Back to Journals » Infection and Drug Resistance » Volume 12
The Pathogenesis Of Streptococcus anginosus In Aerobic Vaginitis
Authors Tao Z ![]() , Zhang L, Zhang Q, Lv T, Chen R, Wang L, Huang Z, Hu L, Liao Q
, Zhang L, Zhang Q, Lv T, Chen R, Wang L, Huang Z, Hu L, Liao Q
Received 19 August 2019
Accepted for publication 31 October 2019
Published 4 December 2019 Volume 2019:12 Pages 3745—3754
DOI https://doi.org/10.2147/IDR.S227883
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Zhi Tao,1,2 Lei Zhang,2 Qiongqiong Zhang,1,2 Tao Lv,2 Rui Chen,1,2 Lijun Wang,2 Zhenyu Huang,1,2 Long Hu,1 Qinping Liao2
1School of Clinical Medicine, Tsinghua University, Beijing 100084, People’s Republic of China; 2Department of Obstetrics and Gynecology, Beijing Tsinghua Changgung Hospital, School of Clinical Medicine, Tsinghua University, Beijing 102218, People’s Republic of China
Correspondence: Qinping Liao
Department of Obstetrics and Gynecology, Beijing Tsinghua Changgung Hospital, School of Clinical Medicine, Tsinghua University, Litang Road No. 168, Changping District, Beijing 102218, People’s Republic of China
Tel/Fax + 86 10 5611 8901
Email [email protected]
Background: Aerobic vaginitis (AV) is a newly defined type of bacterial vaginitis, but its pathogenesis is not yet clear. Streptococcus anginosus appears as an emerging pathogen in recent case reports, and colonizes in vagina of patients with AV. In this study, we investigate the pathogenesis of S. anginosus in AV.
Materials and methods: (1) We collected 41 vaginal specimens from 21 healthy, fertile women with normal vaginal flora (NM), 10 with bacterial vaginosis (BV) and 10 with AV; their microbiome structure was analysed by 16S rRNA gene sequencing. (2) S. anginosus and vaginal epithelial cells were cocultured in vitro, and cytotoxicity was tested by an LDH kit. (3) The S. anginosus virulence gene sag was knocked out, and the cytotoxicity of the mutant in vaginal epithelial cells was tested.
Results: (1) The microbiome structure of AV was dramatically different from that of BV and NM. The predominant genera of the three groups were Streptococcus spp., Gardnerella spp. and Lactobacillus spp. Streptococcus spp. were significantly more abundant in AV than in BV (95% CI [0.1391, 0.8676], P<0.01) and NM (95% CI [0.1391, 0.8676], P<0.01). (2) S. anginosus was the dominant species in AV (95% CI [0.04672, 0.1097], P<0.01). (3) The mean cytotoxicity of S. anginosus in vaginal epithelial cells was 58.34% for the wild type (WT) and 16.43% for the mutant; this difference was significant (95% CI [−60.55, −23.28], P<0.01).
Conclusion: S. anginosus was the predominant microorganism in patients with AV in our study. S. anginosus caused vaginal epithelial cell lysis, indicating that S. anginosus is an AV pathogen. The S. anginosus virulence gene sag is vital for vaginal epithelial cell lysis.
Keywords: Streptococcus anginosus, aerobic vaginitis, vaginal microbiome, gene knockout, next-generation sequencing technology, NGS
Plain Language Summary
Aerobic vaginitis (AV), which is an infection caused by aerobic bacteria, was newly defined by Donders in 2002 and diagnosed according to Donders’ score. However, not all AV cases are symptomatic or can be cured by antibiotics targeting aerobic bacteria. Therefore, study on the pathogen and pathogenesis of AV is very important. However, the data are limited. In this study, we found that S. anginosus was dominant and pathogenic in a subset of AV patients. All the results in our study are novel and unpublished.
Introduction
Aerobic vaginitis (AV) was first defined by Donders in 2002 as a type of vaginal infection caused by aerobic bacteria, and AV is diagnosed according to Donders’ score criteria.1 Patients with AV present with vaginal itching, purulent discharge with or without a rotten odour, and/or red vaginal wall. AV is not a new entity, but the second description of a type of bacterial vaginitis, which was first reported by Scheffey in 19562 and named desquamative inflammatory vaginitis (DIV) by Gardner in 1968;3,4 Both AV and DIV is distinct from bacterial vaginosis (BV). There is no difference between severe AV and DIV.5 Previous studies have shown that AV is associated with miscarriage,6 increased risk of amnionitis,7 placental histological inflammation,8 premature rupture of membranes,9 preterm delivery,6,10 foetal funisitis,8 and major cervical cytological abnormalities.11,12 The prevalence of AV is reported to be 5–27.6% in various areas.6,9,12–17 Thus, it is vital to identify AV so that standardized treatments can be developed to avoid complications.
Like BV, AV is considered an endogenous infection,18 but the causal pathogens of AV have not been studied as thoroughly as those of BV. Escherichia coli, Streptococcus spp., Staphylococcus aureus, coagulase-negative staphylococci, Staphylococcus epidermidis, and Enterococcus faecalis have been found to colonize in significant numbers in AV patients.1,14,15 However, the pathogenesis of aerobic bacteria in AV is not yet clear. Further data are limited. Facilitated by the progression of sequencing technology and bioinformatics analysis, studies on the vaginal microbiome have increased in recent years. The work of the National Institutes of Health (NIH) has illustrated new insights into correlations between the vaginal microbiome and the general health of women.19 To date, no study on the pathogen of AV by next-generation sequencing (NGS) has been performed.
Except for E. coli, all of the colonizing bacteria in AV stated above are cocci. In this study, we concentrated on purely symptomatic AV caused by cocci. Our research focused on identifying pathogens by high-throughput 16S rRNA gene sequencing with newly designed primers.
Samples And Methodology
Subject Selection
Forty-one nonpregnant fertile women were enrolled in our study, in which there were 10 women with symptomatic AV diagnosed by Donders’ score,1 10 women with BV diagnosed by Nugent score20 (≥7ʹ) and 21 women with normal vaginal microbiota (NM). All of the participants were informed about the study, signed an informed consent form and were then examined by an appointed attending doctor in Beijing Tsinghua Changgung Hospital from July 2017 to September 2018. This study was reviewed and approved by the Ethics Committee Board at the hospital. All participants were aged from 18 to 50 years and matched the following criteria: (1) did not use antibiotics or immunosuppressants during the last 30 days; (2) had no vaginal intercourse or vaginal procedures in the preceding week; (3) were free of systemic diseases such as infectious diseases, diabetes, autoimmune diseases, and cancer; (4) had at least one of the following clinical characteristics: itching, purulent discharge, dyspareunia, rotten odour, vaginal wall reddening (for AV only); (5) presence of cocci or chained cocci with or without parabasal epithelial cells in vaginal smears (for AV only).
Sample Collection And Preparation
The vaginal discharge of each participant was collected with two sterile cotton swabs of the lateral upper vaginal wall. One swab was examined with Donders’ score and Gram stain, and the other swab was diluted in 1 mL of sterile PBS for total DNA extraction, kept in an icebox, and then stored at −80°C within one hour.
Total DNA Extraction, PCR Amplification And 16S rRNA Gene Sequencing
Total DNA from the vaginal discharge was extracted according to the manufacturer’s protocol (TIANamp Bacteria DNA Kit, TIANGEN BIOTECH, Beijing, China). The extracted DNA concentration of each sample varied from 47.7–283.6 ng/mL as tested by a NANODROP ONE (Thermo Scientific), and the purity of the DNA was consistent.
The V1-V2 hypervariable regions of the bacterial 16S rRNA gene were amplified by BIO-RAD in a 50 μL reaction system including 30 cycles. Each reaction contained 20 pmol of each of the primers (27Fʹ: 5ʹ-AGRGTTYGATYCTGGCTCAG-3ʹ; 338Rʹ: 5ʹ-GCTGCCTCCCGTAGGAGT-3ʹ), 25 μL of 2Xtaq PCR Master Mix (TIANGEN), 2 μL of DNA template and 19 μL of double-distilled water (ddwater; TIANGEN). Each sample had a pair of specific barcodes flanking primers 27Fʹ and 338Rʹ. Thermocycling was initiated by a 3 min incubation at 94°, followed by cycling parameters of 94°C for 30 s, 55°C for 30 s, and 72°C for 30 s and terminated by a 5 min incubation at 72°C. All PCR samples were tested by 1% agarose gel electrophoresis (400–500 bp). After generating sequence libraries with a TruSeq® DNA PCR-Free Sample Preparation kit (Illumina, USA), the concentration and purity were quantified with an Agilent 2100 Qubit 2.0 bioanalyzer and a BIO-RAD CFX96. Finally, the libraries were sequenced on the Illumina HiSeq 2500 platform using the 250 paired-end protocol.
Processing Of Sequencing Data
After sequencing, the raw data (Figures S1 and S2) were filtered according to the QIIME (V1.7.0) quality control process. Then, the paired reads of the clean data were spliced, and the sequences were merged with FLASH software (V1.2.7). Demultiplexing was performed with in-house R scripts, according to the sample-specific barcode. A 16S sequence reference index was built using the command “bowtie2-build”, with default parameters. All reads were aligned against the prebuilt index using bowtie2, with a parameter of “bowtie2–local”. Alignments were associated with taxa by a sequence-id-to-taxonomy map, provided by the reference database, using a custom Perl script. Unique reads were counted for each taxon, and the abundance was calculated for all taxa. Species with an abundance of less than 1% or read number less than 5 were excluded. For reference databases, we used the SILVA database (SSU 128 Ref NR 99 version from https://www.arb-silva.de). All taxonomic data are summarized at the species level. The data of all samples was available from https://bigd.big.ac.cn/gsa/ (CRA002031).
Construction Of Δsag ATCC33397
S. anginosus (ATCC33397=NCTC10713) was purchased from ATCC (01077) and cultured in brain heart infusion (BHI; BD) broth and agar. The primers used in this paper are listed in Table 1, and the working concentration for all primers was 10 pmol/μL. The erythromycin resistance gene fragment was amplified from plasmid pJDC9 (provided by Jing-Ren Zhang, Tsinghua University) with primers erm-F2 and erm-R2. After preparation of ATCC33397 genomic DNA, up- and downstream regions of sag gene clusters were amplified with primers Up-F1, Up-R1, Down-F3, and Down-R3. Each PCR contained 10 μL of buffer, 4 μL of dNTPs, 0.5 μL of STAR, 40 ng of genomic DNA template and ddwater to produce a final volume of 50 μL. PCRs were performed under the following conditions: 98°C for 10 min, [98°C for 30 s, 55°C for 1 min/kb, 72°C 3 min] 33 cycles, and 72°C for 5 min (PrimeSTAR DNA polymerase was purchased from Takara). All PCR fragments were purified according to the Magen Pure DNA Mini kit manufacturer’s protocol. Next, fusion PCR of the up/downstream regions of sag gene clusters and the erythromycin resistance gene fragment was performed. The first round in the fusion PCR process included 12 cycles, and the PCR volume was 50 μL comprising 10 μL of buffer, 4 μL of dNTPs, 0.5 μL of STAR, DNA template (upstream:erm:downstream=1:1:1>800 ng) and ddwater. The PCR conditions were the same as those described above except for the 12 cycles. The second round of the fusion PCR process included 25 cycles. The reaction contained 10 μL of buffer, 4 μL of dNTPs, 1 μL of Up-F1 primer, 1 μL of Down-R3 primer, 0.5 μL of STAR, 10 μL of purified up+erm+down DNA template and 23.5 μL of ddwater. The PCR conditions were the same as described above except for the 25 cycles. Finally, the fusion PCR product was purified and stored at −20°C. All DNA fragments were verified by 1% agarose gel electrophoresis (Figure 1A–I).
 |
Table 1 Primers Used In This Study |
 |
Figure 1 The PCR fragments for the knockout of the sag gene and the S. anginosus (ATCC33397/NCTC10713) growing on Blood Agar Plate (BAP). (A) The up+erm+down fragment after fusion PCR; (B) is the up+erm fragment and (C) is the erm+down fragment. (D) The sag fragment of ATCC33397 (WT); (E and F) are erm fragments; (G) is upstream and (H) is downstream of the sag coding area. (I) The sag fragment of the mutant amplified from Δsag ATCC33397 with primers sag-F-primer and sag-R-primer shown in Table 1. (J) The wild type, withαhaemolytic activity, and (K) is the mutant with no sag gene and no haemolytic activity. |
Transformation of ATCC33397 was conducted using a competence-stimulated peptide (CSP; synthesized by ChinaPeptides), as previously reported.21 First, 100 μg/mL of CSP and fusion PCR product was added to 200 μL of a diluted overnight culture of ATCC33397 (in BHI broth), which was then incubated at 37°C in 5% CO2 for 3 h. Then, the incubation was continued at room temperature for 2 h. Finally, the transformation mixture was plated on BHI agar plates containing 100 μg/mL erythromycin to select transformants (Figure 1J and K).
Coculture Of S. Anginosus And Vaginal Epithelial Cells
VK2/E6E7 cell line was purchased from ATCC (CRL-2616) and cultured in Keratinocyte-serum free medium (K-SFM, Gibco) with 0.1 ng/mL human recombinant EGF (PeproTech), 0.05 mg/mL bovine pituitary extract (Science cell) and 44.1 mg/L Calcium chloride at 37°C and 5% CO2. The epithelial cell(VK2/E6E7) cytotoxicity produced by S. anginosus (ATCC33397) was determined with an LDH assay kit. VK2/E6E7 cells were cultured with wild-type ATCC33397 and the mutant Δsag ATCC33397 at a concentration of 106 colony forming units (cfu)/mL for 1 h at 37°C and 5% CO2 (initial MOI: 1).
Statistical Analysis
The sample number was calculated using PASS software with type I error=0.05 and target power=0.8 according to the AV microorganism proportions reported by Donders.1
The 16S rRNA gene sequencing data were converted to proportions to determine the percentage of the microbiome. The predominant species were defined as the largest number of microorganisms, equal to the highest percentage of reads. Wilcox ranksum test was used to analyse the variance in abundance between and within groups.
The cytotoxicity data were expressed as the mean±SEM and analysed for significance by t-test (a P value <0.05 was used as the threshold of significance). All experiments were repeated three times.
Results
The Clinical Characteristics Of The AV Patients
The 10 patients enrolled in our study presented at least one AV symptom, such as purulent discharge, itching, dyspareunia, rotten odour and vaginal wall reddening. The most frequent symptom was purulent discharge (9/10), the second most frequent was itching (7/10), and the third was vaginal wall reddening (4/10). All patients with dominant S. anginosus (7/10) presented purulent discharge, and 5 of these patients complained of itching (5/7). All patients found to have vaginal wall reddening during the gynaecological examination had a greater abundance of S. anginosus than of other species (Table 2).
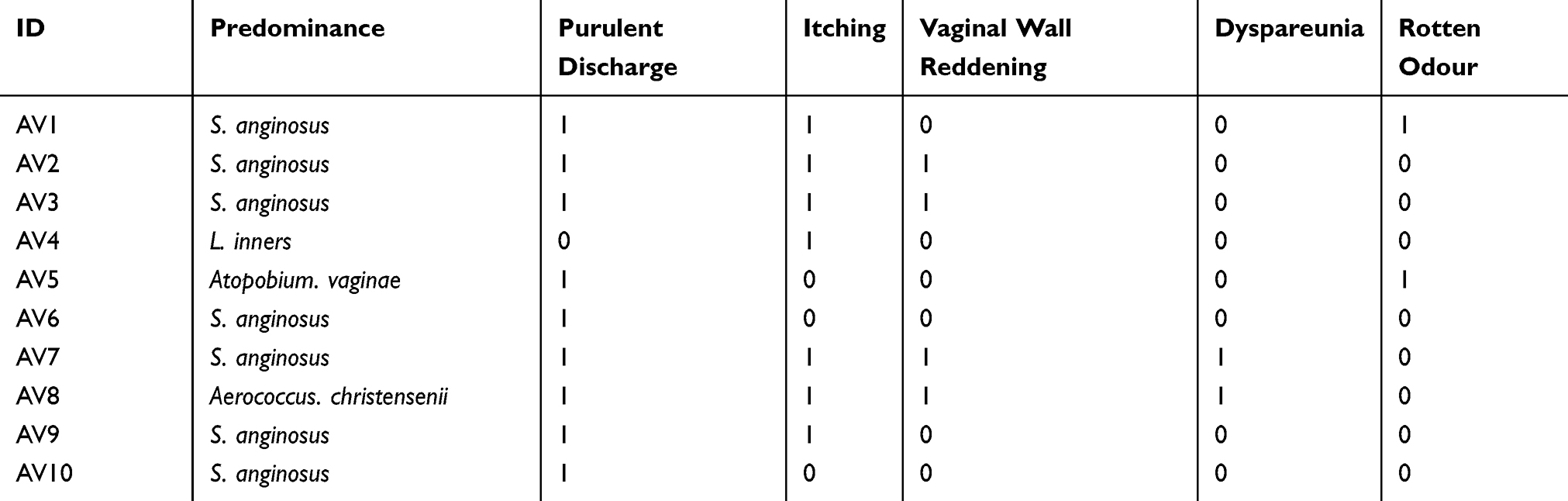 |
Table 2 The Symptoms Of Each Patient With AV. “1” Represents Yes, Meaning The Patient Had The Characteristic, And “0” Represents No |
The Structure Of The Symptomatic AV, BV, And NM Vaginal Microbiome
The clustering analysis of AV, BV and NM sequencing data indicated a distinct community between three groups based on the operational taxonomic unit (OTU)-taxon-origin (Figures S5–S8). AV mostly correlated with S. anginosus, BV mostly correlated with Gardnerella vaginalis, and NM clustered with Lactobacillus crispatus, L. gasseri, L. inners and L. jensenii (Figure 2). The dendrogram showed that the distance of the AV and BV samples was larger than that of the NM samples, but most of the AV or BV samples were close to each other (Figure 3).
 |
Figure 2 The clustering analysis of AV, BV and NM. The relevance of species to each sample is shown. The deeper the colour is, the more relevant the sample is to the species. |
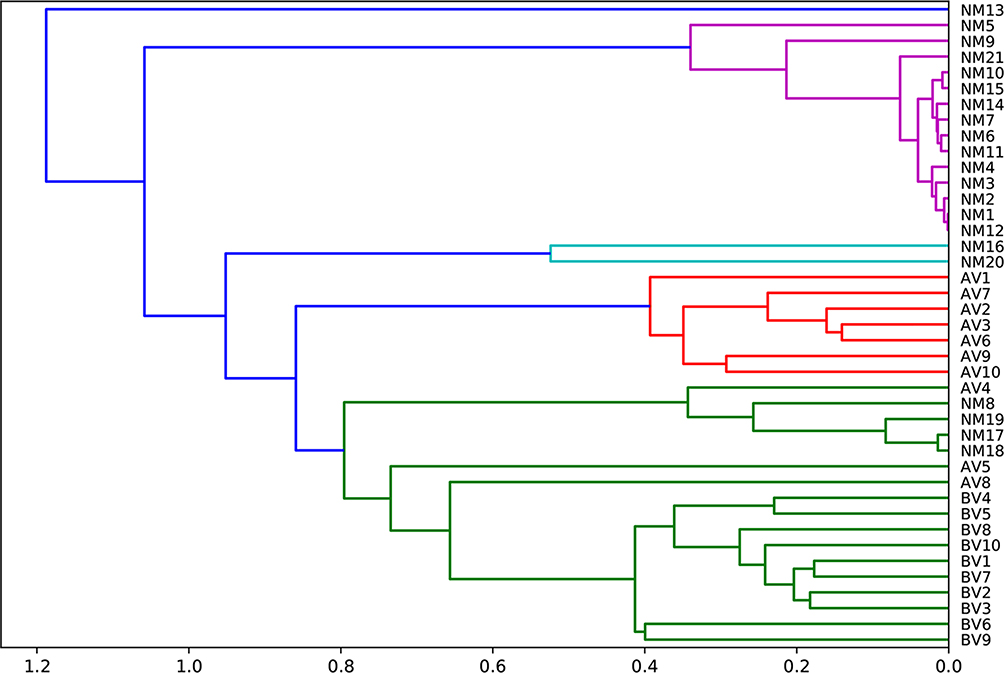 |
Figure 3 The dendrograms of AV, BV and NM samples. The dimensions of the tree show the distance of the samples. |
Streptococcus, Lactobacillus and Gardnerella were the predominant genera in AV, NM and BV, respectively (Figures 4 and S3). Streptococcus was significantly more abundant in AV than in BV and NM (P<0.01) (Figure 5). S. anginosus was the dominant species in AV, as shown in Figure 6, and it was barely present in NM and BV samples (Figures 6 and S4). Compared with NM, both AV and BV were associated with a significant decrease in Lactobacillus crispatus (Figures 6 and S4). The comparison of the abundance of each species in AV revealed that S. anginosus was significantly more abundant than other species (P<0.01). (Figure 7)
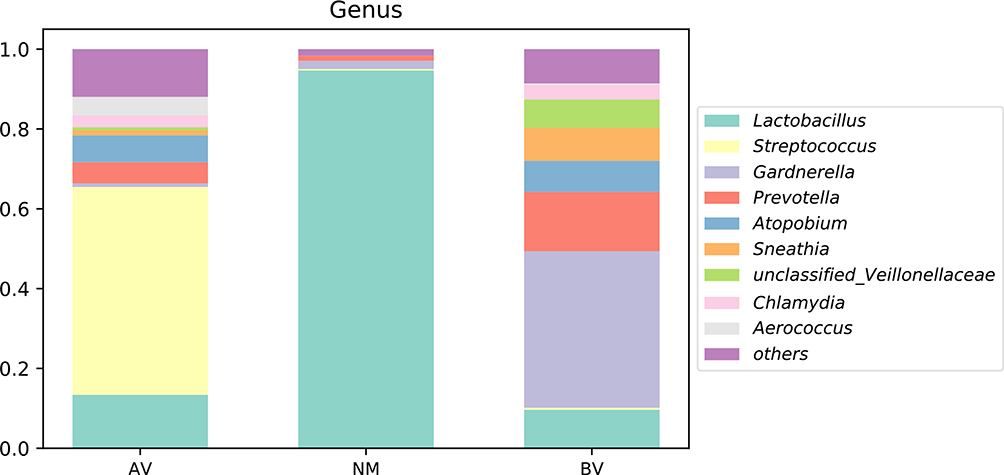 |
Figure 4 The average abundance of genera in AV, NM and BV samples. The total abundance of each group is 1. |
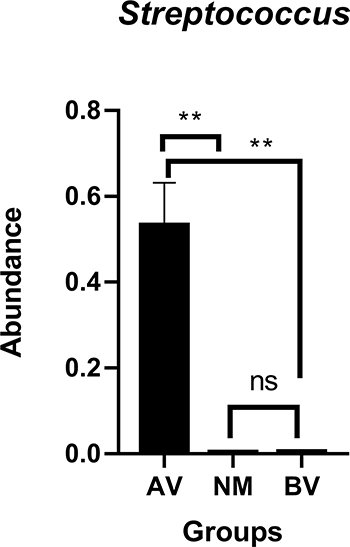 |
Figure 5 Comparison of the abundance of Streptococcus between AV, NM and BV samples. **p<0.01. |
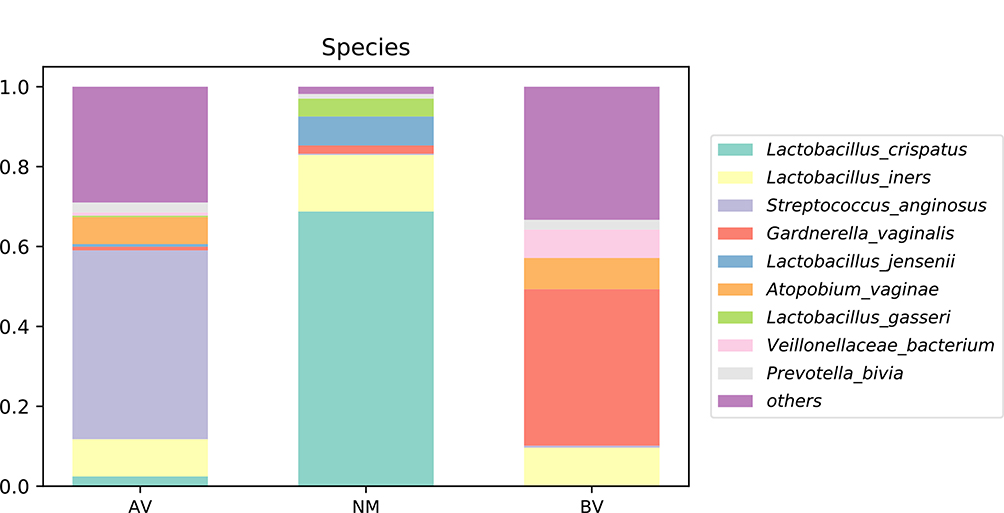 |
Figure 6 The average abundance of species in AV, NM and BV samples. The total abundance of each group is 1. |
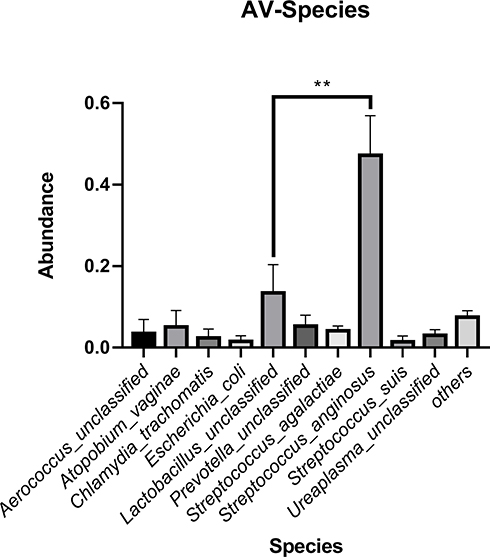 |
Figure 7 The average abundance of each species in the AV microbiome. S. anginosus was significantly more abundant than other species. **p<0.01. |
The Cytotoxicity Of Streptococcus In Vaginal Epithelial Cells
S. anginosus (WT) was cytotoxic to vaginal epithelial cells. The cytotoxicity of the WT and the mutant was (58.34±6.712)% and (16.43±6.712)% respectively. Though the mutant S. anginosus grew at the same rate as the wild type, the cytotoxicity difference was significant (P<0.01) (Figures 8 and 9).
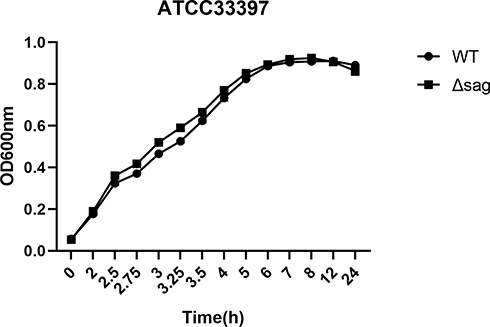 |
Figure 8 The growth curves of S. anginosus (ATCC33397) and the Δsag mutant. |
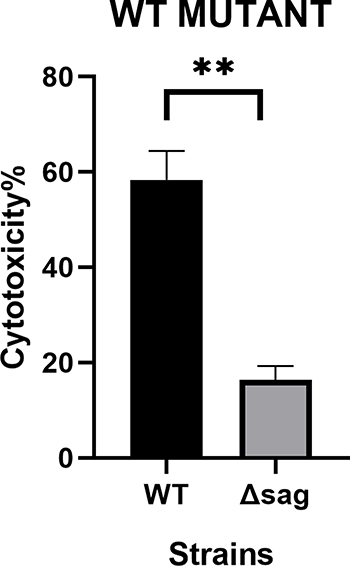 |
Figure 9 The cytotoxicity of S. anginosus (ATCC33397) and the Δsag mutant was significantly different. **p<0.01. |
Discussion
Fertile women are the most important category of AV patients, but not all of these patients are symptomatic.5 A previous study found that 53.85% of AV cases involved mixed infections, including BV, trichomonas vaginitis (TV) or vulvovaginal candidiasis (VVC) .14 We noticed that patients with BV, TV or VVC frequently score above 3ʹ (according to Donders’ score criteria), and the treatment of other infections affects the AV score. Therefore, single-cause symptomatic AV needs further attention. We recruited symptomatic women with pure AV caused by cocci and found that purulent discharge was present in 90% and itching was present in 70%, which is higher than that of all types of AV.1
Consistent with a previous study,15 the 16S rRNA sequencing of symptomatic fertile women with AV in our study showed a dramatic predominance of Streptococcus spp. However, at the species level, S. anginosus was more abundant than Streptococcus agalactiae in AV patients in our study. Relative to previous studies, this is a new discovery.18
S. anginosus is a commensal strain belonging to the S. anginosus group (SAG) in the human oral cavity, genitourinary tract, and gastrointestinal tract that was not recognized as a pathogen until nearly 20 years. Case reports about S. anginosus as an emerging pathogen associated with purulent infections have become increasingly frequent.22 S. anginosus accounted for 82% of the 245 patient-unique strains isolated from hospitalized patients by an automated system during a 37-month investigation in a tertiary care teaching hospital in Israel.23 S. anginosus is an opportunistic pathogen that is able to survive in acid conditions,24 adhere to exposed basement membranes of human and cultured epithelial cells,25,26 and lyse erythrocytes and some epithelial cell types.27 Streptolysin S (SLS) is an important S. anginosus virulence factor encoded by the sag gene cluster, including sagA1/A2, B-I.22,27–32 S. anginosus ATCC33397 was cytotoxic to vaginal epithelial cells in our study, and the toxicity was significantly reduced by sag gene ablation.
All patients in whom S. anginosus was the dominant species, presented purulent discharge, and 5 of these patients complained of itching. Furthermore, all patients found to have vaginal wall reddening during gynaecological examination had a greater abundance of S. anginosus than other species. Based on the above results, we hypothesized that S. anginosus causes purulent discharge and vaginal wall reddening in patients with AV, which is associated with damage to the vaginal epithelial wall. Furthermore, the pathogenic characteristic of S. anginosus matched the AV patient symptoms.
Our results suggest that S. anginosus may be a pathogenic organism that causes AV and may be associated with purulent discharge. However, we should collect more symptomatic subjects to verify the predominance and isolate clinical strains to test their pathogenic genes and cytotoxicity. In addition, more research is needed to further indicate the virulence activity of SLS from S. anginosus in the pathogenesis of AV.
Conclusion
S. anginosus is the predominant microorganism in patients with symptomatic AV and caused the lysis of vaginal epithelial cells, which indicates that S. anginosus is an AV pathogen. The S. anginosus virulence gene sag is vital for vaginal epithelial cell lysis.
Ethics Approval
All procedures of this study involving human subjects were reviewed and approved by the Ethics Committee Board of Beijing Tsinghua Changgung Hospital in 2016 (File NO 19201-0-01). We confirm that this study was conducted in accordance with the Declaration of Helsinki.
Acknowledgments
We acknowledge the funding of Natural Science Foundation of China (NSFC; grant no. 81671409) and Beijing Municipal Administration of Hospitals Clinical Medicine Development of Special Funding (grant no. XMLX201605), and the contributions of public experimental platform of Beijing Tsinghua Changgung Hospital and Jing-Ren Zhang Lab of Tsinghua University. We also thank Xiaoqian Wang, Ying Liu, Jun Liu, Xi Yang, Ke Ma, Meng Zhao, and Ying Wang to help talking to patients about the study and collecting samples. Thank our families for all their helps.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Donders GG, Vereecken A, Bosmans E, Dekeersmaecker A, Salembier G, Spitz B. Definition of a type of abnormal vaginal flora that is distinct from bacterial vaginosis: aerobic vaginitis. BJOG. 2002;109(1):34–43. doi:10.1111/j.1471-0528.2002.00432.x
2. Scheffey LC, Rakoff AE, Lang WR. An unusual case of exudative vaginitis (hydrorrhea vaginalis) treated with local hydrocortisone. Am J Obstet Gynecol. 1956;72(1):208–211. doi:10.1016/S0002-9378(16)37537-8
3. Gardner HL. Desquamative inflammatory vaginitis: a newly defined entity. Am J Obstet Gynecol. 1968;102(8):1102–1105. doi:10.1016/0002-9378(68)90399-2
4. Gardner HL. Desquamative inflammatory vaginitis. Am J Obstet Gynecol. 1969;104(8):1225. doi:10.1016/S0002-9378(16)34307-1
5. Donders GGG, Bellen G, Grinceviciene S, Ruban K, Vieira-Baptista P. Aerobic vaginitis: no longer a stranger. Res Microbiol. 2017;168(9–10):845–858. doi:10.1016/j.resmic.2017.04.004
6. Donders GG, Van Calsteren K, Bellen G, et al. Predictive value for preterm birth of abnormal vaginal flora, bacterial vaginosis and aerobic vaginitis during the first trimester of pregnancy. BJOG. 2009;116(10):1315–1324. doi:10.1111/j.1471-0528.2009.02237.x
7. Curzik D, Drazancic A, Hrgovic Z. Nonspecific aerobic vaginitis and pregnancy. Fetal Diagn Ther. 2001;16(3):187–192. doi:10.1159/000053906
8. Rezeberga D, Lazdane G, Kroica J, Sokolova L, Donders GG. Placental histological inflammation and reproductive tract infections in a low risk pregnant population in Latvia. Acta Obstet Gynecol Scand. 2008;87(3):360–365. doi:10.1080/00016340801936487
9. Han C, Li H, Han L, et al. Aerobic vaginitis in late pregnancy and outcomes of pregnancy. Eur J Clin Microbiol Infect Dis. 2018;38(4).
10. Lamont RF, Taylor-Robinson D. The role of bacterial vaginosis, aerobic vaginitis, abnormal vaginal flora and the risk of preterm birth. BJOG. 2010;117(1):
11. Jahic M, Mulavdic M, Hadzimehmedovic A, Jahic E. Association between aerobic vaginitis, bacterial vaginosis and squamous intraepithelial lesion of low grade. Med Arch. 2013;67(2):94–96. doi:10.5455/medarh.2013.67.94-96
12. Vieira-Baptista P, Lima-Silva J, Pinto C, et al. Bacterial vaginosis, aerobic vaginitis, vaginal inflammation and major Pap smear abnormalities. Eur J Clin Microbiol Infect Dis. 2016;35(4):657–664. doi:10.1007/s10096-016-2584-1
13. Tomusiak A, Heczko PB, Janeczko J, Adamski P, Pilarczyk-Zurek M, Strus M. Bacterial infections of the lower genital tract in fertile and infertile women from the southeastern Poland. Ginekol Pol. 2013;84(5):352–358.
14. Fan A, Yue Y, Geng N, Zhang H, Wang Y, Xue F. Aerobic vaginitis and mixed infections: comparison of clinical and laboratory findings. Arch Gynecol Obstet. 2013;287(2):329–335. doi:10.1007/s00404-012-2571-4
15. Rumyantseva TA, Bellen G, Savochkina YA, Guschin AE, Donders GG. Diagnosis of aerobic vaginitis by quantitative real-time PCR. Arch Gynecol Obstet. 2016;294(1):109–114. doi:10.1007/s00404-015-4007-4
16. Wang ZL, Fu LY, Xiong ZA, et al. Diagnosis and microecological characteristics of aerobic vaginitis in outpatients based on preformed enzymes. Taiwan J Obstet Gynecol. 2016;55(1):40–44. doi:10.1016/j.tjog.2015.06.012
17. Vieira-Baptista P, Grinceviciene S, Bellen G, et al. Genital tract infections in an isolated community: 100 women of the principe Island. Infect Dis Obstet Gynecol. 2017;2017:3058569. doi:10.1155/2017/3058569
18. Kaambo E, Africa C, Chambuso R, Passmore JS. Vaginal microbiomes associated with aerobic vaginitis and bacterial vaginosis. Front Public Health. 2018;6:78. doi:10.3389/fpubh.2018.00078
19. Huang B, Fettweis JM, Brooks JP, Jefferson KK, Buck GA. The changing landscape of the vaginal microbiome. Clin Lab Med. 2014;34(4):747–761. doi:10.1016/j.cll.2014.08.006
20. Nugent RP, Krohn MA, Hillier SL. Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. J Clin Microbiol. 1991;29(2):297–301.
21. Havarstein LS, Hakenbeck R, Gaustad P. Natural competence in the genus Streptococcus: evidence that streptococci can change pherotype by interspecies recombinational exchanges. J Bacteriol. 1997;179(21):6589–6594. doi:10.1128/jb.179.21.6589-6594.1997
22. Kawaguchi Y, Tabata A, Nagamune H, Ohkura K. Molecular analysis of Streptococcus anginosus-derived SagA peptides. Anticancer Res. 2014;34(8):4627–4631.
23. Siegman-Igra Y, Azmon Y, Schwartz D. Milleri group streptococcus–a stepchild in the viridans family. Eur J Clin Microbiol Infect Dis. 2012;31(9):2453–2459. doi:10.1007/s10096-012-1589-7
24. Sasaki M, Kodama Y, Shimoyama Y, Ishikawa T, Kimura S. Aciduricity and acid tolerance mechanisms of Streptococcus anginosus. J Gen Appl Microbiol. 2018;64(4):174–179.
25. Kodama Y, Ishikawa T, Shimoyama Y, Sasaki D, Kimura S, Sasaki M. The fibronectin-binding protein homologue Fbp62 of Streptococcus anginosus is a potent virulence factor. Microbiol Immunol. 2018;62(10):624–634. doi:10.1111/mim.v62.10
26. Allen BL, Hook M. Isolation of a putative laminin binding protein from Streptococcus anginosus. Microb Pathog. 2002;33(1):23–31. doi:10.1006/mpat.2002.0510
27. Asam D, Mauerer S, Spellerberg B. Streptolysin S of Streptococcus anginosus exhibits broad-range hemolytic activity. Med Microbiol Immunol. 2015;204(2):227–237. doi:10.1007/s00430-014-0363-0
28. Tabata A, Nakano K, Ohkura K, et al. Novel twin streptolysin S-like peptides encoded in the sag operon homologue of beta-hemolytic Streptococcus anginosus. J Bacteriol. 2013;195(5):1090–1099. doi:10.1128/JB.01344-12
29. Asam D, Mauerer S, Walheim E, Spellerberg B. Identification of beta-haemolysin-encoding genes in Streptococcus anginosus. Mol Oral Microbiol. 2013;28(4):302–315. doi:10.1111/omi.12026
30. Flaherty RA, Puricelli JM, Higashi DL, Park CJ, Lee SW. Streptolysin S promotes programmed cell death and enhances inflammatory signaling in epithelial keratinocytes during Group A streptococcus infection. Infect Immun. 2015;83(10):4118–4133. doi:10.1128/IAI.00611-15
31. Molloy EM, Casjens SR, Cox CL, et al. Identification of the minimal cytolytic unit for streptolysin S and an expansion of the toxin family. BMC Microbiol. 2015;15:141. doi:10.1186/s12866-015-0464-y
32. Bauer R, Mauerer S, Spellerberg B. Regulation of the beta-hemolysin gene cluster of Streptococcus anginosus by CcpA. Sci Rep. 2018;8(1):9028. doi:10.1038/s41598-018-27334-z
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.