Back to Journals » Journal of Pain Research » Volume 14
The Spinal α7-Nicotinic Acetylcholine Receptor Contributes to the Maintenance of Cancer-Induced Bone Pain
Authors Yang T, Zhou Y, Zhang W, Zhang L, Chen S, Chen C, Gao F, Yang H, Manyande A, Wang J, Tian Y, Tian X
Received 12 October 2020
Accepted for publication 23 December 2020
Published 15 February 2021 Volume 2021:14 Pages 441—452
DOI https://doi.org/10.2147/JPR.S286321
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Robert B. Raffa
Ting Yang1 1, Yaqun Zhou1 1, Wen Zhang1 1, Longqing Zhang1 1, Shuping Chen1 1, Chao Chen1 1, Feng Gao1 1, Hui Yang1 1, Anne Manyande2 2, Jie Wang3 3, Yuke Tian1 1, Xuebi Tian1 1
1Department of Anesthesiology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, People’s Republic of China; 2School of Human and Social Sciences, University of West London, London, UK; 3State Key Laboratory of Magnetic Resonance and Atomic and Molecular Physics, Key Laboratory of Magnetic Resonance in Biological Systems, Wuhan Center for Magnetic Resonance, Wuhan Institute of Physics and Mathematics, Chinese Academy of Sciences, Wuhan, Hubei, People’s Republic of China
Correspondence: Xuebi Tian
Department of Anesthesiology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Jiefang Avenue 1095#, Wuhan, Hubei, 430030, People’s Republic of China
Tel +862783663423
Fax +862783662853
Email [email protected]
Introduction: Cancer-induced bone pain (CIBP) is acknowledged as a multifactorial chronic pain that tortures advanced cancer patients, but existing treatment strategies for CIBP have not been satisfactory yet. Investigators have demonstrated that the activation of α 7-nAChRs exerts analgesic effects in some chronic pain models. However, the role of spinal α 7-nAChRs in CIBP remains unknown. This study was designed to investigate the role of α 7-nAChRs in a well-established CIBP model induced by Walker 256 rat mammary gland carcinoma cells.
Methods: The paw withdrawal threshold (PWT) of the ipsilateral hind paw was measured using von Frey filament. The expressions of spinal α 7-nAChRs and NF-κB were measured with Western blotting analysis. Immunofluorescence was employed to detect the expression of α 7-nAChRs and co-expressed of α 7-nAChRs with NeuN or GFAP or Iba1.
Results: Experiment results showed that the expression of spinal α 7-nAChRs was significantly downregulated over time in CIBP rats, and in both CIBP rats and sham rats, most of the α 7-nAChRs located in neurons. Behavioral data suggested PNU-282,987, a selective α 7-nAChRs agonist, dose-dependently produced analgesic effect and positive allosteric modulator could intensify its effects. Further, repeated administration of PNU-282,987 reversed the expression of α 7-nAChRs, inhibited the nuclear factor kappa B (NF-κB) signaling pathway, and attenuates CIBP-induced mechanical allodynia state as well.
Conclusion: These results suggest that the reduced expression of spinal α 7-nAChRs contributes to the maintenance of CIBP by upregulating NF-κB expression, which implying a novel pharmacological therapeutic target for the treatment of CIBP.
Keywords: cancer-induced bone pain, α 7-nAChR, NF-κB, PNU-282,987
Introduction
Cancer-induced bone pain (CIBP) is usually the trickiest for clinicians for its complicated mechanisms, which involved immune cells, nerves and mechanosensitive receptors.1–3 Statistically, more than 70% of cancer patients suffer pain4 in which CIBP is the most common type of pain, and their quality of life is reduced markedly because of poor mood, the ability to walk5 and sleep.5–7 The World Health Organization (WHO) guidelines recommend a stepladder approach using non-opioids, weak and then strong opioids for the relief to CIBP,8 but the problem is still inadequately addressed, and there remains a great challenge to effectively alleviate the patients’ pain because of limited effects and/or intolerable side effects.9,10 Given that the existing treatment for CIBP remains deficient according to its known mechanisms, the specific cellular and molecular mechanisms underlying CIBP remain a pressing need to do more to understand.
Nicotine acetylcholine receptors (nAChRs) exist extensively in the central and peripheral nervous system as well as on immune cells, having been explored as a pain relief strategy in the past three decades. Alpha7-nAChR, one of the nicotinic acetylcholine receptors, is composed of five α7 subunits per receptor, possessing the characteristic of high calcium permeability and rapid desensitization in response to the stimulation from high concentration agonist11,12 compared with other nAChRs subtypes. They are reported to be beneficial for retinal diseases,13 psychiatric disorders,14 and some central nervous system disorders.15 The α7-nAChR has been shown to be distributed in the pain transmission pathway and evidences are accumulated that α7-nAChR activation has been related to analgesic effects in some animal models.16–21 However, so far, the role of spinal α7-nAChR in CIBP has not been studied yet. Nuclear factor kappa-B (NF-κB) is a heterodimer composed of p65 and p50 or p52 subunits. NF-κB is thought to control many genes related to nociceptive mediators.22,23 Accumulating evidence indicates that activation of NF-κB enhanced pain perception, while inhibition of NF-κB reduced pain, including cancer-induced bone pain.24–26 Lipopolysaccharide activating α7-nAChR blocked NF-κB nuclear translocation.27 Here in this study, we tested whether α7-nAChR and NF-κB signaling pathways were involved in the maintenance of CIBP and its possible mechanisms.
Materials and Methods
Animals and Ethical Statement
One hundred and forty-three virgin female Sprague–Dawley rats (200–220 g, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, China) that are highly susceptible to Walker 256 rat mammary gland carcinoma cells (from our laboratory storage and approved by the research ethics committee of Tongji hospital) were seed in this study. Sex selection for building CIBP model depends on tumor tissue of origin and epidemiological characteristics of relative cancer between men and women.28 The rats were housed in groups of one to three per cage under controlled conditions (24 ± 0.5°C, relative humidity was about 55%, 12 h alternating light-dark cycle) and given food and water ad libitum. All experimental protocols were acquired approval from the Institutional Animal Care and Use Committee of Huazhong University of Science and Technology and conform to guidelines of the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Preparation of Carcinoma Cells
Walker 256 rat mammary gland carcinoma cells were taken out from the liquid nitrogen tank, quickly recovered in a 37°C water bath, and then diluted to 4 × 107 cells/mL. For obtaining more tumor cells quickly, the diluted cells (4 × 107 cells/mL, 1 mL) were inoculated into the abdominal cavity of female SD rats. About 5 to 7 days later, carcinoma cells were harvested from the peritoneal fluid. Then, cells were diluted to 4×107cells/mL with sterile phosphate buffer saline (PBS) and placed on ice for tumor cells implantation (TCI).
Establishment of Tibial CIBP Models
CIBP model was performed as described previously.29,30 First of all, under deep anesthesia with sodium pentobarbital (50 mg/kg, i.p.), the skin of the operation area was disinfected and a small incision was made to expose the proximal right tibia. And then, a hole was drilled on the tibia and Walker 256 cells (4 × 107 cells/mL, 10μL) harvesting from the peritoneal fluid were slowly injected into the tibial hole using a 10 μL Hamilton syringe. The sham group was applied the same operation, but injectedwith10 μL PBS instead. The syringe was then detained for at least 2 minutes to ensure adequate diffusion of carcinoma cells in the bone cavity. After that, the hole was sealed with bone wax as soon as the syringe was gotten out. Last of all, the wound was stitched with 3–0 silk thread. Animals were kept warm on a heated pad until recovering from anesthesia. Rats that successfully constructed a cancer-induced pain model not only had changes in the paw withdrawal threshold but also the bone destruction at the tumor injection site; however, the injury caused by NS injection of sham rats healed several days later.
Drug Administration
We modified the previous intrathecal catheterization (i.t.) methods31,32 and performed it 7 days before establishing CIBP models. In simple terms, the rats were anesthetized using sodium pentobarbital (50 mg/kg, i.p.), and then a PE-10 polyethylene catheter (PE-10 tube) was implanted into the subarachnoid space from the spinal cord intervertebral space between L5 and L6. If there were no signs of a tail-flick response or cerebrospinal fluid in the catheter when inserting the catheter and a sudden paralysis of both legs after an i.t. injection of 2% lidocaine (10μL), the rats were eliminated. Alpha7-nAChR agonist PNU-282,987 and α7-nAChR antagonist Methyllycaconitine citrate (MLA) were purchased from Med Chem Express (Monmouth Junction, NJ, USA), and α7-nAChR positive allosteric modulator (PAM) PNU-120,596 were purchased from Sigma (Sigma-Aldrich, St. Louis, USA). The first two drugs were dissolved in 0.9% saline and the last one was dissolved in 5% DMSO. The dose of drugs was determined according to previous reports.20,33,34 The specific protocol was as follows: for acute treatment, a single dose of PNU-282,987 (0.5, 0.25, 0.1 mg/kg, i.t.) was given on day14 after the TCI; for chronic treatment, PNU-282,987 was given from day 14 to day 18 once daily. MLA (10μg/10μL, i.t.) or PNU-120,596 (4μg/10μL, i.t.) was given 30 min before PNU-282,987 (0.5mg/kg, i.t.) injection.
Behavioral Tests
In order to evaluate mechanical allodynia, we stimulated the rats regarding the paw withdrawal threshold (PWT) of the ipsilateral hind paw using von Frey filament as described previously.35 In short, each rat was placed in a single transparent chamber on a metal mesh floor and acclimated to the environment for 20–30 min before behavioral testing. The testing was started at a 2 g filament. A positive response was defined as an abrupt withdrawal, licking, and shaking of the tested paw. Once a positive response happened, the von Frey filament was replaced with a next lower one; otherwise, the next higher one was applied. Finally, the lowest numerical value of von Frey filament eliciting a positive response was recorded as the PWT. All behavioral tests on animals were carried out by a blinded investigator.
Immunofluorescence
Under appropriate anesthetic depth with sodium pentobarbital (50 mg/kg, i.p.), each animal was accepted perfusion transcardially with about 250 mL 4% icy paraformaldehyde (PFA) after 250 mL PBS. The next steps were to harvest L4 to L5 spinal segments, post-fix these samples in 4% PFA, and dehydrate them in 30% sucrose solution overnight at 4°C. After that the tissues were embedded in O.C.T. and sectioned 20 μm thick in a cryostat (CM1900, Leica, Wetzlar, Germany), last of all, keep the sections at −20°C.
Single Immunostainings
The spinal sections were incubated with 3‰ Triton X-100 for 15 min and blocked with 5% donkey serum for 1 hour, and then placed at 4°C for a night with rabbit anti-α7 nAChR antibody (1:50, 21,379-1-AP, Proteintech Group, Wuhan, China). The sections were incubated with IFKine Green Affinipure Donkey Anti-Rabbit IgG (1:100, A24221, Abbkine) for 2 hours at 37°C and required 5 washes (7 minutes each) with PBST. Sections were drained excess PBST and covered the coverslips after adding 50% glycerol. Photomicrographs were captured by a fluorescence microscope (DM2500, Leica, Wetzlar, Germany).
Double-Labeling Procedures
Spinal sections were incubated with a mixture of rabbit anti-α7 nAChR antibody (1:50, 21,379-1-AP, Proteintech Group, Wuhan, China), and mouse anti-neuronal nuclei (NeuN) antibody (1:200; MAB377; EMD Millipore, Billerica, MA; neuronal marker), mouse anti-glial fibrillary acidic protein (GFAP) antibody (1:200; 3670; Cell Signaling Technology, Danvers, MA; astrocytic marker), or goat anti-ionized calcium-binding adapter molecule 1 (Iba1) antibody (1:200; ab5076; Abcam; microglial marker), respectively. The next day, slices were washed with PBST 5 times, then incubated with a mixture of IFKine Green Affinipure Donkey Anti-Rabbit IgG (1:100, A24221, Abbkine) and IFKine Green Affinipure Donkey Anti-Mouse IgG (1:200, A24411, Abbkine) or IFKine Green Affinipure Donkey Anti-Goat IgG (1:200, A24431, Abbkine) for 2 hours at room temperature. The other steps were the same as the single immunostainings described above.
The number of α7-nAChRs and double-labeled α7-nAChRs/NeuN or Iba 1 or GFAP-positive cells in the dorsal horn (laminae I–IV) was counted by Software Image-Pro Plus Version 6.0 for Windows™ (MediaCybernetics, Bethesda, USA). Three slices were randomly selected from each rat for counting. The average value of each rat was used as five independent values for further statistical analysis. Then, we calculated the percentage of positive cells co-expressed with NeuN or GFAP or Iba1 in the total positive cells.
Western Blot Analysis
Western blot analysis was performed to quantify the expression of aimed protein as described previously.36 Under appropriate anesthesia with sodium pentobarbital (50 mg/kg, i.p.), the lumbar enlargements were obtained and homogenized in radio-immunoprecipitation assay (RIPA) lysis buffer containing inhibitor cocktail. The supernatant was kept following centrifugation (16,000×g, 4°C, 15 min, 2 times) and the protein concentration was determined using the Bradford method and standardized. The protein samples were heated for 15 min at 100°C with sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) buffer. About 40 μg aimed proteins were separated by 10% sodium dodecyl sulfate-PAGE electrophoresis and then transferred to polyvinylidene fluoride (PVDF) membranes (IPVH00010, EMD Millipore). The membranes were blocked with 5% skim milk in Tris-buffered saline and Tween 20 (TBST, 0.1%) for 2 h at room temperature, and subsequently incubated overnight at 4°C with anti-α7 nAChR antibody (1:1000, 21,379-1-AP, Proteintech Group, Wuhan, China) or p65 (1:1000, 10,745-1-AP, Proteintech Group, Wuhan, China) and rabbit anti-GAPDH antibody (1:5000; AS1039; Aspen), respectively. The protein bands were washed in TBST for 4 times (8 minutes each) and incubated with HRP goat anti-rabbit (1:5000; A21020; Abbkine) for 2 h at 37°C. After that, the protein bands were emerged using Super Lumia ECL Plus HRP Substrate Kit (K22030; Abbkine). Finally, we calculated the mean intensities of selected areas and normalized them to values of GAPDH by Image Lab software (BIO-RAD, Chemi DocXRS1, USA).
Statistical Analysis
All results are presented as the mean standard error of the mean (SEM) and performed by GraphPad Prism version 8. PWTs were analyzed by one-way ANOVA followed by Bonferroni post hoc test; Two-way analysis of variance with repeated measures, followed by Bonferroni post hoc test was used for Western blot, immunochemistry data. NF-κB Western blot data before drugs administration were analyzed by unpaired t-test; p < 0.05 was considered statistically significant.
Results
Expression and Cell Localization of α7-nAChRs in Spinal Cord of CIBP Rats
The experimental design is shown in Figure 1A. Firstly, we tested the expression of α7-nAChRs on cancer-induced bone pain (CIBP) rats. As expected, mechanical allodynia was established on Walker 256 tumor cells implanted rats (d7: F (2,15) =40.76, p<0.0001; t=6.774, p<0.0001; d14:F (2,15) =40.85, p<0.0001; t=7.46, p<0.0001;d21: F (2,15) =55.02, p<0.0001; t=8.327, p<0.0001) (Figure 1B). And then the spinal expression of α7-nAChRs was tested by western-blot. Compared with sham rats, the expression of α7-nAChRs was significantly decreased on day14 and day21 after tumor cells implantation (F(4,20)=10.23, p=0.0001; t14=3.86, p=0.0098; t21=5.099p=0.0005) (Figure 1C and D). To figure out cellular localization of α7-nAChRs in lumbar spinal cord dorsal horn, we used double stain of anti-α7-nAChRs and anti-neuron marker (NeuN), anti-astrocyte marker (GFAP), and anti-microglia marker (Iba1). As presented in Figure 2A and B, α7-nAChRs were predominantly colocalized with NeuN, only a few with GFAP and Iba1 in the spinal cord, both in CIBP and sham rats. With the results tallied, it was about 82.5% α7-nAChRs in sham rats and 75.2% α7-nAChRs in CIBP rats were expressed in neurons, and less than 5% of α7-nAChRs were expressed on activated microglia and activated astrocytes, the rest might be expressed on cells such as oligodendrocyte, immune cells, inactive microglia and inactive astrocytes (Figure 2C and D).
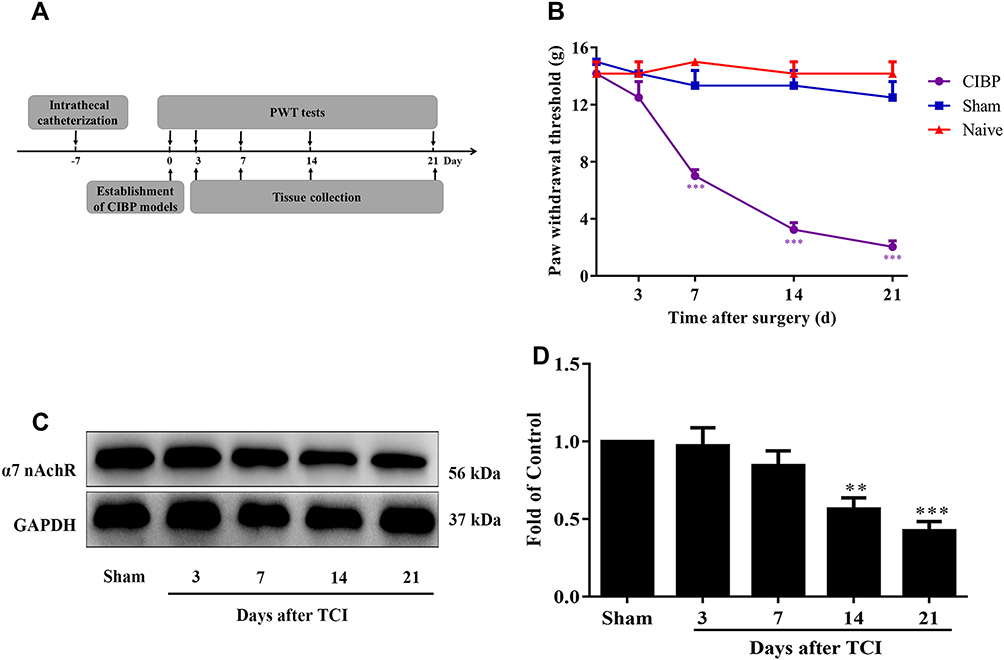 |
Figure 1 Spinal α7-nAChRs was downregulated in CIBP rats. (A) Schematic diagram of the experimental procedure. (B) The ipsilateral PWTs in response to von Frey filaments decreased from day 7 to day 21 in CIBP rats. In contrast, the changes of PWTs in naive and sham rats were not observed in the 21days (***p< 0.001 versus the naive group, n = 6 per group). (C, D) Western blot analysis indicated α7-nAChRs protein level was gradually downregulated in CIBP rats (**P <0.01, ***P < 0.001 versus the sham group, n = 5 per group). Abbreviations: α7-nAChR, alpha-7 nicotinic acetylcholine receptor; CIBP, cancer-induced bone pain; PWTs, paw withdrawal thresholds; TCI, tumor cells implantation. |
 |
Figure 2 Expression and cellular localization of α7-nAChRs in spinal cord dorsal horn. (A, B) Photomicrographs of α7-nAChRs (green) double fluorescence labeling with neuronal marker NeuN (red), astrocyte marker GFAP (red), and microglial marker Iba1 (red) in the spinal cord at day 21 after TCI (n = 5 per group). Our results revealed that α7-nAChRs were co-expressed predominantly with NeuN, only a few with GFAP and Iba1 in the spinal cord, both in CIBP and normal rats. In addition, immunofluorescence data showed that the immunoreactivity of α7-nAChRs was decreased in the spinal cord dorsal horn on day 21 after TCI. (C, D) It was about 82.5% α7-nAChRs in sham rats and 75.2% α7-nAChRs in CIBP rats were expressed in neurons, and less than 5% of α7-nAChRs were expressed on activated microglia and activated astrocytes. Abbreviations: α7-nAChR, alpha-7 nicotinic acetylcholine receptor; CIBP, cancer-induced bone pain; PWTs, paw withdrawal thresholds; TCI, tumor cells implantation. |
Pharmacological Activation of α7-nAChRs Produced Dose-Dependent Analgesic Effects in CIBP Rats
The experimental design is shown in Figure 3A. To explore the analgesic effect of spinal α7-nAChRs in CIBP rats by pharmacological methods, firstly, a single dose of α7-nAChR agonist, PNU-282,987 was intrathecally administrated on day14 after TCI. And the behavioral tests were performed at 0.5, 1, 1.5, 2, 2.5 and 3 h after injection. When compared with the normal saline given rats, 0.5 and 0.25 mg/kg PNU-282,987 treatment notably relieved cancer-induced bone pain, lasting from 0.5 h(F (4, 17) = 12.71, P<0.0001; t0.5mg/kg=5.801, p=0.0002; t0.25mg/kg= 4.687, p= 0.0021) to 2.5 h (F (4, 17) = 612.71, P = 0.0021; t0.5mg/kg=4.171, p=0.0064; t0.25mg/kg = 3.436, p = 0.0315) (Figure 3B). However, compared with that of the NS group, treatment of 0.1 mg/kg PNU-282,987 had no obvious influence on PWT. When α7-nAChR antagonist, the MLA (i.t.10μg/10μL) was administrated 30 min before PNU-282,987 (i.t., 0.5mg/kg) injection, the analgesic effect of PNU-282,987 could be totally blocked (0.5 h:F(4,17)=12.71, P<0.0001; t=5.139, p=0.0008; 1 h:F(4,17)=15.99, P<0.0001; t=5.975, p=0.0002; 1.5 h:F(4,17)=15.22, P<0.0001; t=5.75, p=0.0002; 2h: F(4,17)=12.02, P<0.0001; t=4.848, p=0.0015; 2.5 h:F(4,17)=6.656, P=0.0021; t=3.725, p=0.0168) (Figure 3B). These results indicated that there were dose-dependent analgesic effects by pharmacological activation of α7-nAChRs.
 |
Figure 3 Pharmacologically activating α7-nAChRs produce analgesic effect in CIBP rats. (A) Schematic diagram of the experimental procedure. (B) Single-dose PNU-282,987 (i.t., 0.5, 0.25 and 0.1 mg/kg) was given on day 21 after TCI. The PWTs were tested at 0.5h, 1h, 1.5h, 2h, 2.5h, and 3h after drug injection. PNU-282,987 at the dosage of 0.25 and 0.5 mg/kg dramatically reversed the PWTs in CIBP rats, lasting from 0.5h to2.5 h. The MLA (i.t.10μg/10μL) was administrated 30 min before PNU-282,987 (i.t., 0.5mg/kg) injection. Results showed that MLA could block the analgesic effect ofPNU-282,987.(*P < 0.05, **P < 0.01, ***P < 0.001 versus the NS group; #p<0.05, ##p<0.01, ###p< 0.001 versus the group treated with PNU-282,987 0.5 mg/kg, n =5 in 0.5mg/kg PNU group, MLA+0.5mg/kg PNU group; n =4 in 0.25mg/kg PNU group, 0.1mg/kg PNU group and NS group; PNU=PNU-282,987).(C) PNU-282,987 (i.t.,0.1mg/kg) and PNU-120,596 (i.t., 4μg/10μL) on its own had no obvious analgesic effect in CIBP rats, respectively. However, the analgesic effect of PNU-282,987 (i.t., 0.1mg/kg) was intensified at least at 0.5h and 1.5h by 30min pretreatment of PNU-120,596 (i.t., 4μg/10μL). (*P < 0.05, **P < 0.01 versus the DMSO group, n=4 per group). Abbreviations: α7-nAChR, alpha-7 nicotinic acetylcholine receptor; CIBP, cancer-induced bone pain; PWTs, paw withdrawal thresholds; TCI, tumor cells implantation; MLA, methyllycaconitine; i.t., intrathecal catheterization. |
Positive Allosteric Modulator Intensified Analgesic Effects of α7-nAChRs Agonist in CIBP Rats
Next, we verified whether α7-nAChR positive allosteric modulator PNU-120,596 could intensify the analgesic effect of PNU-282,987. A relative lower dose of PNU-282,987 (0.1mg/kg) or 4μg/10μL PNU-120,596 was intrathecally administrated. When compared with rats given only lower dose of PNU-282,987, the analgesic effect of PNU-282,987 (i.t., 0.1mg/kg) on rats with pretreatment of α7-nAChR positive allosteric modulator PNU-120,596, was enhanced at least at 0.5 h (F(3,12)=7.73, p=0.0039; t=4.68, p=0.0032), 1.5 h(F(3,12)=4.765, p=0.0206; t=3.772, p=0.00160) (Figure 3C).These results showed that positive allosteric modulator PNU-120,596 could intensify the analgesic effect of PNU-282,987. Collectively, these results suggested positive allosteric modulator intensifies analgesic effects of α7-nAChR agonist in CIBP rats.
Repeated Activation of α7-nAChRs Reversed Downregulated Expression of α7-nAChRs in Spinal Cord and Mechanical Allodynia State in CIBP Rats
The experimental design is shown in Figure 4A. We tested analgesic effect of chronic treatment 0.5mg/kg PNU-282,987 in CIBP rats. PNU-282,987 was administered once a day from day 14 to day 18 after tumor cells implantation. When compared with controlled rats, the PWTs were increased distinctly in CIBP rats treated with PNU-282,987 at day 14 (F (3, 16) = 89.33, p<0.0001; t=4.366, p=0.0029) day 15 (F (3, 16) = 78.96, P<0.0001; t=6.215, p<0.0001), day 16 (F (3, 16) = 50.75, P<0.0001; t= 6.335, p <0.0001), day 17 (F (3, 16) = 42.48, p<0.0001; t=6.2, p<0.0001) and day 18 (F (3, 16) = 44.6, P<0.0001; t=5.511, p=0.0002) after tumor cells implantation (Figure 4B).
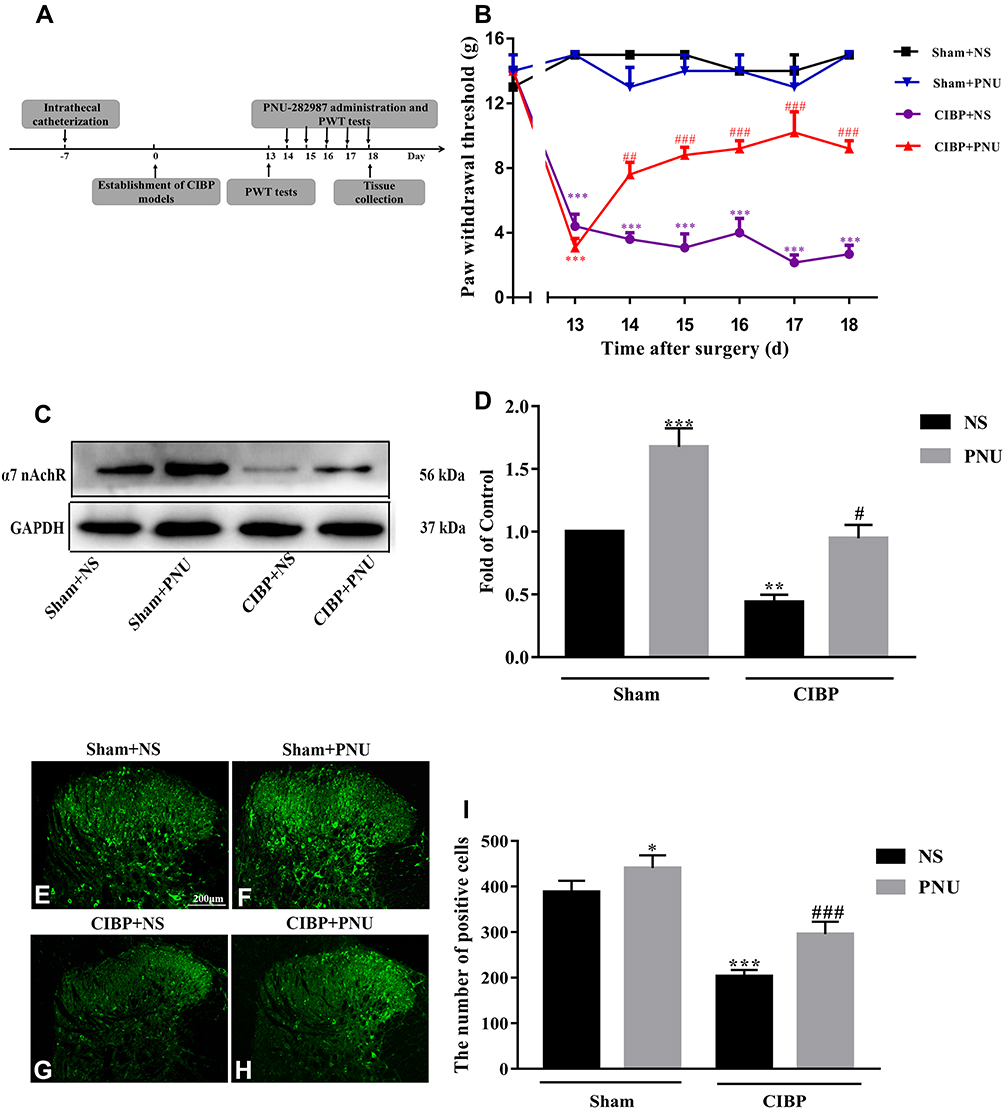 |
Figure 4 Repeat activating α7-nAChRs reversed downregulation expression of α7-nAChRs in spinal cord and mechanical allodynia state in CIBP rats. (A) Schematic diagram of the experimental procedure. (B) The dose of 0.5mg/kg PNU-282,987 was administered once daily from day 14 to day 18 after tumor cells implantation. The behavioral data was acquired at 2.0h after PNU-282,987 injection. End results showed that repeated injection of PNU-282,987 (0.5mg/kg, i.t.) alleviated the mechanical allodynia in CIBP rats. However, no obvious changes of PWTs were observed in CIBP + NS group (***p< 0.001 versus the sham + NS group; ##P < 0.01, ###p< 0.001 versus the CIBP + NS group, n=5 per group; PNU=PNU-282,987).(C, D) Western blot data showed that PNU-282,987 upregulated the protein expression of α7-nAChRs not only in CIBP rats but in sham rats (**P < 0.01, ***P < 0.001 versus the Sham+NS group, #p<0.05 versus the CIBP+NS group, n = 5 per group; PNU=PNU-282,987). (E–I) Photomicrographs showed that PNU-282,987 reversed CIBP-induced downregulation of α7-nAChRs immunoreactivity in the spinal cord in CIBP and normal rats. (*P < 0.05, ***P < 0.001 versus the Sham+NS group; ###P < 0.001 versus the CIBP+NS group, n=5 per group; PNU=PNU-282,987). PNU-282,987 (i.t., 0.5 mg/kg) or NS (10μL) was administrated once a day from day 14 to day 18 after TCI. All samples were harvested after PWT tests (PNU=PNU-282,987). Abbreviations: α7-nAChR, alpha-7 nicotinic acetylcholine receptor; CIBP, cancer-induced bone pain; PWTs, paw withdrawal thresholds; TCI, tumor cells implantation; NS, normal saline; i.t., intrathecal catheterization. |
To explore whether repeated administration of PNU-282,987 played a role in analgesia by restoring the expression of α7-nAChRs, the expression of α7-nAChRs in the spinal cord of CIBP rats was tested by Western blot and immunofluorescence. All rats were conducted with PNU-282,987 (i.t.,0.5 mg/kg) or normal saline chronically from day 14 to day 18 after TCI. In CIBP rats, α7-nAChRs protein level was significantly increased compared to rats treated with normal saline (F (1,16) = 45.33, P=0.0090; t=3.766, p= 0.0101). Further, in sham rats, the protein expression of α7-nAChRs also higher in PNU-282,987 treated rats than NS treated rats (F (1,16) = 45.33, P=0.0090; t=4.99, p=0.0008) (Figure 4C and D). In Figure 4E–I, immunofluorescence staining results were consistent with the Western blot data, either in sham group (F(1, 16) =45.59, p<0.0001; t=3.472, p=0.0189) or CIBP group (F(1, 16) =45.59, p<0.0001; t=6.077, p<0.00010). Collectively, repeated activation of α7-nAChRs could upregulate α7-nAChRs expression and reverse mechanical allodynia in CIBP rats.
Repeated Activation of α7-nAChRs Inhibited Spinal NF-κB Signaling Pathway in CIBP Rats
To further explore the possible post-receptor mechanism of α7-nAChRs, we tested the spinal expression of NF-κB-p65 protein. Firstly, we tested the protein level of NF-κB-p65 in CIBP rats. When compared with sham rats, the expression level of NF-κB-p65 significantly increased in CIBP rats on day 21 after TCI (t=5, df=8, p=0.0011) (Figure 5A and B). Then, we evaluate its expression level after repeated PNU-282,987 (i.t. 0.5mg/kg) treatment for 5 days (from day 14 to day 18 after TCI). In CIBP rats when treated with PNU-282,987, NF-κB-p65 protein level was obviously below the NS group (F (1,16) = 14.84, P=0.0014; t=5.786, p= 0.002). And in sham rats, the expression level of NF-κB-p65 did not change whenever PNU-282,987 was given or not (F (1,4) = 22.54, P=0.0090; t= 2.124, p= 0.6053) (Figure 5C and D). Therefore, it is likely that the spinal NF-κB signaling pathway got involved in the post-receptor mechanism in the analgesic effect of PNU-282,987.
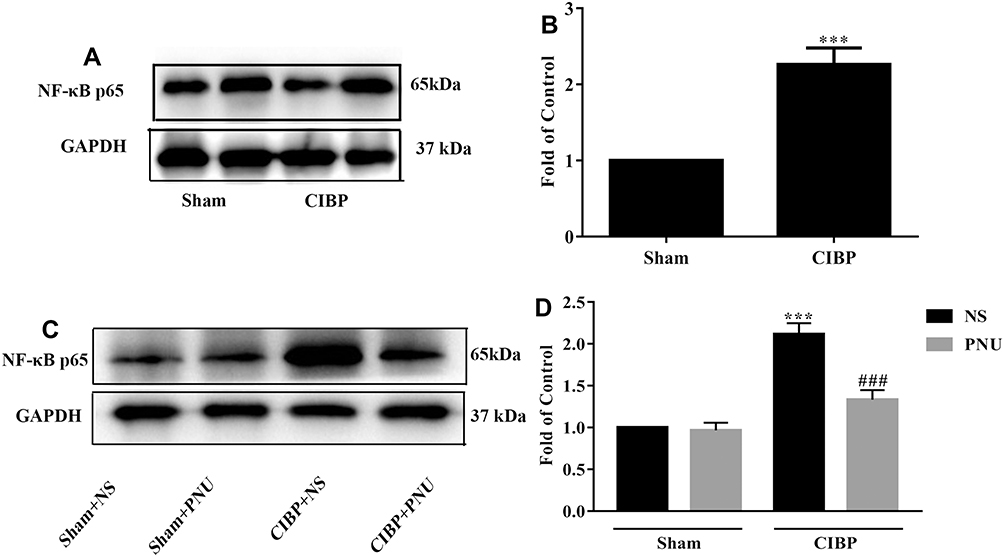 |
Figure 5 Repeat activating α7-nAChRs inhibit spinal NF-κB Signaling Pathway in CIBP rats. Western blot analysis indicated NF-κB-p65 protein level increased on day 21 after TCI (***P <0.01 versus the sham group, n =5 per group) (A, B). (C, D) Western blot analysis showed that after PNU-282,987 (i.t. 0.5mg/kg) treatment for 5 days (from day 14 to day 18 after TCI), the protein expression of NF-κB-p65 was downregulated (***P <0.001versus the Sham+NS group, ###p< 0.001 versus the CIBP+NS group, n = 5 per group; PNU=PNU-282,987). Abbreviations: α7-nAChR, alpha-7 nicotinic acetylcholine receptor; NF-κB, nuclear factor kappa B; CIBP, cancer-induced bone pain; PWTs, paw withdrawal thresholds; TCI, tumor cells implantation; NS, normal saline; i.t., intrathecal catheterization. |
Discussion
The treatment of cancer-induced bone pain (CIBP) remains a major clinical challenge for its elusive mechanisms. The α7-nAChRs are widely expressed in the pain transmission pathway and could be a potential non-opioid target for analgesia. Here in this study, by taking a CIBP model, we found spinal α7-nAChRs contribute to the maintenance of CIBP.
The results revealed that spinal α7-nAChRs mainly located in neurons and the expression of α7-nAChR reduced at least from day 14 in a CIBP model. Previously, investigators also found that the expression level of spinal α7-nAChRs was downregulated in terms of both mRNA and protein in a rat oxaliplatin-induced neuropathic pain model.18 These findings indicate that the downregulation of α7-nAChRs may be a vital contributor to the maintenance of chronic pain. Activation of α7-nAChRs by its agonists has been reported to produce analgesic effects in several chronic pain models17–19,37–40 but lacks efficacy in some acute pain.41–43 Here in the CIBP model, activation of α7-nAChRs exerted analgesic effects in a dose-dependent manner and positive allosteric modulator PNU-120,596 intensified the effects of PNU-282,987. Thus, α7-nAChRs could be a potential non-opioid target for CIBP. The reasons why α7-nAChR agonist was valid in chronic pain, but invalid in some acute pain models have not to be revealed. One of the greatest possibilities could be its indirect action, by targeting NR2B-containing NMDA receptor, which is a key molecular of central sensitization44,45 and also largely involved in chronic pain state including CIBP.46,47 Activation of α7-nAChRs inhibited phosphorylation of NR2B in the spinal dorsal horn,34 and reduced the amplitude of NMDA mediated postsynaptic currents through a direct interaction with the NR2B subunit of the NMDAR.48 As a result, the degree of central sensitization was lowered.
Another finding was that repeated activation of α7-nAChRs could partially reverse mechanical allodynia state in CIBP rats. One reason could be restoring the expression of α7-nAChRs. The protein level of α7-nAChRs in CIBP rats was restored after repeated intrathecal treatment of PNU-282,987. Our findings were very consistent with some other studies. Christensen et al have discovered repeated administration of α7-nAChR agonists could widely increase α7-nAChRs expression in the juvenile rat brain.49 Investigators demonstrated chronic PNU-282,987 treatment augments α7-nAChRs expression in sciatic nerve, DRG and spinal cord of sham rats18 as well. The exact mechanisms for upregulation of α7-nAChRs after repeated activation are not quite clear. Some researchers considered that, in the continuous presence of nAChR agonist, nAChRs might be locked in the membrane, the turnover and degradation pathway is reduced or prevented.50 And some investigators thought it is about receptor trafficking, subunit maturation and assembly, changes in subunit stoichiometry and nAChR conformational changes.51 Another possible reason for chronic administration of α7-nAChR agonist to alleviate mechanical allodynia in CIBP rats could be by inhibiting NF-κB signaling pathway. The following relevant intracellular signaling pathways may be involved. Janus kinase 2 (JAK2)/signal transducer and activator of transcription 3 (STAT3) may participate in the prevention of NF-kB/p65 activity after α7-nAChRs activation.52 Besides this, JAK2/phosphatidylinositol-3 kinase (PI3K)/protein kinase B (AKT) pathway53 and cyclooxygenase2 (COX2)/prostaglandin E2 (PGE2) pathway54 are also involved. Specific as follows: the activation of α7-nAChRs results in enhancement of JAK2-mediated tyrosine-phosphorylation of PI3K, AKT as well as activation of cAMP response element-binding protein (CREB).55–57 Besides these, α7-nAChRs activation leads to stimulation of adenylate cyclase-1 (AC1) which then increases cAMP level, in turn, protein kinase A (PKA) is stimulated and CREB is activated ultimately.58 Subsequently, activated CREB competes with NF-kB for limited amounts of CREB binding protein (CBP)/p300 to inhibit NF-kB activity.55–57 In this study, NF-κB-p65 protein level was upregulated on day 21 after TCI; however, after chronic administration with PNU-282987, the protein expression of NF-κB-p65 was downregulated, suggesting that activation of α7-nAChRs reveres mechanical allodynia in CIBP model by inhibition of NF-κB signaling pathway.
In summary, we demonstrated that spinal α7-nAChRs contribute to the maintenance of CIBP, activation of α7-nAChRs produced a powerful analgesic effect. A positive allosteric modulator could intensify its effects. And recovery of α7-nAChRs expression attenuated CIBP-induced mechanical allodynia, at least partly, by inhibiting the NF-κB signaling pathway in this study. Thus, α7-nAChRs could be a potential non-opioid pharmacological target for CIBP.
Conclusion
Our research demonstrated that spinal α7-nAChRs were downregulated which mediated pain hypersensitivity in bone cancer pain, and restoring diminished α7-nAChRs attenuates CIBP-induced mechanical allodynia, at least partly, by inhibiting the NF-κB signaling pathway in this study. Therefore, this new knowledge indicated that spinal α7-nAChRs may become a valuable target for the therapy of CIBP.
Acknowledgments
This work was financially supported by grants from the National Natural Science Foundation of China (81571053 to Y. T. and 81974170 to X.T.), and Youth Innovation Promotion Association of Chinese Academy of Sciences (Y6Y0021004 to J. W.).
Disclosure
The authors declare that they have no conflicts of interest for this work.
References
1. Kane CM, Hoskin P, Bennett MI. Cancer induced bone pain. BMJ. 2015;350(jan29 7):h315. doi:10.1136/bmj.h315
2. Yoneda T, Hiasa M, Okui T. Crosstalk between sensory nerves and cancer in bone. Curr Osteoporos Rep. 2018;16(6):648–656. doi:10.1007/s11914-018-0489-x
3. Falk S, Dickenson AH. Pain and nociception: mechanisms of cancer-induced bone pain. J Clin Oncol. 2014;32(16):1647–1654. doi:10.1200/JCO.2013.51.7219
4. van den Beuken-van Everdingen MH, de Rijke JM, Kessels AG, Schouten HC, van Kleef M, Patijn J. Prevalence of pain in patients with cancer: a systematic review of the past 40 years. Ann Oncol. 2007;18(9):1437–1449. doi:10.1093/annonc/mdm056
5. Dibai-Filho AV, Ferreira VT, Kelly de Oliveira A, Gomes CA, Melo ES, Maria de Almeida A. Assessing the impact of pain on the life of breast cancer survivors using the brief pain inventory. J Phys Ther Sci. 2015;27(5):1361–1363. doi:10.1589/jpts.27.1361
6. Khan L, Uy C, Nguyen J, et al. Self-reported rates of sleep disturbance in patients with symptomatic bone metastases attending an outpatient radiotherapy clinic. J Palliat Med. 2011;14(6):708–714. doi:10.1089/jpm.2010.0491
7. Parker KP, Berger AM
8. WHO. WHO’s cancer pain ladder for adults. 2015. Available from: http://www.who.int/cancer/palliative/painladder/en/.
9. Harris JD. Management of expected and unexpected opioid-related side effects. Clin J Pain. 2008;24(Suppl 10):S8–S13. doi:10.1097/AJP.0b013e31816b58eb
10. Weber M, Huber C. Documentation of severe pain, opioid doses, and opioid-related side effects in outpatients with cancer: a retrospective study. J Pain Symptom Manage. 1999;17(1):49–54. doi:10.1016/S0885-3924(98)00103-1
11. Radcliffe KA, Dani JA, Pidoplichko VI. Variations in desensitization of nicotinic acetylcholine receptors from hippocampus and midbrain dopamine areas. Eur J Pharmacol. 2000;393(1–3):31–38. doi:10.1016/S0014-2999(00)00003-0
12. Devillers-Thiéry A, Bertrand D
13. Hackett SF, Seidel C, Abraham S, et al. The nicotinic cholinergic pathway contributes to retinal neovascularization in a mouse model of retinopathy of prematurity. Invest Ophthalmol Vis Sci. 2017;58(2):1296–1303. doi:10.1167/iovs.16-20670
14. Hołuj M, Potasiewicz A, Kos T, Popik P, Arias HR, Nikiforuk A. 3-furan-2-yl-N-p-tolyl-acrylamide, a positive allosteric modulator of the α7 nicotinic receptor, reverses schizophrenia-like cognitive and social deficits in rats. Neuropharmacology. 2017;113(Pt A):188–197. doi:10.1016/j.neuropharm.201
15. Zhou Y, Liu PR, Zhang Y, Diao S. Electroacupuncture alleviates surgery-induced cognitive dysfunction by increasing α7-nAChR expression and inhibiting inflammatory pathway in aged rats. Neurosci Lett. 2017;659:1–6. doi:10.1016/j.neulet.2017.08.043
16. Munro G, Hansen R, Erichsen H, Timmermann D, Christensen J, Hansen H. The alpha7 nicotinic ACh receptor agonist compound B and positive allosteric modulator PNU-120596 both alleviate inflammatory hyperalgesia and cytokine release in the rat. Br J Pharmacol. 2012;167(2):421–435. doi:10.1111/j.1476-5381.2012.02003.x
17. Shi G-N, Liu Y-L, Lin H-M, et al. Involvement of cholinergic system in suppression of formalin-induced inflammatory pain by cobratoxin. Acta Pharmacol Sin. 2011;32(10):1233–1238. doi:10.1038/aps.2011.65
18. Mannelli LD, Pacini A, Matera C, et al. Involvement of α7 nAChR subtype in rat oxaliplatin-induced neuropathy: effects of selective activation. Neuropharmacology. 2014;79(p):37–48. doi:10.1016/j.neuropharm.2013.10.034
19. Pacini A, Di Cesare Mannelli L, Bonaccini L, Ronzoni S, Bartolini A, Ghelardini C. Protective effect of alpha7 nAChR: behavioural and morphological features on neuropathy. Pain. 2010;150(3):542–549. doi:10.1016/j.pain.2010.06.014
20. Gong S, Liang Q, Zhu Q, et al. Nicotinic acetylcholine receptor alpha7 subunit is involved in the cobratoxin-induced antinociception in an animal model of neuropathic pain. Toxicon. 2015;93:31–36. doi:10.1016/j.toxicon.2014.11.222
21. Meyer EM, Damaj MI, Martin BR. The antinociceptive effects of α7 nicotinic agonists in an acute pain model. Neuropharmacology. 2000;39(13):2785–2791. doi:10.1016/S0028-3908(00)00139-8
22. Makarov SS. NF-kappaB as a therapeutic target in chronic inflammation: recent advances. Mol Med Today. 2000;6(11):441–448. doi:10.1016/S1357-4310(00)01814-1
23. Luo J, Sun T, Jia M, Li H, Li K, Fu Z. Small interfering RNA-mediated knockdown of NF-kappaBp65 attenuates neuropathic pain following peripheral nerve injury in rats. Eur J Pharmacol. 2012;682(1–3):79–85. doi:10.1016/j.ejphar.2012.02.017
24. Zhou YL, Jiang GQ, Wei J, et al. Enhanced binding capability of nuclear factor-kappaB with demethylated P2X3 receptor gene contributes to cancer pain in rats. Pain. 2015;156(10):1892–1905. doi:10.1097/j.pain.0000000000000248
25. Song ZP, Xiong BR, Guan XH, et al. Minocycline attenuates bone cancer pain in rats by inhibiting NF-kappaB in spinal astrocytes. Acta Pharmacol Sin. 2016;37(6):753–762. doi:10.1038/aps.2016.1
26. Wang Y, Ni H, Li H, et al. Nuclear factor kappa B regulated monocyte chemoattractant protein-1/chemokine CC motif receptor-2 expressing in spinal cord contributes to the maintenance of cancer-induced bone pain in rats. Mol Pain. 2018;14:1744806918788681. doi:10.1177/1744806918788681
27. Patel H, McIntire J, Ryan S, Dunah A, Loring R. Anti-inflammatory effects of astroglial alpha7 nicotinic acetylcholine receptors are mediated by inhibition of the NF-kappaB pathway and activation of the Nrf2 pathway. J Neuroinflammation. 2017;14(1):192. doi:10.1186/s12974-017-0967-6
28. Slosky LM, Largent-Milnes TM, Vanderah TW. Use of animal models in understanding cancer-induced bone pain. Cancer Growth Metastasis. 2015;8:47–62.
29. Fu Q, Shi D, Zhou Y, et al. MHC-I promotes apoptosis of GABAergic interneurons in the spinal dorsal horn and contributes to cancer induced bone pain. Exp Neurol. 2016;286:12–20. doi:10.1016/j.expneurol.2016.09.002
30. Chen SP, Zhou YQ, Wang XM, et al. Pharmacological inhibition of the NLRP3 inflammasome as a potential target for cancer-induced bone pain. Pharmacol Res. 2019;147:104339. doi:10.1016/j.phrs.2019.104339
31. Tall JM, Zhao C
32. Lee H. Effects of co-administration of intrathecal nociceptin/orphani FQ and opioid antagonists on formalin-induced pain in rats. Yonsei Med J. 2013;54(3):763–771. doi:10.3349/ymj.2013.54.3.763
33. Sun R, Zhang W, Bo J, et al. Spinal activation of alpha7-nicotinic acetylcholine receptor attenuates posttraumatic stress disorder-related chronic pain via suppression of glial activation. Neuroscience. 2017;344:243–254. doi:10.1016/j.neuroscience.2016.12.029
34. Zhang W, Liu Y, Hou B, Gu X, Ma Z. Activation of spinal alpha-7 nicotinic acetylcholine receptor attenuates remifentanil-induced postoperative hyperalgesia. Int J Clin Exp Med. 2015;8(2):1871–1879.
35. Guan X, Fu Q, Xiong B, et al. Activation of PI3Kgamma/Akt pathway mediates bone cancer pain in rats. J Neurochem. 2015;134(3):590–600. doi:10.1111/jnc.13139
36. Zhou Y-Q, Chen S-P, Liu D-Q, et al. The role of spinal GABAB receptors in cancer-induced bone pain in rats. J Pain. 2017;18(8):933–946. doi:10.1016/j.jpain.2017.02.438
37. Robson Costa EM, Motta MN, Manjavachi MC, Calixto JB. Activation of the alpha-7 nicotinic acetylcholine receptor (α7 nAchR) reverses referred mechanical hyperalgesia induced by colonic inflammation in mice. Neuropharmacology. 2012;63(5):798–805. doi:10.1016/j.neuropharm.2012.06.004
38. Liu Q, Liu C, Jiang L, et al. alpha7 nicotinic acetylcholine receptor-mediated anti-inflammatory effect in a chronic migraine rat model via the attenuation of glial cell activation. J Pain Res. 2018;11:1129–1140. doi:10.2147/JPR.S159146
39. Freitas KC, Carroll FI, Negus SS. Effects of nicotinic acetylcholine receptor agonists in assays of acute pain-stimulated and pain-depressed behaviors in rats. J Pharmacol Exp Ther. 2015;355(2):343–352. doi:10.1124/jpet.115.226803
40. Medhurst SJ, Hatcher JP, Hille CJ, et al. Activation of the alpha7-nicotinic acetylcholine receptor reverses complete freund adjuvant-induced mechanical hyperalgesia in the rat via a central site of action. J Pain. 2008;9(7):580–587. doi:10.1016/j.jpain.2008.01.336
41. Freitas K, Carroll FI, Damaj MI. The antinociceptive effects of nicotinic receptors alpha7-positive allosteric modulators in murine acute and tonic pain models. J Pharmacol Exp Ther. 2013;344(1):264–275. doi:10.1124/jpet.112.197871
42. Papke RL, Bagdas D, Kulkarni AR, et al. The analgesic-like properties of the alpha7 nAChR silent agonist NS6740 is associated with non-conducting conformations of the receptor. Neuropharmacology. 2015;91:34–42. doi:10.1016/j.neuropharm.2014.12.002
43. Bagdas D, Wilkerson JL, Kulkarni A, et al. The alpha7 nicotinic receptor dual allosteric agonist and positive allosteric modulator GAT107 reverses nociception in mouse models of inflammatory and neuropathic pain. Br J Pharmacol. 2016;173(16):2506–2520. doi:10.1111/bph.13528
44. Ali DW, Salter MW. NMDA receptor regulation by Src kinase signalling in excitatory synaptic transmission and plasticity. Curr Opin Neurobiol. 2001;11(3):336–342. doi:10.1016/S0959-4388(00)00216-6
45. Petrenko AB, Yamakura T, Baba H, Shimoji K. The role of N-methyl-D-aspartate (NMDA) receptors in pain: a review. Anesth Analg. 2003;97(4):1108–1116. doi:10.1213/01.ANE.0000081061.12235.55
46. Gu X, Zhang J, Ma Z, et al. The role of N-methyl-D-aspartate receptor subunit NR2B in spinal cord in cancer pain. Eur J Pain. 2010;14(5):496–502. doi:10.1016/j.ejpain.2009.09.001
47. Yang H, Yan H, Li X, et al. Inhibition of connexin 43 and phosphorylated NR2B in spinal astrocytes attenuates bone cancer pain in mice. Front Cell Neurosci. 2018;12:129. doi:10.3389/fncel.2018.00129
48. Zhou X, Zong Y, Zhang R, et al. Differential modulation of GABAA and NMDA Receptors By An Α7-Nicotinic Acetylcholine Receptor Agonist In Chronic Glaucoma. Front Mol Neurosci. 2017;10:422. doi:10.3389/fnmol.2017.00422
49. Christensen DZ, Mikkelsen JD, Hansen HH, Thomsen MS. Repeated administration of alpha7 nicotinic acetylcholine receptor (nAChR) agonists, but not positive allosteric modulators, increases alpha7 nAChR levels in the brain. J Neurochem. 2010;114(4):1205–1216. doi:10.1111/j.1471-4159.2010.06845.x
50. Buisson B, Bertrand D. Nicotine addiction: the possible role of functional upregulation. Trends Pharmacol Sci. 2002;23(3):130–136. doi:10.1016/S0165-6147(00)01979-9
51. Govind AP, Vezina P, Green WN. Nicotine-induced upregulation of nicotinic receptors: underlying mechanisms and relevance to nicotine addiction. Biochem Pharmacol. 2009;78(7):756–765. doi:10.1016/j.bcp.2009.06.011
52. de Jonge WJ, van der Zanden EP, The FO, et al. Stimulation of the vagus nerve attenuates macrophage activation by activating the Jak2-STAT3 signaling pathway. Nature Immunology. 2005;6(8):844–851.
53. Shaw S, Bencherif M, Marrero MB. Janus kinase 2, an early target of alpha 7 nicotinic acetylcholine receptor-mediated neuroprotection against abeta-(1–42) amyloid. J Biol Chem. 2002;277(47):44920–44924. doi:10.1074/jbc.M204610200
54. Takahashi HK, Iwagaki H, Hamano R, Yoshino T, Tanaka N, Nishibori M. alpha7 nicotinic acetylcholine receptor stimulation inhibits lipopolysaccharide-induced interleukin-18 and −12 production in monocytes. J Pharmacol Sci. 2006;102(1):143–146. doi:10.1254/jphs.SC0060074
55. Wang H, Liao H, Ochani M, et al. Cholinergic agonists inhibit HMGB1 release and improve survival in experimental sepsis. Nat Med. 2004;10(11):1216–1221. doi:10.1038/nm1124
56. Shimada K, Sugano N, Ito K, Murai S. Nicotine inhibits the production of inflammatory mediators in U937 cells through modulation of nuclear factor-kappaB activation. Biochem Biophys Res Commun. 1998;252(1):25–28. doi:10.1006/bbrc.1998.9599
57. Yoshikawa H, Kurokawa M, Ozaki N, et al. Nicotine inhibits the production of proinflammatory mediators in human monocytes by suppression of I-kappaB phosphorylation and nuclear factor-kappaB transcriptional activity through nicotinic acetylcholine receptor alpha7. Clin Exp Immunol. 2006;146(1):116–123. doi:10.1111/j.1365-2249.2006.03169.x
58. Cheng Q, Yakel JL. Activation of alpha7 nicotinic acetylcholine receptors increases intracellular cAMP levels via activation of AC1 in hippocampal neurons. Neuropharmacology. 2015;95:405–414. doi:10.1016/j.neuropharm.2015.04.016
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.