD/R were abolished by Tβ4 treatment (P<0.05; Figure 7).
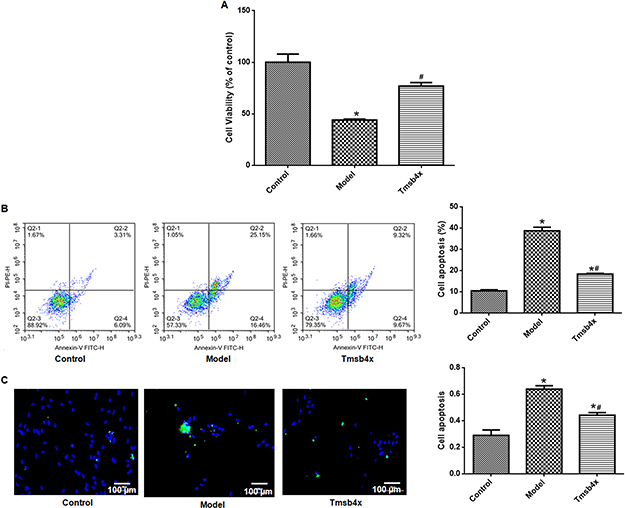 |
Figure 6 Exogenous Tβ4 prevents OGD/R-induced apoptosis. (A) Evaluation of cell viability with the CCK-8 assay without or with 10 ng/mL Tβ4 treatment. (B) Detection of apoptotic cells by flow cytometry. Left: flow cytometry plots; right: quantitative analysis. (C) Detection of apoptotic cells with the TUNEL assay. Left: representative images (apoptotic cells are green and cell nuclei are blue); right: quantitative analysis. Data shown as mean ± SD. *P<0.05 vs control; #P<0.05 vs OGD/R. |
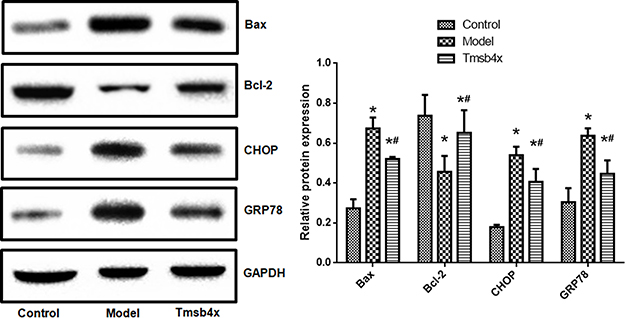 |
Figure 7 Exogenous Tβ4 reverses OGD/R-induced changes in ER and apoptosis-related protein expression. Upper panel: representative immunoblots; lower panel: quantitative analysis. Data shown as mean ± SD. *P<0.05 vs control; #P<0.05 vs OGD/R. |
Discussion
This study investigated the therapeutic potential of Tβ4 for preventing ischemia/reperfusion-associated brain injury using an in vitro OGD/R model established with primary cortical neurons. Our results demonstrate that both Tβ4 overexpression and exogenous Tβ4 application prevented OGD/R-induced apoptosis and ER stress in the neurons. These results provide experimental evidence for the efficacy of Tβ4 treatment for cerebral ischemia-reperfusion injury.
Tβ4 has a variety of pharmacological activities including the stimulation of angiogenesis,13 nerve axon repair,14 and inhibition of the inflammatory response,15 and is therefore a candidate molecule for the repair and regeneration of vascular nerve units following cerebral ischemia. Tβ4 expression after myocardial infarction inhibits myocardiocyte apoptosis and restores cardiac function;16 it was also found to be upregulated in the hippocampus after focal and global cerebral ischemia.17 Additionally, intraperitoneal injection of synthetic Tβ4 increased vascular density in the ischemic penumbra and accelerated nerve axon repair and functional recovery.18
Tβ4 promotes the migration, proliferation, and apoptosis of endothelial progenitor cells (EPCs) through the phosphoinositide 3-kinase (PI3K)/Akt/endothelial nitric oxide synthase pathway. Blocking PI3K/Akt signaling reduced the effect of Tβ4 on EPCs.19 Bone marrow stem cells participate in vascular regeneration following stroke.20 However, local inflammation after stroke can injure EPCs. Tβ4 suppresses inflammation-induced EPC apoptosis, providing a microenvironment that promotes post-stroke vascular regeneration.20 Combined with findings from other studies,19,21 our data indicate that Tβ4 not only stimulates vascular regeneration but also enhances neuronal survival after stroke.
Neuronal apoptosis is one of the main forms of cerebral ischemia-reperfusion injury. Bcl-2 family proteins play a key role in regulating apoptosis.22 The anti-apoptotic Bcl-2 is mainly localized in the outer membrane of mitochondria, nuclear membrane, and ER. Bcl-2 blocks the release of cytochrome c and inhibits the downstream apoptotic cascade.23 Bax is present in the cytoplasm and forms a heterodimer with Bcl-2, thereby blocking its activity and stimulating the cytochrome c cascade.24 Downregulation of Bcl-2 and upregulation of Bax have been reported in cerebral ischemic injury25 and were observed in our OGD/R model, which is consistent with previous findings.26 Tβ4 overexpression or exogenous application increased the level of Bcl-2 while decreasing that of Bax, indicating that apoptosis was suppressed.
GRP78 is a heat shock protein family member that functions as a chaperone in the ER. Under physiological conditions GRP78 participates in the folding, assembly, and transport of newly synthesized polypeptide chains27 but upon cellular stress caused by ischemia, hypoglycemia, or calcium imbalance or oxidative stress, polypeptide chains degenerate and accumulate in the ER, with a concomitant increase in GRP78 expression.28 The upregulation of GRP78 in the kidney, brain, retina, lung, myocardium, and other tissues during ischemia-reperfusion29 counters ER stress.30,31
CHOP is a basic leucine zipper transcription factor belonging to the C/EBP family of CCAATP enhancer-linked proteins. CHOP is normally expressed at a low level but is upregulated under ER stress,27 causing cell cycle arrest or apoptosis; conversely, loss of CHOP function protects cells against apoptosis.32 Our results demonstrate that GRP78 and CHOP levels were increased by OGD/R but were decreased by Tβ4 treatment. These results confirm that Tβ4 inhibits apoptosis induced by ischemic injury. Moreover, the TEM analysis revealed that ER structure, which was undermined by OGD/R, was restored by Tβ4 overexpression. Thus, Tβ4 counters the ER stress response to ischemia-reperfusion by modulating the expression of GRP78, CHOP, Bcl-2, Bax, and other proteins, thereby preventing tissue injury.
In vivo studies have demonstrated the protective effects of Tβ4 against ischemic brain injury in adult and aged animals; moreover, an optimal dose of 3.75 mg/kg was suggested for preventing post-stroke tissue damage.7,8 Here we showed that Tβ4 relieves ER stress in cortical neurons induced by OGD/R and describe the possible underlying mechanisms. Whether Tβ4 has a similar effect in vivo remains to be determined.
Conclusion
The results of this study demonstrate that Tβ4 prevents OGD/R-induced ER stress-dependent apoptosis in neurons. Our findings provide evidence for the clinical applicability of Tβ4 in the treatment of cerebral ischemia-reperfusion injury.
Acknowledgment
This work was supported by the grants from Industrial Technology and Research and Development Fund of Qingyuan City (2017A022).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Philippart R, Brunet-Bernard A, Clementy N, et al. Oral anticoagulation, stroke and thromboembolism in patients with atrial fibrillation and valve bioprosthesis. The Loire Valley Atrial Fibrillation Project. Thromb Haemost. 2016;115(5):1056–1063. doi:10.1160/TH16-01-0007
2. Smith EE, Saposnik G, Biessels GJ, et al. Prevention of stroke in patients with silent cerebrovascular disease: a scientific statement for healthcare professionals from the American Heart Association/American Stroke Association. Stroke. 2017;48(2):e44–e71. doi:10.1161/STR.0000000000000116
3. Chong WC, Shastri MD, Eri R. Endoplasmic reticulum stress and oxidative stress: a vicious nexus implicated in bowel disease pathophysiology. Int J Mol Sci. 2017;18(4):E771.
4. Shi B, Ding Q, He X, et al. Tbeta4-overexpression based on the piggyBac transposon system in cashmere goats alters hair fiber characteristics. Transgenic Res. 2017;26(1):77–85.
5. Ziegler T, Bahr A, Howe A, et al. Tbeta4 increases neovascularization and cardiac function in chronic myocardial ischemia of normo- and hypercholesterolemic pigs. Mol Ther. 2018;26(7):1706–1714.
6. Akpinar H, Naziroglu M, Ovey IS, Cig B, Akpinar O. The neuroprotective action of dexmedetomidine on apoptosis, calcium entry and oxidative stress in cerebral ischemia-induced rats: contribution of TRPM2 and TRPV1 channels. Sci Rep. 2016;6:37196.
7. Morris DC, Cui Y, Cheung WL, et al. A dose-response study of thymosin beta4 for the treatment of acute stroke. J Neurol Sci. 2014;345(1–2):61–67.
8. Morris DC, Cheung WL, Loi R, et al. Thymosin beta4 for the treatment of acute stroke in aged rats. Neurosci Lett. 2017;659:7–13.
9. Ji H, Xu L, Wang Z, Fan X, Wu L. Effects of thymosin beta4 on oxygenglucose deprivation and reoxygenationinduced injury. Int J Mol Med. 2018;41(3):1749–1755.
10. Song Z, Chen H, Xu W, Wu S, Zhu G. Basolateral amygdala calpain is required for extinction of contextual fear-memory. Neurobiol Learn Mem. 2018;155:180–188. doi:10.1016/j.nlm.2018.08.004
11. Zhu G, Wang X, Wu S, Li Q. Involvement of activation of PI3K/Akt pathway in the protective effects of puerarin against MPP+-induced human neuroblastoma SH-SY5Y cell death. Neurochem Int. 2012;60(4):400–408. doi:10.1016/j.neuint.2012.01.003
12. Li J, Yang S, Zhu G. Postnatal calpain inhibition elicits cerebellar cell death and motor dysfunction. Oncotarget. 2017;8(50):87997–88007. doi:10.18632/oncotarget.21324
13. Trenkwalder T, Deindl E, Bongiovanni D, et al. Thymosin-beta4-mediated therapeutic neovascularization: role of the PI3K/AKT pathway. Expert Opin Biol Ther. 2015;15(Suppl 1):S175–S185. doi:10.1517/14712598.2015.1011122
14. Pipes GT, Yang J. Cardioprotection by thymosin beta 4. Vitam Horm. 2016;102:209–226. doi:10.1016/bs.vh.2016.04.004
15. Lee SI, Yi JK, Bae WJ, Lee S, Cha HJ, Kim EC. Thymosin beta-4 suppresses osteoclastic differentiation and inflammatory responses in human periodontal ligament cells. PLoS One. 2016;11(1):e0146708. doi:10.1371/journal.pone.0146708
16. Santra M, Chopp M, Santra S, et al. Thymosin beta 4 up-regulates miR-200a expression and induces differentiation and survival of rat brain progenitor cells. J Neurochem. 2016;136(1):118–132. doi:10.1111/jnc.13394
17. Morris DC, Chopp M, Zhang L, Lu M, Zhang ZG. Thymosin beta4 improves functional neurological outcome in a rat model of embolic stroke. Neuroscience. 2010;169(2):674–682. doi:10.1016/j.neuroscience.2010.05.017
18. Zhang J, Zhang ZG, Li Y, et al. Thymosin beta4 promotes oligodendrogenesis in the demyelinating central nervous system. Neurobiol Dis. 2016;88:85–95. doi:10.1016/j.nbd.2016.01.010
19. Qiu FY, Song XX, Zheng H, Zhao YB, Fu GS. Thymosin beta4 induces endothelial progenitor cell migration via PI3K/Akt/eNOS signal transduction pathway. J Cardiovasc Pharmacol. 2009;53(3):209–214. doi:10.1097/FJC.0b013e318199f326
20. Seto SW, Chang D, Jenkins A, Bensoussan A, Kiat H. Angiogenesis in ischemic stroke and angiogenic effects of Chinese Herbal medicine. J Clin Med. 2016;5(6):E56
21. Tan WKY, Purnamawati K, Pakkiri LS, et al. Sources of variability in quantifying circulating thymosin beta-4: literature review and recommendations. Expert Opin Biol Ther. 2018;18(sup1):141–147.
22. Carthy CM, Yanagawa B, Luo H, et al. Bcl-2 and Bcl-xL overexpression inhibits cytochrome c release, activation of multiple caspases, and virus release following coxsackievirus B3 infection. Virology. 2003;313(1):147–157.
23. Um HD. Bcl-2 family proteins as regulators of cancer cell invasion and metastasis: a review focusing on mitochondrial respiration and reactive oxygen species. Oncotarget. 2016;7(5):5193–5203.
24. Wang Q, Zhang L, Yuan X, et al. The relationship between the Bcl-2/Bax proteins and the mitochondria-mediated apoptosis pathway in the differentiation of adipose-derived stromal cells into neurons. PLoS One. 2016;11(10):e0163327.
25. Terashi T, Otsuka S, Takada S, et al. Neuroprotective effects of different frequency preconditioning exercise on neuronal apoptosis after focal brain ischemia in rats. Neurol Res. 2019;41:1–9.
26. Liu Y, Wu X, An J, et al. Glaucocalyxin B protects against oxygen-glucose-deprivation/reperfusion-induced neuronal injury in PC-12 cells. J Cell Biochem. 2019;120(4):6137–6144.
27. Lee AS. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods. 2005;35(4):373–381.
28. Ayaub EA, Kolb PS, Mohammed-Ali Z, et al. GRP78 and CHOP modulate macrophage apoptosis and the development of bleomycin-induced pulmonary fibrosis. J Pathol. 2016;239(4):411–425.
29. Zhao Y, Han Y, Bu DF, et al. Reduced AKT phosphorylation contributes to endoplasmic reticulum stress-mediated hippocampal neuronal apoptosis in rat recurrent febrile seizure. Life Sci. 2016;153:153–162.
30. Virrey JJ, Dong D, Stiles C, et al. Stress chaperone GRP78/BiP confers chemoresistance to tumor-associated endothelial cells. Mol Cancer Res. 2008;6(8):1268–1275.
31. Flodby P, Li C, Liu Y, et al. The 78-kD glucose-regulated protein regulates endoplasmic reticulum homeostasis and distal epithelial cell survival during lung development. Am J Respir Cell Mol Biol. 2016;55(1):135–149.
32. Dong B, Zhang X, Fan Y, Cao S, Zhang X. GPR4 knockout improves renal ischemia-reperfusion injury and inhibits apoptosis via suppressing the expression of CHOP. Biochem J. 2017;474(24):4065–4074.
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.