Back to Journals » Cancer Management and Research » Volume 13
The Preliminary Study for Postoperative Radiotherapy Survival Associated with RUNX3 and TLR9 Expression in Lung Cancer
Authors Gao C, Qiao T ![]() , Yuan S, Zhuang X
, Yuan S, Zhuang X ![]()
Received 11 February 2021
Accepted for publication 23 April 2021
Published 4 June 2021 Volume 2021:13 Pages 4497—4507
DOI https://doi.org/10.2147/CMAR.S305452
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ahmet Emre Eşkazan
Caixia Gao, Tiankui Qiao, Sujuan Yuan, Xibing Zhuang
Jinshan Hospital Center for Tumor Diagnosis & Therapy, Shanghai, 201508, People’s Republic of China
Correspondence: Tiankui Qiao
Jinshan Hospital of Fudan University, No. 1508 Longhang Road, Jinshan District, Shanghai, 201508, People’s Republic of China
Tel +86 189 3077 8786
Email [email protected]
Background: Many studies have reported that the inflammatory immune response related to TLR9 signaling activation participates in tumor development and affects the treatment outcome. RUNX3 functions as a tumor suppressor by regulating DNA methylation. RUNX3 protein plays an important role in TGF-β signaling pathway that is involved in tumor growth inhibition and apoptosis. At present, radiotherapy is still an important treatment in lung cancer, which induces immune response and affects the therapeutic outcome. The role of TLR9 signaling activation and RUNX3 in this process is not clear.
Methods: In this study, we investigated the expression of TLR9 in tumor and RUNX3 in surrounding tissues by immunohistochemical methods and analyzed the relationship on postoperative survival in lung cancer.
Results: We found that the high expression of TLR9 was the risk factor in postoperative survival of lung cancer with no difference in lifetime. The high expression of RUNX3 in lung cancer with TLR9 signaling activation was in favor of progression-free survival and overall survival in postoperative radiotherapy. It suggested that RUNX3 played an important role in lung cancer radiotherapy. In order to determine the effect of RUNX3 in lung cancer radiation with TLR9 signaling activation, we introduced 5-Aza-2ʹ-deoxycytidine (5-Aza-CdR) and exposed lung cancer A459 cells repeatedly. The high expression of RUNX3 especially RUNX3-B in cells treated with 5-Aza-CdR was observed. We examined that 5-Aza-CdR induced more cell blocking in G2/M phase in combining irradiation.
Conclusion: The result implied that it was feasible to improve radiosensitivity of lung cancer with TLR9 signaling activation by increasing RUNX3 expression, and 5-Aza-CdR was an option in this process.
Keywords: TLR9 signaling activation, RUNX3 expression, 5-Aza-CdR, radiotherapy, lung cancer
Introduction
Although radiotherapy is one of the most effective treatments for non-small-cell lung carcinoma, local recurrence and metastasis are the main reasons for the failure of clinical treatment. There is an urgent need to explore the relationship of radiotherapy and tumor development for lung cancer treatment currently.
TLR9 is a member of toll-like receptor family, binding DNA in bacteria and viruses. TLR9 is a pattern recognition receptor connecting innate and adaptive immune systems.1 The researchers have demonstrated that TLR9 is expressed in multiple cancer cells.2 The signaling activation causing inflammatory cytokine response such as TNFa, IL-1, IL-6, IL-8 promotes tumor proliferation and microenvironment development, contributing to a huge impact on clinical tumor treatment.3,4 Ren et al reported that the activation of TLR9 signal in lung cancer cells upregulated inflammatory cytokines, MMP-2, chemokines, and immunosuppressive factor (IL-10) levels, leading to significant enhancement of tumor proliferation, invasion and metastasis.5,6 CpG-oligodeoxynucleotide induced TLR9 activation increasing prostate cancer metastasis has been confirmed.7 And the correlation of TLR9 high expression and poor tumor grading or later tumor staging had been reported in prostate cancer, glioma and esophagus tumor.8–10 RUNX3 is a protein-coding gene that functions as a tumor suppressor and regulates cell development, apoptosis, signaling transduction, etc.11,12 Recently it was reported that RUNX3 has a regulatory function on tumor immunity, whose upregulation can facilitate the expression of T-bet and promote Th1 immune response.13–15 Irradiation triggering signaling cascade and leading to upregulation of more than 60 genes involved in cell cycle regulation, DNA damage and repair, apoptosis, oncogenes and tumor suppressor genes, cell metabolism, etc have reported.16 And how TLR9 signal activation and RUNX3 expression influence radiotherapy outcome have rarely been reported. In this study, we surveyed the effect of TLR9 in tumor tissues and RUNX3 in adjacent carcinoma tissues on postoperative radiotherapy survival in lung cancer. 5-Aza-2ʹ-deoxycytidine (5-Aza-CdR) is one of the inhibitors of DNA methyltransferase. Studies have shown that epigenetic changes of DNA methylation and histone modification affect tumor development and treatment.17 It is believed that DNA methyltransferase could inhibit transcription of target genes by forming transcription repressors with histone methyltransferase at promoter region.18 The application of 5-Aza-CdR and the analog 5-azacitidine in tumor chemoradiotherapy has been reviewed.19,20 We had determined the radiation resistance in lung cancer A549 cells exposed repeatedly in preliminary study, which TLR9 expression upregulated.21 In the study we would investigate the role of RUNX3 in 5-Aza-CdR combined radiotherapy for cells with TLR9 signal activation and further elucidated that it was significant in raising RUNX3 expression for lung cancer radiotherapy.
Materials and Methods
Clinical Information Related to the Cases
Sixty-three patients were enrolled in the study for tumor resection and postoperative chemoradiotherapy in JinShan Hospital of Fudan University from March 1, 2010 to December 31, 2011. Fifty-two males and 11 females were selected. The oldest patient was 76 years old, and the youngest was 36 years old. The median age was 63 years. Forty-two cases had a smoking history. All patients received a preoperative pathological diagnosis. Sixty-one patients accepted lobectomy and lymph node dissection while two patients took unilateral pneumonectomy. Thirty-six squamous carcinomas apart from three sarcomatoid carcinomas and 27 adenocarcinomas, including three adeno-squamous carcinomas were diagnosed pathologically after tumor resection. There were 28 cases of mediastinal lymph node metastasis. The postoperative tumor tissues of 63 patients were selected as the effective specimens for this experiment. This was conducted in accordance with the Declaration of Helsinki. The study was approved by the Ethics Committee of Jinshan Hospital, Fudan University (Shanghai, China), together with the confirmation that all patients whose tissue was used provided written informed consent.
Thirty-six patients received radiotherapy within three months postoperatively. The conditions for postoperative radiotherapy were as follows: positive edge-cutting and/or mediastinal lymph node metastasis, KPS score ≥85 and FEV 1.0 of pulmonary function ≥80%. The signature confirmation for postoperative radiotherapy was signed. Intensity-modulated radiation therapy (IMRT) was adopted for postoperative radiotherapy. The target drawings referred to the NCCN non-small-cell lung cancer clinical practice guidelines. Clinical target volume (CTV) put on 5–8 mm externally as the planning target area (PTV). Ninety-five percent isodose line covering the 100% volume of PTV was approved. The dose limits of dangerous organs was as follows: the side lung V30 ≤25%, total lung V20 ≤25%, mean dose of the side lung ≤18 Gy, heart maximum dose ≤45 Gy, spinal cord ≤45 Gy. The plan was administrated by 6 MV X-ray for the total dose 50 Gy/25 Fx, adding up to 56–60 Gy/28-30 Fx for the patient's positive surgical margin. Thirty-six patients completed postoperative radiotherapy with no serious side effects.
The follow-up began three months after surgical treatment and/or chemoradiotherapy and ended in February 2015. It was once every three months for two years and every six months for two years. The median for follow-up was 38 months. The time from the first visit to tumor recurrence was determined as progressive-free survival (PFS). The time from the visit ending or death in follow-up was defined as overall survival (OS). Forty-eight patients died of lung cancer and one non-neoplastic death. Twelve patients were alive except for four survival with tumor recurrence and two lost. The ratio of follow-up was 96.8%. The outpatient booking for follow-up was preferred, letter or telephone visit as a supplement.
Expression of TLR9 and RUNX3 in Cancer Tissues
The wax blocks of lung cancer tissues were cut into sections of 4 µm, fixed and labeled. After being deparaffinized, hydrated and antigen retrieval, the sections were treated with 3% hydrogen peroxide and 1% FBS. Diluted primary antibody (Rabbit Anti-TLR9 polyclonal antibody 1:1000 and Rabbit Anti-RUNX3 monoclonal antibody 1:1000, Abcam, US) was added and incubated at 4 °C overnight. HRP-Polymer Antimouse Immunohistochemistry (IHC) Kit (KIT-5930 Maixin Biology, Fuzhou, China) was incubated for 30 min at room temperature, followed by diaminobenzidine for color development. Finally the sections were counterstained with hematoxylin and mounted with Permount. The scores were determined by the proportion of stained cells and intensity. The cells binding TLR9 protein in tumor tissues and RUNX3 protein in paracarcinoma tissues was counted respectively. The proportion of stained cells was graded as follows: 0 (no stained cells), 1 (10% stained cells), 2 (10–50% stained cells) and 3 (50% stained cells). The intensity of the stain was recorded on a scale of 0 (no staining), 1 (weak staining, light yellow), 2 (moderate staining, yellowish brown) and 3 (strong staining, brown). The staining index was calculated as follows: staining index = staining intensity + score of stained cells. Tissue sections with staining index over 4 were defined as high expression and staining index less than 4 as low expression.
Preparation for Lung Cancer A549 Cell with TLR9 Signal Activation and the Experimental Groups
Human lung cancer A549 cell line from Shanghai Tumor Hospital, Fudan University (Shanghai, People’s Republic of China) was approved by the Ethics Committee of Jinshan Hospital, which was used for the preparation of the target cell line based on our previous study. A549 cell in logarithmic growth was exposed 6 MV X-ray, source to surface distance (SSD=100 cm) irradiation with dose rate 2 Gy/min and once 6.37 Gy for 10 exposures.21 The irradiation was performed once again until 63.7 Gy. And this cell line was designated as lung cancer A549 cell with high-TLR9 expression (A549high-TLR9). 5-Aza-CdR (Merck Drugs & Biotechnology, Sigma-Aldrich, Germany) was prepared for experimental cells in 0.5, 1.0, 2.0, 5.0 and 10 μmol/L, respectively. The absorbance was measured by microplate spectrophotometer at 450 nm. According the measurement, 5.0 μmol/L 5-Aza-CdR and 10 μg/mL chloroquine (CQ) were adopted for the treatment in experimental cells.
The cells were divided into six groups: A549 group, A549high-TLR9 group, A549high-TLR9/IR (cell irradiation) group, A549high-TLR9+CQ/IR (chloroquine combining irradiation) group, A549high-TLR9+5-Aza-CdR/IR (5-Aza-CdR combining irradiation) group, A549high-TLR9+5-Aza-CdR (5-Aza-CdR treatment) group. The cells were administrated in serum-free medium with 10 μg/mL chloroquine and 5 μmol/L 5-Aza-CdR respectively before IR 30 min.
Detection of TLR9 and RUNX3 Protein by Western blotting
After cell disruption and centrifugation, protein extracts concentration were determined by the BCA method and separated by 10% SDS-PAGE, transferred to polyvinylidene difluoride (PVDF) membranes (EMD Millipore, Billerica, MA, USA). The membrane was blocked with 10% nonfat dry milk in Tris-buffered saline (TBS) containing 0.1% Tween-20 (TBS-T) for 2.5 h at room temperature and then incubated with the following primary antibodies at 4°C overnight: Rabbit TLR9 monoclonal antibody 1:5000 (Abcam, USA), Rabbit RUNX3 monoclonal antibody 1:1000 (Abcam), Mouse GAPDH monoclonal antibody 1:3500 (Proteintech, USA). Membranes were washed with TBS-T three times and probed with secondary antibodies for two hours at room temperature. Protein bands were visualized after incubation with chemiluminescence HRP substrate (EMD Millipore).
Cycle Distribution of Cells in Experimental Groups
The cells were exposed with 6 MV X-ray, a single dose of 10 Gy. The cell culture was replaced once after irradiation 24 h. The cells irradiated 0 h, 12 h, 24 h, 36 h, 48 h were harvested and fixed with 75% ethanol at 4°C overnight, then incubated with 500 μL propidium iodide/RNase staining buffer (BD Biosciences) at room temperature for 15 min. After centrifugation and PBS washing, the cell cycle reagent PI (50 mg/mL) was added to suspended cells avoiding light at 4°C for 20 min. The cell cycle was determined by flow cytometer.
Statistical Analysis
SPSS 17.0 statistical software (SPSS, Inc., Chicago, IL, USA) was used in the study. The one-way variance analysis of TLR9 and RUNX3 and clinicopathological features was tested by chi-squared test. The risk assessment of factors on postoperative survival of lung cancer was measured by logistic regression. The prediction of factors associated on postoperative radiotherapy survival was examined by Cox regression. The life tables were used for calculating the survival time related to TLR9 and RUNX3. The measurement data showed on x±s and analysis of variance was adopted in comparison of the groups. The statistical figures were made by GraphPad Prism 5.0 software. P<0.05 was regarded as statistically significant.
Results
The Relationship of TLR9 and RUNX3 and Postoperative Radiotherapy Survival in Lung Cancer
The expression of TLR9 in tumor tissues and RUNX3 adjacent to cancer was shown in Figure 1. Forty-three and 28 of 63 patients presented positive expression of TLR9 and RUNX3 respectively. The relationship of TLR9 and RUNX3 and clinicopathological characteristics by one-way variance analysis was presented in Table 1. The results displayed that the high expression of TLR9 in cancer tissues emerged in later T stage significantly (χ2=8.620, P=0.013), which the high expression of RUNX3 in paracarcinoma tissues was likely to in earlier T stage (χ2=5.920, P=0.052). RUNX3 expression in non-squamous carcinoma showed statistic difference, the chi-squared value being 4.200 and P=0.040. The expression of TLR9 and RUNX3 in tumor differentiation, N staging, and postoperative radiotherapy had no distinction.
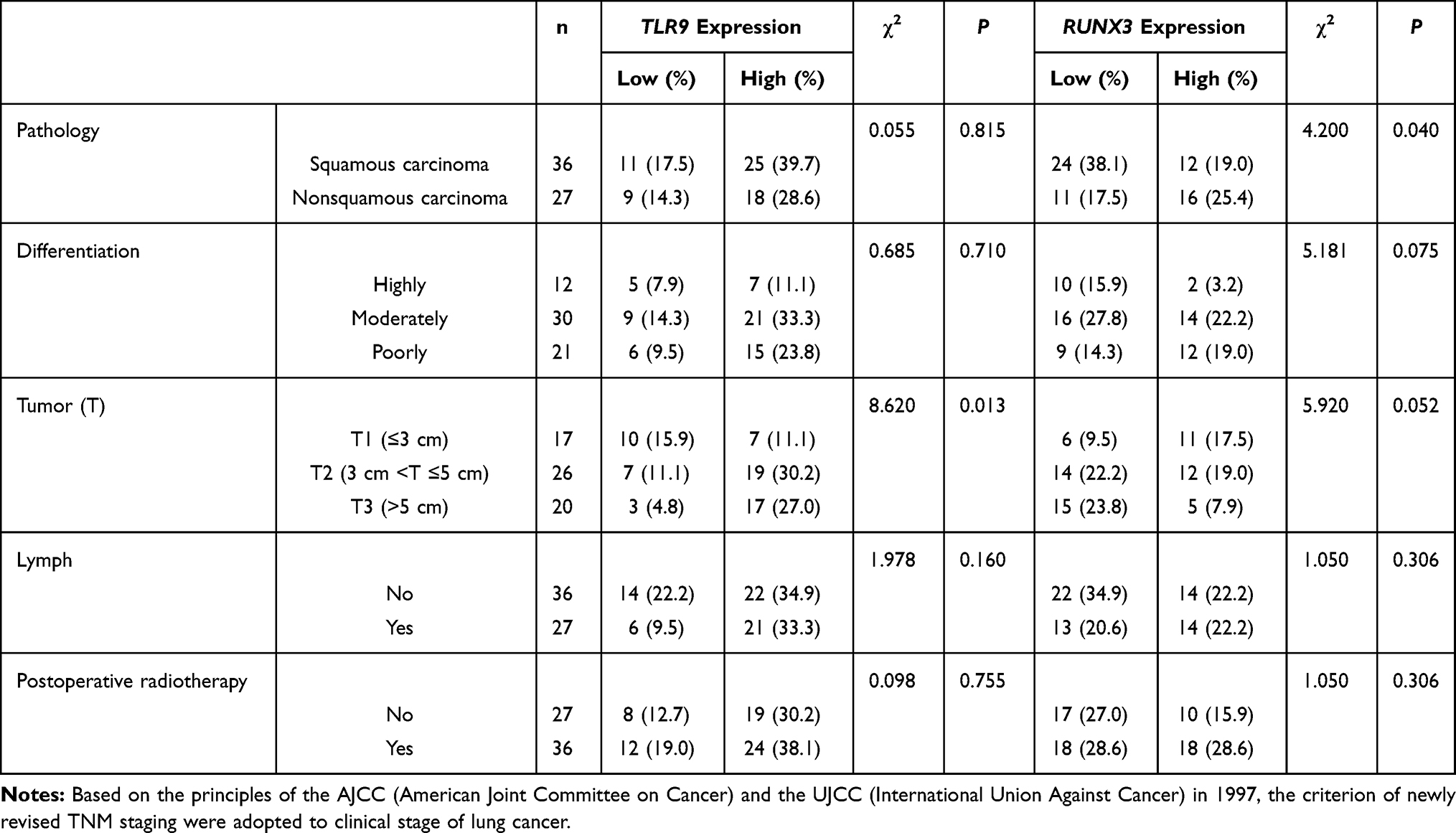 |
Table 1 The One-way Analysis of Variance on TLR9 and RUNX3 Expression in Lung Cancer Tissues |
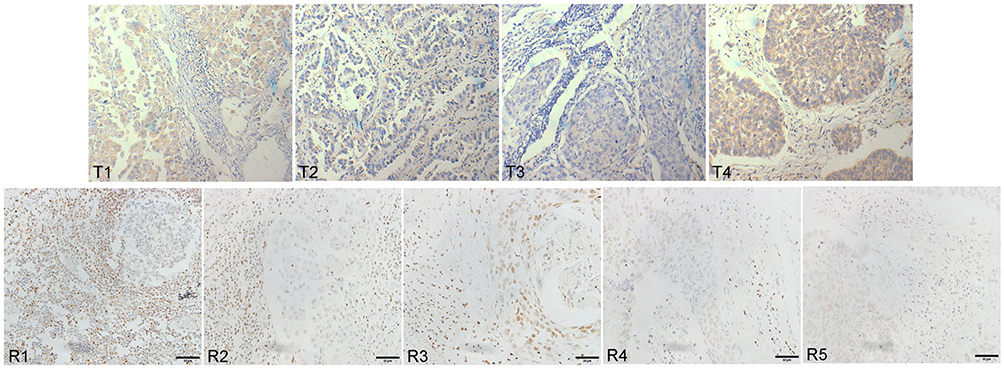 |
Figure 1 The expression of TLR9 and RUNX3 in lung cancer tissues (immunohistochemical SP method). Expression of TLR9 in tumor tissues (T1-T4) and RUNX3 in adjacent to cancer (R1-R5) was examined with DAB magnification 400Ẋ. Compared with normal bronchoalveolar cells, tumor cells became larger in volume and abnormally clumped together. TLR9 protein stained yellow or brown distributed equality in cytoplasm of tumor cells with unusual karyomegaly, multileaf nuclear and nuclear membrane thickened, etc. RUNX3 protein stained yellow or brown granular distributed in nucleus. |
Then we conducted the risk assessment on TLR9, RUNX3 and above clinical features for postoperative survival in lung cancer by logistic regression, shown in Table 2. The result revealed that the relative risk value associated with tumor T staging and TLR9 expression in the model was 2.716 and 5.057, respectively. It suggested that the high expression of TLR9 was harmful for postoperative survival in lung cancer with the P-value 0.019 significant. The median time of PFS and OS associated with TLR9 expression was shown in Table 2, with no difference in postoperative or radiotherapy patients. The cumulative survival ratio on TLR9 expression in postoperative or radiotherapy patients was exhibited in Figure 2. Subsequently the median survival time of PFS and OS in postoperative radiotherapy associated with TLR9 and RUNX3 expression was analyzed and shown in Table 3. The statistical value of PFS and OS in postoperative radiotherapy related RUNX3 expression and negative TLR9 was 1.905 and 4.224, respectively, P=0.168 and 0.040. With positive TLR9 it was 6.097 and 6.521, respectively, P=0.014 and 0.011 significantly, as shown in Figure 3. This result implied that the patients with TLR9 signaling activation was a survival benefit from postoperative radiotherapy in RUNX3 high expression. With Cox regression for PFS on lung cancer radiotherapy, the risk evaluation of tumor differentiation, T staging and RUNX3 in equation in Table 3 was statistic significance, P=0.026, 0.055 and 0.001, respectively. The relative risk was 2.109, 1.861 and 0.178 with 95%CI for Exp(B) (1.093–4.071), (0.987–3.509) and (0.067–0.475). The result illuminated that the high expression of RUNX3 was helpful for postoperative radiotherapy survival in lung cancer. It was meaningful for upregulating RUNX3 expression to improve radiotherapy outcome in lung cancer.
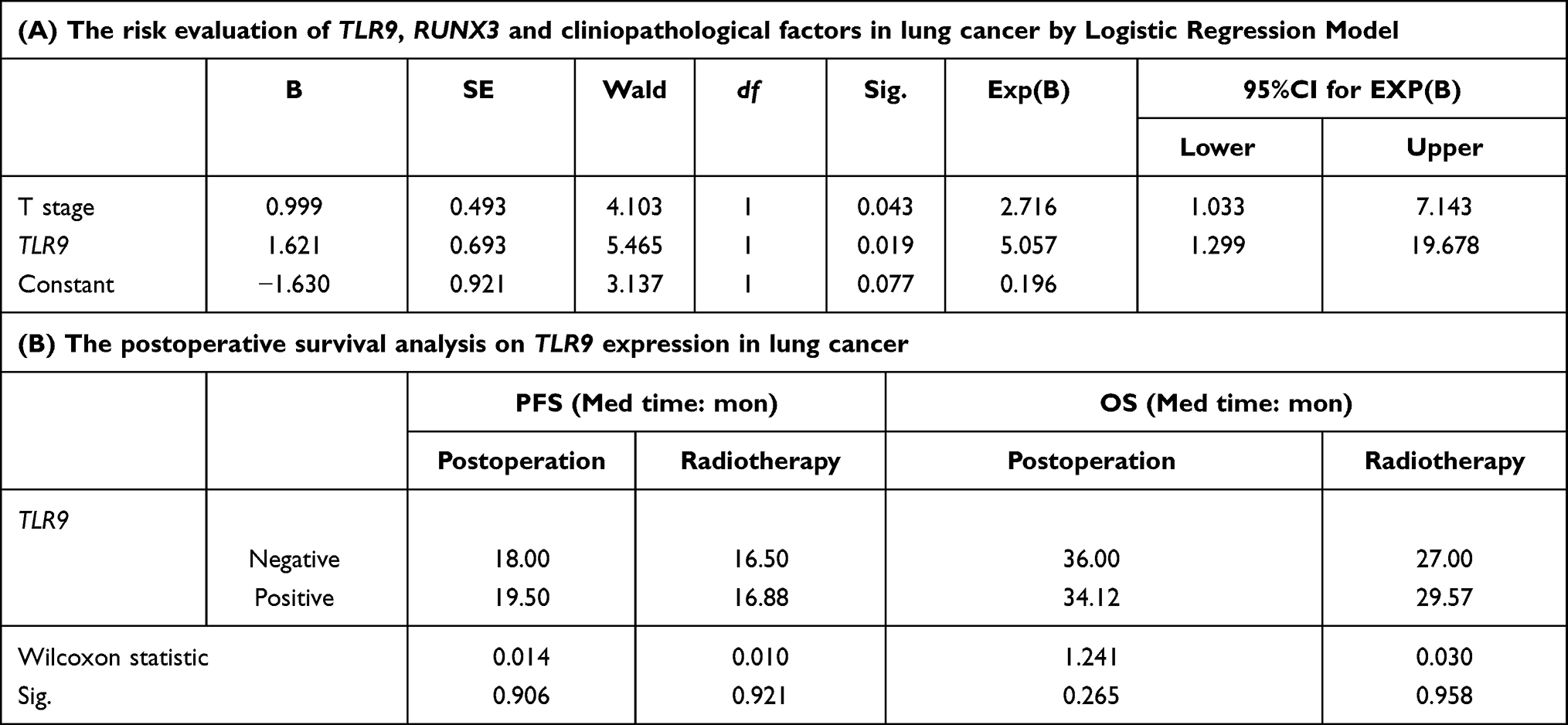 |
Table 2 The Correlation of TLR9 Expression in Cancer Tissues and Postoperative Survival |
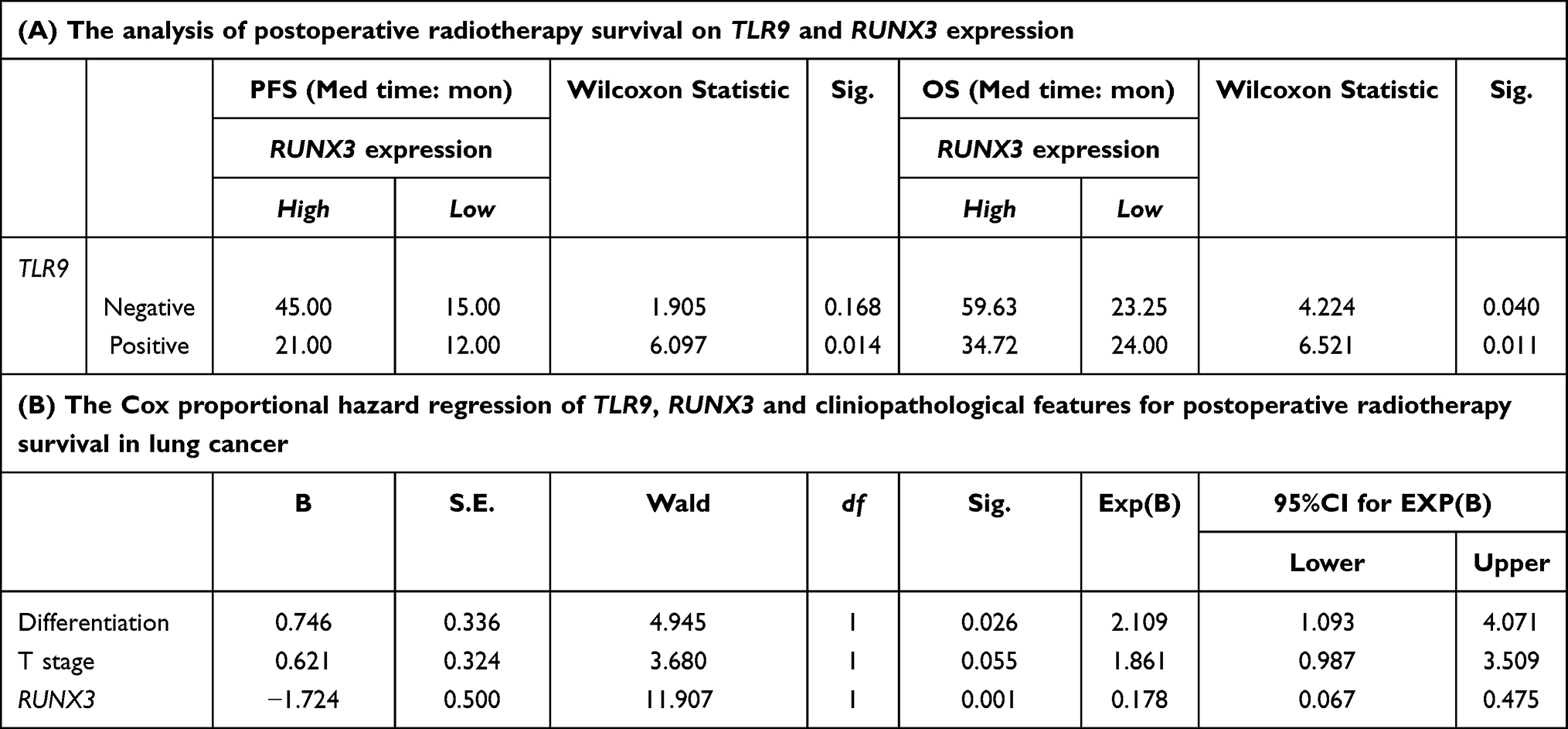 |
Table 3 The Effect of TLR9 and RUNX3 Expression on Postoperative Radiotherapy Survival of Lung Cancer |
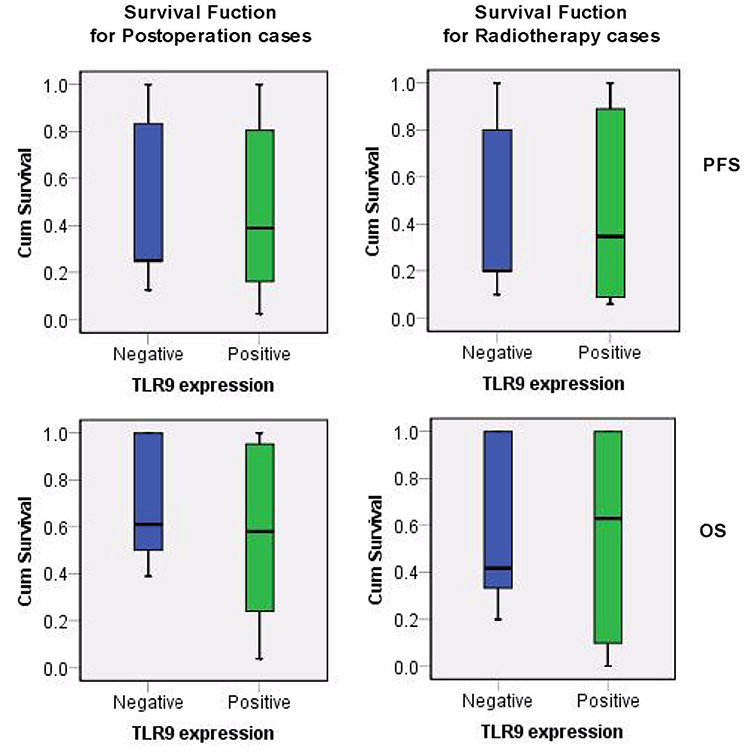 |
Figure 2 The survival function of lung cancer postoperation related TLR9 expression. X axis was identified for TLR9 status, Y axis indicated the cumulative survival of PFS and OS on lung cancer postoperation and radiotherapy respectively. Note: PFS and OS refer to progression-free survival and overall survival respectively. |
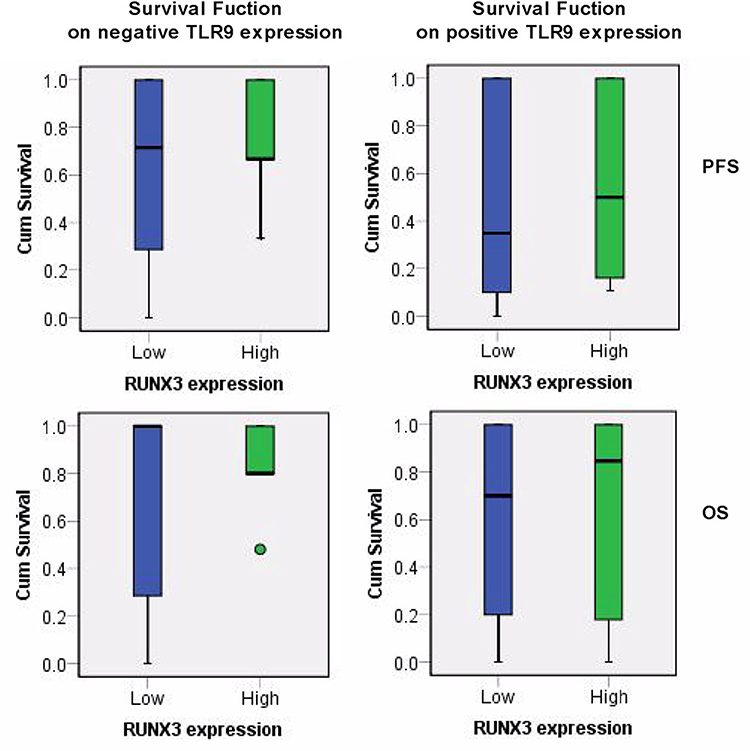 |
Figure 3 The survival function of lung cancer radiotherapy related TLR9 and RUNX3 expression. X axis was identified for RUNX3 status, Y axis indicated the cumulative survival of PFS and OS on lung cancer radiotherapy with negative and positive TLR9 expression. Note: PFS and OS refer to progression-free survival and overall survival respectively. |
RUNX3 Expression in Experimental Cells Before and After Radiotherapy
In order to demonstrate the effect of RUNX3 in lung cancer radiotherapy, the levels of TLR9 and RUNX3 protein in experiment cells was detected by Western blotting (shown in Figure 4). Based on RUNX3 antibody instructions and double-band pattern, RUNX3-A and RUNX3-B standing for the band with larger and smaller molecular weight, respectively were adopted to data analysis in the study. The results displayed the levels of RUNX3 protein in 5-Aza-CdR treatment cells was increased especially RUNX3-B before irradiation. Compared with A549high-TLR9 cells, RUNX3-B high expression in 5-Aza-CdR treatment cells was significance, with the t value 3.785, P<0.05. The levels of RUNX3 protein in cells postirradiation were decreased with no difference. TLR9 expression in A549high-TLR9 and A549 cells was statistical significance, t=4.081 and P<0.01. It was the explanation for TLR9 signal activation in A549 cell exposure repeatedly.
 |
Figure 4 RUNX3 and TLR9 expression in cells before and after radiotherapy. The levels of RUNX3 and TLR9 protein in experimental cells were detected by Western blotting. RUNX3-A and RUNX3-B standard for the larger and smaller molecular weight band separately. The levels of TLR9 protein in the A549high-TLR9 cell was significantly more than in the A549 cell, t=4.081 and P<0.01. It indicated that TLR9 signaling activated in A549 cells exposure repeatedly. The result displayed that RUNX3 protein especially RUNX3-B was increased in 5-Aza-CdR treatment cells. In comparison with the A549high-TLR9 cell, the increasing levels of RUNX3-B protein was significant, with the t value 3.785 and P<0.05. The decreased levels of RUNX3 protein in cells after irradiation was observed with no difference. Abbreviations: IR, irradiation; A549high-TLR9 cell, A549 cell with TLR9 signal activation; 5-Aza-CdR, 5-Aza-2ʹ-deoxycytidine; CQ, chloroquine. Notes: *t=3.785, P<0.05; **t=4.081, P<0.01. |
Cycle Distribution Related RUNX3 Upregulation in Lung Cancer Radiotherapy
The distribution of the cell cycle in experiment was tested, shown in Figure 5A. Compared with the cell irradiation only, the cell proportion in G2/M and G1/G0 in A549high-TLR9+5-Aza-CdR/IR group was 44.8±1.76 and 45.73±1.67, respectively with the t values 14.15 and 13.52, P<0.001 equally. There was no change in cell distribution in 5-Aza-CdR treatment and the irradiation only group. The fewer G2/M cells and more G1/G0 cells in the A549high-TLR9+CQ/IR group than that in A549high-TLR9/IR group were examined. Furthermore we examined 5-Aza-CdR induced cells blocking in G2/M with maximization at irradiation 24 h and maintained for 36 h and 48 h after radiotherapy, as shown in Figure 5B. The distribution of S cells in chloroquine combining irradiation was more than in the A549high-TLR9/IR group at 12 h with the t values 11.04 and P<0.001. Meanwhile we saw fewer G2/M cells and many more G1/G0 cells in A549high-TLR9+CQ/IR group than the irradiated cells alone.
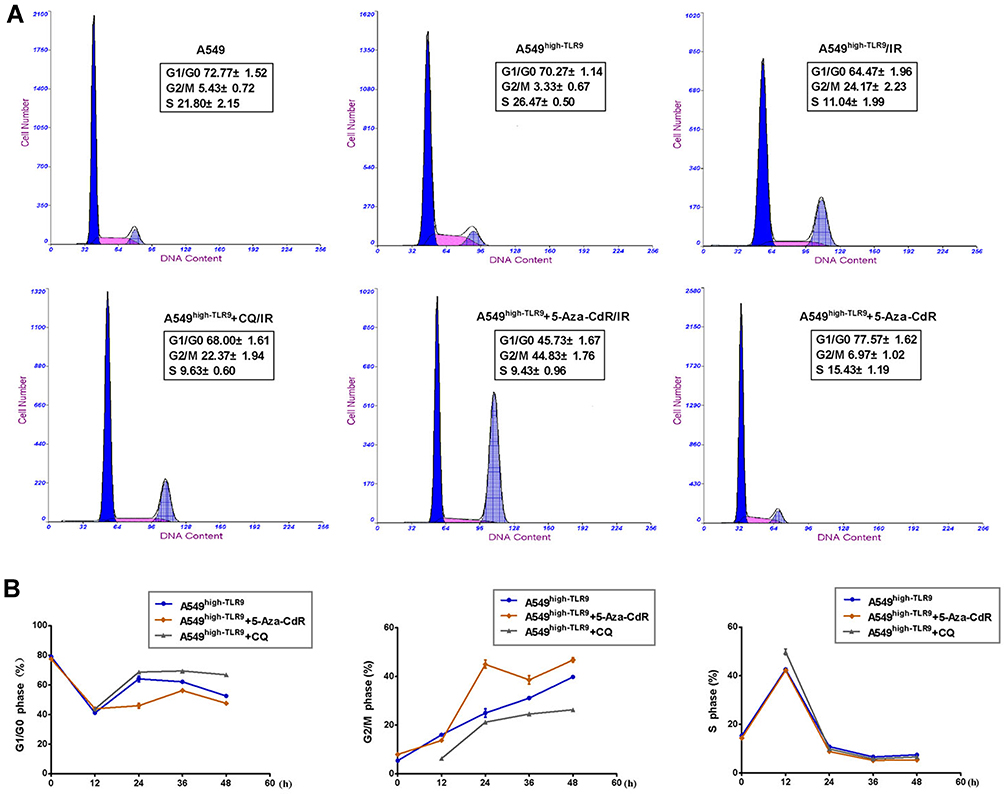 |
Figure 5 Comparison of cell distribution on experimental cells before and after radiotherapy. The cycle distribution in experimental cells before and after radiotherapy (A). The proportion of G2/M cells in 5-Aza-CdR treatment at irradiation 24 h was 44.83±1.76, twice that of solely irradiated cells significantly, t=15.52 and P<0.001. It maintained for irradiation 36 h and 48 h (B). The G1/G0 cells in 5-Aza-CdR adjuvant radiotherapy remained low for irradiation 48 h. The cells ratio in S phase in chloroquine combining radiotherapy at irradiation 12 h was 15.43±1.19, which was significantly more than the solely irradiated cells, P<0.001. The result was that after irradiation more cells kept in G2/M phase in 5-Aza-CdR treatment group as well more cells in S phase in chloroquine combining radiotherapy. |
Discussion
Radiotherapy is a common clinical treatment for lung cancer patients. TLR9 is a transmembrane protein. It is activated by binding unmethylated CpG sequences in DNA molecules to TIR domain.22,23 The researchers have demonstrated that irradiation leads to DNA damage and cells debris and proteins associated with hypoxia and necrosis activates TLR9 signal cascades by endogenous DAMPs, which promotes downstream pro-inflammatory cytokines synthesis. The result contributes to the alternative tumor microenvironment and host immunity and influences clinical treatment outcomes.24–26 RUNX3 encodes a heterodimer comprised of a Runt domain containing α subunit and a DNA-binding β subunit.27,28 It is reported that the high expression of RUNX3 in CD8+ T cells is essential to suppress transcription of CD4 silencer and regulates the expression of CD8+ T-cell specific genes.29–31 The differentiation of Th1 cells is determined by transcription factor T-bet and the induction of RUNX3 expression. The interaction between transcription factor T-bet and RUNX3 causes large production of IFN-γ and the silence of IL4 gene.15,32 However, the low expression of RUNX3 due to DNA methylation at CpG site has also been demonstrated in various tumors.33 For these reasons, the aim of this study was to investigate whether the high expression of RUNX3 influences irradiation immune responses related TLR9 signal activation and enhanced radiotherapy effects.
We characterized the expression of TLR9 in tumor tissues and RUNX3 in paracarcinoma tissues and explored the relationship with postoperative radiotherapy survival in lung cancer. The result present that TLR9 expression and tumor T staging among clinicopathologic features were the variables of relative risk in postoperative survival of lung cancer with statistic value 5.057 and 2.716, respectively. It suggested that TLR9 signal activation was the disadvantage of postoperative survival in lung cancer. And we observe no difference in PFS and OS of postoperative or radiotherapy survival in lung cancer. However patients with RUNX3 high expression had the better PFS and OS in postoperative radiotherapy, either positive or negative TLR9 expression. This result confirmed that RUNX3 expression in paracarcinoma tissues of lung cancer mediated irradiation effects and impact the survival on postoperative radiotherapy. The analysis of Cox regression on PFS in lung cancer radiotherapy revealed that the relative risk on tumor differentiation, tumor T staging, and RUNX3 was 2.109, 1.861, and 0.178, respectively. It was certain that the high expression of RUNX3 was significant to improve radiotherapy outcomes associated with TLR9 activation.
To test the effect of RUNX3 on lung cancer radiotherapy related TLR9 signal activation, we introduced 5-Aza-CdR and lung cancer A549 cell was exposed repeatedly and detected the expression of TLR9 and RUNX3 in experimental cells. The increasing levels of TLR9 protein obviously in A549high-TLR9 cells was observed. We also examined the levels of RUNX3 protein especially RUNX3-B in 5-Aza-CdR treatment cells was higher than in A549high-TLR9 cells. Furthermore we found that the G2/M cells in 5-Aza-CdR combining radiotherapy was sevenfold to the 5-Aza-CdR treatment group and twofold to the cell irradiated solely, respectively. It was maximized at irradiation 24 h and kept the irradiation advantage until 36 h and 48 h. It is known that the G2/M cells has the most sensitivity to irradiation. It was remarkable that 5-Aza-CdR treatment combined radiotherapy made more cells block in the G2/M phase and enhanced irradiation effect. In contrast, we observed the S cells prolonged in chloroquine combining radiotherapy, which was insensitive to X-rays. It suggested that only inhibiting TLR9 signaling was unable to improve the radiation resistance related TLR9 signaling activation. And this result also conformed to our first clinical investigation. Considering the higher proportion of G2/M cells in 5-Aza-CdR adjuvant radiotherapy we thought that RUNX3 expression upregulation played a key role in this process. It showed the radiosensitization of 5-Aza-CdR in lung cancer with TLR9 signaling activation by upregulating RUNX3 expression. We concluded that it was necessary to raise RUNX3 expression to promote the irradiation effect in lung cancer with TLR9 signaling activation.
Conclusion
In summary, TLR9 signaling activation involved in tumor progression and influenced the postoperative radiotherapy survival in lung cancer. It was notable that the increasing levels of RUNX3 protein could contribute to reverse radioresistance in this process and improve the survival related lung cancer radiotherapy. Of course, 5-Aza-CdR was considerable.
Acknowledgment
The experimental design and data analysis were directed by Dr Qiao. Zhuangxibing took part in the medical records collection in this study, Yuansujun participated in the experimental process. The authors are grateful for their hard work.
Disclosure
The authors report no competing interests in this work.
References
1. Chen K, Huang J, Gong W, Iribarren P, Dunlop NM, Wang JM. Toll-like receptors in inflmmation, infection and cancer. Int Immunopharmacol. 2007;7:1271–1285. doi:10.1016/j.intimp.2007.05.016
2. Droemann D, Albrecht D, Gerdes J, et al. Human lung cancer cells express functionally active Toll-like receptor 9. Respir Res. 2005;6:1. doi:10.1186/1465-9921-6-1
3. Merrell MA, Ilvesaro JM, Lehtonen N, et al. Toll-like receptor 9 agonists promote cellular invasion by increasing matrix metalloproteinase activity. Mol Cancer Res. 2006;4:437–447. doi:10.1158/1541-7786.MCR-06-0007
4. Takala H, Kauppila JH, Soini Y, et al. Toll-like receptor 9 is a novel biomarker for esophageal squamous cell dysplasia and squamous cell carcinoma progression. J Innate Immun. 2011;3:631–638. doi:10.1159/000329115
5. Ren T, Xu L, Jiao S, et al. TLR9 signaling promotes tumor progression of human lung cancer cell In vivo. Pathol Oncol Res. 2009;15(4):623–630. doi:10.1007/s12253-009-9162-0
6. Xu L, Wang C, Wen Z, et al. CpG oligodeoxynucleotides enhance the efficacy of adoptive cell transfer using tumor infiltrating lymphocytes by modifying the Th1 polarization and local infiltration of Th17 cells. Clin Dev Immunol. 2010;2010:410893. doi:10.1155/2010/410893
7. Kundu SD, Lee C, Billips BK, et al. The toll-like receptor pathway: a novel mechanism of infection-induced carcinogenesis of prostate epithelial cells. Prostate. 2008;68(2):223–229. doi:10.1002/pros.20710
8. Väisänen MR, Väisänen T, Jukkola-Vuorinen A, et al. Expression of toll-like receptor-9 is increased in poorly differentiated prostate tumors. Prostate. 2010;70(8):817–824. doi:10.1002/pros.21115
9. Kauppila JH, Takala H, Selander KS, Lehenkari PP, Saarnio J, Karttunen TJ. Increased Toll-like receptor 9 expression indicates adverse prognosis in oesophageal adenocarcinoma. Histopathology. 2011;59(4):643–649. doi:10.1111/j.1365-2559.2011.03991.x
10. Wang C, Cao S, Yan Y, et al. TLR9 expression in glioma tissues correlated to glioma progression and the prognosis of GBM patients. BMC Cancer. 2010;10:415. doi:10.1186/1471-2407-10-415
11. Bae SC, Choi JK. Tumor suppressor activity of RUNX3. Oncogene. 2004;23(24):4336–4340. doi:10.1038/sj.onc.1207286
12. Bangsow C, Rubins N, Glusman G, et al. The RUNX3 gene-sequence, structure and regulated expression. Gene. 2001;279(2):22l–232. doi:10.1016/S0378-1119(01)00760-0
13. Milner JJ, Toma C, Yu B, et al. Runx3 programs CD8+ T cell residency in non-lymphoid tissues and tumors. Nature. 2017;552(7684):253–257. doi:10.1038/nature24993
14. Mami-Chouaib F, Blanc C, Corgnac S, et al. Resident memory T cells, critical components in tumor immunology. J ImmunoTher Cancer. 2018;6:87. doi:10.1186/s40425-018-0399-6
15. Djuretic IM, Levanon D, Negreanu V, Groner Y, Rao A, Ansel KM. Transcription factors T-bet and Runx3 cooperate to activate IFN-γ and silence IL4 in T helper type 1 cells. Nat Immunol. 2006;8(7):145–153.
16. Torres-Roca JF, Eschrich S, Zhao H, et al. Prediction of radiation sensitivity using a gene expression classifier. Cancer Res. 2005;65:7169–7176. doi:10.1158/0008-5472.CAN-05-0656
17. Esteller M. Epigenetic gene silencing in cancer: the DNA hypermethylome. Hum Mol Genet. 2007;16(1):R50–R59. doi:10.1093/hmg/ddm018
18. Kim HJ, Kim JH, Chie EK, et al. DNMT (DNA methyltransferase) inhibitors radiosensitize human cancer cells by suppressing DNA repair activity. Radiat Oncol. 2012;7(1):39. doi:10.1186/1748-717X-7-39
19. Valdez BC, Li Y, Murray D, Corn P, Champlin RE, Andersson BS. 5-Aza-2′-deoxycytidine sensitizes busulfan-resistant myeloid leukemia cells by regulating expression of genes involved in cell cycle checkpoint and apoptosis. Leuk Res. 2010;34(3):364–372. doi:10.1016/j.leukres.2009.08.014
20. Oing C, Verem I, Mansour WY, Bokemeyer C, Dyshlovoy S, Honecker F. 5-Azacitidine exerts prolonged pro-apoptotic effects and overcomes cisplatin-resistance in non-seminomatous germ cell tumor cells. Int J of Mol Sci. 2019;20:21. doi:10.3390/ijms20010021
21. Yan L, Xu G, Qiao T, Chen W, Yuan S, Li X. CpG ODN 7909 increases radiation sensitivity of radiation-resistant human lung adenocarcinoma cell line by overexpression of toll-like receptor 9. Cancer Biother Radiopharm. 2013;28(7):559–564. doi:10.1089/cbr.2012.1450
22. Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol. 2003;21:335–376. doi:10.1146/annurev.immunol.21.120601.141126
23. Hoshino K, Kaisho T, Lwabe T, Takeuchi O, Akira S. Differential involvement of IFN-beta in Toll-like receptor stimulated dendritic cell activation. Int Immunol. 2002;14(10):1225–1231. doi:10.1093/intimm/dxf089
24. Johnson GB, Brunn GJ, Tang AH, Platt JL. Evolutionary clues to the functions of the Toll-like family as surveillance receptors. Trends Immunol. 2003;24(1):19–24.
25. Peter M, Bode K, Lipford GB, Eberle F, Heeg K, Dalpke AH. Characterization of suppressive oligodeoxynucleotides that inhibit Toll-like receptor 9 mediated activation of innate immunity. Immunology. 2008;123(1):118–128. doi:10.1111/j.1365-2567.2007.02718.x
26. Serafini P, De Santo C, Marigo I, et al. Derangement of immune responces by myeloid suppressor cells. Cancer Immunol Lmmunother. 2004;53(2):64–72. doi:10.1007/s00262-003-0443-2
27. Yano T, Ito K, Fukamachi H, et al. The RUNX3 tumor suppressor upregulates Bim in gastric epithelial cells undergoing transfoming growth factor β-induced apoptosis. Mol Cell Biol. 2006;26:4474–4488. doi:10.1128/MCB.01926-05
28. Durst KL, Hiebert SW. Role of RUNX family members in transcriptional repression and gene silencing. Oncogene. 2004;23(24):4220–4224. doi:10.1038/sj.onc.1207122
29. Ohno S, Sato T, Kohu K, et al. Runx proteins are involved in regulation of CDl22, Ly49 family and IFN-γ expression during NK cell differentiation. Int Immunol. 2008;20(1):7l–79. doi:10.1093/intimm/dxm120
30. Taniuchi I, Littman DR. Epigenetic gene silencing by Runx proteins. Oncogene. 2004;23(24):4341–4345. doi:10.1038/sj.onc.1207671
31. Sato T, Ohno S, Hayashi T, et al. Dual functions of Runx proteins for reactivating CD8 and silencing CD4 at the commitment process into CD8 thymocytes. Immunity. 2005;22(3):317–328. doi:10.1016/j.immuni.2005.01.012
32. Schulz EG, Mariani L, Radbruch A, Höfer T. Sequential polarization and imprinting of type l T helper lymphocytes by interferon-γ and interleukin-12. Immunity. 2009;30(5):673–683. doi:10.1016/j.immuni.2009.03.013
33. Wolff EM, Liang G, Cortez CC, et al. RUNX3 methylation reveals that bladder tumors are older in patients with a history of smoking. Cancer Res. 2008;68(15):6208–6214. doi:10.1158/0008-5472.CAN-07-6616
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.