Back to Journals » Drug Design, Development and Therapy » Volume 18
Therapeutic Potential of Natural Resources Against Endometriosis: Current Advances and Future Perspectives
Authors Gu X, Zhou H, Miao M, Hu D, Wang X, Zhou J, Teichmann AT, Yang Y, Wang C
Received 20 February 2024
Accepted for publication 9 July 2024
Published 21 August 2024 Volume 2024:18 Pages 3667—3696
DOI https://doi.org/10.2147/DDDT.S464910
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Xia Gu,1,2,* Hui Zhou,1,* Mengyue Miao,1 Daifeng Hu,1 Xinyue Wang,3 Jing Zhou,4 Alexander Tobias Teichmann,1 Youzhe Yang,1,5 Chunyan Wang1
1Sichuan Provincial Center for Gynaecology and Breast Diseases, The Affiliated Hospital of Southwest Medical University, Luzhou, 646000, People’s Republic of China; 2Department of Gynaecology and Obstetrics, Leshan People’s Hospital, Leshan, 614003, People’s Republic of China; 3The Basic Medical College, Army Medical University, Chongqing, 400038, People’s Republic of China; 4Department of Endocrinology, Chengdu Third People’s Hospital, Chengdu, 610014, People’s Republic of China; 5Academician (Expert) Workstation of Sichuan Province, Luzhou, 646000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Youzhe Yang; Chunyan Wang, The Affiliated Hospital of Southwest Medical University, NO. 25 Taiping Street, Jiangyang District, Luzhou, 646000, People’s Republic of China, Email [email protected]; [email protected]
Abstract: Endometriosis (EMS) is defined as the appearance, growth, infiltration, and repeated bleeding of endometrioid tissue (glands and stroma) outside the uterus cavity, which can form nodules and masses. Endometriosis is a chronic inflammatory estrogen-dependent disease and occurs in women of reproductive age. This disorder may significantly affect the quality of life of patients. The pathogenic processes involved in the development and maintenance of endometriosis remain unclear. Current treatment options for endometriosis mainly include drug therapy and surgery. Drug therapy mainly ties to the use of non-steroidal anti-inflammatory drugs (NSAIDs) and hormonal drugs. However, these drugs may produce adverse effects when used for long-term treatment of endometriosis, such as nausea, vomiting gastrointestinal reactions, abnormal liver and kidney function, gastric ulcers, and thrombosis. Although endometriosis lesions can be surgically removed, the disease has a high recurrence rate after surgical resection, with a recurrence rate of 21.5% within 2 years and 40% to 50% within 5 years. Thus, there is an urgent need to develop alternative or additional therapies for the treatment of endometriosis. In this review, we give a systematic summary of therapeutic multiple component prescriptions (including traditional Chinese medicine and so on), bioactive crude extracts of plants/herbs and purified compounds and their newly found mechanisms reported in literature in recent years against endometriosis.
Keywords: endometriosis, treatment, multiple component prescriptions, extracts of plants/herbs, purified compounds, mechanisms
Introduction
The definition of endometriosis (EMS) is the presence of endometrial tissue outside of the uterine cavity. It is considered as an estrogen-dependent chronic inflammatory disorder that primarily affects pelvic organs, including the ovaries. Although endometriosis is one of the most common benign gynecological disorders, the pathogenesis of endometriosis remains unclear. To date, several theories have been established to explain the origination of endometriosis, the most popular and widely accepted one is Sampson’s retrograde menstruation theory.1 It is now widely accepted that the onset and progression of this disease are associated with estrogen dominance and progesterone resistance,2 a pro-inflammatory pelvic microenvironment,3 increased neovascularization,4 apoptosis dysregulation,5 cell proliferation dysregulation,6 fibrosis,7 migration,8 invasion,9 dysregulation of oxidative stress and anti-oxidative stress,10 and alterations in epigenetic factors11 (Figure 1). Endometriosis can spread like a sandstorm in the ovaries, fallopian tubes, pelvic cavity and intestines. The clinical manifestations include chronic pelvic pain, dysuria, progressively increasing dysmenorrhea, gastrointestinal symptoms, painful intercourse, painful bowel movements during menstruation, infertility, and irregular menstruation.12–17 Based on the histopathology and anatomic location of endometriosis, it can be classified into four subtypes: ovarian endometriosis, superficial endometriosis, deep infiltrative endometriosis (DIE),18 and other sites of endometriosis.19 Current clinical options for the treatment of endometriosis mainly include pharmacological and surgical treatments. The following drugs are commonly used for the treatment of endometriosis: non-steroidal anti-inflammatory drugs (NSAIDs),20,21 combined oral contraceptives pills (COCP),22,23 progesterone,24 mifepristone (selective progesterone receptor modulators, SPRM),25 danazol (pseudo-menopausal therapy),26 gonadotropin-releasing hormone agonist (GnRH-a),27 aromatase inhibitors,28 gonadotropin-releasing hormones (GnRH) antagonists.29,30 These drugs often generate serious adverse effects when used for long-term treatment of endometriosis, such as nausea, vomiting gastrointestinal reactions, abnormal liver and kidney function, gastric ulcers, thrombosis, depression, headache, breast tenderness, weight gain, irregular spotting of vaginal bleeding and dryness, hot flashes, loss of libido and bone.22,24–28,30–32 In addition, drug therapy is ineffective in some patients. The specific mechanisms of action and adverse side effects of the mentioned drugs are shown in Table S1. The purpose of surgical treatment is to remove endometriotic lesions and restore the normal anatomical structure. When the symptoms are not relieved after drug treatment, the local lesions are aggravated, the fertility function is not recovered, and the ovarian endometriosis cysts are larger, it is suitable for surgical treatment. The surgical methods mainly include fertility-preserving surgery, ovary-preserving surgery, and radical surgery (Table S2).29 Clinically, few patients choose radical surgery, and almost all patients choose fertility-preserving surgery, which may lead to a high postoperative recurrence rate. Unlike other ovarian cysts, ovarian chocolate cysts are firmly adherent to the ovarian cortex and stroma, which can make surgical removal of the ovarian cyst wall difficult and lead to inadvertent removal of normal ovarian tissue, ultimately resulting in decreased ovarian reserve function. Even though endometriosis lesions can be surgically removed, the recurrence rate is high, with a 2-year recurrence rate of 21.5% and a 5-year recurrence rate of 40% - 50%.33 Surgery remains the gold standard for the final diagnosis, but the risks of surgery must be weighed against the potential decrease in ovarian reserve, especially in the case of bilateral endometriomas and previous ovarian surgery.31,34 Obviously, the current clinical drug therapy and surgical treatment have been unable to meet the individual treatment needs of patients with endometriosis. Therefore, there is an urgent need to develop new drugs that can achieve higher efficacy, fewer side effects and can be used as long-term treatment options.
 |
Figure 1 Molecular mechanisms and factors involved in the occurrence and development of endometriosis. |
Some therapeutic mixtures or crude preparations made from natural materials, especially from plants, have been used to combat diseases for a long time in human history. The basis of these synergistic effects of the mixtures or crude preparations is bioactive ingredients. With the development of antibiotics in the mid-twentieth century, purified compounds have become more typical in drug formulations. These compounds obtained from natural sources, eg, plants, animals, minerals, and microorganisms, are grouped as natural products (NPs) scientifically. In order to adapt to the environment and improve the competitiveness of survival, organisms interact with the environment to form extremely complex chemical structures in the body after hundreds of millions of years of evolution. These molecules may increase their competitive advantages in the ecosystem in which they operate.35 Natural products have long been a source of new drug development.36 First of all, the use of natural products as “as they are” to treat various diseases has been extensively documented in literature and books since ancient times. Secondly, with the development of industrial technology and the progress of drug development, it has become possible to extract and isolate components with clear structures and special biological activities from plants.37 Moreover, these compounds with clear structures and special biological activities have great potential to be modified. According to the needs of clinical medication, some of its structure can be changed emphatically, so as to improve the characteristics of such compounds.38 Thus, natural products can be used as a powerful pool to produce new therapeutic drugs.36
In this review, a literature search was conducted in the database of Pubmed and Medline. Various terms “endometriosis”, “treatment”, “medication”, “natural product”, “herb/herbal”, “extract”, “phytochemical”, “compound” and “bioactive compound” were used as key words for relevance. The articles were published in English from 2017 to 2023 and were original ones. This review is divided into three parts, namely, multiple component prescriptions (including traditional Chinese medicine and so on), crude extracts of plants/herbs, and purified bioactive compounds and their newly found mechanisms reported in literature against endometriosis.
Multiple Component Prescriptions
Cai’s Neiyi Prescription (CNYP)
Cai’s Neiyi Prescription (CNYP) is created by professor Xiaosun Cai from China. Based on Gui Zhi Fu Ling Pill, CNYP is mixed with shanjiapian (pangolin tablets), zhaojiaoci (spina gleditsiae), and eshu (rhizoma curcumae). Cai’s Neiyi Prescription mainly consists of the following traditional Chinese medicine, baifuling (chinaroot greenbrier rhizome), guizhi (cassia twig), shuizhi (leech), chishao (radix paeoniae rubra), eshu (rhizoma curcumae), shijianchuan (Chinese sage herb), mudanpi (cortex moutan), zhaojiaoci (spina gleditsiae), taoren (peach seed) (Table S3).39 Li et al found that Cai’s Neiyi Prescription reduced the expression of interleukin-1β (IL-1β), IL-6, transforming growth factor-β (TGF-β), and tumor necrosis factor-α (TNF-α) in human endometriotic stromal cells and rat model of endometriosis with autologous transplantation. In addition, CNYP could reduce the expression of anti-apoptotic protein B-cell lymphoma-2 (Bcl-2), and also could upregulate the expression of pro-apoptotic proteins Bcl-2 associated X protein (Bax) and caspase-7 by inhibiting the activation of the ERK/2 signaling pathway.40 In conclusion, CNYP may exert anti-endometriosis effects by inhibiting the inflammatory response and promoting apoptosis.
Danefukang Jiangao
Danefukang Jiangao (DEFK) is a common Chinese patent medicine for the treatment of endometriosis and adenomyosis, which has the effects of activating blood circulation, regulating menstruation, relieving pain, and softening and dispersing nodules.41 DEFK mainly contains radix salviae miltiorrhizae, rhizoma curcumae, sanleng, rhizoma cyperi, rhizoma corydalis (Table S3).42 Zhong et al randomly divided 174 patients with endometriosis into an experimental group (DEFK group) and a control group (mifepristone group), with 87 patients in each group. The levels of inflammatory cytokines (TNF-α, IL-6), and carbohydrate antigen-125 (CA-125) were significantly downregulated in the DEFK group. The patients in DEFK group also showed a higher quality of life (QOL) scores, a lower depression and anxiety self-assessment scale (SDS and SAS) scores compared with the control group (mifepristone).43
Guizhi Fuling Decoction
Guizhi fuling Decoction (GZFLD) is one of the classic Chinese herbal prescriptions, mainly used to treat gynecological diseases. The prescription of GZFLD consists of five herbs: poria cocos, persicae semen, paeoniae radix, cinnamon cortex, and moutan cortex (Table S3).44 Gao et al predicted the potential signaling pathways and targets of GZFLD in the treatment of endometriosis through bioinformatics methods. Further experiments were carried out in vitro and in vivo. According to the results, GZFLD could reduce the expression of inflammatory cytokines (IL-6, IL-1β, TNF-α) in ectopic endometriosis rat model. Meanwhile, the effect of GZFLD on hyperalgesia was supported by a hot plate experiment. What’s more, the nerve fiber density of ectopic endometrial tissue of rats was reduced after the treatment of GZFLD. They hypothesized that GZFLD might relieve endometriosis related pain by reducing the nerve fiber density and exert anti-inflammatory effects through the downregulation of the PI3K/Akt signaling pathway.45
Huangzhi Neiyi Capsule
Huangzhi Neiyi Capsule (HZNYC) is a Chinese medicinal preparation developed by professor Min Xia of Chongqing Hospital of Traditional Chinese Medicine and is mostly used clinically for the treatment of endometriosis. HZNYC is mainly composed of three traditional Chinese medicines, namely, cooked rhubarb, leech, and Cyperus rotundus, which have the functions of activating blood circulation, resolving blood stasis, and relieving pain (Table S3).46 Vascularization plays a vital role in the pathogenesis of endometriosis by providing nutrients and oxygen for the growth of endometriotic lesions. Neovascularization of endometriotic lesions, including angiogenesis, vasculogenesis, and inosculation, is a complex process. Neovascularization involves several growth factors and cytokines, such as fibroblast growth factor (FGF), platelet-derived endothelial cell growth factor (PD-ECGF), angiopoietin (Ang)1/2, matrix metalloproteinase-2 (MMP-2) and matrix metalloproteinase-9 (MMP-9), among which vascular endothelial growth factor (VEGF) is the most vital irritant.4 Ectopic endometrium is in a hypoxic environment during ectopic implantation. Under hypoxic conditions, hypoxia-inducible factor-1 alpha (HIF-1α) fails to undergo ubiquitination and degradation, then undergoes nuclear translocation to bind to promoters on the vascular endothelial growth factor gene, finally increasing the expression of HIF-1α and VEGF in endometriotic lesions.47 Liu et al found that HZNYC could reduce the expression of mRNA of HIF-1α by regulating the HIF-1α/VEGF signaling pathway, which in turn reduced the expression of VEGF in the peritoneal fluid and endometriotic lesions of Sprague-Dawley (SD) rats endometriosis model.48 In conclusion, HZNYC inhibits the progress of endometriosis by suppressing neovascularization.
NeiyiKangfu Tablet
NeiyiKangfu Tablet (NYKFT) is a traditional Chinese preparation made by the Affiliated Hospital of Chengdu University of Traditional Chinese Medicine. The preparation is mainly composed of bichihua, sanguoteng, cooked rhubarb, peach kernel, three prune, coix seed, soil turtle worm, turtle, and other medicinal materials (Table S3). It mainly has the effects of clearing dampness and removing blood stasis, dispersing knot, and relieving pain.49 Raf kinase inhibitory protein (RKIP) is a widely expressed negative regulator of the RAF/MEK/ERK signaling pathway. By directly binding RAF-1 and MEK, RKIP interferes with the formation of the kinase-substrate complex, thereby inhibiting the phosphorylation of MEK, ultimately plays a role in inhibiting RAF/MEK/ERK signaling pathway. It is well known that the RAF/MEK/ERK pathway plays a key role in controlling cell proliferation and protecting cells from apoptosis.49,50 Wen et al demonstrated that NYKFT could promote the expression of Raf kinase inhibitory protein in a dose-dependent manner in vivo and in vitro. The upregulated expression of RKIP inhibited the RAF/MEK/ERK signaling pathway, resulting in the increased expression of Bax and cleaved caspase-3, and the decreased expression of Bcl-2. In addition, NYKFT treatment decreased the secretion of pro-inflammatory cytokines IL-6 and TNF-α in EMS mice compared with the control group.51
Neiyi Prescription of QIU
Neiyi Prescription of QIU (NYPQ) was founded by the late professor Xiaomei Qiu, a famous Chinese doctor. NYPQ is mainly used to treat endometriosis and adenomyosis. It has the effects of dispersing knot and relieving pain, promoting blood circulation, and removing blood stasis. It consists of the following components: baristellia, honeysuckle vine, red vine, Prunella japonicus, hedyotis vulgaris, white hair vine, corydalis yanhusuo, purple salvia miltiorrhiza, willebrand, barley malt, fried hawthorn, fragrant tea vegetable, conch mountain (Table S3).52 Autophagy and angiogenesis are important processes in the development of endometriosis.4,53 Angiogenesis provides nutritional support for the implantation, proliferation, and survival of endometriotic lesions. It is widely believed that the growth of neovascularization is closely related to vascular endothelial growth factor. Beclin 1 and light chain 3 (LC3) are both specific markers of autophagy. Under normal conditions, LC3 protein is present in the cytoplasm as type I. When autophagy is activated, LC3-I is recruited to autophagosomes and subsequently converted to LC3-II.54 Therefore, the level of autophagy could be assessed by comparing the amount of LC3-II or the ratio of LC3-II to LC3-I. Yang et al demonstrated that NYPQ could exert an antiangiogenic effect by reducing VEGF expression in a rat model of surgically induced endometriosis. In addition, NYPQ could induce cell apoptosis by promoting the expression of autophagy-related protein Beclin 1, and by increasing the amount of LC3-II or the ratio of LC3-II/I, ultimately inhibiting the progression of endometriotic lesions. In addition, they observed that NYPQ could promote the apoptosis of human umbilical vein endothelial cells (HUVECs) and inhibit the tubular formation of HUVECs by flow cytometry and lumen formation assay. Anti-angiogenesis and apoptotic effects of NYPQ might be achieved through the activation of the PPARγ/nuclear factor kappa B (NF-κB) signaling pathway.55
Shaofu Zhuyu Decoction
Shaofu Zhuyu Decoction (SZD) is one of the common herbal formulas for the treatment of dysmenorrhea, with the effect of invigorating blood circulation and eliminating blood stasis, warming menstruation, and relieving pain. It consists of ten herbs: zingiberis rhizoma, cortex cinnamomi, corydalis rhizoma, trogopteri feces, angelicae sinensis radix, fructus, radix paeoniae rubra, typhae pollen, myrrha, rhizoma chuanxiong, and foeniculi (Table S3).56 Zhu et al showed that cell proliferation was inhibited, while apoptosis was promoted after SZD was applied to rats with endometriosis. Moreover, SZD treatment reduced the expression level of HIF-1α and its subsequent transcriptional target VEGF, which in turn reduced the angiogenesis and decreased the volume of endometriotic lesions.57
Tokishakuyakusan
Tokishakuyakusan (TSS) is a Japanese traditional herbal medicine. TSS consists of six herbal components including lanceae rhizoma, cnidii rhizoma, paeoniae radix, angelicae, axtractylodis, and hoelen (Table S3).58 Nagira et al observed that TSS inhibited the development of endometriotic lesions in surgically induced BALB/c mice. And the expression of IL-33 mRNA was decreased after the treatment of TSS. Through hot plate assay, they also demonstrated that TSS alleviated hyperalgesia in a mouse model of endometriosis.59 Takeuchi et al illustrated that TSS and one of its components, ferulic acid (FA), reduced IL-8 and VEGF production in human endometrial stromal cells (HESCs).60 In conclusion, TSS inhibits the progress of endometriosis due to anti-inflammatory and anti-angiogenic pharmacological activities.
Crude Extracts of Plants/Herbs
Achillea Cretica L. Aerial Extract
Achillea cretica L. (AC) is a medicinal plant. The extract of AC mainly consists of flavonoids and phenolic acids. Bina et al found that the extract of AC could reduce the volume of ectopic endometrial lesions on a rat model of endometriosis. Furthermore, the expression levels of inflammatory factors (IL-6, TNF-α) and VEGF were decreased.61
Basil (Ocimum Basilicum L.) Leaves Extract
Basil (Ocimum basilicum L.) leaves contain high levels of flavonoids. The extract of basil leaves exhibited anti-proliferative, anti-inflammatory, and antioxidant activities.62 By establishing a mouse model of endometriosis, A’Yuni et al demonstrated that basil leaves extract could inhibit the progression of endometriosis by reducing the expression of VEGF in the vascular endothelium of endometriosis tissues.63
Boswellia Serrata Extract
Boswellia serrata (BS) belongs to a branch of the family Burseraceae. Boswellia serrata is well documented for its anti-inflammatory activity.64 Most of its biological activity is attributed to pentacyclic triterpenoids. Nuclear factor red lineage related factor 2 (Nrf2) regulates the transcription of endogenous antioxidant enzymes. Under basal conditions, the Kelch-like ECH-associated protein 1 (Keap1) forms a complex with Nrf2, leading to ubiquitination and subsequent degradation of Nrf2. Under conditions of oxidative stress, Keap1-CUL3-E3 ubiquitin ligase undergoes conformational changes, which disrupt the binding of Nrf2 to Keap1. Free Nrf2 translocates to the nucleus and transcribes antioxidant enzyme genes, NAD(P)H quinone oxidoreductase (NQO1) and heme oxygenase-1 (HO-1).65 R. D’Amico et al demonstrated that after the treatment of BS gum resin extract on the Sprague-Dawley rat model of endometriosis, the expression of Nrf2, HO-1, and NQO-1 proteins in ectopic lesions was increased compared with that of the model group. The study also demonstrated that oral BS gum resin extract therapy could reduce Bcl-2 expression and increase the level of Bax in ectopic endometrial lesions. Moreover, it could increase the cleavage rate of caspase-9 and the cleavage of apoptotic marker PARP.66
Calligonum Comosum (Escanbil) Extract
Calligonum comosum (Escanbil) is a plant that mainly grows in desert areas,67 with various pharmacological activities, such as anti-proliferation, pro-apoptosis,68 anti-oxidation,69 anti-cancer70 and anti-inflammation.71 Kiani’s team demonstrated that calligonum comosum extract could inhibit the migration and tubular formation of human dermal microvascular endothelial cells (HDMEC). Calligonum comosum extract inhibited the growth of ectopic endometrial lesions, the formation of blood vessels and the infiltration of immune cells (macrophages and lymphocytes) in the animal model.72
Cyperi Rhizoma Extract
Cyperi rhizome is a medicinal plant, belonging to the rhizome of Cyperus rotundus L. Cyperi rhizome grows naturally all over the world, especially in tropical, subtropical, and temperate regions. Cyperi rhizome is also a common traditional Chinese herbal medicine.73 Cyperi rhizoma extract (CRE) exhibits a wide range of biological properties, including anti-inflammatory, anti-platelet, antioxidant, pro-apoptotic, anti-proliferative, and estrogenic properties.74–77 The adhesion molecules p-calmodulin and MMP-2 are involved in the regulation of cell-cell adhesion, cell-ECM adhesion, and adhesion of endometriotic cells and peritoneal mesothelial cells during the development of endometriosis. In addition, their expression is upregulated in endometriotic lesions.78–80 Neurotrophins, a family of nerve growth factors, such as nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophic factor 3 (NT-3), and neurotrophic factor 4/5 (NT-4/5), are considered to be endometriosis-associated pain factors. These factors are overexpressed in endometriosis.81 Ahn et al demonstrated that CRE could significantly downregulate the mRNA expression of the adhesion molecules p-cadherin and MMP-2 in 12Z cells. In addition, the mRNA expression of neurotrophins such as BDNF, NGF, NT-3, and NT-4/5 was significantly decreased after the treatment with CRE in 12Z cells. CRE inhibited the progression of endometriosis through the regulation of the Akt and NF-κB pathways.82
Extract of Polygala Fallax Hemsl
Polygala fallax Hemsl (PFH) is generally recognized as a unique edible and medicinal plant in China. In the Guilin city of Guangxi province of China, people often stew their roots with meat to treat endometriosis as a dietary therapy.83 Zhong’s team isolated and cultured human ectopic endometrial stromal cells (HEcESCs) from fresh ectopic endometrial tissue (ovarian endometrial tumor wall) of patients diagnosed with ovarian endometriosis. Through flow cytometry, Western blotting, transwell assay and some other experiments, they found that PFH extract (total flavonoids of Polygala fallax Hemsl, PFHF) could inhibit the proliferation, migration, invasion, and promote apoptosis of HEcESCs by inhibiting the PI3K/Akt signaling pathway.84
Pueraria Flower Extract
Pueraria lobata is a basic traditional Chinese herb. Pueraria flower is rich in isoflavones. Its health benefits partially may be due to isoflavones in the extract.85–87 Kim et al showed that pueraria flower extract (PFE) exposure attenuated the adhesion of immortalized human endometriotic cells (11Z and 12Z cells) to human mesothelial cells Met5A. Furthermore, treatment with PFE significantly inhibited 11Z and 12Z cells migration through scratch (wound healing) assay and transwell-migration assay.88 Previous relevant studies showed that MMP-2 and MMP-9 play a critical role in the progression of adhesion, invasion, and neovascularization of ectopic endometrial tissue.79,80,89 The protein and mRNA levels of MMP-2 and MMP-9 were decreased after treated with PFE in endometriosis cells. Besides, in vivo study, PFE treatment significantly inhibited the formation of endometriotic lesions in autologous transplanted mouse models of endometriosis. Kim et al summarized that PFE inhibited the adhesion and migration of endometriotic cells and suppressed the establishment of ectopic endometrial lesion tissue by downregulating MMP-2 and MMP-9 levels, and by regulating the ERK1/2 signaling pathway.88
Saponin Extract
Korean red ginseng is commonly used in traditional oriental medicine, and it possesses various pharmacological activities such as anti-inflammatory,90 autophagy-promoting,91 antioxidant,92 and antitumor93 effects. Increasing evidence suggests that aberrant microRNA (miRNA) expression is associated with the pathogenesis of endometriosis.94–96 Cleaved caspase-3 is a traditional marker of activated cell death.97,98 Park’s team demonstrated that saponin extract exposure dramatically decreased miR-21-5p expression in HESCs and upregulated caspase-3 expression, which in turn promoted cell apoptosis. HESCs transfected with miR-21-5p inhibitor could upregulate the expression of caspase-3 and could promote apoptosis in HESCs. They also analyzed that modulating the expression level of miRNA might be one of the molecular mechanisms by which saponin extract exerted its anti-endometriosis effect.99
Zingiber Officinale Roscoe (Ginger) Extract
Zingiber officinale Roscoe (Ginger) is classified into the Zingiberaceae family and is a spice commonly used in the daily diet. Ginger is rich in several chemical constituents and shows various pharmacological properties, including anti-inflammatory, antibacterial, anticancer, and antioxidant properties.100 The biological activities of ginger are mainly due to its phenols. Filho et al suggested that ginger extract reduced the volume of ectopic endometrial lesions in Wistar rats, a model of surgically induced endometriosis. However, the expression level of IL-6 and TNF-α in the peritoneal lavage fluid was not affected.101 This result contradicts the conclusion of Mao’s team in their review that ginger extract has anti-inflammatory property.100 The reason for the difference in results might be the short exposure time and the low dose of ginger extract used in their experiments.101
Extract of Urtica Dioica L
Urtica dioica L. is a perennial herb in the family Urticaceae.102 Ilhan et al found that flavonoids from Urtica dioica L. decreased the expression levels of TNF-α, VEGF, and IL-6 in the peritoneal fluid of Wistar albino rats endometriosis model, reduced the adhesion score of ectopic endometrial lesions and decreased the volume of ectopic lesions. They concluded that the anti-endometriosis effect of the methanolic extract of Urtica dioica L. was mainly attributed to its flavonoids.103
Purified Bioactive Compounds
Apigenin
Apigenin (Figure 2) is isolated from vegetables and fruits, including bergamot, parsley and orange.104 Apigenin has a wide range of biological properties, such as antitumor,105 anti-inflammatory,106 and pro-apoptotic107 properties. Park et al demonstrated that apigenin could inhibit the proliferation and induce apoptosis of human endometriosis cells. Treatment of apigenin on vaginal mucosa-derived epithelial endometriotic cells (VK2/E6E7) and endocervix-derived endometriotic cells (End1/E6E7) cells resulted in the loss of mitochondrial membrane potential, increased calcium in the cytoplasm, elevated the expression of the pro-apoptotic protein Bax, and the release of cytochrome c (Cyt c).108 Released Cyt c and apoptotic protease activating factor-1 (Apaf-1) form a multimeric complex, then the caspase recruitment domain (CARD) at the amino terminus of Apaf-1 recruits the cytoplasmic caspase-9 precursor to form a new complex. This new complex activates downstream caspase-3 and caspase-7 to cause apoptosis (Figure 3).97 In protein synthesis, the endoplasmic reticulum is also involved in the processing and transmission of apoptotic signals. Apigenin treatment increased the production of ROS and the expression of endoplasmic reticulum stress proteins in VK2/E6E7 and End1/E6E7 cells.108 The protein kinase RNA-like ER kinase (PERK), eukaryotic translation initiation factor 2α (eIF2α), glucose regulatory protein (GRP78), and growth arrest and DNA damage 153 (GADD153/CHOP) are all endoplasmic reticulum stress-related proteins. In VK2/E6E7 and End1/E6E6 cells, the phosphorylation levels of PERK and eIF2α were upregulated, and even GADD153 and total GRP78 proteins were increased after the treatment of apigenin.108 When endoplasmic reticulum stress (ERS) is severe and persistent, it can trigger apoptosis by activating the endoplasmic reticulum (ER) pro-apoptotic cysteine protease caspase-12 and inhibiting the expression of Bcl-2.98 Park et al also found that the pro-apoptotic effect of apigenin on endometriosis cells and the inhibitory effect on cell proliferation were achieved through modulating MAPK and PI3K/Akt signaling pathways.108
 |
Figure 2 Structures of bioactive compounds (part 1) against endometriosis. |
 |
Figure 3 In the mitochondrial pathway, the death signal activates members of the Bcl-2 family, such as Bax. Bax can be inserted into mitochondrial membrane, causing changes in mitochondrial membrane permeability, and releasing Cyt c. Released Cyt c and Apaf-1 form a multimeric complex, then the CARD at the amino terminus of Apaf-1 recruits the cytoplasmic caspase-9 precursor to form a new complex. This new complex activates downstream caspase-3 and caspase-7 and finally cause apoptosis. |
Carnosic Acid and Rosmarinic Acid
The phenolic diterpene carnosic acid (CA, Figure 2) and the polyphenol rosmarinic acid (RA, Figure 2) are abundantly identified from rosemary (Rosmarinus Officinalis) leaves.109,110 Previous studies have demonstrated that this plant shows antioxidant,110 anti-inflammatory,111 anti-cancer properties.112 Ferella et al proposed that CA and RA could significantly inhibit the proliferation of Telomerase-immortalized human endometrial stromal cells (T-HESC) and primary endometrial stromal cells. CA induced cell cycle arrest at the G2/M phase, but CA had no effect on the change of intracellular reactive oxygen species (ROS) level. RA reduced intracellular ROS accumulation and exerted antioxidant properties. CA and RA significantly reduced the area of ectopic endometrial lesions in a mouse model of surgically induced endometriosis.113
Chrysin
Chrysin (Figure 2) is a flavonoid obtained from natural materials such as honey, propolis, flat mushrooms, and passion flowers.114,115 Chrysin has a variety of biological functions, including antiproliferative,116 anti-inflammatory,117 and antioxidant118 effects. Ryu et al showed that chrysin might exert anti-proliferative and pro-apoptotic effects on human endometriosis cells by regulating endoplasmic reticulum stress and the PI3K/Akt signaling pathways. They also found that chrysin varied the cell cycle ratio by increasing the proportion of End1/E6E7 cells and VK2/E6E7 cells in the sub-G1 phase and decreasing the proportion of cells in the G0/G1 phase. Cytoplasmic calcium levels, ROS, and a key protein of endoplasmic reticulum stress GRP78, were significantly increased in chrysin-treated endometriosis cells.119 The endoplasmic reticulum is the main site of intracellular protein synthesis and a major reservoir of Ca2+. Oxidative stress and disturbance of endoplasmic reticulum calcium metabolism may disrupt endoplasmic reticulum homeostasis, leading to endoplasmic reticulum stress, which then induces the unfolded protein response (UPR) to adapt to these changes. When ERS is severe and persistent, UPR eventually activates the expression of endoplasmic reticulum pro-apoptotic caspase-12. Activated caspase-12 can further shear caspase-3, eventually inducing apoptosis (Figure 4).98
 |
Figure 4 Oxidative stress and disturbance of endoplasmic reticulum calcium metabolism may disrupt endoplasmic reticulum homeostasis, leading to endoplasmic reticulum stress, which then induces the UPR to adapt to these changes. If UPR continue to accumulate in the ER, GRP78 can be released from IRE1, ATF6 and PERK. Furthermore, GRP78 binds to unfolded proteins to activate three signaling proteins, namely PERK, ATF6, and IRE1, which are responsible for ERs and signaling transmission. When ER stress is persistent and severe, it promotes cell apoptosis by upregulating the expression of GADD153/CHOP, by activating caspase-12, and by inhibiting Bcl-2 expression. |
Curcumol
Curcumol (Figure 2) is a dominating constituent isolated from the root of the herbal plant turmeric.120 Curcumol possesses significant pharmacological activities such as anti-cancer,121 anti-inflammatory,122 and anti-fibrotic.123 Wang et al investigated that curcumol inhibited the progression of endometriosis by inhibiting the JAK2/STAT3 pathway in vivo and in vitro.124 JAK2/STAT3 is an important signaling axis in the cell proliferation, apoptosis, and inflammation. Cytokines such as IL-6 and interferon can activate JAK2/STAT3 signaling pathway and phosphorylate STAT3. The phosphorylated STAT-STAT dimer enters the nucleus and then participates in the transcriptional regulation of related target genes, such as Bcl-2 and caspase-3, ultimately leading to the sustained proliferation of ectopic endometrium and enhanced resistance to apoptosis.124–126 Curcumol reduced the production of inflammatory cytokines such as TNF-α, IL-6, and IL-1β of ectopic endometrial stromal cells (EESCs). This team also demonstrated that curcumol suppressed the proliferation of EESCs, accelerated cell apoptosis, and attenuated cell migration activity by MTT assay, wound healing assay, and flow cytometry. Curcumol also promoted the upregulation of apoptosis-related proteins Bax and caspase-3 and the downregulation of the expression of anti-apoptotic protein Bcl-2 in ectopic lesions in rats with autologous transplants of endometriosis. Curcumol also reduced the expression of TNF-α, IL-6, and IL-1β in the peritoneal fluid of rats with endometriosis.124
Daidzein-Rich Isoflavone Aglycones
Soy glycosides are isoflavone glycosides that are extracted from soybeans. However, they have high hydrophilicity and high molecular weight, which cannot be absorbed directly by the intestine. To be absorbed, they need to be degraded by the glucosidase in the intestinal flora. Soy glycosides are transformed into Daidzein-rich isoflavone aglycones (DRIAs) after removal of the sugar chain, which can be absorbed rapidly and efficiently by the intestine.127 NF-κB dimers (p50 and p65) bind to IκB to form a complex. This complex is retained in the cytosol at rest. IL-1β stimulation causes activation of the IκB kinases (IKKs) complex, then IKKs activation leads to IκB ubiquitination and subsequent proteasomal degradation. This results in the release of the nuclear factor-κB dimer (p50 and RelA/p65), which is translocated to the nucleus for regulation of transcription.128,129 Activation of the NF-κB pathway contributes to the elevation of inflammatory cytokines, such as IL-6 and IL-8130 (Figure 5). The results of Takaoka et al suggested that DRIAs might exert anti-inflammatory and proliferation-inhibiting effects by inhibiting phosphorylation of IκB and nuclear translocation of p65. In the endometriosis mouse model, DRIAs-containing diets significantly reduced the percentage of Ki-67-positive cells in endometriotic lesions.131 Bulun’s group proposed that estrogen stimulates the expression of cyclooxygenase-2 (COX-2) and the production of prostaglandin E2 (PGE2) through estrogen receptors (ERs). Then PGE2 can stimulate the expression of the aromatase gene in endometrial stromal cells. Ultimately, aromatase locally converts cholesterol to estradiol, thus forming a vicious cycle.2 Takaoka et al stated that the expression of estrogen receptor β (ERβ) in ovarian endometrial stromal cells (OESCs) was suppressed at the gene and protein levels after DRIAs treatment, which in turn inhibited the expression of COX-2 and the production of PGE2. The decreased expression of PGE2 results in the decrease of aromatase activity.131 In conclusion, because of the inhibition of ERβ-mediated proliferative and anti-apoptotic effects in endometriosis stromal cells, DRIAs are expected to be a new therapeutic candidate drug for endometriosis.
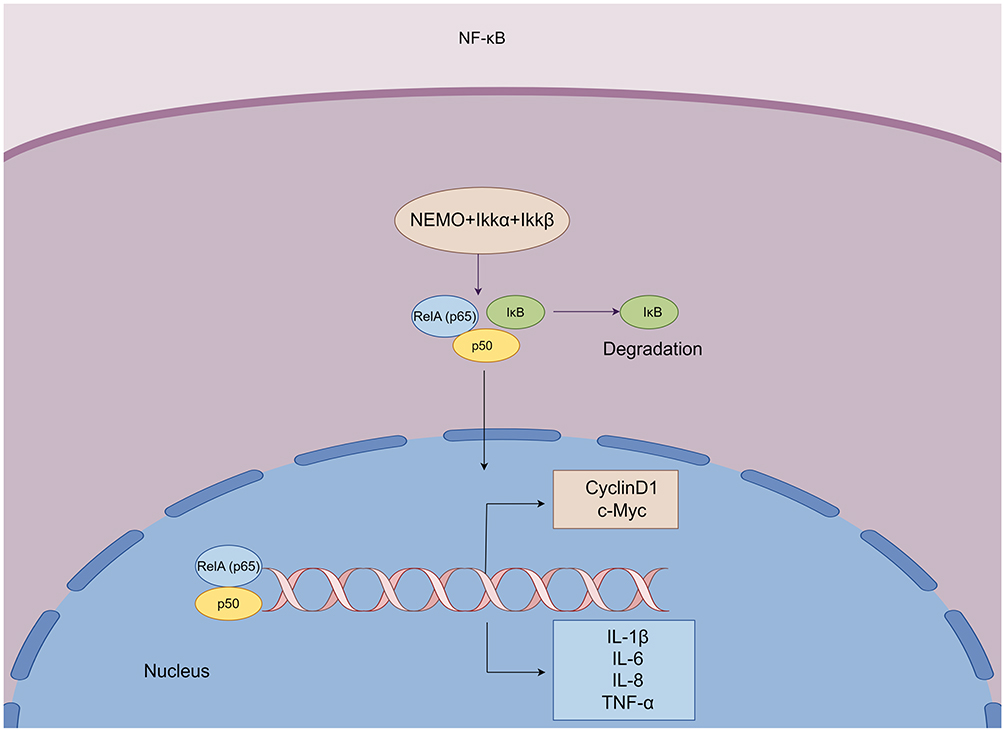 |
Figure 5 NF-κB dimers (p50 and p65) bind to IκB to form a complex. The activation of IκB kinases (IKKs) complex leads to IκB ubiquitination and subsequent proteasomal degradation. This results in the release of the nuclear factor-κB dimer (p50 and RelA/p65), which is translocated to the nucleus for regulation of transcription. Activation of the NF-κB pathway induces transcription of target genes, such as CyclinD1, c-Myc, IL-1β, IL-6, TNF-α, etc. |
Dehydrocostus Lactone
Dehydrocostus lactone (DHL, Figure 2) is a natural sesquiterpene lactone, mainly extracted and purified from the root of the medicinal plant Aucklandia lappa.132 Dehydrocostus lactone exerts different pharmacological benefits, including pro-apoptotic,133 anti-proliferative,134 anti-fibrotic,135 and anti-inflammatory.135 It is well known that initiation of the caspase cascade is essential for cell apoptosis. Woo et al showed that DHL induced apoptosis of the human endometriotic 12Z cells by activating the caspase cascade.136 PGE2 is a pivotal factor causing inflammation and pain allergy in patients with endometriosis.137 COX-2 is the key enzyme for the biosynthesis of PGE2 by arachidonic acid.138 DHL exposure significantly reduced the expression of PGE2 and COX-2 in 12Z cells.136 Neurotrophins, known as endometriosis-related pain factors, are a family of nerve growth factors such as BDNF, NGF, NT-3, and NT-4/5, which are overexpressed in patients with endometriosis.81 Treatment with DHL significantly reduced the expression of NGF, NT-3, BDNF, and NT-4/5 in 12Z cells.136 Previous studies have demonstrated an increase in M2 macrophages in lesions of patients with endometriosis.139 Activated M2 macrophages promote tissue remodeling and angiogenic activity.140 In the tumor microenvironment, M2 macrophages have been shown to induce angiogenesis, metastasis, and immune escape through the production of various factors such as VEGF, MMP-2, MMP-9 and IL-10.141,142 In contrast, monocyte chemotactic protein-1 (MCP-1), as a chemokine, is responsible for the recruitment of macrophages to endometriotic lesions.143 Huang et al found that alternate activation of macrophages (M2 polarization) was significantly inhibited after the treatment with dehydroxylactone. DHL stimulated the decreased expression of CD206 and TREM-2, as the markers of the endometriosis-associated macrophage M2 phenotype, as well as the decreased expression of MCP-1 and M2-related molecules (IL-10, VEGF, MMP-2 and MMP-9).136 These effects of DHL might be achieved through inhibition of the Akt and NF-κB pathways.136
Delphinidin
Delphinidin (Figure 2) is originally isolated from brightly colored pigments of vegetables and fruits.144 Delphinidin is an anthocyanin with anti-inflammatory, anti-angiogenic, antioxidant, and anti-cancer properties.144–146 Delphinidin exhibited anti-proliferative and pro-apoptotic effects in a dose-dependent manner on human endometriosis-derived cell lines by regulating the cell cycle according to the results of Park S. The immunofluorescence intensity of proliferating cell nuclear antigen (PCNA) was decreased after the treatment of delphinidin in both cell lines.147 In addition, the decrease in mitochondrial membrane potential, the gradual increase in intracellular calcium ion levels, and cellular mitochondrial dysfunction occurred with the increasing doses of delphinidin treatment, ultimately led to endometrial cell apoptosis.147 When the intracytoplasmic Ca2+ concentration increases, it can activate calcium-dependent proteases and the opening of transition pores, then affect mitochondrial permeability and result in the loss of mitochondrial membrane potential.97,148–150 When mitochondrial membrane potential begins to decline, mitochondria release cytochrome c into the cytoplasm. Cyt c and Apaf-1 are released to form a multimeric complex. CARD at the amino terminus of Apaf-1 recruits the cytoplasmic caspase-9 precursor to form a new complex. This new complex initiates the caspase cascade reaction and activates downstream caspase-3 and caspase-7, thereby inducing apoptosis.97 Park et al showed that the antiproliferative effect of delphinidin was achieved through the regulation of the PI3K/Akt and MAPK signaling pathways.147
Dimethylchrysin
Dimethylchrysin (Figure 2) is the totally methylated form of Chrysin. Kaempferia parviflora (K. parviflora, KP) are good sources of dimethylchrysin, which is extensively used in Thai and Lao folk medicine.151,152 Park et al observed that dimethylchrysin had anti-proliferative and apoptotic effects on VK2/E6E7 and End1/E6E7 cells.152 Treatment of dimethylchrysin in two endometriosis cell lines led to reduced cell proliferation by inactivating the PI3K/Akt pathway. Dimethylchrysin also accelerated cell apoptosis through DNA breakage and cell cycle arrest. In addition, dimethylchrysin could mediate mitochondrial and endoplasmic reticulum apoptotic pathways by inducing ROS production, intracellular calcium ion dysregulation, and mitochondrial membrane potential loss in human endometriosis-derived cell lines.152 Oxidative stress and disturbance of endoplasmic reticulum calcium metabolism may disrupt ER homeostasis, then lead to the production of unfolded proteins in ER. If unfolded proteins continue to accumulate in the ER, GRP78 (also known as BIP) can be released from inositol-requiring kinase/endonuclease 1 (IRE1), activating transcription factor 6 (ATF6) and protein kinase activated by PERK. Furthermore, GRP78 binds to unfolded proteins to activate three signaling proteins, namely PERK, ATF6, and IRE1, which are responsible for ERs and signaling transmission. However, when ER stress is persistent and severe, UPR is insufficient to restore ER function, it promotes cell apoptosis by upregulating the expression of GADD153/CHOP, by activating caspase-12, and by inhibiting Bcl-2 expression.98 Their study also demonstrated that the expression levels of GRP78 and IRE1α were increased in a dose-dependent manner after the treatment of dimethylchrysin.152 Dimethylchrysin holds promise to inhibit the progression of endometriosis.
Ellagic Acid
Ellagic acid (EA, Figure 2) belongs to the group of polyphenol. EA can be found in various vegetables, fruits, and nutmeat.153 It possesses a wide range of biological activities and has a variety of mechanisms of action. EA exhibits pro-apoptotic, anti-angiogenic, anti-proliferative, anti-inflammatory, antioxidant, anti-migration, and anti-invasion effects.154,155 Cell migration, attachment, proliferation, and invasion are closely related to the pathogenesis of endometriosis. And simultaneously they are involved in the onset, development, and growth of ectopic endometrial lesions.53 Mc Cormack et al demonstrated that EA affected the cell cycle progression of T-HESCs by arresting the cell cycle in the G2/M phase. Besides, EA inhibited the migration and adhesion of endometrial epithelial cells (ECC-1) and T-HESCs. Taken together, EA can suppress the progression of endometriosis.156
Epigallocatechin-3-Gallate
Epigallocatechin gallate (EGCG, Figure 6) is found with a high concentration in green tea, and exerting a wide range of pharmacological benefits. Kamal et al summarized in a review that EGCG had anti-angiogenic, anti-fibrotic, antiproliferative, antioxidant, and pro-apoptotic effects.157 E-cadherin mediates abnormal cell adhesion function. Reduced expression of E-cadherin can loosen cell adhesion, facilitate cell movement, and accelerate cell metastasis and invasion.158 Guan et al showed that EGCG was effective in inhibiting the progression of endometriosis. EGCG reduced the size of the ectopic endometrium in an endometriosis nude mouse model. Guan proposed the mechanism of the anti-endometriosis effect of EGCG was the demethylation of the E-cadherin promoter region, resulting in the upregulation of E-cadherin expression.159
 |
Figure 6 Structures of bioactive compounds (part 2) against endometriosis. |
Flavokawain a
Kava-kava, known as the piper methysticum, is a medicinal plant belonging to the pepper family. Flavokawain A (FK A, Figure 6) is a chalcone isolated from kava-kava.160 Flavokawains mainly have anti-cancer161 and anti-inflammatory162 biological activities. Wei et al established a rat model of endometriosis by autologous transplantation. After the treatment of FK A, the inflammatory cytokines in peritoneal fluid and plasma, such as IL-1β, IL-8, nitric oxide (NO), COX-2, and PGE2 were decreased. In addition, FK A could promote the expression of apoptosis-related proteins Bax and cleaved caspase-3, and also reduce the expression of anti-apoptotic protein Bcl-2 in ectopic endometrial tissues. The expression of VEGF in peritoneal fluid and plasma of rats treated with FK A was decreased too. The authors proposed that FK A can exert anti-inflammatory, pro-apoptotic, and anti-angiogenic effects by inhibiting the overactivation of PI3K/Akt and NF-κB signaling pathways, ultimately inhibiting the progression of endometriosis.163
Gamma Oryzanol
Gamma oryzanol (GO, Figure 6) is mainly found in rice bran and rice bran oil.164,165 Gamma oryzanol has abundant biological activities, including antioxidant165,166 and anti-inflammatory.167 Eisalou et al demonstrated that GO decreased steroidogenic factor 1 (SF1) expression and upregulated BECN1 expression in autologous transplanted rat endometriotic lesions.168 SF1, also known as NR5A1, is an orphan nuclear receptor, and there is no clear natural ligand for SF1. SF1 plays a key role in regulating the steroid synthesis pathway in the ovary. SF1 can bind to the promoters of several steroidogenic genes, such as steroid synthesis acute regulatory protein (STAR) and aromatase, which can result in the production of high amounts of estradiol.2 It is well known that increased estradiol synthesis promotes the proliferation of endometriotic lesions, inhibits cell apoptosis, and promotes the occurrence and development of endometriosis. Beclin1 is a specific marker of autophagy.169,170 In conclusion, GO may inhibit the progression of endometriosis by decreasing estrogen production and increasing cell autophagy.
Ginsenoside PPD
Ginseng is a traditional herbal medicine, while ginsenosides PPD (Figure 6) is the dominating bioactive component of the Panax genus.171 Ginsenoside PPD has significant biological activity, such as anti-inflammatory172 and anti-cancer.173 There are strong evidences that estrogen dependence, progesterone resistance, and immune system defects, play important roles in the development of endometriosis.2,53 Cellular autophagy is another type of programmed cell death. Under the influence of external environmental factors, cells use lysosomes to degrade their own damaged, degenerated, or senescent macromolecules as well as organelles (such as mitochondria, and endoplasmic reticulum). Autophagy is a self-protective mechanism and plays a vital role in regulating cell survival and death.174 Mei et al found that estrogen could inhibit autophagy in endometrial stromal cells (ESCs) by up-regulating the expression of chemokine CXCL12 and CXCR4 (the receptor of CXCL12).175 Zhang et al believed that increased expression of estrogen and estrogen receptor α (ERα) inhibited autophagy and apoptosis in EESCs, however, PPD could reverse this inhibition. PPD treatment significantly downregulated the expression of ERα and upregulated the expression of progesterone receptor (PR). Subsequently, PPD induced a high level of autophagy in EESCs, upregulated the expression of pro-apoptotic molecules, Bax/Bcl-2 antagonist/killer (Bak), and downregulated the expression of anti-apoptotic molecules (Bcl-2/Bcl-xL). What’s more, progesterone can prevent the inhibitory effect of estrogen on autophagy in EESCs. PPD could decrease the expression of ERα and increase the expression of PRα in ectopic lesions in a mice model of endometriosis, then lead to autophagy. In addition, PPD could upregulate granzyme B and perforin, activating natural killer (NK) cells. The activated NK cells exerted cytotoxic effects on EESCs, enhanced the immune surveillance of ectopic lesions, and inhibited the progression of endometriotic lesions.176
Hesperidin and Nerolidol
Hesperidin (Figure 6) is a bioflavonoid that is abundant in citrus fruits, including lemons, oranges and grapefruits.177,178 Nerolidol (Figure 6) exists in the essential oils of many plants with a floral aroma.179,180 Hesperidin and nerolidol have a variety of physiological actions, including anti-inflammatory effects, antioxidant, and other effects.177,179–181 Like vitamin C, hesperidin exerts its anti-inflammatory effect by inhibiting the NF-κB signaling pathway and reducing inflammatory cytokines, such as TNF-α, IL-6, and IL-1β.182 Nerolidol also exerts anti-inflammatory properties by reducing the levels of TNF-α and IL-1β, as well as by reducing the number of polymorphonuclear cells.183 Treatment with hesperidin or nerolidol resulted in obvious regression of surgically induced endometriosis foci in rats. Hesperidin or nerolidol treatment resulted in an obvious decrease in malondialdehyde (oxidant) levels, a significant increase in glutathione levels, and superoxide dismutase (antioxidant enzyme) activity. Hesperidin or nerolidol also improved the pathological status of inflammatory cell infiltration, tissue necrosis and hemorrhage, and vascular congestion in endometriotic lesions in a rat model of endometriosis.184 Since inflammation and oxidative stress have been considered as underlying factors in the pathophysiological mechanisms of endometriosis, hesperidin or nerolidol may provide a new strategy for the prevention and treatment of inflammatory diseases such as endometriosis.
Isoliquiritigenin
Isoliquiritigenin (ISL, Figure 7) is a natural chalcone isolated from the roots of Glycyrrhiza spp.185 Previous studies showed the antioxidant,186 anti-inflammatory,187 anti-fibrosis,188 and anticancer189 effects of ISL. Epithelial mesenchymal transition (EMT) refers to the morphological transformation of epithelial cells into a fibroblastic or mesenchymal cell phenotype, with loss of cell polarity, cytoskeletal rearrangement, and increased migratory motility. The expression level of E-cadherin is negatively correlated with the invasive and metastatic ability of cells.158,190 Since E-cadherin has a stronger adhesive ability than N-cadherin, a decrease in E-cadherin expression and an increase in N-cadherin expression on the cell membrane surface increase cell motility and the ability to invade and metastasize.191 Snail and Slug can competitively bind to the E-box sequence of the proximal promoter of E-cadherin gene with SMAD-associated protein 1 (SIP1), thereby inhibiting the expression of E-cadherin and inducing EMT in cells.158 Hsu et al found that the expression of E-cadherin was upregulated, however N-cadherin, Slug, and Snail could be reduced after isoliquiritigenin treatment, then inhibited EMT of End1/E6E7 and ectopic endothelial cells, thereby inhibiting migration of cells in vitro and in vivo. They also demonstrated that Bcl-2 and estrogen receptor β were decreased, while pro-apoptotic protein Bax and cleaved caspase-3 were upregulated in vivo. The expressions of IL-1β, IL-6, and VEGF in serum and ectopic endometrial tissues were decreased after ISL treatment.192
 |
Figure 7 Structures of bioactive compounds (part 3) against endometriosis. |
Kaempferol
Kaempferol (KP, Figure 7) is a dietary flavonoid mainly found in tea, some medicinal plants, fruits and vegetables.193 Previous studies showed that kaempferol has anti-inflammatory,194 anticancer,195 and anti-angiogenic196 biological properties. Presented by Zhao et al, the protein expression of PTEN was upregulated, while the expression level of MMP-9 was decreased in Ishikawa cells treated with kaempferol.197 In the PI3K/Akt signaling pathway, PTEN can convert phosphatidylinositol 3,4,5-triphosphate (PIP3) to phosphatidylinositol 4.5-bisphosphate (PIP2), thereby preventing the phosphorylation of Akt, and then inhibiting the activation of PI3K/Akt/mTOR signaling pathway (Figure 8).198 They also demonstrated that kaempferol exposure to Ishikawa cells inhibited cell migration and invasion by using scratch assay (wound healing) and perforation assay.197
 |
Figure 8 PI3K produces the second messenger PIP3 in cells, which then activates Akt. The activated Akt can inhibit the TSC1/2 complex, thus activating mTORC1 and inhibiting autophagy. In the PI3K/Akt signaling pathway, PTEN converts PIP3 to PIP2, thereby inhibiting Akt phosphorylation. Furthermore, the activation of PI3K/Akt/mTOR signaling pathway is inhibited. Caspase-9, Myc, CCND1, Bcl-2 and NO are downstream target genes of the PI3K/Akt/mTOR signaling pathway. |
Luteolin
Luteolin (Figure 7) is a naturally occurring flavonoid that is widely distributed in flowers, vegetables, herbs, and spices.199,200 Cyclin D1 (CCND1), cyclin E1 (CCNE1), cyclin-dependent kinase 2 (CDK2), and cyclin-dependent kinase 4 (CDK4) are all cell cycle regulation-related proteins/genes, while CCND1 and CCNE1 are key molecules for G1 phase progression.201,202 Cyclin D1 can bind to either CDK4 or CDK6. Cyclin D1 overexpression allows cells to rapidly pass the G1/S checkpoint, resulting in a shortened S phase. In contrast, blocking or knocking down Cyclin D1 blocks the cell cycle and induces apoptosis. Cyclin E1 is the rate-limiting factor for the transition between G1 and S phases. Cyclin binds to the corresponding CDK to form a complex that promotes Rb protein phosphorylation, then the phosphorylated Rb releases the nuclear transcription factor E2F, which is free to enter the nucleus and promotes the transcriptional expression of downstream genes, allowing cells to move from G1 phase to S phase.203–205 The anti-proliferative and pro-apoptotic effects of luteolin on endometriosis have been demonstrated by Park et al in vivo and in vitro. Treatment with luteolin, VK2/E6E7 and End1/E6E7 cells were arrested in the sub-G0/G1 phase. Luteolin decreased the mRNA expression of CCNE1, CDK2, and CDK4, which in turn inhibited cell proliferation by preventing cell cycle progression.206 Activation of the pro-apoptotic proteins recombinant Bak and Bax can insert into the mitochondrial membrane, causing changes in mitochondrial membrane permeability, loss of transmembrane potential, then releasing of Cyt c. Cyt c can form a multimeric complex with Apaf-1, which can recruit the cytoplasmic caspase-9 precursor through the CARD at the amino terminus of Apaf-1 to form a complex. This complex initiates the caspase cascade reaction, which activates downstream caspase-3 and caspase-7 to cause cell apoptosis.97,207 The endoplasmic reticulum is the main site of intracellular protein synthesis and a major reservoir of Ca2+. Within the endoplasmic reticulum, disruption of calcium homeostasis or excessive accumulation of endoplasmic reticulum proteins induces caspase-12 expression on the endoplasmic reticulum membrane. Activated caspase-12 further shears caspase-3 and triggers apoptosis.98 There is a correlation between the endoplasmic reticulum and the mitochondrial apoptotic pathway. When the intracytoplasmic Ca2+ concentration is elevated, it can activate calcium-dependent proteases, and affect mitochondrial permeability as well as membrane potential, thereby promoting apoptosis.97,148 Park et al illustrated that luteolin could also lead to cell apoptosis by causing intracellular calcium and ROS accumulation, by activating the pro-apoptotic proteins Bax and Bak.206 They suggested that these effects of luteolin were mediated through the regulation of PI3K/AKT and MAPK signaling pathways.206
Myricetin
Myricetin (MYR, Figure 7) belongs to flavonol, and commonly presents in fruits, vegetables, wines, berries and teas. Myricetin has a variety of pharmacological activities, including anti-inflammatory, anticancer, anti-angiogenesis, and anti-proliferation.208,209 Park et al showed that myricetin had anti-proliferative and pro-apoptotic effects in endometriosis.210 Myricetin inhibited cell proliferation by reducing CCNE1 expression according to the results of Park et al. They also suggested that myricetin could inhibit the expression of Bcl-2 and induce the expression of Bax by regulating the MAPK signaling pathway. Moreover, myricetin induced apoptosis of endometriosis patient-derived cells (VK2/E6E7 and End1/E6E7) by mediating ROS, calcium ion accumulation, and causing changes in mitochondrial membrane potential (MMP).210
Naringenin and Naringin
Naringenin (NGN, Figure 7) is a flavonoid compound, it is the aglycone of naringin. Naringenin can be found in citrus fruits (grapefruit, oranges) and tomatoes. Naringenin has a variety of biological activities.211 Nrf2 regulates the transcription of endogenous antioxidant enzymes, such as glutamate cysteine ligase (GCL) and glutathione (GSH). Furthermore, Nrf2 can protect against oxidative damage caused by oxidative stress and inflammation.212 Under basal conditions, Keap1 forms a complex with Nrf2, leading to ubiquitination and subsequent degradation of Nrf2. Under conditions of oxidative stress, Keap1-CUL3-E3 ubiquitin ligase undergoes conformational changes that disrupt the binding of Nrf2 to Keap1. Free Nrf2 translocates to the nucleus and transcribes antioxidant enzyme (NQO1, HO-1) genes.65 Nrf2 and its downstream molecules (NQO1, HO-1) were significantly elevated in endometriotic lesions, and these antioxidants mitigated cell death by scavenging ROS (O2− and ·OH).213 Kapoor et al illustrated that naringin treatment on endometrial cells significantly inhibited the expression of the Nrf2 protein and its downstream HO-1 and NQO1. In other words, a decrease in antioxidant enzymes means an increase in ROS. ROS can disrupt protein, lipid, and DNA structures, ultimately leading to cell apoptosis.213,214 The accumulation of ROS in the cell leads to unfolded protein response and induces endoplasmic reticulum stress. Kapoor et al demonstrated that naringenin significantly increased the loss of mitochondrial membrane potential and shifted the MMP toward depolarization. They also showed that naringenin decreased the expression levels of TNF-α, NO, MMP-2, and MMP-9. Naringin could accelerate mitochondria-mediated cells apoptosis by generating mitochondrial membrane potential depolarization, by increasing the expression level of Bax, by releasing Cyt c and activating caspase-3 in VK2/E6E7 and End1/E6E7 cells.215 Taking together, naringin promoted apoptosis by inducing mitochondrial membrane depolarization, endoplasmic reticulum stress, ROS production, and regulating MAPK and PI3K/Akt pathways. In addition, naringin also exhibited anti-inflammatory and anti-invasive effects. These results suggest that naringenin can delay the progression and development of endometriosis.
Nobiletin
Nobiletin (Figure 7), a polymethoxyflavonoid isolated from citrus peel,216 has a wide range of biological effects and plays an interesting role in anticancer,217 anti-inflammatory,218 and anti-oxidation.219 Activation of the NF-κB pathway induces transcription of target genes, such as Cyclin D1, c-Myc, IL-1β, IL-6, TNF-α, etc.128,129,220,221 Wei et al found that nobiletin significantly reduced the gene expression levels of IL-6, IL-1β, TNF-α, matrix metalloproteinase-1 (MMP-1), and matrix metalloproteinase-3 (MMP-3). Compared with untreated endometriosis mice, the expression levels of PCNA and VEGF were significantly reduced, however, E-cadherin was significantly increased in the nobiletin-treated group. The effects of nobiletin on endometriosis might be attributed to the inhibition of the activity of IKKs.222
Oleuropein
Oleuropein (Figure 7) is a polyphenolic compound extracted from olive.223 ERβ can increase the proliferation of ectopic endometrial lesions and prevent apoptosis, thus promoting the progression of endometriosis.2 Park et al found that oleuropein could selectively inhibit the activity of ERβ, but had no effect on the activity of ERα. In addition, oleuropein reduced the expression of proliferation-related indicator Ki-67, promoted the expression of cleaved caspase-3 and reduced the expression of cellular inflammatory factors, thus the endometriosis was alleviated.224
Quercetin
Quercetin (Figure 9) is a polyphenolic flavonoid compound chemically.225 Quercetin is abundantly identified from fruits and vegetables, taking cilantro, grapes, apples, onions, and cherries as examples.226,227 The upregulation of CCND1 is tightly associated with the pathogenesis of endometriosis.228 CCND1 is the G1/S checkpoint of the cell cycle, and reduced expression of CCND1 is associated with G1 phase arrest.229 Park et al observed that quercetin induced cell cycle arrest in the G0/G1 phase by decreasing the expression of CCND1 protein and mRNA level, which in turn inhibited the proliferation of VK2/E6E7 and End1/E6E7 cells. Quercetin also induced apoptosis through the accumulation of ROS and the loss of mitochondrial membrane potential. Park et al suggested that the anti-proliferative and pro-apoptotic effects of quercetin might be achieved through regulation of MAPK and PI3K/Akt signaling pathway transduction.6 Jamali et al showed that the combined application of quercetin and metformin exerted anti-inflammatory effects by decreasing the inflammatory cytokine TNF-α.230 Increased expression of antioxidant enzymes can attenuate the oxidative stress response.213 Quercetin could promote the expression levels of the Nrf2 gene and enhance the activity of its downstream target gene/antioxidant enzyme (NQO1).230 Kim et al demonstrated that mTOR played a negative regulatory role in the autophagy pathway.231 In a surgically induced endometriosis model, the expression of the mTOR gene was reduced in ectopic endometriotic lesion tissues, and gene expression levels of autophagy markers (Beclin 1 and ATG5) were significantly increased.230
 |
Figure 9 Structures of bioactive compounds (part 4) against endometriosis. |
Resveratrol
Resveratrol (Figure 9) is a naturally occurring polyphenolic compound which is extensively present in grapes, red wine, and peanuts.232 Resveratrol possesses pro-apoptotic,233 anti-angiogenic,234 anti-inflammatory,235 antioxidant236 and anti-tumor237 biological properties. Cell migration is associated with the aggressiveness of endometriosis,238 which depends on the ratio of matrix metalloproteinase (MMP)/TIMP1 (tissue inhibitor of metalloproteinase-1). Khodarahmian et al summarized that resveratrol could reduce the expression of VEGF, TNF-α protein, and mRNA levels in the eutopic endometrium of patients with endometriosis.239 Madanes et al illustrated that resveratrol could promote apoptosis by increasing the expression level of cleaved caspase-3, and could exert an anti-angiogenic effect by reducing the expression of VEGF and angiotensin converting enzyme-1 in 12Z and ST-T1b cells. Madanes et al also demonstrated that resveratrol treatment could exert an anti-invasive effect by decreasing the MMP-2/TIMP1 ratio in primary endometriotic stromal cells, 12Z and ST-T1b cells. Resveratrol could suppress cell migration.240 In conclusion, resveratrol reduces angiogenesis, migration, invasion, and inflammatory response in vitro and in vivo.
Scutellarin
Scutellarin (Figure 9) is classified as a flavonoid obtained from the medicinal plant Erigeron breviscapus (Vant). Hand-Mazz.241 Scutellarin has been shown to have a variety of pharmacological effects, including anticancer, antioxidant, anti-proliferative, anti-invasive, anti-angiogenic, and anti-inflammatory effects.242–247 Like the eutopic endometrium, the ectopic endometrium also undergoes periodic bleeding and shedding under the regulation of estrogen, resulting in tissue damage. Platelets are involved in the repair of damaged tissue. Once tissue injury or trauma occurs, platelets are the first to reach the trauma site, initiating hemostasis, inflammation, and cell proliferation. In addition, they participate in the repair process of tissue remodeling. Activated platelets secrete a large number of bioactive molecules, including various cytokines and growth factors, such as VEGF, epidermal growth factor (EGF), TGF-β, and platelet-derived growth factor (PDGF).248 TGF-β induces the TGF-β/Smad signaling pathway, which is a critical pathway for regulating EMT. Activated platelets also drive transdifferentiation of fibroblasts to myofibroblasts (FMT) and smooth muscle metaplasia (SMM).249 Ding et al found that scutellarin could effectively restrain the development and progression of endometriosis by inhibiting the aggregation and activation of platelets, by inhibiting tissue proliferation, angiogenesis, and fibrosis.250
Silibinin
Silibinin (Figure 9) is isolated from the Silybum marianum (milk thistle) plant.251,252 Silibinin has numerous biological activities, principally such as antioxidant,253 anti-migration,254 anticancer255 and anti-inflammatory256 effects. Mirzaei’s group divided 70 patients with endometriosis into an experimental group (silymarin group) and a control group (placebo group). Their results showed that silymarin decreased IL‐6 expression of serum and reduced the size of endometriotic lesions.257 Ham et al demonstrated that silibinin exhibited anti-inflammatory, anti-proliferative, and pro-apoptotic effects in vitro and in vivo. The mRNA levels of TNF-α, IL-1β, and IL-6 in ectopic endometriotic lesions were decreased after intraperitoneal injection of silibinin in a mouse model of endometriosis. Increased cytoplasmic calcium ion concentration, disrupted mitochondrial membrane potential, and increased oxidative stress after the treatment of silibinin in VK2/E6E7 and End1/E6E7 cells were observed.258 Oxidative stress, and disturbance of endoplasmic reticulum calcium metabolism may lead to elevated expression of endoplasmic reticulum stress-related proteins such as IRE1, ATF6, and PERK, which are responsible for ERS and signal transmission. When ERS is severe and persistent, the expression of GADD153/CHOP and caspase-12 can be upregulated, ultimately inducing apoptosis.98 According to the results of Ham et al, silibinin could elevate the expression levels of both IRE1α and PERK in human endometriosis-derived cells. Meanwhile, their downstream molecules, such as eIF2α and GADD153/CHOP, were also elevated, ultimately resulted in apoptosis. Additionally, they suggested that silibinin could inhibit the proliferation of human endometriosis cells by regulating the MAPK signaling pathway.258
Tetramethylpyrazine
Tetramethylpyrazine (TMP, Figure 9), also known as ligustrazine, is a compound originally isolated from the Chinese herbal medicine chuanxiong.259 TMP exerts the effects of inhibition of platelet aggregation,260 antitumor,261 anti-angiogenesis,262 antioxidant,263 and anti-inflammation.259 Previous studies have shown that activated platelets contribute to EMT and FMT in endometriosis cells.249 E-cadherin is a marker for epithelial-like cells, while vimentin and α-smooth muscle actin (α-SMA) are markers for mesenchymal-like cells.158,264 Activated platelets can release TGF-β, which is associated with fibrosis.248,265 TGF-β can activate the expression of EMT-related transcription factors through the TGF-β/Smad signaling pathway, which in turn promotes cell invasion and metastasis.249 Type I collagen (Col-I) is a marker of collagen fibers. And α-SMA is a marker of smooth muscle cells and myofibroblasts, which are the important effector cells in the process of fiber formation. Fibroblasts can transform into myofibroblasts under the stimulation of TGF-β. Myofibroblasts can upregulate the expression of α-SMA and stimulate the production of Col-I, which in turn drives cell contraction and collagen production, ultimately promoting tissue fibrosis.266,267 TMP treatment increased the E-cadherin expression in a dose dependent manner. TMP could significantly inhibit the aggregation of platelets, the expression of TGF-β, α-SMA, and Col-I in vivo. Meanwhile, the weight of ectopic endometrial lesions and the degree of fibrosis were reduced in endometriosis mouse models. Taken these results together, TMP can slow the progression of endometriosis and fibrosis by inhibiting platelet activation, platelet-induced TGF-β, EMT, FMT, cell contraction, and collagen production.268
Ursolic Acid
Ursolic acid (UA, Figure 9) is a pentacyclic triterpenoid purified from medicinal plants. It is also extensively present in various fruits and vegetables.269 UA has several pharmacological activities, principally including antioxidant, anti-inflammatory, and antitumor effects.270 Stimulation with COX-2 results in overexpression of PGE2, a key promotor of the expression of STAR and aromatase. Abnormal expression of STAR and aromatase will lead to high levels of estradiol production, which may promote the occurrence and development of endometriosis.2 Li et al showed that UA had anti-proliferation, pro-apoptosis, and anti-angiogenesis effects. Treatment with UA significantly decreased COX-2 expression and subsequent PGE2 production in human primary endometriotic stromal cells, while significantly enhanced the expression of caspase-3. Meanwhile, VEGF was decreased after the treatment of UA. The result of capillary formation experiments also supported the inhibitory effect of UA on angiogenesis. In addition, the negative impact of UA against endometriosis may attribute to the regulation of the JNK and p38 signaling pathways.271
Wogonin
Scutellaria baicalensis is one of the most common herbs in traditional Chinese medicine. Wogonin (WG, Figure 9), known as a kind of flavonoid, is abundantly identified from the root of Scutellaria baicalensis.272,273 It has a wide range of biological properties, taking antitumor,273 anti-inflammatory,274 and pro-apoptotic275 properties as examples. Estrogen enhances the proliferation and survival of endometriotic tissues.2 Ferella et al investigated that WG could significantly inhibit the expression of estrogen receptor α in THESC and could induce cell cycle arrest at the G2/M phase, thereby inhibiting cell proliferation. Additionally, WG increased intracellular ROS accumulation and promoted cell apoptosis. WG significantly reduced the area of ectopic endometrial lesions in vivo.113
Discussion and Conclusion
Endometriosis is a benign disease and usually occurs in women of childbearing age.276 The common clinical symptoms of endometriosis are pelvic pain, discomfort during intercourse, pelvic masses, and infertility.12,15 The common management options in clinical practice currently are pharmacotherapy and surgery.29 The majority of conventional pharmacotherapy treatments are hormonal drugs and NSAIDs. Hormonal drugs are used to induce pseudopregnancy, pseudomenopause, or suppress ovarian function.32 However, these drug treatments interfere with ovulation and prevent conception in patients of reproductive age who have reproductive needs. Hormonal drugs can generate serious adverse effects when used for a long-term treatment.22,24–28,31,32 Side effects such as gastrointestinal reactions and neurological symptoms may occur after the use of NSAIDs.20 Even though endometriosis lesions can be surgically removed, the disease has a high recurrence rate after surgical removal.33 On the other hand, the ovarian cortex may be damaged during surgery, with the postoperative decline in ovarian reserve function.29,31 Additionally, the surgery is an invasive procedure into the abdominal cavity, which may inevitably lead to pelvic and/or abdominal adhesions. Pelvic and/or abdominal adhesions are also detrimental to the patient’s pregnancy. Thus, the development of novel, effective, safe, low side effects and affordable drugs for individualized treatment is urgently needed, preferably ones that can be used for a long time.
Natural materials are a vital source for the discovery and development of new drugs. As described in Table S3–S6, multi-component herbal preparations, extracts of medicinal plants, and purified compounds from plants/herbs, have been shown to target a variety of cellular processes, such as cell proliferation, apoptosis, inflammation, autophagy, oxidative stress, adhesion, migration, invasion, fibrosis, estrogen and progesterone receptor expression, and angiogenesis, which are critical in the pathogenesis of endometriosis. The anti-endometriosis effects involve plentiful molecular mechanisms and targets, including anti-proliferative (CCNE1, CDK2, and CDK4), anti-inflammatory (PGE2, COX-2, TNF-α, NO, IL-1β, IL-6, and IL-8), anti-angiogenic (VEGF, matrix metalloproteinase and Ang-1), pro-apoptotic (Bax, Bak, Bcl-2, Bcl-xL), autophagy (Beclin 1, LC3-II/LC3-I), endoplasmic reticulum stress, endoplasmic reticulum apoptotic pathway, mitochondrial apoptotic pathway and caspase cascade, anti-migratory invasion (MMP-2, MMP-9, TIMP1, E-cadherin, N-cadherin, Snail and Slug), regulation of estrogen and progesterone receptors, and anti-fibrotic (E-calmodulin, α-SMA, type I collagen, wave proteins and fibronectin). The signaling pathways involved in these effects are mainly the Hippo pathway, TGF-β/Smad signaling pathway, NF-κB, MAPK, PI3K/Akt, and JAK2/STAT3 signaling pathway. Medicinal plant extracts and multi-component herbal preparations contain various bioactive compounds. Therefore, they have infinite potential as multi-targets drugs. They may also affect multiple signaling pathways simultaneously, which could synergistically enhance their efficacy in the treatment of endometriosis. As mentioned above, the structural modification of bioactive lead compounds also has enormous potential to fight diseases.
Even the bioactive extracts and compounds mentioned in this review have shown interesting efficacy in the treatment of endometriosis, but most of the studies are based on preclinical cellular experiments and animal model experiments, namely, few clinical trials have been conducted. Almost all the animal experiments covered in this review were carried out in mice or rats, and these artificial animal models of endometriosis do not fully simulate the development of endometriosis, because there are obvious differences in the physiological mechanisms of reproduction between humans and animals. Therefore, more researches in the future are also needed to establish models in suitable animals, such as orangutans, which have similar physiological structures and physiological mechanisms as humans. Furthermore, serious adverse effects have not been elucidated in depth. In another aspect, most plant-derived bioactive compounds have low stability and bioavailability in vivo in their natural form, which make their effective use in patients with endometriosis challenging. More excitingly, some basic parameters of Chinese herbal medicines and natural compounds, such as chemicals, targets, oral bioavailability and blood-brain barrier, can be retrieved in the Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP) (Table S7). In conclusion, more detailed comprehensive pharmacodynamic and pharmacokinetic researches are extremely necessary in the future, as well as further clinical trials to confirm the efficacy and safety of these therapeutic herbal preparations, extracts and compounds.
Funding
This study was supported by the grants from the Doctoral Research Initiation Fund of the Affiliated Hospital of Southwest Medical University (No. 18061), the Fund for High-level Talents in Luzhou City (No. 02/00180095, 02/00180117), the grants for Application of Simulation Based Mastery Learning (SBML) Teaching Method in Standardized Training of Gynecological Laparoscopic Technique (2022 Municipal Guiding Science and Technology Plan Project). Figures 1–4 and 7 in this review were designed and created by Figdraw (www.figdraw.com).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wang Y, Nicholes K, Shih IM. The origin and pathogenesis of endometriosis. Annu Rev Pathol. 2020;15:71–95. doi:10.1146/annurev-pathmechdis-012419-032654
2. Bulun SE, Yilmaz BD, Sison C, et al. Endometriosis. Endocr Rev. 2019;40(4):1048–1079. doi:10.1210/er.2018-00242
3. Lai ZZ, Yang HL, Ha SY, et al. Cyclooxygenase-2 in Endometriosis. Int J Biol Sci. 2019;15(13):2783–2797. doi:10.7150/ijbs.35128
4. Laschke MW, Menger MD. Basic mechanisms of vascularization in endometriosis and their clinical implications. Hum Reprod Update. 2018;24(2):207–224. doi:10.1093/humupd/dmy001
5. Samimi M, Pourhanifeh MH, Mehdizadehkashi A, Eftekhar T, Asemi Z. The role of inflammation, oxidative stress, angiogenesis, and apoptosis in the pathophysiology of endometriosis: basic science and new insights based on gene expression. J Cell Physiol. 2019;234(11):19384–19392. doi:10.1002/jcp.28666
6. Park S, Lim W, Bazer FW, Whang KY, Song G. Quercetin inhibits proliferation of endometriosis regulating cyclin D1 and its target microRNAs in vitro and in vivo. J Nutr Biochem. 2019;63:87–100. doi:10.1016/j.jnutbio.2018.09.024
7. Hirakawa T, Nasu K, Miyabe S, et al. β-catenin signaling inhibitors ICG-001 and C-82 improve fibrosis in preclinical models of endometriosis. Sci Rep. 2019;9(1):20056. doi:10.1038/s41598-019-56302-4
8. Li H, Ma RQ, Cheng HY, Ye X, Zhu HL, Chang XH. Fibrinogen alpha chain promotes the migration and invasion of human endometrial stromal cells in endometriosis through focal adhesion kinase/protein kinase B/matrix metallopeptidase 2 pathway. Biol Reprod. 2020;103(4):779–790. doi:10.1093/biolre/ioaa126
9. Qi S, Yan L, Liu Z, et al. Melatonin inhibits 17β-estradiol-induced migration, invasion and epithelial-mesenchymal transition in normal and endometriotic endometrial epithelial cells. Reprod Biol Endocrinol. 2018;16(1):62. doi:10.1186/s12958-018-0375-5
10. Amini L, Chekini R, Nateghi MR, et al. The effect of combined vitamin C and Vitamin E supplementation on oxidative stress markers in women with endometriosis: a randomized, triple-blind placebo-controlled clinical trial. Pain Res Manag. 2021;2021:5529741. doi:10.1155/2021/5529741
11. Koninckx PR, Ussia A, Adamyan L, Wattiez A, Gomel V, Martin DC. Pathogenesis of endometriosis: the genetic/epigenetic theory. Fertil Steril. 2019;111(2):327–340. doi:10.1016/j.fertnstert.2018.10.013
12. Taylor HS, Adamson GD, Diamond MP, et al. An evidence-based approach to assessing surgical versus clinical diagnosis of symptomatic endometriosis. Int J Gynaecol Obstet. 2018;142(2):131–142. doi:10.1002/ijgo.12521
13. Flores I, Abreu S, Abac S, Fourquet J, Laboy J, Ríos-Bedoya C. Self-reported prevalence of endometriosis and its symptoms among Puerto Rican women. Int J Gynaecol Obstet. 2008;100(3):257–261. doi:10.1016/j.ijgo.2007.08.010
14. Martire FG, Lazzeri L, Conway F, et al. Adolescence and endometriosis: symptoms, ultrasound signs and early diagnosis. Fertil Steril. 2020;114(5):1049–1057. doi:10.1016/j.fertnstert.2020.06.012
15. Agarwal SK, Chapron C, Giudice LC, et al. Clinical diagnosis of endometriosis: a call to action. Am J Obstet Gynecol. 2019;220(4):354.e1–354.e12. doi:10.1016/j.ajog.2018.12.039
16. Huchon C, Aubry G, Ploteau S, Fauconnier A. Specific clinical signs suggestive of endometriosis (excluding adenomyosis) and questionnaires of symptoms, pain and quality of life: CNGOF-HAS Endometriosis Guidelines. Gynecol Obstet Fertil Senol. 2018;46(3):168–176. doi:10.1016/j.gofs.2018.02.022
17. DiVasta AD, Vitonis AF, Laufer MR, Missmer SA. Spectrum of symptoms in women diagnosed with endometriosis during adolescence vs adulthood. Am J Obstet Gynecol. 2018;218(3):324.e1–324.e11. doi:10.1016/j.ajog.2017.12.007
18. Rolla E. Endometriosis: advances and controversies in classification, pathogenesis, diagnosis, and treatment. F1000Res. 2019;8. doi:10.12688/f1000research.14817.1
19. Menni K, Facchetti L, Cabassa P. Extragenital endometriosis: assessment with MR imaging. A pictorial review. Br J Radiol. 2016;89(1060):20150672. doi:10.1259/bjr.20150672
20. Brown J, Crawford TJ, Allen C, Hopewell S, Prentice A. Nonsteroidal anti-inflammatory drugs for pain in women with endometriosis. Cochrane Database Syst Rev. 2017;1(1):Cd004753. doi:10.1002/14651858.CD004753.pub4
21. Allen C, Hopewell S, Prentice A. Non-steroidal anti-inflammatory drugs for pain in women with endometriosis. Cochrane Database Syst Rev. 2005;4:Cd004753.
22. Brown J, Crawford TJ, Datta S, Prentice A. Oral contraceptives for pain associated with endometriosis. Cochrane Database Syst Rev. 2018;5(5):Cd001019. doi:10.1002/14651858.CD001019.pub3
23. Hee L, Kettner LO, Vejtorp M. Continuous use of oral contraceptives: an overview of effects and side-effects. Acta Obstet Gynecol Scand. 2013;92(2):125–136. doi:10.1111/aogs.12036
24. Buggio L, Somigliana E, Barbara G, Frattaruolo MP, Vercellini P. Oral and depot progestin therapy for endometriosis: towards a personalized medicine. Expert Opin Pharmacother. 2017;18(15):1569–1581. doi:10.1080/14656566.2017.1381086
25. Fu J, Song H, Zhou M, et al. Progesterone receptor modulators for endometriosis. Cochrane Database Syst Rev. 2017;7(7):Cd009881. doi:10.1002/14651858.CD009881.pub2
26. Godin R, Marcoux V. Vaginally administered danazol: an overlooked option in the treatment of rectovaginal endometriosis? J Obstet Gynaecol Can. 2015;37(12):1098–1103. doi:10.1016/S1701-2163(16)30075-5
27. Bedaiwy MA, Allaire C, Alfaraj S. Long-term medical management of endometriosis with dienogest and with a gonadotropin-releasing hormone agonist and add-back hormone therapy. Fertil Steril. 2017;107(3):537–548. doi:10.1016/j.fertnstert.2016.12.024
28. Garzon S, Laganà AS, Barra F, et al. Aromatase inhibitors for the treatment of endometriosis: a systematic review about efficacy, safety and early clinical development. Expert Opin Investig Drugs. 2020;29(12):1377–1388. doi:10.1080/13543784.2020.1842356
29. Kalaitzopoulos DR, Samartzis N, Kolovos GN, et al. Treatment of endometriosis: a review with comparison of 8 guidelines. BMC Women's Health. 2021;21(1):397. doi:10.1186/s12905-021-01545-5
30. Vannuccini S, Clemenza S, Rossi M, Petraglia F. Hormonal treatments for endometriosis: the endocrine background. Rev Endocr Metab Disord. 2022;23(3):333–355. doi:10.1007/s11154-021-09666-w
31. Dunselman GA, Vermeulen N, Becker C, et al. ESHRE guideline: management of women with endometriosis. Hum Reprod. 2014;29(3):400–412. doi:10.1093/humrep/det457
32. Falcone T, Flyckt R. Clinical management of endometriosis. Obstet Gynecol. 2018;131(3):557–571. doi:10.1097/AOG.0000000000002469
33. Guo SW. Recurrence of endometriosis and its control. Hum Reprod Update. 2009;15(4):441–461. doi:10.1093/humupd/dmp007
34. Practice Committee of the American Society for Reproductive Medicine. Endometriosis and infertility: a committee opinion. Fertil Steril. 2012;98(3):591–598. doi:10.1016/j.fertnstert.2012.05.031
35. Mishra BB, Tiwari VK. Natural products: an evolving role in future drug discovery. Eur J Med Chem. 2011;46(10):4769–4807. doi:10.1016/j.ejmech.2011.07.057
36. Koehn FE, Carter GT. The evolving role of natural products in drug discovery. Nat Rev Drug Discov. 2005;4(3):206–220. doi:10.1038/nrd1657
37. Zhang L, Song JK, Kong LL, et al. The strategies and techniques of drug discovery from natural products. Pharmacol Ther. 2020;216:107686. doi:10.1016/j.pharmthera.2020.107686
38. Atanasov AG, Zotchev SB, Dirsch VM, Supuran CT. Natural products in drug discovery: advances and opportunities. Nat Rev Drug Discov. 2021;20(3):200–216. doi:10.1038/s41573-020-00114-z
39. Hang YY, Tan L, Cao Y, Zhuang MF, Jin YL. Mechanism of Cai’s Neiyi Prescription regulating TRIM24/NLRP3 inflammasome-mediated endometriotic pyroptosis in rats. Shanghai J TraditChin Med. 2021;55(09):74–80.
40. Li SD, Zhu YP, Zhang TT, Hang YY, Chen Q, Jin YL. Cai’s Neiyi Prescription promotes apoptosis and inhibits inflammation in endometrial stromal cells with endometriosis through inhibiting USP10. Biotechnol Appl Biochem. 2019;66(2):231–239. doi:10.1002/bab.1715
41. Su HN, Liu J, He K, Liu LH, Li CX. Effects of Danefukang decocted extract on the expression of VEGF and TNF-α in endometriosis rats. Pharmacology and Clinics of Chinese Materia Medica. 2013;29(06):128–131.
42. Fu XY, Wei SB, Feng TT. Different effects of Danefukang Decoction on the expression of MMP-2 and VEGF in eutopic and ectopic endometrium of adenomyoma mice. Chinese Traditional Patent Medicine. 2013;35(11):2523–2525.
43. Zhong YC, Zhou XF, Hou CM, Li WP. Effect of danefukang on symptoms and biomarkers in women with endometriosis. Taiwan J Obstet Gynecol. 2019;58(2):218–222. doi:10.1016/j.tjog.2019.01.009
44. Dai YF, Qiang WJ, Yu XK, et al. Guizhi Fuling Decoction inhibiting the PI3K and MAPK pathways in breast cancer cells revealed by HTS(2) technology and systems pharmacology. Comput Struct Biotechnol J. 2020;18:1121–1136. doi:10.1016/j.csbj.2020.05.004
45. Gao Y, Ji WJ, Lu M, et al. Systemic pharmacological verification of Guizhi Fuling decoction in treating endometriosis-associated pain. J Ethnopharmacol. 2022;297:115540. doi:10.1016/j.jep.2022.115540
46. Zhang LM, Zhao L, Chen L, Xia M. A clinical study on treating endometriosis with Huangzhi Neiyi capsule. Clinical Journal of Chinese Medicine. 2017;9(30):90–92.
47. Ahluwalia A, Tarnawski AS. Critical role of hypoxia sensor--HIF-1α in VEGF gene activation. Implications for angiogenesis and tissue injury healing. Curr Med Chem. 2012;19(1):90–97. doi:10.2174/092986712803413944
48. Liu HL, Sun XL, Zhao Y, Xia M, Wang CX. Anti-angiogenesis effect and mechanism study of Huangzhi Neiyi capsule in a rat endometriosis model. J Int Med Res. 2020;48(1):300060519899767. doi:10.1177/0300060519899767
49. Wei SB, Cao YF, Wang Y. Effect of Neiyi Kangfu suppository on expressions of cytochrome C and survivin in ectopic and eutopic endometrium in rats with endometriosis. Zhongguo Zhong Xi Yi Jie He Za Zhi. 2008;28(2):139–141.
50. Yeung K, Seitz T, Li S, et al. Suppression of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature. 1999;401(6749):173–177. doi:10.1038/43686
51. Wen Y, Fan LX, Pang LL, et al. NeiyiKangfu tablets control the progression of endometriosis through inhibiting RAF/MEK/ERK signal pathway by targeting RKIP. Gynecol Endocrinol. 2022;38(12):1136–1146. doi:10.1080/09513590.2022.2162495
52. Yang HD, Xia MT, Wu YP, Wang XE. Effect of medicated serum of Qiu’s Neiyi Recipe on angiogenic ability of human umbilical vein endothelial cells. Chinese Archives of Traditional Chinese Medicine. 2016;34(10):2545–2547.
53. Hung SW, Zhang R, Tan Z, Chung JPW, Zhang T, Wang CC. Pharmaceuticals targeting signaling pathways of endometriosis as potential new medical treatment: a review. Med Res Rev. 2021;41(4):2489–2564.
54. Mizushima N. Methods for monitoring autophagy. Int J Biochem Cell Biol. 2004;36(12):2491–2502. doi:10.1016/j.biocel.2004.02.005
55. Yang HD, Zhu QF, Li H, Jiang XL, Xu XQ, Guo Y. Effect of Neiyi Prescription of QIU on autophagy and angiogenic ability of endometriosis via the PPARγ/NF-κB signaling pathway. Arch Gynecol Obstet. 2022;306(2):533–545.
56. Chen YH, Mao HY, Wu QS, et al. Mechanism of Shaofu Zhuyu Decoction in treatment of endometriosis-associated dysmenorrhea with syndrom of cold coagulation and blood stasis based on MSK1/2. China Journal of Chinese Materia Medica. 2022;47:17 4674–4681.
57. Zhu GH, Jiang CH, Yan X, Zhao S, Xu DJ, Cao Y. Shaofu Zhuyu decoction regresses endometriotic lesions in a rat model. Evid Based Complement Alternat Med. 2018;2018:3927096. doi:10.1155/2018/3927096
58. Nagamatsu T, Fujii T, Schust DJ, et al. Tokishakuyakusan, a traditional Japanese medicine (Kampo) mitigates iNKT cell-mediated pregnancy loss in mice. Am J Reprod Immunol. 2018;80(4):e13021. doi:10.1111/aji.13021
59. Nagira K, Taniguchi F, Nakamura K, et al. Tokishakuyakusan, a Kampo medicine, attenuates endometriosis-like lesions and hyperalgesia in murine with endometriosis-like symptoms. Am J Reprod Immunol. 2019;82(5):e13182. doi:10.1111/aji.13182
60. Takeuchi A, Koga K, Tokita Y, et al. The effects of tokishakuyakusan, a traditional Japanese medicine (kampo), ferulic acid and paeoniflorin, on human endometriotic stromal cells and peritoneal macrophages. J Reprod Immunol. 2020;139:103104. doi:10.1016/j.jri.2020.103104
61. Bina F, Daglia M, Santarcangelo C, et al. Phytochemical profiling and ameliorative effects of Achillea cretica L. on rat model of endometriosis. J Ethnopharmacol. 2020;254:112747. doi:10.1016/j.jep.2020.112747
62. Romano R, De Luca L, Aiello A, et al. Basil (Ocimum basilicum L.) Leaves as a Source of Bioactive Compounds. Foods. 2022;11:20. doi:10.3390/foods11203212
63. A’Yuni DQ, A S, Widjiati W. Ethanol extract of basil (Ocimum Basilicum L.) leaves inhibits endometriosis growth in a mouse model by modulating vascular endothelial growth factor (VEGF) expression. J Med Life. 2023;16(8):1224–1230.
64. Siddiqui MZ. Boswellia serrata, a potential antiinflammatory agent: an overview. Indian J Pharm Sci. 2011;73(3):255–261. doi:10.4103/0250-474X.93507
65. Jaramillo MC, Zhang DD. The emerging role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev. 2013;27(20):2179–2191.
66. D’Amico R, Impellizzeri D, Cordaro M, et al. Regulation of apoptosis and oxidative stress by oral boswellia serrata gum resin extract in a rat model of endometriosis. Int J Mol Sci. 2022;23:23.
67. Soliman S, Mohammad M, El-Keblawy A, et al. Mechanical and phytochemical protection mechanisms of Calligonum comosum in arid deserts. PLoS One. 2018;13(2):e0192576. doi:10.1371/journal.pone.0192576
68. Alehaideb Z, AlGhamdi S, Yahya WB, et al. Anti-proliferative and pro-apoptotic effects of calligonum comosum (l’her.) methanolic extract in human triple-negative MDA-MB-231 breast cancer cells. J Evid Based Integr Med. 2020;25:2515690x20978391. doi:10.1177/2515690X20978391
69. Abdel-Sattar EA, Mouneir SM, Asaad GF, Abdallah HM. Protective effect of Calligonum comosum on haloperidol-induced oxidative stress in rat. Toxicol Ind Health. 2014;30(2):147–153. doi:10.1177/0748233712452601
70. Alzahrani AJ. Potent antioxidant and anticancer activities of the methanolic extract of Calligonum comosum (L’Her) fruit hairs against human hepatocarcinoma cells. Saudi J Biol Sci. 2021;28(9):5283–5289. doi:10.1016/j.sjbs.2021.05.053
71. Liu XM, Zakaria MN, Islam MW, et al. Anti-inflammatory and anti-ulcer activity of Calligonum comosum in rats. Fitoterapia. 2001;72(5):487–491. doi:10.1016/S0367-326X(01)00271-4
72. Kiani K, Rudzitis-Auth J, Scheuer C, et al. Calligonum comosum (Escanbil) extract exerts anti-angiogenic, anti-proliferative and anti-inflammatory effects on endometriotic lesions. J Ethnopharmacol. 2019;239:111918. doi:10.1016/j.jep.2019.111918
73. Seo EJ, Lee DU, Kwak JH, Lee SM, Kim YS, Jung YS. Antiplatelet effects of Cyperus rotundus and its component (+)-nootkatone. J Ethnopharmacol. 2011;135(1):48–54. doi:10.1016/j.jep.2011.02.025
74. Taheri Y, Herrera-Bravo J, Huala L, et al. Cyperus spp.: a review on phytochemical composition, biological activity, and health-promoting effects. Oxid Med Cell Longev. 2021;2021:4014867. doi:10.1155/2021/4014867
75. Lee CH, Hwang DS, Kim HG, et al. Protective effect of Cyperi rhizoma against 6-hydroxydopamine-induced neuronal damage. J Med Food. 2010;13(3):564–571. doi:10.1089/jmf.2009.1252
76. Wang FK, Song X, Ma SS, et al. Cyperus rotundusThe treatment role of L. to triple-negative breast cancer cells. Biosci Rep. 2019;39:6.
77. Kim HG, Hong J, Huh Y, et al. Cyperi Rhizoma inhibits the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced reduction in nigrostriatal dopaminergenic neurons in estrogen-deprived mice. J Ethnopharmacol. 2013;148(1):322–328. doi:10.1016/j.jep.2013.04.038
78. Chen GT, Tai CT, Yeh LS, Yang TC, Tsai HD. Identification of the cadherin subtypes present in the human peritoneum and endometriotic lesions: potential role for P-cadherin in the development of endometriosis. Mol Reprod Dev. 2002;62(3):289–294. doi:10.1002/mrd.10121
79. Shaco-Levy R, Sharabi S, Benharroch D, Piura B, Sion-Vardy N. Matrix metalloproteinases 2 and 9, E-cadherin, and beta-catenin expression in endometriosis, low-grade endometrial carcinoma and non-neoplastic eutopic endometrium. Eur J Obstet Gynecol Reprod Biol. 2008;139(2):226–232. doi:10.1016/j.ejogrb.2008.01.004
80. Kim JH, Yang YI, Ahn JH, Lee JG, Lee KT, Choi JH. Deer (Cervus elaphus) antler extract suppresses adhesion and migration of endometriotic cells and regulates MMP-2 and MMP-9 expression. J Ethnopharmacol. 2012;140(2):391–397. doi:10.1016/j.jep.2012.01.032
81. Kobayashi H, Yamada Y, Morioka S, Niiro E, Shigemitsu A, Ito F. Mechanism of pain generation for endometriosis-associated pelvic pain. Arch Gynecol Obstet. 2014;289(1):13–21. doi:10.1007/s00404-013-3049-8
82. Ahn JH, Choi JM, Kang ES, et al. The anti-endometriotic effect of cyperi rhizoma extract, inhibiting cell adhesion and the expression of pain-related factors through akt and NF-kB pathways. Medicina. 2022;58:3.
83. Liang HL, Liu BY, Wu C, et al. Effects of light intensity on the growth of Polygala fallax Hemsl (Polygalaceae). Front Plant Sci. 2022;13:985628. doi:10.3389/fpls.2022.985628
84. Zhong CM, Ju GCH, Yang SF, Zhao XP, Chen JX, Li N. Total flavonoids of polygala fallax hemsl induce apoptosis of human ectopic endometrial stromal cells through PI3K/AKT/Bcl-2 signaling pathway. Gynecol Obstet Invest. 2023;88(4):197–213. doi:10.1159/000530104
85. Wong KH, Li GQ, Li KM, Razmovski-Naumovski V, Chan K. Kudzu root: traditional uses and potential medicinal benefits in diabetes and cardiovascular diseases. J Ethnopharmacol. 2011;134(3):584–607. doi:10.1016/j.jep.2011.02.001
86. Takano A, Kamiya T, Tsubata M, Ikeguchi M, Takagaki K, Kinjo J. Oral toxicological studies of pueraria flower extract: acute toxicity study in mice and subchronic toxicity study in rats. J Food Sci. 2013;78(11):T1814–21. doi:10.1111/1750-3841.12263
87. Yao M, Liao Y, Li GQ, Law FC, Tang Y. Quantitative analysis of two isoflavones in Pueraria lobata flowers from eleven Chinese provinces using high performance liquid chromatography. Chin Med. 2010;5:14. doi:10.1186/1749-8546-5-14
88. Kim JH, Woo JH, Kim HM, Oh MS, Jang DS, Choi JH. Anti-endometriotic effects of pueraria flower extract in human endometriotic cells and mice. Nutrients. 2017;9:3.
89. Li T, Li YG, Pu DM. Matrix metalloproteinase-2 and −9 expression correlated with angiogenesis in human adenomyosis. Gynecol Obstet Invest. 2006;62(4):229–235. doi:10.1159/000094426
90. Ahn H, Han B, Hong E, et al. Korean Red Ginseng attenuates ultraviolet-mediated inflammasome activation in keratinocytes. J Ginseng Res. 2021;45(3):456–463. doi:10.1016/j.jgr.2021.02.002
91. Kim JK, Shin KK, Kim H, et al. Korean Red Ginseng exerts anti-inflammatory and autophagy-promoting activities in aged mice. J Ginseng Res. 2021;45(6):717–725. doi:10.1016/j.jgr.2021.03.009
92. Choi JH, Jang M, Kim EJ, et al. Korean Red Ginseng alleviates dehydroepiandrosterone-induced polycystic ovarian syndrome in rats via its antiinflammatory and antioxidant activities. J Ginseng Res. 2020;44(6):790–798. doi:10.1016/j.jgr.2019.08.007
93. Baek KS, Yi YS, Son YJ, et al. Comparison of anticancer activities of Korean Red Ginseng-derived fractions. J Ginseng Res. 2017;41(3):386–391. doi:10.1016/j.jgr.2016.11.001
94. Brichant G, Laraki I, Henry L, Munaut C, Nisolle M. New therapeutics in endometriosis: a review of hormonal, non-hormonal, and non-coding RNA treatments. Int J Mol Sci. 2021;22:19. doi:10.3390/ijms221910498
95. Bjorkman S, Taylor HS. MicroRNAs in endometriosis: biological function and emerging biomarker candidates. Biol Reprod. 2019;100(5):1135–1146. doi:10.1093/biolre/ioz014
96. Wu JN, Huang HY, Huang W, Wang L, Xia XM, Fang XL. Analysis of exosomal lncRNA, miRNA and mRNA expression profiles and ceRNA network construction in endometriosis. Epigenomics. 2020;12(14):1193–1213. doi:10.2217/epi-2020-0084
97. Yao N, Li YJ, Zhang DM, et al. B4G2 induces mitochondrial apoptosis by the ROS-mediated opening of Ca(2+)-dependent permeability transition pores. Cell Physiol Biochem. 2015;37(3):838–852. doi:10.1159/000430212
98. He Q, Zhou XJ, Liu Y, et al. Titanium dioxide nanoparticles induce mouse hippocampal neuron apoptosis via oxidative stress- and calcium imbalance-mediated endoplasmic reticulum stress. Environ Toxicol Pharmacol. 2018;63:6–15. doi:10.1016/j.etap.2018.08.003
99. Park JH, Lee SK, Kim MK, et al. Saponin extracts induced apoptosis of endometrial cells from women with endometriosis through modulation of miR-21-5p. Reprod Sci. 2018;25(2):292–301. doi:10.1177/1933719117711263
100. Mao QQ, Xu XY, Cao SY, et al. Bioactive compounds and bioactivities of Ginger (Zingiber officinale Roscoe). Foods. 2019;8:6. doi:10.3390/foods8060185
101. Filho JMM, Neto JN, Gomes L, et al. Zingiber officinale Roscoe (Ginger) as a complementary option for clinical treatment of endometriosis: an experimental study in rats. J Med Food. 2021;24(4):342–347. doi:10.1089/jmf.2019.0320
102. Dhouibi R, Affes H, Ben salem M, et al. Screening of pharmacological uses of Urtica dioica and others benefits. Prog Biophys Mol Bio. 2020;150:67–77. doi:10.1016/j.pbiomolbio.2019.05.008
103. Ilhan M, Ali Z, Khan IA, Taştan H, Küpeli Akkol E. Bioactivity-guided isolation of flavonoids from Urtica dioica L. and their effect on endometriosis rat model. J Ethnopharmacol. 2019;243:112100. doi:10.1016/j.jep.2019.112100
104. Hostetler GL, Ralston RA, Schwartz SJ. Flavones: food sources, bioavailability, metabolism, and bioactivity. Adv Nutr. 2017;8(3):423–435. doi:10.3945/an.116.012948
105. Imran M, Aslam Gondal T, Atif M, et al. Apigenin as an anticancer agent. Phytother Res. 2020;34(8):1812–1828. doi:10.1002/ptr.6647
106. Park CH, Min SY, Yu HW, et al. Effects of Apigenin on RBL-2H3, RAW264.7, and HaCaT cells: anti-allergic, anti-inflammatory, and skin-protective activities. Int J Mol Sci. 2020;21:13.
107. Yang JL, Pi CC, Wang GH. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomed Pharmacother. 2018;103:699–707. doi:10.1016/j.biopha.2018.04.072
108. Park S, Lim W, Bazer FW, Song G. Apigenin induces ROS-dependent apoptosis and ER stress in human endometriosis cells. J Cell Physiol. 2018;233(4):3055–3065. doi:10.1002/jcp.26054
109. Ou JY, Huang JQ, Wang MF, Ou SY. Effect of rosmarinic acid and carnosic acid on AGEs formation in vitro. Food Chem. 2017;221:1057–1061. doi:10.1016/j.foodchem.2016.11.056
110. Birtić S, Dussort P, Pierre FX, Bily AC, Roller M. Carnosic acid. Phytochemistry. 2015;115:9–19. doi:10.1016/j.phytochem.2014.12.026
111. Satoh T, Trudler D, Oh C, Lipton S. Potential therapeutic use of the rosemary diterpene carnosic acid for alzheimer’s disease, parkinson’s disease, and Long-COVID through NRF2 activation to counteract the NLRP3 inflammasome. Antioxidants. 2022;11:1.
112. Moore J, Yousef M, Tsiani E. Anticancer effects of rosemary (Rosmarinus officinalis L.) extract and rosemary extract polyphenols. Nutrients. 2016;8(11):731. doi:10.3390/nu8110731
113. Ferella L, Bastón JI, Bilotas MA, et al. Active compounds present inRosmarinus officinalis leaves andScutellaria baicalensis root evaluated as new therapeutic agents for endometriosis. Reprod Biomed Online. 2018;37(6):769–782. doi:10.1016/j.rbmo.2018.09.018
114. Mani R, Natesan V. Chrysin: sources, beneficial pharmacological activities, and molecular mechanism of action. Phytochemistry. 2018;145:187–196. doi:10.1016/j.phytochem.2017.09.016
115. Naz S, Imran M, Rauf A, et al. Chrysin: pharmacological and therapeutic properties. Life Sci. 2019;235:116797. doi:10.1016/j.lfs.2019.116797
116. Raina R, Afroze N, Kedhari Sundaram M, et al. Chrysin inhibits propagation of HeLa cells by attenuating cell survival and inducing apoptotic pathways. Eur Rev Med Pharmacol Sci. 2021;25(5):2206–2220. doi:10.26355/eurrev_202103_25253
117. Liao TY, Ding L, Wu P, et al. Chrysin Attenuates the NLRP3 inflammasome cascade to reduce synovitis and pain in KOA rats. Drug Des Devel Ther. 2020;14:3015–3027. doi:10.2147/DDDT.S261216
118. Li XY, Li S, Wang Q, Fu JJ, Yang YJ. Chrysin ameliorates sepsis-induced cardiac dysfunction through upregulating Nfr2/heme oxygenase 1 pathway. J Cardiovasc Pharmacol. 2021;77(4):491–500. doi:10.1097/FJC.0000000000000989
119. Ryu S, Bazer FW, Lim W, Song G. Chrysin leads to cell death in endometriosis by regulation of endoplasmic reticulum stress and cytosolic calcium level. J Cell Physiol. 2019;234(3):2480–2490. doi:10.1002/jcp.26770
120. Wei W, Rasul A, Sadiqa A, et al. Curcumol: from Plant Roots to Cancer Roots. Int J Biol Sci. 2019;15(8):1600–1609. doi:10.7150/ijbs.34716
121. Zhang JD, Su G, Tang ZW, et al. curcumol exerts anticancer effect in cholangiocarcinoma cells via down-regulating CDKL3. Front Physiol. 2018;9:234. doi:10.3389/fphys.2018.00234
122. Zheng Y, Wang L, Wang JH, Liu LL, Zhao TJ. Effect of Curcumol on NOD-like receptor thermoprotein domain 3 inflammasomes in liver fibrosis of mice. Chin J Integr Med. 2021;2021:1.
123. Yuan WW, Sun H, Yu L, Wang JB. Curcumol inhibits keloid fibroblast proliferation and collagen synthesis through the ERK signaling pathway. Nan Fang Yi Ke Da Xue Xue Bao. 2021;41(5):687–693. doi:10.12122/j.issn.1673-4254.2021.05.08
124. Wang Y, Nie XB, Liu SJ, Liu J, Bian WH. Curcumol attenuates endometriosis by inhibiting the JAK2/STAT3 signaling pathway. Med Sci Monit. 2022;28:e934914. doi:10.12659/MSM.934914
125. Xin P, Xu XY, Deng CJ, et al. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int Immunopharmacol. 2020;80:106210. doi:10.1016/j.intimp.2020.106210
126. Hu XY, Li J, Fu MR, Zhao X, Wang W. The JAK/STAT signaling pathway: from bench to clinic. Signal Transduct Target Therap. 2021;6(1):402. doi:10.1038/s41392-021-00791-1
127. Izumi T, Piskula MK, Osawa S, et al. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J Nutr. 2000;130(7):1695–1699. doi:10.1093/jn/130.7.1695
128. Liu T, Zhang L, Joo D, Sun SC. NF-κB signaling in inflammation. Signal Transduct Target Therap. 2017;2:17023. doi:10.1038/sigtrans.2017.23
129. Napetschnig J, Wu H. Molecular basis of NF-κB signaling. Annu Rev Biophys. 2013;42:443–468. doi:10.1146/annurev-biophys-083012-130338
130. Wu M, Zhang Y. MiR-182 inhibits proliferation, migration, invasion and inflammation of endometrial stromal cells through deactivation of NF-κB signaling pathway in endometriosis. Mol Cell Biochem. 2021;476(3):1575–1588. doi:10.1007/s11010-020-03986-2
131. Takaoka O, Mori T, Ito F, et al. Daidzein-rich isoflavone aglycones inhibit cell growth and inflammation in endometriosis. J Steroid Biochem Mol Biol. 2018;181:125–132. doi:10.1016/j.jsbmb.2018.04.004
132. Hu B, Wu FF, Shi ZL, et al. Dehydrocostus lactone attenuates osteoclastogenesis and osteoclast-induced bone loss by modulating NF-κB signalling pathway. J Cell Mol Med. 2019;23(8):5762–5770. doi:10.1111/jcmm.14492
133. Long HY, Huang QX, Yu YY, et al. Dehydrocostus lactone inhibits in vitro gastrinoma cancer cell growth through apoptosis induction, sub-G1 cell cycle arrest, DNA damage and loss of mitochondrial membrane potential. Arch Med Sci. 2019;15(3):765–773. doi:10.5114/aoms.2018.73128
134. Cai H, Qin XS, Yang CH. Dehydrocostus lactone suppresses proliferation of human chronic myeloid leukemia cells through Bcr/Abl-JAK/STAT signaling pathways. J Cell Biochem. 2017;118(10):3381–3390. doi:10.1002/jcb.25994
135. Xiong Y, Cui XC, Zhou YJ, et al. Dehydrocostus lactone inhibits BLM-induced pulmonary fibrosis and inflammation in mice via the JNK and p38 MAPK-mediated NF-κB signaling pathways. Int Immunopharmacol. 2021;98:107780. doi:10.1016/j.intimp.2021.107780
136. Woo JH, Ahn JH, Jang DS, Choi JH. Effect of dehydrocostus lactone isolated from the roots of aucklandia lappa on the apoptosis of endometriotic cells and the alternative activation of endometriosis-associated macrophages. Am J Chin Med. 2019;47(6):1289–1305. doi:10.1142/S0192415X19500666
137. Smyth E, Grosser T, Wang M, Yu Y, FitzGerald G. Prostanoids in health and disease. J Lipid Res. 2009;S423–8. doi:10.1194/jlr.R800094-JLR200
138. Murakami M, Kudo I. Recent advances in molecular biology and physiology of the prostaglandin E2-biosynthetic pathway. Prog Lipid Res. 2004;43(1):3–35. doi:10.1016/S0163-7827(03)00037-7
139. Laganà AS, Salmeri FM, Ban Frangež H, Ghezzi F, Vrtačnik-Bokal E, Granese R. Evaluation of M1 and M2 macrophages in ovarian endometriomas from women affected by endometriosis at different stages of the disease. Gynecol Endocrinol. 2020;36(5):441–444. doi:10.1080/09513590.2019.1683821
140. Miller JE, Ahn SH, Marks RM, et al. IL-17A modulates peritoneal macrophage recruitment and M2 polarization in endometriosis. Front Immunol. 2020;11:108. doi:10.3389/fimmu.2020.00108
141. Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11(10):889–896. doi:10.1038/ni.1937
142. Li HL, Huang N, Zhu WK, et al. Modulation the crosstalk between tumor-associated macrophages and non-small cell lung cancer to inhibit tumor migration and invasion by ginsenoside Rh2. BMC Cancer. 2018;18(1):579. doi:10.1186/s12885-018-4299-4
143. Gazzaniga S, Bravo AI, Guglielmotti A, et al. Targeting tumor-associated macrophages and inhibition of MCP-1 reduce angiogenesis and tumor growth in a human melanoma xenograft. J Invest Dermatol. 2007;127(8):2031–2041. doi:10.1038/sj.jid.5700827
144. Barkallah M, Nzoughet-Kouassi J, Simard G, et al. Enhancement of the anti-angiogenic effects of delphinidin when encapsulated within small extracellular vesicles. Nutrients. 2021;13:12. doi:10.3390/nu13124378
145. Sharma A, Choi HK, Kim YK, Lee HJ. Delphinidin and its glycosides’ war on cancer: preclinical perspectives. Int J Mol Sci. 2021;22:21. doi:10.3390/ijms222111500
146. Chen ZX, Zhang R, Shi WM, et al. The multifunctional benefits of naturally occurring delphinidin and its glycosides. J Agric Food Chem. 2019;67(41):11288–11306. doi:10.1021/acs.jafc.9b05079
147. Park S, Lim W, Song G. Delphinidin induces antiproliferation and apoptosis of endometrial cells by regulating cytosolic calcium levels and mitochondrial membrane potential depolarization. J Cell Biochem. 2019;120(4):5072–5084. doi:10.1002/jcb.27784
148. Chang J, Yang JY, Choi J, Jung HH, Im GJ. Calcium imaging in gentamicin ototoxicity: increased intracellular calcium relates to oxidative stress and late apoptosis. Int J Pediatr Otorhinolaryngol. 2011;75(12):1616–1622. doi:10.1016/j.ijporl.2011.09.027
149. Rizzuto R, Pinton P, Ferrari D, et al. Calcium and apoptosis: facts and hypotheses. Oncogene. 2003;22(53):8619–8627. doi:10.1038/sj.onc.1207105
150. Bauer TM, Murphy E. Role of mitochondrial calcium and the permeability transition pore in regulating cell death. Circ Res. 2020;126(2):280–293. doi:10.1161/CIRCRESAHA.119.316306
151. Ochiai W, Kobayashi H, Kitaoka S, et al. Effect of the active ingredient of Kaempferia parviflora, 5,7-dimethoxyflavone, on the pharmacokinetics of midazolam. J Nat Med. 2018;72(3):607–614. doi:10.1007/s11418-018-1184-z
152. Park W, Park MY, Song G, Lim W. 5,7-Dimethoxyflavone induces apoptotic cell death in human endometriosis cell lines by activating the endoplasmic reticulum stress pathway. Phytother Res. 2020;34(9):2275–2286. doi:10.1002/ptr.6677
153. Derosa G, Maffioli P, Sahebkar A. Ellagic acid and its role in chronic diseases. Adv Exp Med Biol. 2016;928:473–479.
154. Ríos J, Giner R, Marín M, Recio M. A pharmacological update of ellagic acid. Planta Med. 2018;84(15):1068–1093. doi:10.1055/a-0633-9492
155. Wang F, Chen JY, Xiang DB, Lian XJ, Wu CR, Quan J. Ellagic acid inhibits cell proliferation, migration, and invasion in melanoma via EGFR pathway. Am J Transl Res. 2020;12(5):2295–2304.
156. Mc Cormack BA, Bilotas MA, Madanes D, Ricci AG, Singla JJ, Barañao RI. Potential use of ellagic acid for endometriosis treatment: its effect on a human endometrial cell cycle, adhesion and migration. Food Funct. 2020;11(5):4605–4614. doi:10.1039/D0FO00267D
157. Kamal DAM, Salamt N, Zaid SSM, Mokhtar MH. Beneficial effects of green tea catechins on female reproductive disorders: a review. Molecules. 2021;26:9. doi:10.3390/molecules26092675
158. Vergara D, Simeone P, Franck J, et al. Translating epithelial mesenchymal transition markers into the clinic: novel insights from proteomics. EuPA Open Proteom. 2016;10:31–41. doi:10.1016/j.euprot.2016.01.003
159. Guan QH, Shi WJ, Zhou LS, Tao AL, Li L. Effect of epigallocatechin-3-gallate on the status of DNA methylation of E-cadherin promoter region on endometriosis mouse. J Obstet Gynaecol Res. 2020;46(10):2076–2083. doi:10.1111/jog.14358
160. Abu N, Ho WY, Yeap SK, et al. The flavokawains: uprising medicinal chalcones. Cancer Cell Int. 2013;13(1):102. doi:10.1186/1475-2867-13-102
161. Palko-łabuz A, Kostrzewa-Susłow E, Janeczko T, et al. Cyclization of flavokawain B reduces its activity against human colon cancer cells. Hum Exp Toxicol. 2020;39(3):262–275. doi:10.1177/0960327119882986
162. Kwon DJ, Ju SM, Youn GS, Choi SY, Park J. Suppression of iNOS and COX-2 expression by flavokawain A via blockade of NF-κB and AP-1 activation in RAW 264.7 macrophages. Food Chem Toxicol. 2013;58:479–486. doi:10.1016/j.fct.2013.05.031
163. Wei Z, Gu X, Zhang JR, et al. Beneficial biological effects of Flavokawain A, a chalcone constituent from kava, on surgically induced endometriosis rat model. J Ethnopharmacol. 2024;318(Pt A):116896. doi:10.1016/j.jep.2023.116896
164. Yu S, Nehus ZT, Badger TM, Fang N. Quantification of vitamin E and gamma-oryzanol components in rice germ and bran. J Agric Food Chem. 2007;55(18):7308–7313. doi:10.1021/jf071957p
165. Ma YR, Xiang ST, Jiang WW, et al. Gamma-oryzanol protects human liver cell (L02) from hydrogen peroxide-induced oxidative damage through regulation of the MAPK/Nrf2 signaling pathways. J Food Biochem. 2022;46(7):e14118. doi:10.1111/jfbc.14118
166. Bumrungpert A, Chongsuwat R, Phosat C, Butacnum A. Rice bran oil containing gamma-oryzanol improves lipid profiles and antioxidant status in hyperlipidemic subjects: a randomized double-blind controlled trial. J Altern Complement Med. 2019;25(3):353–358. doi:10.1089/acm.2018.0212
167. Xu H, Dai ZH, He GL, et al. Gamma-oryzanol alleviates intervertebral disc degeneration development by intercepting the IL-1β/NLRP3 inflammasome positive cycle. Phytomedicine. 2022;102:154176. doi:10.1016/j.phymed.2022.154176
168. Eisalou MY, Farahpour MR. Effectiveness of Gamma Oryzanol on prevention of surgical induced endometriosis development in rat model. Sci Rep. 2022;12(1):2816. doi:10.1038/s41598-022-06883-4
169. Shi BH, Ma MQ, Zheng YT, Pan YY, Lin XH. mTOR and Beclin1: two key autophagy-related molecules and their roles in myocardial ischemia/reperfusion injury. J Cell Physiol. 2019;234(8):12562–12568. doi:10.1002/jcp.28125
170. Houtman J, Freitag K, Gimber N, Schmoranzer J, Heppner FL, Jendrach M. Beclin1-driven autophagy modulates the inflammatory response of microglia via NLRP3. EMBO j. 2019;38:4. doi:10.15252/embj.201899430
171. Hou MQ, Wang RF, Zhao SJ, Wang ZT. Ginsenosides in Panax genus and their biosynthesis. Acta Pharm Sin B. 2021;11(7):1813–1834. doi:10.1016/j.apsb.2020.12.017
172. Han X, Song J, Lian LH, et al. Ginsenoside 25-OCH(3)-PPD promotes activity of LXRs To Ameliorate P2X7R-Mediated NLRP3 inflammasome in the development of hepatic fibrosis. J Agric Food Chem. 2018;66(27):7023–7035. doi:10.1021/acs.jafc.8b01982
173. Teng B, Jiang JG, Zhao LJ, et al. Ginsenoside PPD’s antitumor effect via down-regulation of mTOR revealed by super-resolution imaging. Molecules. 2017;22:3. doi:10.3390/molecules22030486
174. Glick D, Barth S, Macleod KF. Autophagy: cellular and molecular mechanisms. J Pathol. 2010;221(1):3–12. doi:10.1002/path.2697
175. Mei J, Zhu XY, Jin LP, Duan ZL, Li DJ, Li MQ. Estrogen promotes the survival of human secretory phase endometrial stromal cells via CXCL12/CXCR4 up-regulation-mediated autophagy inhibition. Hum Reprod. 2015;30(7):1677–1689. doi:10.1093/humrep/dev100
176. Zhang B, Zhou WJ, Gu CJ, et al. The ginsenoside PPD exerts anti-endometriosis effects by suppressing estrogen receptor-mediated inhibition of endometrial stromal cell autophagy and NK cell cytotoxicity. Cell Death Dis. 2018;9(5):574. doi:10.1038/s41419-018-0581-2
177. Tejada S, Pinya S, Martorell M, et al. Potential anti-inflammatory effects of hesperidin from the genus citrus. Curr Med Chem. 2018;25(37):4929–4945. doi:10.2174/0929867324666170718104412
178. Li C, Schluesener H. Health-promoting effects of the citrus flavanone hesperidin. Crit Rev Food Sci Nutr. 2017;57(3):613–631. doi:10.1080/10408398.2014.906382
179. Dong JR, Chang WW, Chen SM. Nerolidol inhibits proliferation of leiomyoma cells via reactive oxygen species-induced DNA damage and downregulation of the ATM/Akt pathway. Phytochemistry. 2021;191:112901. doi:10.1016/j.phytochem.2021.112901
180. de Souza E, Gomes M, Dos Santos Lima B, et al. Nerolidol-beta-cyclodextrin inclusion complex enhances anti-inflammatory activity in arthritis model and improves gastric protection. Life Sci. 2021;265:118742. doi:10.1016/j.lfs.2020.118742
181. Man MQ, Yang B, Elias PM. Benefits of hesperidin for cutaneous functions. Evid Based Complement Alternat Med. 2019;2019:2676307. doi:10.1155/2019/2676307
182. Ren H, Hao JL, Liu TT, et al. Hesperetin suppresses inflammatory responses in lipopolysaccharide-induced RAW 264.7 Cells via the inhibition of NF-κB and Activation of Nrf2/HO-1 Pathways. Inflammation. 2016;39(3):964–973. doi:10.1007/s10753-016-0311-9
183. Fonsêca DV, Salgado PR, de Carvalho FL, et al. Nerolidol exhibits antinociceptive and anti-inflammatory activity: involvement of the GABAergic system and proinflammatory cytokines. Fundam Clin Pharmacol. 2016;30(1):14–22. doi:10.1111/fcp.12166
184. Melekoglu R, Ciftci O, Eraslan S, Cetin A, Basak N. The beneficial effects of nerolidol and hesperidin on surgically induced endometriosis in a rat model*. Gynecol Endocrinol. 2018;34(11):975–980. doi:10.1080/09513590.2018.1469611
185. Zhao TT, Xu YQ, Hu HM, Gong HB, Zhu HL. Isoliquiritigenin (ISL) and its formulations: potential antitumor agents. Curr Med Chem. 2019;26(37):6786–6796. doi:10.2174/0929867325666181112091700
186. Shi DD, Yang JL, Jiang YM, Wen LG, Wang ZB, Yang B. The antioxidant activity and neuroprotective mechanism of isoliquiritigenin. Free Radic Biol Med. 2020;152:207–215. doi:10.1016/j.freeradbiomed.2020.03.016
187. Zeng J, Chen YZ, Ding R, et al. Isoliquiritigenin alleviates early brain injury after experimental intracerebral hemorrhage via suppressing ROS- and/or NF-κB-mediated NLRP3 inflammasome activation by promoting Nrf2 antioxidant pathway. J Neuroinflammation. 2017;14(1):119. doi:10.1186/s12974-017-0895-5
188. Liao Y, Tan RZ, Li JC, et al. Isoliquiritigenin attenuates UUO-induced renal inflammation and fibrosis by inhibiting Mincle/Syk/NF-Kappa B signaling pathway. Drug Des Devel Ther. 2020;14:1455–1468. doi:10.2147/DDDT.S243420
189. Peng F, Du QH, Peng C, et al. A review: the pharmacology of isoliquiritigenin. Phytother Res. 2015;29(7):969–977. doi:10.1002/ptr.5348
190. Yang J, Antin P, Berx G, et al. Guidelines and definitions for research on epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2020;21(6):341–352. doi:10.1038/s41580-020-0237-9
191. Loh CY, Chai JY, Tang TF, et al. The E-Cadherin and N-cadherin switch in epithelial-to-mesenchymal transition: signaling, therapeutic implications, and challenges. Cells. 2019;8:10.
192. Hsu YW, Chen HY, Chiang YF, Chang LC, Lin PH, Hsia SM. The effects of isoliquiritigenin on endometriosis in vivo and in vitro study. Phytomedicine. 2020;77:153214. doi:10.1016/j.phymed.2020.153214
193. Ren J, Lu YF, Qian YH, Chen BZ, Wu T, Ji G. Recent progress regarding kaempferol for the treatment of various diseases. Exp Ther Med. 2019;18(4):2759–2776. doi:10.3892/etm.2019.7886
194. Alam W, Khan H, Shah MA, Cauli O, Saso L. Kaempferol as a dietary anti-inflammatory agent: current therapeutic standing. Molecules. 2020;25:18. doi:10.3390/molecules25184073
195. Imran M, Salehi B, Sharifi-Rad J, et al. Kaempferol: a key emphasis to its anticancer potential. Molecules. 2019;24:12. doi:10.3390/molecules24122277
196. Liang F, Han YX, Gao H, et al. Kaempferol identified by zebrafish assay and fine fractionations strategy from dysosma versipellis inhibits angiogenesis through VEGF and FGF pathways. Sci Rep. 2015;5:14468. doi:10.1038/srep14468
197. Zhao JD, Wang JT, Liu JX, Li SL, Liu PF, Zhang XD. Effect and mechanisms of kaempferol against endometriosis based on network pharmacology and in vitro experiments. BMC Complement Med Ther. 2022;22(1):254. doi:10.1186/s12906-022-03729-4
198. Alzahrani AS. PI3K/Akt/mTOR inhibitors in cancer: at the bench and bedside. Semin Cancer Biol. 2019;59:125–132. doi:10.1016/j.semcancer.2019.07.009
199. Gendrisch F, Esser PR, Schempp CM, Wölfle U. Luteolin as a modulator of skin aging and inflammation. Biofactors. 2021;47(2):170–180. doi:10.1002/biof.1699
200. Imran M, Rauf A, Abu-Izneid T, et al. Luteolin, a flavonoid, as an anticancer agent: a review. Biomed Pharmacother. 2019;112:108612. doi:10.1016/j.biopha.2019.108612
201. Mende N, Kuchen EE, Lesche M, et al. CCND1-CDK4-mediated cell cycle progression provides a competitive advantage for human hematopoietic stem cells in vivo. J Exp Med. 2015;212(8):1171–1183. doi:10.1084/jem.20150308
202. Zhang X, Hu SJ, Zhang X, et al. MicroRNA-7 arrests cell cycle in G1 phase by directly targeting CCNE1 in human hepatocellular carcinoma cells. Biochem Biophys Res Commun. 2014;443(3):1078–1084. doi:10.1016/j.bbrc.2013.12.095
203. Liu DY, Liu AJ. Superoxide dismutase induces G1-phase cell cycle arrest by down-regulated expression of Cdk-2 and cyclin-E in murine sarcoma S180 tumor cells. Cell Biochem Funct. 2013;31(4):352–359. doi:10.1002/cbf.2912
204. Chen MJ, Cheng AC, Lee MF, Hsu YC. Simvastatin induces G(1) arrest by up-regulating GSK3β and down-regulating CDK4/cyclin D1 and CDK2/cyclin E1 in human primary colorectal cancer cells. J Cell Physiol. 2018;233(6):4618–4625. doi:10.1002/jcp.26156
205. Sherr CJ. D-type cyclins. Trends Biochem Sci. 1995;20(5):187–190. doi:10.1016/S0968-0004(00)89005-2
206. Park S, Lim W, You S, Song G. Ameliorative effects of luteolin against endometriosis progression in vitro and in vivo. J Nutr Biochem. 2019;67:161–172. doi:10.1016/j.jnutbio.2019.02.006
207. Ruiz-Vela A, Korsmeyer SJ. Proapoptotic histone H1.2 induces CASP-3 and −7 activation by forming a protein complex with CYT c, APAF-1 and CASP-9. FEBS Lett. 2007;581(18):3422–3428. doi:10.1016/j.febslet.2007.06.049
208. Song XMT, Tan L, Wang M, et al. A review of the most recent research. Biomed Pharmacother. 2021;134:111017. doi:10.1016/j.biopha.2020.111017
209. Gupta G, Siddiqui MA, Khan MM, et al. Current pharmacological trends on myricetin. Drug Res. 2020;70(10):448–454. doi:10.1055/a-1224-3625
210. Park S, Song G, Lim W. Myricetin inhibits endometriosis growth through cyclin E1 down-regulation in vitro and in vivo. J Nutr Biochem. 2020;78:108328. doi:10.1016/j.jnutbio.2019.108328
211. Mir IA, Tiku AB. Chemopreventive and therapeutic potential of ”naringenin”, a flavanone present in citrus fruits. Nutr Cancer. 2015;67(1):27–42. doi:10.1080/01635581.2015.976320
212. Marcellin L, Santulli P, Chouzenoux S, et al. Alteration of Nrf2 and Glutamate Cysteine Ligase expression contribute to lesions growth and fibrogenesis in ectopic endometriosis. Free Radic Biol Med. 2017;110:1–10. doi:10.1016/j.freeradbiomed.2017.04.362
213. Iwabuchi T, Yoshimoto C, Shigetomi H, Kobayashi H. Oxidative stress and antioxidant defense in endometriosis and its malignant transformation. Oxid Med Cell Longev. 2015;2015:848595. doi:10.1155/2015/848595
214. Kapoor R, Sirohi VK, Gupta K, Dwivedi A. Naringenin ameliorates progression of endometriosis by modulating Nrf2/Keap1/HO1 axis and inducing apoptosis in rats. J Nutr Biochem. 2019;70:215–226. doi:10.1016/j.jnutbio.2019.05.003
215. Park S, Lim W, Bazer FW, Song G. Naringenin induces mitochondria-mediated apoptosis and endoplasmic reticulum stress by regulating MAPK and AKT signal transduction pathways in endometriosis cells. Mol Hum Reprod. 2017;23(12):842–854. doi:10.1093/molehr/gax057
216. Li S, Yu H, Ho CT. Nobiletin: efficient and large quantity isolation from Orange peel extract. Biomed Chromatogr. 2006;20(1):133–138. doi:10.1002/bmc.540
217. Han SH, Han JH, Chun WJ, Lee SS, Kim HS, Lee JW. Nobiletin inhibits non-small-cell lung cancer by inactivating WNT/β-catenin signaling through downregulating miR-15-5p. Evid Based Complement Alternat Med. 2021;2021:7782963. doi:10.1155/2021/7782963
218. Güvenç M, Cellat M, Gökçek İ, et al. Nobiletin attenuates acetaminophen-induced hepatorenal toxicity in rats. J Biochem Mol Toxicol. 2020;34(2):e22427. doi:10.1002/jbt.22427
219. Cajas YN, Cañón-Beltrán K, Ladrón de Guevara M, et al. Antioxidant nobiletin enhances oocyte maturation and subsequent embryo development and quality. Int J Mol Sci. 2020;21:15. doi:10.3390/ijms21155340
220. Hayden MS, Ghosh S. Shared principles in NF-kappaB signaling. Cell. 2008;132(3):344–362. doi:10.1016/j.cell.2008.01.020
221. Oeckinghaus A, Ghosh S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb Perspect Biol. 2009;1(4):a000034. doi:10.1101/cshperspect.a000034
222. Wei X, Shao X. Nobiletin alleviates endometriosis via down-regulating NF-κB activity in endometriosis mouse model. Biosci Rep. 2018;38(3):3. doi:10.1042/BSR20180470
223. Gorzynik-Debicka M, Przychodzen P, Cappello F, et al. Potential health benefits of olive oil and plant polyphenols. Int J Mol Sci. 2018;19:3. doi:10.3390/ijms19030686
224. Park Y, Cho YJ, Sung N, et al. Oleuropein suppresses endometriosis progression and improves the fertility of mice with endometriosis. J Biomed Sci. 2022;29(1):100. doi:10.1186/s12929-022-00883-2
225. Anand David AV, Arulmoli R, Parasuraman S. Overviews of biological importance of quercetin: a bioactive flavonoid. Pharmacogn Rev. 2016;10(20):84–89. doi:10.4103/0973-7847.194044
226. Rauf A, Imran M, Khan IA, et al. Anticancer potential of quercetin: a comprehensive review. Phytother Res. 2018;32(11):2109–2130. doi:10.1002/ptr.6155
227. Almeida AF, Borge GIA, Piskula M, et al. Bioavailability of quercetin in humans with a focus on interindividual variation. Compr Rev Food Sci Food Saf. 2018;17(3):714–731. doi:10.1111/1541-4337.12342
228. Ping SM, Ma CB, Liu P, et al. Molecular mechanisms underlying endometriosis pathogenesis revealed by bioinformatics analysis of microarray data. Arch Gynecol Obstet. 2016;293(4):797–804. doi:10.1007/s00404-015-3875-y
229. Qie S, Diehl JA. Cyclin D1, cancer progression, and opportunities in cancer treatment. J Mol Med. 2016;94(12):1313–1326. doi:10.1007/s00109-016-1475-3
230. Jamali N, Zal F, Mostafavi-Pour Z, Samare-Najaf M, Poordast T, Dehghanian A. Ameliorative effects of quercetin and metformin and their combination against experimental endometriosis in rats. Reprod Sci. 2021;28(3):683–692. doi:10.1007/s43032-020-00377-2
231. Kim YC, Guan KL. mTOR: a pharmacologic target for autophagy regulation. J Clin Invest. 2015;125(1):25–32. doi:10.1172/JCI73939
232. Galiniak S, Aebisher D, Bartusik-Aebisher D. Health benefits of resveratrol administration. Acta Biochim Pol. 2019;66(1):13–21. doi:10.18388/abp.2018_2749
233. Wu H, Chen L, Zhu FF, Han X, Sun LD, Chen KP. The cytotoxicity effect of resveratrol: cell cycle arrest and induced apoptosis of breast cancer 4T1 cells. Toxins. 2019;11:12. doi:10.3390/toxins11120731
234. Popescu M, Bogdan C, Pintea A, Rugină D, Ionescu C. Antiangiogenic cytokines as potential new therapeutic targets for resveratrol in diabetic retinopathy. Drug Des Devel Ther. 2018;12:1985–1996. doi:10.2147/DDDT.S156941
235. Meng TT, Xiao DF, Muhammed A, Deng JY, Chen L, He JH. Anti-inflammatory action and mechanisms of resveratrol. Molecules. 2021;26:1. doi:10.3390/molecules26010229
236. Zhou DD, Luo M, Huang SY, et al. Effects and mechanisms of resveratrol on aging and age-related diseases. Oxid Med Cell Longev. 2021;2021:9932218. doi:10.1155/2021/9932218
237. Rauf A, Imran M, Butt MS, Nadeem M, Peters DG, Mubarak MS. Resveratrol as an anti-cancer agent: a review. Crit Rev Food Sci Nutr. 2018;58(9):1428–1447. doi:10.1080/10408398.2016.1263597
238. Hseu YC, Yang TY, Li ML, et al. Chalcone flavokawain A attenuates TGF-β1-induced fibrotic pathology via inhibition of ROS/Smad3 signaling pathways and induction of Nrf2/ARE-mediated antioxidant genes in vascular smooth muscle cells. J Cell Mol Med. 2019;23(2):775–788. doi:10.1111/jcmm.13973
239. Khodarahmian M, Amidi F, Moini A, et al. A randomized exploratory trial to assess the effects of resveratrol on VEGF and TNF-α 2 expression in endometriosis women. J Reprod Immunol. 2021;143:103248. doi:10.1016/j.jri.2020.103248
240. Madanes D, Meresman G, Valla SA, et al. Resveratrol impairs cellular mechanisms associated with the pathogenesis of endometriosis. Reprod Biomed Online. 2022;44(6):976–990. doi:10.1016/j.rbmo.2022.02.008
241. Wang LP, Ma Q. Clinical benefits and pharmacology of scutellarin: a comprehensive review. Pharmacol Ther. 2018;190:105–127. doi:10.1016/j.pharmthera.2018.05.006
242. Han J, Wang P, Xia X, et al. EGR1 promoted anticancer effects of Scutellarin via regulating LINC00857/miR-150-5p/c-Myc in osteosarcoma. J Cell Mol Med. 2021;25(17):8479–8489. doi:10.1111/jcmm.16809
243. Mo J, Yang RH, Li F, et al. Scutellarin protects against vascular endothelial dysfunction and prevents atherosclerosis via antioxidation. Phytomedicine. 2018;42:66–74. doi:10.1016/j.phymed.2018.03.021
244. Zeng S, Chen L, Sun Q, et al. Scutellarin ameliorates colitis-associated colorectal cancer by suppressing Wnt/β-catenin signaling cascade. Eur J Pharmacol. 2021;906:174253. doi:10.1016/j.ejphar.2021.174253
245. Lv WL, Liu Q, An JH, Song XY. Scutellarin inhibits hypoxia-induced epithelial-mesenchymal transition in bladder cancer cells. J Cell Physiol. 2019;234(12):23169–23175. doi:10.1002/jcp.28883
246. Long LL, Li YB, Yu S, et al. Scutellarin prevents angiogenesis in diabetic retinopathy by downregulating VEGF/ERK/FAK/Src pathway signaling. J Diabetes Res. 2019;2019:4875421. doi:10.1155/2019/4875421
247. Peng L, Wen L, Shi QF, et al. Scutellarin ameliorates pulmonary fibrosis through inhibiting NF-κB/NLRP3-mediated epithelial-mesenchymal transition and inflammation. Cell Death Dis. 2020;11(11):978. doi:10.1038/s41419-020-03178-2
248. Barrientos S, Stojadinovic O, Golinko MS, Brem H, Tomic-Canic M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008;16(5):585–601. doi:10.1111/j.1524-475X.2008.00410.x
249. Zhang Q, Duan J, Liu X, Guo SW. Platelets drive smooth muscle metaplasia and fibrogenesis in endometriosis through epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation. Mol Cell Endocrinol. 2016;428:1–16. doi:10.1016/j.mce.2016.03.015
250. Ding D, Cai X, Zheng H, Guo SW, Liu X. Scutellarin suppresses platelet aggregation and stalls lesional progression in mouse with induced endometriosis. Reprod Sci. 2019;26(11):1417–1428. doi:10.1177/1933719118817661
251. Abenavoli L, Capasso R, Milic N, Capasso F. Milk thistle in liver diseases: past, present, future. Phytother Res. 2010;24(10):1423–1432. doi:10.1002/ptr.3207
252. Jouhari S, Mohammadzadeh A, Soltanghoraee H, et al. Effects of silymarin, cabergoline and letrozole on rat model of endometriosis. Taiwan J Obstet Gynecol. 2018;57(6):830–835. doi:10.1016/j.tjog.2018.10.011
253. Wei PS, Li X, Wang S, et al. Silibinin Ameliorates Formaldehyde-Induced Cognitive Impairment by Inhibiting Oxidative Stress. Oxid Med Cell Longev. 2022;2022:5981353. doi:10.1155/2022/5981353
254. Si L, Fu J, Liu W, et al. Silibinin inhibits migration and invasion of breast cancer MDA-MB-231 cells through induction of mitochondrial fusion. Mol Cell Biochem. 2020;463(1–2):189–201. doi:10.1007/s11010-019-03640-6
255. Verdura S, Cuyàs E, Ruiz-Torres V, et al. Lung Cancer Management with Silibinin: a Historical and Translational Perspective. Pharmaceuticals. 2021;14:6. doi:10.3390/ph14060559
256. Trappoliere M, Caligiuri A, Schmid M, et al. Silybin, a component of sylimarin, exerts anti-inflammatory and anti-fibrogenic effects on human hepatic stellate cells. J Hepatol. 2009;50(6):1102–1111. doi:10.1016/j.jhep.2009.02.023
257. Mirzaei N, Jahanian Sadatmahalleh S, Rouholamin S, Nasiri M. A randomized trial assessing the efficacy of Silymarin on endometrioma-related manifestations. Sci Rep. 2022;12(1):17549. doi:10.1038/s41598-022-22073-8
258. Ham J, Kim J, Bazer FW, Lim W, Song G. Silibinin-induced endoplasmic reticulum stress and mitochondrial dysfunction suppress growth of endometriotic lesions. J Cell Physiol. 2019;234(4):4327–4341. doi:10.1002/jcp.27212
259. Jiang RD, Xu JQ, Zhang YZ, Zhu XM, Liu JC, Tan YR. Ligustrazine alleviate acute lung injury through suppressing pyroptosis and apoptosis of alveolar macrophages. Front Pharmacol. 2021;12:680512. doi:10.3389/fphar.2021.680512
260. Sheu JR, Kan YC, Hung WC, Ko WC, Yen MH. Mechanisms involved in the antiplatelet activity of tetramethylpyrazine in human platelets. Thromb Res. 1997;88(3):259–270. doi:10.1016/S0049-3848(97)00253-3
261. Yang SJ, Wu SD, Dai WL, et al. Tetramethylpyrazine: a review of its antitumor potential and mechanisms. Front Pharmacol. 2021;12:764331. doi:10.3389/fphar.2021.764331
262. Zhu XL, Shan Y, Yu MS, et al. Tetramethylpyrazine ameliorates peritoneal angiogenesis by regulating VEGF/Hippo/YAP signaling. Front Pharmacol. 2021;12:649581. doi:10.3389/fphar.2021.649581
263. Chen JC, Tian JJ, Ge HF, Liu RH, Xiao JB. Effects of tetramethylpyrazine from Chinese black vinegar on antioxidant and hypolipidemia activities in HepG2 cells. Food Chem Toxicol. 2017;109(Pt 2):930–940. doi:10.1016/j.fct.2016.12.017
264. Brabletz S, Schuhwerk H, Brabletz T, Stemmler M. Dynamic EMT: a multi-tool for tumor progression. EMBO J. 2021;40(18):e108647. doi:10.15252/embj.2021108647
265. Zhou WJ, Yang HL, Shao J, et al. Anti-inflammatory cytokines in endometriosis. Cell Mol Life Sci. 2019;76(11):2111–2132. doi:10.1007/s00018-019-03056-x
266. Pardali E, Sanchez-Duffhues G, Gomez-Puerto M, Ten Dijke P. TGF-β-induced endothelial-mesenchymal transition in fibrotic diseases. Int J Mol Sci. 2017;18:10. doi:10.3390/ijms18102157
267. Biernacka A, Dobaczewski M, Frangogiannis NG. TGF-β signaling in fibrosis. Growth Factors. 2011;29(5):196–202. doi:10.3109/08977194.2011.595714
268. Huang SH, Xiao FY, Guo SW, Zhang TT. Tetramethylpyrazine retards the progression and fibrogenesis of endometriosis. Reprod Sci. 2022;29(4):1170–1187. doi:10.1007/s43032-021-00813-x
269. Khwaza V, Oyedeji OO, Aderibigbe BA. Ursolic acid-based derivatives as potential anti-cancer agents: an update. Int J Mol Sci. 2020;21(16):5920. doi:10.3390/ijms21165920
270. Yin R, Li T, Tian JX, Xi P, Liu RH. Ursolic acid, a potential anticancer compound for breast cancer therapy. Crit Rev Food Sci Nutr. 2018;58(4):568–574. doi:10.1080/10408398.2016.1203755
271. Li JJ, Zeng Z, Chang YJ, et al. Suppressive effects of ursolic acid on human endometriotic stromal cells survival. Gynecol Obstet Invest. 2020;85(1):72–81. doi:10.1159/000502258
272. Ge JL, Yang HH, Zeng YF, Liu YJ. Protective effects of wogonin on lipopolysaccharide-induced inflammation and apoptosis of lung epithelial cells and its possible mechanisms. Biomed Eng Online. 2021;20(1):125. doi:10.1186/s12938-021-00965-6
273. Sharifi-Rad J, Herrera-Bravo J, Salazar L, et al. The therapeutic potential of wogonin observed in preclinical studies. Evidence-Bas Complem Alternat Med. 2021;2021:9935451. doi:10.1155/2021/9935451
274. Khan N, Haseeb A, Ansari M, Devarapalli P, Haynie S, Haqqi T. Wogonin, a plant derived small molecule, exerts potent anti-inflammatory and chondroprotective effects through the activation of ROS/ERK/Nrf2 signaling pathways in human Osteoarthritis chondrocytes. Free Radical Bio Med. 2017;106:288–301. doi:10.1016/j.freeradbiomed.2017.02.041
275. Jiang RB, Jin B, Wan DY, Zhu CH, Feng JG, Gu LH. Therapy effects of wogonin on ovarian cancer cells. Biomed Res Int. 2017;2017:9381513. doi:10.1155/2017/9381513
276. Vercellini P, Viganò P, Somigliana E, Fedele L. Endometriosis: pathogenesis and treatment. Nat Rev Endocrinol. 2014;10(5):261–275. doi:10.1038/nrendo.2013.255
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.