")
Back to Archived Journals » Open Access Insect Physiology » Volume 5
The role of water, ice nucleators, and inoculation in insect cold survival
Authors Rozsypal J
Received 17 March 2015
Accepted for publication 15 July 2015
Published 13 August 2015 Volume 2015:5 Pages 21—30
DOI https://doi.org/10.2147/OAIP.S64952
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Guy Smagghe
Jan Rozsypal,1,2
1Biology Centre of the Academy of Sciences of the Czech Republic, Institute of Entomology, Ceské Budejovice, Czech Republic; 2Graduate School of Science, Osaka City University, Osaka, Japan
Abstract: Over the past century and especially the last few decades a significant progress has been made in our understanding of the mechanisms that underlie the capacity of insects and other ectothermic animals to survive at subzero temperatures. Although we now understand relatively well the complexity of the cold tolerance phenomenon, from the molecular to the ecological level, some aspects remain elusive. This review summarizes current knowledge on the mechanisms of cold tolerance and focuses on the three following issues that remain unsolved in this field: 1) the role of water in preservation of biological structures in organisms that exploit dehydration as a part of their cold survival strategy, 2) the cause of ice nucleation in freeze-intolerant organisms below the temperature range of the activity of ice nucleating proteins and above the temperature of homogeneous nucleation of water, and 3) the capacity of some primarily freeze-intolerant insects to tolerate freezing upon inoculation with ice at high subzero temperatures.
Keywords: insect cold tolerance, dehydration, ice nucleators, inoculation
Introduction
Insects represent one of the most successful groups of organisms and are found in a wide range of different environments. Species inhabiting temperate and polar regions have evolved a variety of adaptations to cope with seasonally changing conditions. Many of these adaptations target, either directly or indirectly, a single substance – water. Some appear to have evolved to modulate the phase behavior of water at low temperatures and others seem to protect various biological structures from the potential damage caused by the phase behavior or the absence of water. The mechanisms that allow insects to survive during winter are referred to as cold tolerance adaptations.
Most insects belong to one of the two basic categories depending on their cold tolerance strategy: freeze-intolerant (also referred to as freeze-avoidant or chill-susceptible) and freeze-tolerant.1–3 The majority of insects belong to the freeze-intolerant category and survive at subzero temperatures in a metastable, supercooled state by depressing the freezing point of their body fluids (supercooling point, SCP). These species cannot tolerate the formation of ice within their bodies. The freeze-intolerant organisms, which keep their body water in a liquid state, even at extremely low temperatures, may be subject to the rapid growth of large ice crystals, which damage their cells and tissues. Freeze-tolerant insects can tolerate the formation of internal ice, provided that the ice crystals are restricted to extracellular spaces.2 In contrast to freeze-intolerant species, the freeze-tolerant species typically have relatively high SCPs, which prevent the explosive growth of ice crystals that occur when freezing at extremely low temperatures takes place.4,5 The ice that forms in the extracellular spaces of freeze-tolerant organisms increases the osmotic pressure of extracellular fluids compared with intracellular fluids, which draws water from the cells. The partial removal of water from cells results in cellular freeze-dehydration and an increase in the concentration of intracellular fluids, which prevents intracellular freezing.6,7
Toward the end of the 20th century, an alternative strategy, described as cryoprotective dehydration, was identified in some edaphic invertebrates, including insects, that have permeable cuticles.8–12 These animals can survive partial dehydration. The removal of water in these species occurs due to the vapor pressure gradient between the ice in frozen soil and the internal supercooled body fluids. The gradient drives the loss of body water to the soil ice until an equilibrium is reached, which results in partial dehydration and an increase in the concentration of body fluids. This depresses the freezing point and prevents internal freezing.6,7 The body water in species relying on cryoprotective dehydration actually freezes outside the body. One extreme dehydration strategy, when an organism can lose as much as 90% of its normal water content, or more, is called anhydrobiosis (a kind of cryptobiosis).13,14 Organisms undergoing dehydration typically accumulate polyhydroxy compounds, such as disaccharides, which are highly effective protectants. Some of these substances may form amorphous glass at very low water contents.15 Glass transition (vitrification) can also occur in organisms that only partially dehydrate and some researchers recognize it as fourth category of cold tolerance.16,17 Species that use any of these three/four strategies must deal with some common issues. Most importantly, they must ensure that their biological structures remain hydrated/protected. Even species employing cryoprotective dehydration must protect their biological structures from damage, which is believed to be possible only in the presence of water or, possibly, protective substances like disaccharides.17
Organisms that rely on the freeze-tolerance strategy typically maintain the freezing point of their body fluids at high subzero temperatures. Freezing at higher temperatures proceeds slower, produces smaller ice crystals, and leaves enough time for the osmotic pressure between freezing extracellular and unfrozen intracellular fluids to equilibrate.7 The main mechanism by which freeze-tolerant organisms maintain the freezing point of their body fluids at higher temperatures is believed to be through the activity of ice nucleators, which are mostly proteinaceous in nature. Ice nucleator proteins have been identified in many freeze-tolerant organisms, from bacteria to vertebrates.18 While the nucleators responsible for freezing in freeze-tolerant organisms are relatively well understood, the cause of nucleation in freeze-intolerant organisms, and in various aqueous solutions and water samples, is unknown. The generally accepted idea, however, is that various unidentified particles may serve as nuclei in these systems.
Many animals overwinter in protected hibernacula, such as soil or litter layer, which provide relatively good protection from extremely low temperatures. However, such microhabitats tend to be wet. In the moist environment, when temperatures drop below zero, the risk of inoculative freezing increases rapidly.19,20 Nevertheless, what may represent a risk to one species can often be considered as an opportunity for another. Since ice itself is probably the best known nucleator, freeze-tolerant invertebrates and some freeze-tolerant vertebrates may seek out overwintering sites which contain ice in order to initiate freezing at high subzero temperatures through inoculation.21–23
The aim of the present review is to discuss several fundamental, yet unsolved, issues surrounding cold tolerance mechanism research. Some of these issues have been debated for decades, others have not received as much attention, although they could be critical factors that affect cold survival in many organisms or be an important component of a survival strategy. This review will first discuss the importance of water in biological systems, its role in dehydration-resistant organisms, and the mechanisms used to protect biological structures at low water contents. It will then focus on the nature of ice nucleators, and the causes of nucleation in freeze-intolerant organisms. The final part of the review will concentrate on the importance of inoculative freezing during cold survival and discuss it as a possible freeze-tolerance mechanism in some organisms.
The role of water in the preservation of biological functions
Water is generally recognized to be the substance that makes biological functions possible. Over the past few decades, a significant progress has been made in our understanding of its role in living organisms. We now perceive water as not just a simple solvent, but as an integral part of biological systems.
Water in cell
The water in cells can be divided into three fractions. Although the terminology varies in the literature, we can distinguish 1) bulk water (also osmotically active or “free” water), 2) hydration water (also interfacial, structural, osmotically inactive, or “bound” water), and 3) confined water (also nanoconfined or vicinal water).24–29 Bulk water is the “ordinary” aqueous solution and is potentially subject to freezing and loss through desiccation.25 Hydration water is in direct contact with the surface of biomolecules, and is believed to be vital for protein, nucleic acid, and membrane function.28–30 Water that is not in contact with any surface, but is in close proximity (on a nanometer scale) or is enclosed between two surfaces, is called confined water.29 The properties of confined water differ from those of bulk water. Many researchers use the term “unfreezable water” for the fraction of water that does not freeze tens of degree Celsius below the equilibrium freezing point, even in the presence of ice. The so-called “unfreezable water” probably includes both hydration water and confined water.26 The confined water has a higher viscosity and is more difficult to remove compared with bulk water. However, some researchers suggest that removal of this fraction of water is not a matter of force, but just a matter of time.26 The low temperature and confined space result in high viscosity, which considerably slows the equilibration time with ice, or even makes it impossible within a relevant time scale, which is why many researchers consider this fraction of water to be “unfreezable”.26
Dehydration
Water is the main constituent of living organisms. Its content ranges from 40% to 90% in invertebrates, with 65%–75% being the most common.31 Many organisms must cope with dehydration, either during overwintering in temperate zones or during dry seasons in the tropics. Resistance to water loss varies among invertebrates. Some are very sensitive and do not survive much below their optimal water content. Others are capable of surviving at various levels of dehydration.31 In particular, those inhabiting dry or desiccating environments may experience extensive dehydration and lose nearly all of their normal water content, and yet still survive, even in a near-anhydrous state.13
All biological structures, under normal conditions, exist within an aqueous environment. Water is thought to be necessary for the correct folding of proteins and maintenance of their functional conformation. It is well known that proteins can be denatured by many factors, such as above- or below-optimal temperature, pressure, pH, and low hydration. Denaturation upon dehydration, however, is reversible for many proteins and such proteins can regain their normal function after rehydration.32 Various proteins maintain their conformation, or at least a residual structure, even in an anhydrous state.33,34 Surprisingly, some enzymes have been shown to exhibit weak activity in pure non-aqueous solvents and were able to catalyze reactions that are not possible in water.35 Nevertheless, it is known that some, even anhydrous proteins, retain a small, residual number of water molecules.36 Even a protein fully “stripped” from its hydration shell may retain several water molecules inside its structure, which could be vital for its function or for regaining normal function upon rehydration. While some proteins are relatively resistant, dehydration may pose a much bigger threat to biological membranes. Liquid-crystalline membranes may transition into the gel phase or lose their integrity due to a transition to the hexagonal phase at above optimal temperatures or under low hydration conditions.37
Anhydrobiosis: advantages and disadvantages
The phase behavior of water at subzero temperatures may pose a serious threat. Organisms that use dehydration to survive adverse conditions evade this threat by removing a substantial portion of body water. Metabolic rate in frozen state is significantly lower compared with supercooled state, but is measurable.38 Since biochemical reactions require water, dehydration results in even more profound decrease in metabolic rate or even makes it non-existent (ametabolism).6,13 Low or suspended metabolism can save energy reserves. Extensive dehydration also prevents the accumulation of potentially toxic products of anaerobic metabolism that may occur in animals surviving in frozen state.39,40 Unlike organisms that overcome unfavorable conditions in hydrated state the organisms capable of extensive dehydration often exhibit tolerance to a wide range of temperatures, even from that of liquid nitrogen to the boiling point of water.13,14 On the other hand, the anhydrobiotic organisms must cope with changes in osmotic pressure and they may be subject to oxidative stress during dehydration.6 Also the absence of water itself may prove deleterious. The organisms employing dehydration have evolved mechanisms to protect their biological structures at low water content.
Mechanisms to protect biological structures at low hydration
The organisms relying on cryoprotective dehydration, and especially anhydrobiosis, typically accumulate relatively high concentrations of substances that can substitute for water. Some disaccharides have these properties, such as trehalose (mostly in animals) and sucrose (in plants).41,42 Especially trehalose is an exceptional stabilizer and can protect proteins and membranes under various stresses including extensive dehydration. The mechanism by which trehalose protects biomolecules and membranes is explained by the “Water Replacement hypothesis” (Figure 1).43,44 The water molecules in the hydration shells of proteins and membranes are replaced by hydrogen bonding with another substance (trehalose).43–46 Trehalose and some other disaccharides can form an amorphous glass at low water content, which further stabilizes the biological structures.15,47 The hydrogen bonding of trehalose to membrane phospholipids and the formation of glass has been found to keep the membrane in a liquid-crystalline state, even at very low water contents, in anhydrobiotic larvae of Polypedilum vanderplanki.48 This protective role has also been identified in some free amino acids. Previous studies have reported that proline in plants and some insects can stabilize various biological structures at low temperatures and low hydration states.49–51 Freeze-tolerant animals and other organisms, which do not dehydrate completely, rely on the non-colligative action of substances like some sugars, polyols, or free amino acids, including trehalose and proline. The mechanism has been described as “preferential hydration and exclusion”.52,53 The solutes in cells compete with water to hydrogen bond to biomolecules and membranes. While solutes are excluded from the vicinity of biomolecules, water hydrogen bonds to the biomolecule, which preferentially hydrates it.
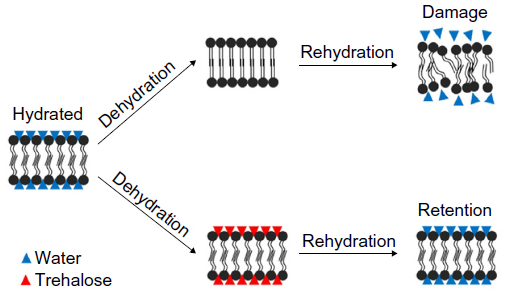 | Figure 1 Water replacement hypothesis. |
Adaptations to dehydration
Some unicellular organisms, which normally do not survive drying, can tolerate extensive freeze-dehydration (lyophilization) when cultured on a medium containing trehalose.54,55 However, no such extensive, artificially induced increase in dehydration tolerance can be found in the literature for complex multicellular organisms, such as insects. Although it is questionable to compare bacteria with multicellular organisms, such a comparison might reveal the mechanism or mechanisms that, in addition to the action of protective substances, are involved in dehydration tolerance in animals. A variety of microorganisms inhabit the most hostile environments on Earth. The reason why proteins and membranes in these organisms do not denature, or denature reversibly, is due to changes in their composition, which results in a more resistant conformation.56 The composition of proteins and membranes in these organisms is a result of selection pressures that have made them more resistant to extreme environmental conditions, such as high temperature, pressure, salinity, and low hydration.56–58 Similar selection pressures on the composition of biomolecules and membranes could be partially responsible for high desiccation tolerance, even in the case of some complex metazoans, such as insects. The “structural” adaptations, in combination with protective mechanisms, may allow the dehydration-tolerant organisms to survive periods of very low water content.
Minimum hydration needed to prevent damage and sustain life
In theory, organisms can lose water until only a single layer of water molecules on the surfaces of biomolecules is left, which is believed to be the critical hydration level required for biological function.30 However, the minimum amount of water needed to keep biological processes operating differs from the amount needed to keep them suspended. Anhydrobiotic and some cryptobiotic/ametabolic organisms defy the definition of life or “being alive” when in a state of suspended animation.59 The “residual” water in anhydrobiotic organisms may thus correspond to, or be even less than, a monolayer of water molecules bound to surfaces of biomolecules and membranes. The hydration level that does not allow biological functions to occur but is sufficient to keep biological structures undamaged could be, at least in theory, very close or even equal to zero. Although “anhydrous” suggests being without water, none of the known anhydrobiotic organisms dehydrate completely.13 More study is needed to clarify the dehydration tolerance limits in anhydrobiotic organisms and whether water is really needed for the preservation of biological structures.
Ice nucleation, ice nucleators, and anti-nucleation
Ice nucleation
Ice nucleation is either homogenous or heterogeneous. Homogenous ice nucleation occurs when sufficient numbers of water molecules aggregate to form an ice crystal (nucleus).60 Homogenous ice nucleation is a stochastic process that depends on temperature and the number of water molecules available to form the initial crystal. The probability of ice nucleation thus increases with decreasing temperature and increasing volume of water. The temperature for homogenous nucleation in small volumes of extremely pure water is close to −40°C.61 Apart from the example of ultra-pure water, all other aqueous solutions, including those in living organisms, are believed to undergo heterogeneous nucleation. With heterogeneous nucleation, a substance other than water serves as a nucleus around which water molecules aggregate. Such substances are called heterogeneous nucleators.5,62
Ice nucleating proteins
Some organic substances and a few inorganic compounds, such as silver iodide, have been shown to have nucleating activity.63 Although many substances are known or thought to exhibit ice nucleating activity, the most studied category of ice nucleators is protein based. The ice nucleating proteins have been identified in various organisms, including insects.64,65 The activity temperature of ice nucleating proteins is approximately −2°C in ice nucleating bacteria and ranges from approximately −5°C to −10°C in other freeze-tolerant organisms.66,67 All freezing events in this range can probably be attributed to the action of proteinaceous ice nucleators.
The proteinaceous nucleators’ mechanism of action is not completely understood. However, ice nucleating proteins typically contain large repeating domains whose structure is believed to resemble that of ice and probably “arranges” water molecules into an ice lattice.68,69
Causes of ice nucleation in freeze-intolerant insects
Freezing below the range of activity of ice nucleating proteins is believed to be due to the activity of other nucleators, of a non-proteinaceous nature. This category of nucleators is often referred to as “impurities” or “dust particles”. They could be responsible for nucleation in freeze-intolerant organisms, as well as in various aqueous solutions and water samples that freeze above the temperature for homogeneous nucleation of water.5,70 Although this explanation is widely accepted by many researchers, some authors question the existence of this category of nucleators. Zachariassen et al showed that homogenous nucleation can occur even at temperatures well above −40°C (the generally accepted temperature for the homogeneous nucleation of pure water).71 This suggests that all freezing below the minimum activity temperature of ice nucleating proteins should be considered homogeneous, and that it is dependent on just the temperature and volume of water. The widely accepted concept of various “particles”, however, cannot be dismissed that easily. This category of nucleators, although poorly defined in biology, is relatively well understood in meteorology where various substances and dust particles have been shown to be responsible for nucleation in clouds.72,73 Both naturally occurring and artificial mineral dusts and other materials have been tested for nucleating activity in atmosphere. Many natural dusts were found to initiate freezing in temperature range between −12°C and −25°C.73,74 Volcanic ash that initiates freezing from −11°C proved to be one of the most effective inorganic nucleators.75 High nucleating activity was described also in artificial dusts. Commercially produced Arizona Test Dust, commonly used to test the efficiency of filters, has been found to be an effective nucleator with activity at approximately −17°C.76
The ice nucleation activity of dust particles could perhaps be tested also in insects. Addition of either natural or artificial dust into insect diet could result in an increase in SCP and thus provide evidence that particles of mineral dust might act as nucleators in animals. However, such evidence would only be indirect and further research would be needed to clarify the real cause of nucleation in freeze-intolerant insects and other animals.
The role of ice nucleators in organisms
Lundheim distinguishes two categories of ice nucleators according to their role in organisms.18 The proteinaceous ice nucleators in freeze-tolerant organisms can be considered “adaptive” because they benefit their users. The nucleators in other organisms, where they do not provide any advantage or even pose a threat, can be considered “incidental”. Adaptive nucleators are most likely part of an organism’s phenotype or survival strategy. In contrast, incidental nucleators are probably of extrinsic origin and could be inadvertently ingested with food.70 The adaptive nucleators are represented mostly by ice nucleating proteins. However, some other substances, even inorganic ones, may belong to this category as well. The calcium phosphate spherules, identified in the malpighian tubules of overwintering, freeze-tolerant goldenrod gall fly (Eurosta solidaginis) larvae, increased the crystallization temperature of Schneider’s insect medium enough to explain the crystallization temperature of the whole larva.77
In freeze-tolerant organisms, nucleators initiate freezing at high subzero temperatures. Previous studies have reported that some freeze-tolerant insects can synthesize their own nucleators, while others may either ingest them along with food or the nucleators are produced by microorganisms that are part of the normal microflora in insect guts.64,65,78–80 In freeze-intolerant species, the presence of ice nucleators is potentially lethal. These insects, which rely on extensive supercooling, must remove ice nucleators from their body to eliminate the risk of their body fluids freezing.70
Antinucleation
There are several known mechanisms for removing ice nucleators or masking their activity. Many freeze-intolerant insects empty their gut before winter to remove various particles that have been ingested along with food and ice nucleating microorganisms that have either been ingested with food or are part of the normal gut microflora. For example, several collembolans and a coleopteran Phyllotreta undulata have been shown to significantly decrease their SCPs upon cessation of feeding and/or evacuation of gut.81–83 Another mechanism is masking the activity of the ice nucleators present in hemolymph. Besides the freezing (and melting) point depression caused by the colligative action of substances like sugars and polyols, a number of insects and other organisms synthesize so-called “antifreeze proteins”.84 The presence of antifreeze proteins (also referred to as thermal hysteresis proteins) manifests as thermal hysteresis, which is the difference between the freezing point and melting point of body fluids. The action of antifreeze proteins is thus non-colligative. Antifreeze proteins are a diverse group that have evolved in parallel in different species.85,86 Despite their diversity, the mechanism of action of these proteins seems to be by adsorption to ice and restricting the growth of ice crystals.87 The antifreeze proteins are also produced in freeze-tolerant species to prevent the growth of individual ice crystals and crystal merging – recrystallization.86
Inoculative freezing
Inoculation in freeze-tolerant insects
Some freeze-tolerant species are known to seek hibernacula that not only protect them from extremely low temperatures, but also initiate freezing at high subzero temperatures through inoculation. A study found that the freeze-tolerant centipede Lithobius forficatus only survived freezing when inoculated with external ice at mild subzero temperatures.88 Freeze-tolerant crane fly larva (Tipula sp.) and freeze-tolerant wooly bear caterpillar (Pyrrharctia isabella) also require inoculation to survive internal freezing.89,90 These arthropods apparently have adopted inoculative freeze-tolerance as their primary strategy for cold survival. Such species, when in dry environment, typically do not supercool lower than 10°C below the equilibrium melting point of their body fluids. These species also exhibit higher survival when frozen inoculatively compared with freezing at the temperature of their SCP. Moreover, they seem to be deficient of mechanisms that could prevent inoculation.90 Even some freeze-tolerant insects that do not require inoculation (rely on internal nucleators) to survive internal freezing may exploit inoculation under certain conditions. Such an example is found in the larvae of the goldenrod gall fly (E. solidaginis).19 The strategy used in this species depends on the water content of the galls. The larvae spontaneously freeze several degree Celsius below the melting point of their body fluids but are inoculated at higher temperatures at low or high gall water content, respectively.
Inoculation in freeze-intolerant insects
In contrast to the species that exploit inoculation as part of their survival strategy, the freeze-intolerant species have evolved several mechanisms to lower the risk of inoculative freezing. Some modify the composition of cuticular lipids or rely on the action of antifreeze proteins, and others spin cocoons that are relatively resistant to ice penetration.91–93
However, some of these insects, although primarily freeze-intolerant (relying on extensive supercooling with SCPs ranging from −15°C to −30°C or lower), have a capacity to tolerate internal ice, provided that freezing occurs inoculatively, just below the melting point of their body fluids.94–97 Codling moth (Cydia pomonella) is a “typical” freeze-intolerant insect. Its larvae have SCPs ranging from −15°C in summer to −26°C in winter and do not survive freezing below their SCP. However, when the larvae are put in contact with ice, they freeze at temperatures as high as −3°C. Such inoculatively frozen larvae have a considerable capacity to survive further cooling to −5°C and exhibit limited survival at temperatures as low as −15°C.97 An extreme cold tolerance is shown by the drosophilid fly Chymomyza costata. The larvae of this fly were first categorized as freeze-intolerant, based on their SCP, which varies between −15°C and −25°C.98 In subsequent studies, a tolerance to inoculative freezing was discovered, and the larvae were found to survive submersion in liquid nitrogen after inoculation and slow cooling.21,99 C. costata larvae probably represent the most complex organism known to survive in liquid nitrogen in a fully hydrated state. The possible mechanisms controlling this incredibly high cold tolerance may be the accumulation of relatively large amounts of proline and trehalose, and/or a glass-like transition that may occur in concentrated solutions of these compounds.100,101 Similar treatment, in combination with cessation of development and feeding with proline, allowed freeze-intolerant Drosophila melanogaster larvae to survive freezing at mild subzero temperatures.102
This “opportunistic” freeze-tolerance does not usually increase survival at subzero temperatures, compared with the primary strategy – supercooling. The lower lethal temperature in such species, when frozen, is, at best, equal to when they are supercooled (with at least the exception of C. costata). Many freeze-intolerant organisms may encounter conditions where even mild freeze-tolerance could increase their chance of survival. The capacity for freeze-tolerance in these species can perhaps be considered in terms of evolutionary or ecological benefit. Such adaptation may have some positive effect on population persistence when abnormal conditions occur. A mere physiological capacity with no clear ecological significance, however, cannot be ruled out either. It is possible that the apparent ability of these insects to tolerate freezing is perhaps in reality an indication of their capacity to tolerate a certain level of cellular dehydration. The amount of ice formed at a given temperature could therefore define the limits for survival. A careful analysis of cold tolerance may prove the capacity to survive inoculation to be more widespread than originally thought among insects and other organisms that primarily rely on supercooling. Further study, however, will be needed to clarify the ecological relevance and the mechanisms that underlie this capacity.
A method to test tolerance to inoculative freezing
The capacity to survive at subzero temperatures, either in a supercooled or in a frozen state, has been tested in many insects and other organisms. However, testing the capacity of apparently freeze-intolerant species to survive inoculation has mostly been neglected. Therefore, I propose a possible method and a cooling protocol to test inoculative freeze-tolerance in insects (Figure 2). This proposed experimental method is a combination and modification of the freeze-tolerance assays used by Koštál et al101 and Rozsypal et al.97 The inoculation is performed by keeping the insect in a moist environment in the presence of ice crystals. The cooling rate required to survive inoculative freezing may differ between species. Nevertheless, 0.1°C/min insured inoculation survival of both C. costata, which can survive subsequent cooling to temperatures as low as that of liquid nitrogen, and D. melanogaster, whose ability to survive in frozen and supercooled states is quite limited.101,102
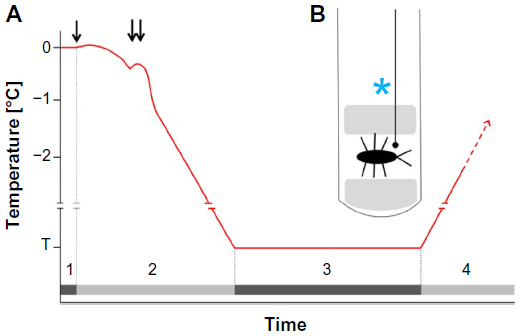 | Figure 2 A possible method to test tolerance to inoculative freezing in insects. |
Conclusion
The role of water as a “molecule of life” is well recognized; however, some of its properties and especially its interactions within organisms remain unknown. While the importance of water in keeping biological processes operating is relatively well understood, its role in the preservation of biological structures in organisms capable of extensive dehydration remains unclear.
The cause of ice nucleation in systems where no known nucleators (ie, ice nucleating proteins) can be found represents another issue that is worthy of our attention. The ice nucleation in freeze-intolerant organisms can perhaps be explained by means of homogeneous nucleation or by the presence of dust particles that are already known to act as nuclei in the atmosphere.
Over the past few decades, some freeze-intolerant insects have been shown to survive freezing of their body fluids when inoculated with ice at temperatures well above their typical SCPs. Although both the mechanism underlying this capacity and ecological significance are unknown, the ability to survive freezing upon inoculation could be relatively widespread among insects that primarily use extensive supercooling as their survival strategy.
Acknowledgments
The author thanks Vladimír Koštál (Institute of Entomology, Biology Centre ASCR, České Budejovice, Czech Republic) and Shin G Goto (Graduate School of Science, Osaka City University, Osaka, Japan) for their helpful comments on the manuscript. The author would also like to thank “Geoffrey” and Timothy Amos for language revision of the manuscript. Thanks also belong to the three anonymous referees for their helpful comments and suggestions. This work was supported by the Japan Society for Promotion of Science (JSPS).
Disclosure
The author reports no conflict of interest in this work.
References
Lee RE Jr. Insect cold-hardiness: to freeze or not to freeze. BioSci. 1989;39(5):308–313. | |
Block W. To freeze or not to freeze? Invertebrate survival of sub-zero temperatures. Funct Ecol. 1991;5(2):284–290. | |
Hawes TC, Bale JS. Plasticity in arthropod cryotypes. J Exp Biol. 2007;210(15):2585–2592. | |
Zachariassen EK. Physiology of cold tolerance in insects. Physiol Rev. 1985;65(4):799–832. | |
Wilson PW, Heneghan AF, Haymet ADJ. Ice nucleation in nature: supercooling point (SCP) measurements and the role of heterogeneous nucleation. Cryobiology. 2003;46(1):88–98. | |
Danks HV. Dehydration in dormant insects. J Insect Physiol. 2000; 46(6):837–852. | |
Ramløv H. Aspects of natural cold tolerance in ectothermic animals. Hum Reprod. 2000;15(5):26–46. | |
Holmstrup M, Westh P. Dehydration of earthworm cocoons exposed to cold: a novel cold hardiness mechanism. J Comp Physiol B. 1994; 164(4):312–315. | |
Holmstrup M, Sømme L. Dehydration and cold hardiness in the Antarctic collembolan Onychiurus arcticus Tullberg 1876. J Comp Physiol B. 1998;168:197–203. | |
Holmstrup M, Bayley M, Ramløv H. Supercool or dehydrate? An experimental analysis of overwintering strategies in small permeable arctic invertebrates. Proc Natl Acad Sci U S A. 2002;99(8):5716–5720. | |
Elnitsky MA, Hayward SAL, Rinehart JP, Denlinger DL, Lee RE Jr. Cryoprotective dehydration and the resistance to inoculative freezing in the Antarctic midge, Belgica antarctica. J Exp Biol. 2008;211(4):524–530. | |
Sørensen JG, Holmstrup M. Cryoprotective dehydration is widespread in Arctic springtails. J Insect Physiol. 2011;57(8):1147–1153. | |
Watanabe M. Anhydrobiosis in invertebrates. Appl Entomol Zool. 2006; 41(1):15–31. | |
Watanabe M, Kikawada T, Minagawa N, Yukuhiro F, Okuda T. Mechanism allowing an insect to survive complete dehydration and extreme temperatures. J Exp Biol. 2002;205(18):2799–2802. | |
Crowe JH, Oliver AE, Tablin F. Is there a single biochemical adaptation to anhydrobiosis? Integr Comp Biol. 2002;42(3):497–503. | |
Wasylyk JM, Tice AR, Baust JG. Partial glass formation: a novel mechanism of insect cryoprotection. Cryobiology. 1988;25(5):451–458. | |
Storey JM, Storey KB. Cold hardiness and freeze tolerance. In: Storey KB, editor. Functional Metabolism: Regulation and Adaptation. Hoboken: John Wiley and Sons Inc.; 2005:473–504. | |
Lundheim R. Physiological and ecological significance of biological ice nucleators. Philos Trans R Soc Lond B Biol Sci. 2002;357(1423):937–943. | |
Layne JR Jr, Lee RE Jr, Huang JL. Inoculation triggers freezing at high subzero temperatures in a freeze-tolerant frog (Rana sylvatica) and insect (Eurosta solidaginis). Can J Zool. 1990;68(3):506–510. | |
Kawarasaki Y, Teets NM, Denlinger DL, Lee RE Jr. Wet hibernacula promote inoculative freezing and limit the potential for cryoprotective dehydration in the Antarctic midge, Belgica antarctica. Polar Biol. 2014;37(6):753–761. | |
Shimada K, Riihamaa A. Cold acclimation, inoculative freezing and slow cooling: essential factors contributing to the freezing tolerance in diapausing larvae of Chymomyza costata (Diptea: Drosophilidae). CryoLetters. 1988;9:5–10. | |
Constanzo JP, Bayuk JM, Lee RE Jr. Inoculative freezing by environmental ice nuclei in the freeze-tolerant wood frog, Rana sylvatica. J Exp Zool. 1999;284(1):7–14. | |
Baker PJ, Costanzo JP, Herlands R, Wood RC, Lee RE Jr. Inoculative freezing promotes winter survival in hatchling diamondback terrapin, Malaclemys terrapin. Can J Zool. 2006;84(1):116–124. | |
Franks F. Unfrozen water: yes; unfreezable water: hardly; bound water: certainly not. An editorial note. Cryoletters. 1986;7:207. | |
Wiggins PM. Role of water in some biological processes. Microbiol Rev. 1990;54(4):432–449. | |
Wolfe J, Bryant G, Koster KL. What is “unfreezable water”, how unfreezable it is and how much is there? Cryoletters. 2002;23(3):157–166. | |
Disalvo EA, Lairion F, Martini F, et al. Structural and functional properties of hydration and confined water in membrane interfaces. BBA Biomembranes. 2008;1778(12):2655–2670. | |
Ball P. Water as an active constituent in cell biology. Chem Rev. 2008; 108(1):74–108. | |
Hong MK, Erramilli S. A dynamic role for water in biological systems. J Biol Phys. 2012;38(1):1–2. | |
Brovchenko I, Oleinikova A. Which properties of spanning network of hydration water enable biological functions? ChemPhysChem. 2008;9(18):2695–2702. | |
Block W. Water or ice? – the challenge for invertebrate cold survival. Sci Prog. 2003;86(1–2):77–101. | |
Prestrelski SJ, Tedeschi N, Arakawa T, Carpenter JF. Dehydration-induced conformational transitions in proteins and their inhibition by stabilizers. Biophys J. 1993;65(2):661–671. | |
Hoaglund-Hyzer CS, Counterman AE, Clemme DE. Anhydrous protein ions. Chem Rev. 1999;99(10):3037–3079. | |
Jarrold MF. Peptides and proteins in vapor phase. Annu Rev Phys Chem. 2000;51:179–207. | |
Klibanov AM. Improving enzymes by using them in organic solvents. Nature. 2001;409(6817):241–246. | |
Dolman M, Halling PJ, Moore BD, Waldron S. How dry are anhydrous enzymes? Measurement of residual and buried 18O-labeled water molecules using mass spectrometry. Biopolymers. 1997;41(3):313–321. | |
Kirk GL, Gruner SM, Stein DL. A thermodynamic model of the lamellar to inverse hexagonal phase transition of lipid membrane-water systems. Biochemistry. 1984;23(6):1093–1102. | |
Irwin JT, Lee RE Jr. Energy and water conservation in frozen vs supercooled larvae of the goldenrod gall fly, Eurosta solidaginis (Fitch) (Diptera: Tephritidae). J Exp Zool. 2002;292(4):345–350. | |
Storey KB, Storey JM. Freeze tolerance in animals. Physiol Rev. 1988;68(1):27–84. | |
Voituron Y, Mouquet N, de Mazancourt C, Clobert J. To freeze of not to freeze? An evolutionary perspective on the cold-hardiness strategies of overwintering ectotherms. Am Nat. 2002;160(2):255–270. | |
Elbein AD, Pan YT, Pastuszak I, Carroll D. New insights on trehalose: a multifunctional molecule. Glycobiology. 2003;13(4):17R–27R. | |
Scott P. Resurrection plants and the secrets of eternal leaf. Ann Bot. 2000;85(2):159–166. | |
Clegg JS, Seitz P, Seitz W, Hazlewood CF. Cellular responses to extreme water loss: the water-replacement hypothesis. Cryobiology. 1982;19(3):306–316. | |
Crowe LM, Crowe JH, Rudolph A, Womersley C, Appel L. Preservation of freeze-dried liposomes by trehalose. Arch Biochem Biophys. 1985;242(1):240–247. | |
Jain NK, Roy I. Effect of trehalose on protein structure. Protein Sci. 2009;18(1):24–36. | |
Crowe JH, Crowe LM, Chapman D. Preservation of membranes in anhydrobiotic organisms: the role of trehalose. Science. 1984;223(4637):701–703. | |
Grasmeijer N, Stankovic M, de Waard H, Frijlink HW, Hinrichs WLJ. Unraveling protein stabilization mechanisms: vitrification and water replacement in a glass transition temperature controlled system. Biochim Biophys Acta. 2013;1834(4):763–769. | |
Sakurai M, Furuki T, Akao K, et al. Vitrification is essential for anhydrobiosis in an African chironomid, Polypedilum vanderplanki. Proc Natl Acad Sci U S A. 2008;105(13):5093–5098. | |
Hare PD, Cress WA. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997;21(2):79–102. | |
Morgan TD, Chippendale GM. Free amino acids of the hemolymph of the southwestern corn borer and the European corn borer in relation to their diapause. J Insect Physiol. 1983;29(10):735–740. | |
Fields PG, Fleurat-Lessard F, Lavenseau L, Febvay G, Peypelut L, Bonnot G. The effect of cold acclimation and deacclimation on cold tolerance, trehalose and free amino acid levels in Sitophilus granaries and Cryptolestes ferrugineus (Coleoptera). J Insect Physiol. 1998;44(10):955–965. | |
Timasheff SN. Protein-solvent preferential interactions, protein hydration, and the modulation of biochemical reactions by solvent components. Proc Natl Acad Sci U S A. 2002;99(15):9721–9726. | |
Shimizu S, Smith DJ. Preferential hydration and the exclusion of cosolvents from protein surfaces. J Chem Phys. 2004;121(2):1148–1154. | |
Gadd GM, Chalmers K, Reed RH. The role of trehalose in dehydration resistance of Sacharomyces cerevisiae. FEMS Microbiol Lett. 1987; 48(1–2):249–254. | |
Zayed G, Roos YH. Influence of trehalose and moisture content on survival of Lactobacilus salivarius subjected to freezedrying and storage. Process Biochem. 2004;39(9):1081–1086. | |
Rothschild LJ, Mancinelli RL. Life in extreme environments. Nature. 2001;409(6823):1092–1101. | |
Jaenicke R, Böhm G. The stability of proteins in extreme environments. Cur Opin Struct Biol. 1998;8(6):738–748. | |
Konings WN, Albers SV, Koning S, Driessen AJM. The cell membrane plays a crucial role in survival of bacteria and archaea in extreme environments. Antonie van Leeuwenhoek. 2002;81(1–4):61–72. | |
Clegg JS. Cryptobiosis – a peculiar state of biological organization. Comp Biochem Physiol B Biochem Mol Biol. 2001;128(4):613–624. | |
Matsumoto M, Saito S, Ohmine I. Molecular dynamics simulation of the ice nucleation and growth process leading to water freezing. Nature. 2002;416(6879):409–413. | |
Lee RE Jr, Constanzo JP. Biological ice nucleation and ice distribution in cold-hardy ectothermic animals. Annu Rev Physiol. 1998;60:55–72. | |
Lee RE Jr. Principles of insect low temperature tolerance. In: Lee RE Jr, Denlinger DL, editors. Insects at Low Temperature. New York: Chapman and Hall; 1991:17–46. | |
Clausse D, Bouabdillah D, Cochet N, Luquet MP, Pulvin S. Ice crystallization induced by silver iodide and bacteria in microsize droplets dispersed within emulsions. Pure Appl Chem. 1991;63(10):1491–1494. | |
Duman JG, Morris JP, Castellino FJ. Purification and composition of an ice nucleating protein from queens of the hornet, Vespula maculata. J Comp Physiol B. 1984;154(1):79–83. | |
Neven LG, Duman JG, Low MG, Sehl LC, Castellino FJ. Purification and characterization of an insect hemolymph lipoprotein ice nucleator: evidence for the importance of phosphatidylinositol and apolipoprotein in the ice nucleator activity. J Comp Physiol B. 1989;159(1):71–82. | |
Gurian-Sherman D, Lindow SE. Bacterial ice nucleation: significance and molecular basis. FASEB J. 1993;7(14):1338–1343. | |
Bale JS. Insect cold hardiness: a matter of life and death. Eur J Entomol. 1996;93(3):369–382. | |
Warren G, Corotto L, Wolber P. Conserved repeats in diverged ice nucleation structural genes from two species of Pseudomonas. Nucleic Acids Res. 1986;14(20):8047–8060. | |
Kajava AV, Lindow SE. A model of the three-dimensional structure of ice nucleation proteins. J Mol Biol. 1993;232(3):709–717. | |
Sømme L. The physiology of cold hardiness in terrestrial arthropods. Eur J Entomol. 1999;96(1):1–10 | |
Zachariassen KE, Kristiansen E, Pedersen SA, Hammel HT. Ice nucleation in solutions and freeze-avoiding insects – homogeneous or heterogeneous? Cryobiology. 2004;48(3):309–321. | |
Cantrell W, Heymsfield A. Production of ice in tropospheric clouds: a review. Bull Amer Meteor Soc. 2005;86(6):795–807. | |
Murray BJ, O’Sullivan D, Atkinson JD, Webb ME. Ice nucleation by particles immersed in supercooled cloud droplets. Chem Soc Rev. 2012;41(19):6519–6554. | |
Zimmermann F, Weinbruch S, Schütz L, et al. Ice nucleation properties of the most abundant mineral dust phases. J Geophys Res. 2008;113:D23204. | |
Fornea AP, Brooks SD, Dooley JB, Saha A. Heterogeneous freezing of ice on atmospheric aerosols containing ash, soot, and soil. J Geophys Res. 2009;114:D13201. | |
Marcolli C, Gedamke S, Peter T, Zobrist B. Efficiency of immersion mode ice nucleation on surrogates of mineral dust. Atmos Chem Phys. 2007;7:5081–5091. | |
Mugnano JA, Lee RE Jr, Taylor RT. Fat body cells and calcium phosphate spherules induce ice nucleation in the freeze-tolerant larvae of the gall fly Eurosta solidaginis (Diptera, Tephritidae). J Exp Biol. 1996; 199(2):465–471. | |
Lee RE Jr, Strong-Gunderson JM, Lee MR, Grove KS, Riga TJ. Isolation of ice nucleating active bacteria from insects. J Exp Zool. 1991;257(1):124–127. | |
Worland MR, Block W. Ice-Nucleating bacteria from the guts of two Sub-Antarctic beetles, Hydromedion sparsutum and Perimylops antarcticus (Perimylopidae). Cryobiology. 1999;38(1):60–67. | |
Dillon RJ, Dillon VM. The gut bacteria of insects: nonpathogenic interactions. Annu Rev Entomol. 2004;49:71–92. | |
Sømme L, Block W. Cold hardiness of Collembola at Signy Island, maritime Antarctic. Oikos. 1982;38(2):168–176. | |
Worland MR, Leinaas HP, Chown SL. Supercooling point frequency distributions in Collembola are affected by moulting. Funct Ecol. 2006;20(2):323–329. | |
Hiiesaar K, Williams I, Luik A, et al. Factors affecting cold hardiness in the small striped flea beetle, Phyllotreta undulata. Entomol Exp Appl. 2009;131(3):278–285. | |
Duman JG. Antifreeze and ice nucleator proteins in terrestrial arthropods. Annu Rev Physiol. 2001;63:327–357. | |
Barrett J. Thermal hysteresis proteins. Int J Biochem Cell Biol. 2001;33(2):105–117. | |
Clark MS, Worland MR. How insects survive the cold: molecular mechanisms – a review. J Comp Physiol B. 2008;178(8):917–933. | |
Davies PL, Baardsnes J, Kuiper MJ, Walker VK. Structure and function of antifreeze proteins. Phil Trans R Soc Lond B. 2002;357(1423):927–935. | |
Tursman D, Duman JD, Knight CA. Freeze tolerance adaptations in the centipede, Lithobius forficatus. J Exp Zool. 1994;268(5):347–353. | |
Gehrken U, Southon TE. Supercooling in a freeze-tolerant larva, Tipula sp. J Insect Physiol. 1992;38(2):131–137. | |
Layne JR Jr, Edgar CL, Medwith RE. Cold hardiness of the Wooly Bear caterpillar (Pyrrharctica isabella Lepidoptera: Arctiidae). Am Midl Nat. 1999;141(2):293–304. | |
Olsen TM, Sass SJ, Li N, Duman JG. Factors contributing to seasonal increases in inoculative freezing resistance in overwintering fire-colored beetle larvae Dendroides canadensis (Pyrochroidae). J Exp Biol. 1998;201(10):1585–1594. | |
Gehrken U. Inoculative freezing and thermal hysteresis in the adult beetles Ips acuminatus and Rhagium inquisitor. J Insect Physiol. 1992;38(7):519–524. | |
Danks HV. The roles of insect cocoons in cold conditions. Eur J Entomol. 2004;101(3):433–437. | |
Fields PG, McNeil JN. Possible dual cold-hardiness strategies in Cisseps fulvicollis Lepidoptera Arctiidae. Can Ent. 1986;118(12):1309–1311. | |
Gehrken U, Strømme A, Lundheim R, Zachariassen KE. Inoculative freezing in overwintering tenebrionid beetle, Bolitophagus reticulatus Panz. J Insect Physiol. 1991;37(9):683–687. | |
Koštál V, Havelka J. Diapausing larvae of the midge Aphidoletes aphidimyza (Diptera: Cecidomyiidae) survive at subzero temperatures in a supercooled state but tolerate freezing if inoculated by external ice. Eur J Entomol. 2000;97(3):433–436. | |
Rozsypal J, Koštál V, Zahradnícková H, Šimek P. Overwintering strategy and mechanisms of cold tolerance in the codling moth (Cydia pomonella). PLoS ONE. 2013;8(4):e61745. | |
Enomoto O. Larval diapause in Chymomyza costata (Diptera: Drosophilidae) II. Frost avoidance. Low Temp Sci B. 1981;39:31–39. | |
Moon I, Fujikawa S, Shimada K. Cryopreservation of Chymomyza larvae (Diptera: Drosophilidae) at −196°C with extracellular freezing. CryoLetters. 1996;17:105–110. | |
Shimada K, Riihamaa A. Cold-induced freezing tolerance in diapausing and non-diapausing larvae of Chymomyza costata (Diptera: Drosophilidae) with accumulation of trehalose and proline. CryoLetters. 1990;11:243–250. | |
Koštál V, Zahradníčková H, Šimek P. Hyperprolinemic larvae of the drosophilid fly, Chymomyza costata, survive cryopreservation in liquid nitrogen. Proc Natl Acad Sci U S A. 2011;108(32):13041–13046. | |
Koštál V, Šimek P, Zahradníčková H, Cimlová J, Štětina T. Conversion of the chill susceptible fruit fly larva (Drosophila melanogaster) to a freeze tolerant organism. Proc Natl Acad Sci U S A. 2012;109(9):3270–3274. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.