Back to Journals » Journal of Inflammation Research » Volume 18
The m6A Modification in T Helper Cells Regulates the Pathogenesis of Autoimmune Diseases
Authors Jiang Q, Wang X, Wu Y, Cui D ![]()
Received 28 April 2025
Accepted for publication 10 September 2025
Published 19 September 2025 Volume 2025:18 Pages 13159—13172
DOI https://doi.org/10.2147/JIR.S537243
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Chaim Putterman
Qi Jiang,1,* Xin Wang,2,* Yingping Wu,3 Dawei Cui4
1Department of Blood Transfusion, Shaoxing People’s Hospital, Shaoxing, 312000, People’s Republic of China; 2Department of Rheumatology and Immunology, Shaoxing People’s Hospital, Shaoxing, 312000, People’s Republic of China; 3Department of Laboratory Medicine, The Fourth Affiliated Hospital, Zhejiang University School of Medicine, Yiwu, People’s Republic of China; 4Department of Blood Transfusion, the First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dawei Cui, Depatmernt of Blood Transfusion, The First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, 310003, People’s Republic of China, Email [email protected] Yingping Wu, Department of Laboratory Medicine, The Fourth Affiliated Hospital, Zhejiang University School of Medicine, Yiwu, People’s Republic of China, Email [email protected]
Abstract: The pathogenesis of autoimmune diseases remains unclear, which is associated with T helper (Th) cell subsets such as Th1, Th2, Treg and Tfh cells. Recent studies have indicated the involvement of N6-methyladenosine (m6A) modification in the pathogenesis of autoimmune diseases, and m6A also affects the differentiation, and function of Th cells. However, few reports focused on the relationship between m6A modification in Th cell subsets and autoimmune diseases. This review summarizes the latest research progress on m6A modification in common autoimmune diseases, specifically highlighting how various m6A-modifying enzymes influence the differentiation and function of Th cells and disease progression. Elucidating the relationship between m6A modification and Th cells in autoimmune diseases may provide a new perspective for disease prevention and targeted therapy. Finally, this review also elaborates on the impact of m6A on the clinical diagnosis and treatment of autoimmune diseases, as well as the challenges that need to be addressed.
Keywords: m6A, T helper cells, treatment, autoimmune diseases
Introduction
N6-methyladenosine (m6A) modification was discovered in mouse hepatoma cells in 1974, but it was not thoroughly investigated until 1977 when methyltransferase-like3 (METTL3) was isolated from HeLa cells by researchers.1,2 Since then, researchers have gradually developed a clearer understanding of m6A methylation. Simply put, m6A methylation is a chemical modification in which a methyl group is added to the nitrogen atom at the sixth position of adenosine in messenger RNA. This modification occurs after RNA synthesis, representing a form of post-transcriptional modification that is dynamic, reversible, and the most common form of mRNA modification (Figure 1). Methyltransferases, also known as m6A “writers”, primarily include METTL3, METTL5, METTL14, METTL16, Wilm’s tumour 1-associated protein (WTAP), ZC3H13, RBM15/15B and VIRMA (also called KIAA1429), and their main role is to add m6A modifications. Demethylases, referred to as m6A methylation “erasers”, mainly consist of fat mass and obesity-associated protein (FTO) and AlkB homologue 5 (ALKBH5), which function to remove m6A modifications. The m6A “readers” mainly comprise YT521-B homology (YTH) domain family (YTHDF1/2/3, YTHDC1/2), insulin-like growth factor 2 mRNA-binding proteins family (IGF2BP1/2/3), heterogeneous nuclear ribonucleoproteins (hnRNP) family (HNRNPC/G/A2B1), and ELAV-like protein 1 (ELAVL1), which recognize these modifications and convert the read information into functional signals.3,4 METTL3 and METTL14 form a stable heterodimeric complex in a 1:1 ratio, where METTL3 serves as the catalytic subunit, while METTL14 is responsible for recognizing target RNA and structurally stabilizing METTL3. WTAP helps mediate the localization of the METTL3 and METTL14 heterodimer on nuclear speckles, acting as a key regulatory molecule of the m6A methylation complex.5 KIAA1429 is the largest scaffold component of this complex, primarily regulating m6A methylation near the 3’ untranslated regions (3’ UTR) and stop codons.6 FTO can reverse m6A modifications in cells, stabilizing the 5’ cap structure of mRNA.7 ALKBH5 regulates RNA metabolism and mRNA export by reducing m6A levels in nuclear speckles.8 The m6A methyltransferases and demethylases complement each other, maintaining m6A methylation in a dynamic and reversible state. When m6A methylation occurs, m6A reader proteins recognize and bind to m6A-modified RNAs, regulating downstream functions and determining their fate. The YTHDF protein family mediates m6A-dependent regulation of mRNA metabolism through selective degradation and translational enhancement. Specifically, YTHDF1 primarily accelerates mRNA translation, YTHDF2 predominantly facilitates mRNA decay, while YTHDF3 exhibits dual functionality in both translation and degradation processes.9 The IGF2BP family is primarily responsible for enhancing mRNA stability and translation10 (Table 1). m6A modifications have been found in various types of RNA, including ribosomal RNA, transfer RNA, and non-coding RNA. These modifications regulate almost all aspects of mRNA metabolism, including export, splicing, translation, localization, and degradation, thereby affecting physiological processes such as metabolism, inflammatory responses, growth, and development. Abnormal m6A modifications can mediate the occurrence of diseases, including those caused by imbalances in helper T cells.
 |
Table 1 The Regulators Involved in m6A Methylation |
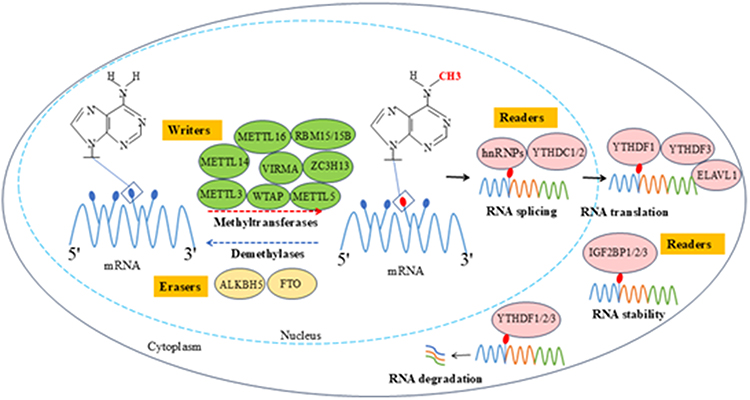 |
Figure 1 The dynamic methylation process of RNA m6A modification. m6A methylation is a dynamic and reversible biological process, regulated by the coordinated actions of methyltransferases (“writers”), demethylases (“erasers”), and methylation recognition proteins (“readers”). Writers (eg, METTL3, METTL14) catalyze the methylation of mRNA bases. Erasers (eg, FTO, ALKBH5) specifically remove methyl groups from mRNA. Readers (eg, YTHDF1/2/3) regulate mRNA stability, splicing, translation, and other processes. |
Th cells are differentiated from naïve CD4+ T cells upon stimulation by specific antigens. Based on the cytokines secreted after activation, they are classified into Th1, Th2, Th9, Th17, Th22, regulatory T cell (Treg), and T follicular helper (Tfh) cells.11 Each subpopulation is activated by specific cytokines and transcription factors, and it exert its effects in autoimmune diseases through the cytokines they secrete. Th1 cells primarily produce interferon-γ (IFN-γ), tumor necrosis factor-α (TNF-α), and interleukin (IL)-2, with T-bet as the main transcription factor for Th1 cell differentiation. Th1 cells play a crucial role in the pathogenesis of inflammatory bowel disease (IBD).12 Reports on the association between Th2 cells and m6A in autoimmune diseases are scarce. Th2 cells play a pivotal role in mediating immune responses to allergic reactions and parasitic infections. The release of inflammatory cytokines such as IL-4, IL-5, and IL-13 leads to bronchial constriction, increased vascular permeability, and inflammatory cell infiltration, resulting in allergic symptoms. IL-4 and IL-13 are the primary inflammatory cytokines in atopic dermatitis (AD).13 Macrophages with IGF2BP2 knockout show a reduced response to IL-4 but an enhanced inflammatory response to lipopolysaccharide (LPS), indicating that IGF2BP2 is involved in regulating macrophage activation in inflammatory diseases.13 Th9 cells secrete IL-9 under the stimulation of transforming growth factor-β (TGF-β) and IL-4. Currently, there are no direct reports linking Th9 cells to m6A in autoimmune diseases. Th17 cells, activated by the combined stimulation of IL-6 and TGF-β, activate their characteristic transcription factor retinoic acid-related orphan receptor γt (RORγt) and secrete inflammatory cytokines such as IL-17A, IL-17F, and IL-21 to exert immune functions. Th17 cells are involved in the disease progression of psoriasis and multiple sclerosis (MS).14,15 IL-22 is the characteristic cytokine of Th22 cells. Under the stimulation of IL-1β, IL-6, IL-23, and TNF-α, the secretion of IL-22, IL-13, IL-26, and TNF-α increases. Th22 cells play a significant role in autoimmune thyroid disease (AITD).16 Treg cells are a T cell subpopulation that exerts a prominent immunosuppressive effect and is one of the crucial factors maintaining immune tolerance in the body. Foxp3 is their characteristic transcription factor. Foxp3 is their characteristic transcription factor. Tregs exerts their effects by secreting inhibitory cytokines such as IL-10 and TGF-β. Tregs are closely associated with various autoimmune diseases such as rheumatoid arthritis (RA) and systemic lupus erythematosus (SLE). Both quantitative deficiency and functional impairment of Tregs have been demonstrated to exacerbate disease progression in autoimmune conditions.17,18 Under the induction of IL-6, IL-12, and IL-27, Tfh cells produce IL-21, which promotes the development of plasma cells and memory B cells. Bcl6 serves as the master transcriptional regulator of Tfh cells. Importantly, Tfh cells play a key role in the occurrence and development of primary Sjögren’s syndrome (pSS).19 This article mainly reviews the manifestations of m6A methylation in common autoimmune diseases (Figure 2) and its relationship with Th1, Th17, Th22, Treg, and Tfh cells. Finally, it summarizes the impact of these findings on clinical practice and treatment, aiming to provide more comprehensive theoretical knowledge for disease treatment (Table 2).
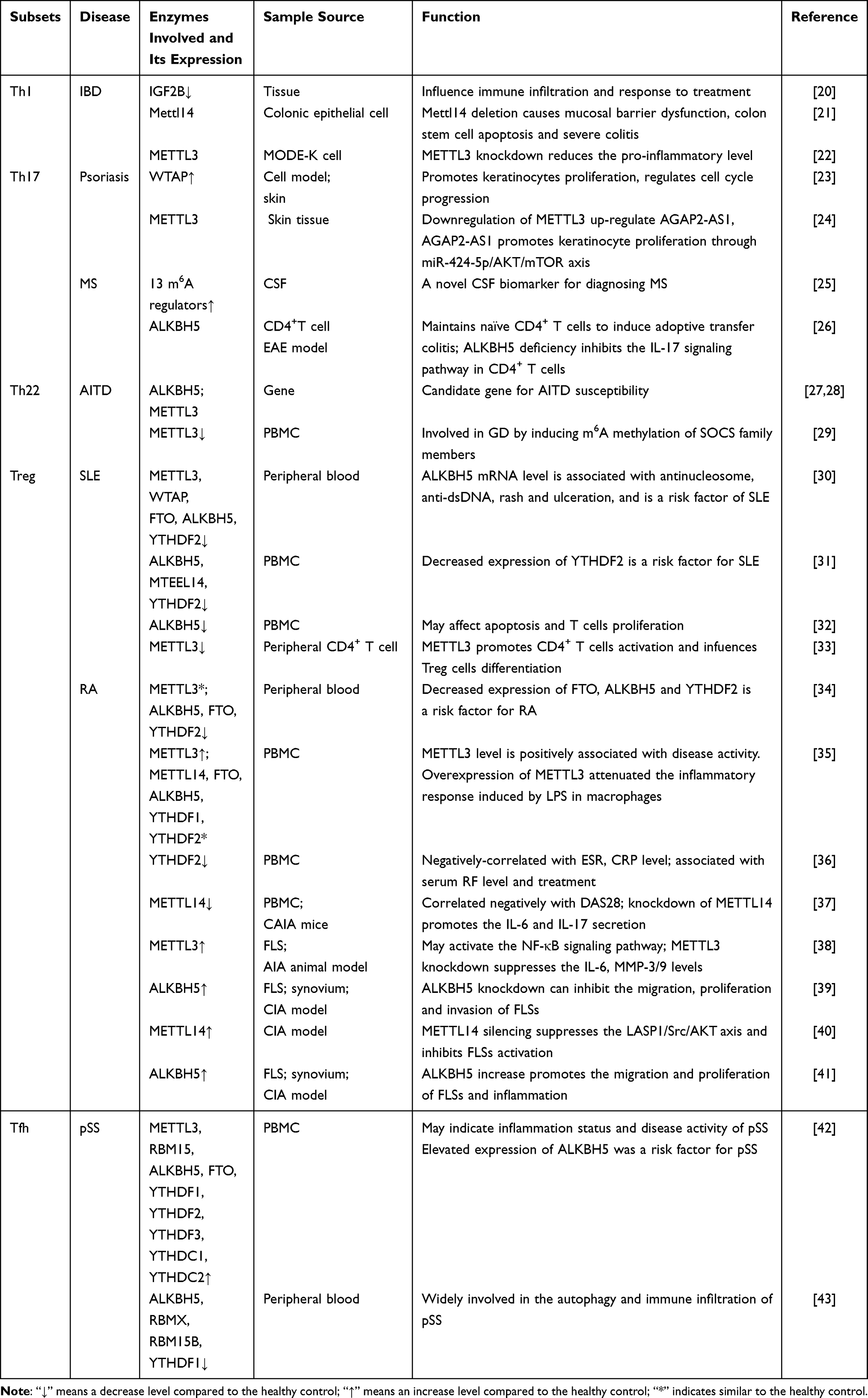 |
Table 2 m6A Modifications in T-Helper Cell Subset Disorders |
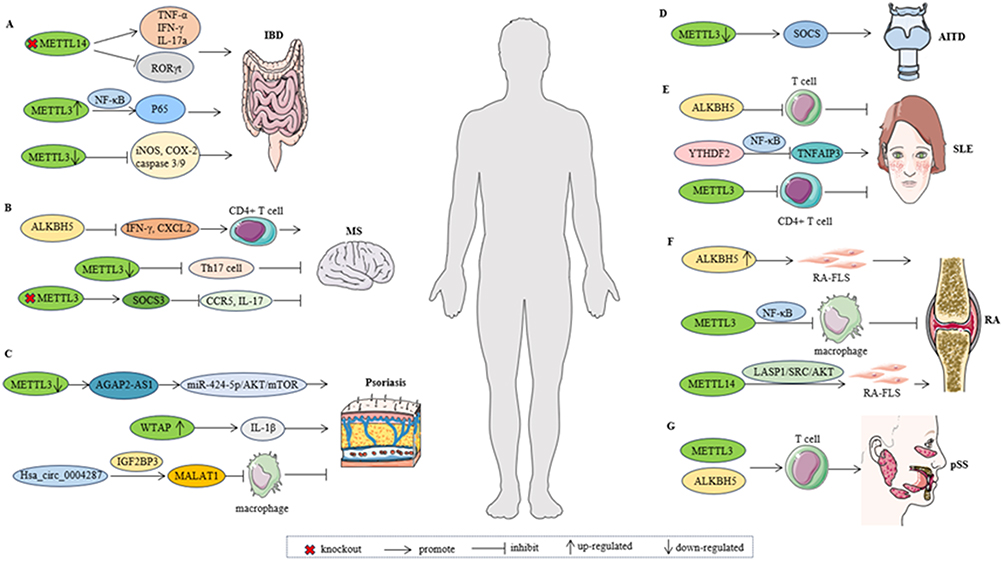 |
Figure 2 The possible mechanism of m6A methylase in various autoimmune diseases. (A). Knockout of Mettl14 in T cells leads to an increase in TNF-α, IFN-γ and IL-17a, a decrease in RORγt expression in Treg cells. Overexpression of METTL3 promotes the phosphorylation of p65. Knockdown of METTL3 reduces caspase 3/9 cleavage, and lowers levels of inflammatory enzymes. (B). ALKBH5 enhances the response of CD4+ T cells and increases the infiltration of neutrophils into the central nervous system by reducing the m6A modification in IFN-γ and CXCL2. The loss of METTL3 in T cells diminishes Th17 cell differentiation. The absence of METTL3 in Th17 cells enhances the stability of SOCS3 mRNA, attenuates the expression of CCR5 and IL-17A. (C). Downregulation of METTL3 increases AGAP2-AS1 expression. Overexpression of WTAP leads to increased expression of IL-1β. Hsa_circ_0004287 binds to IGF2BP3 in an m6A-dependent manner. (D). Knockdown of METTL3 reduces the expression of SOCS family members. (E). ALKBH5 may contribute to SLE pathogenesis by influencing T cell proliferation and apoptosis. Downregulates the level of YTHDF2 through the NF-κB pathway, thereby affecting the progression of SLE. Inhibiting METTL3 in SRBC-immunized mice significantly promotes CD4+ T cell activation. (F). Overexpression of ALKBH5 promotes the function of RA FLSs. Overexpression of METTL3 attenuated the inflammatory response induced by LPS in macrophages. METTL14 promote RA-FLS activation through the LASP1/SRC/AKT signaling pathway. (G). METTL3 and ALKBH5 were closely related to T cells in pSS. Abbreviations: IBD, Inflammatory bowel disease; MS, Multiple sclerosis; AITD, autoimmune thyroid disease; SLE, Systemic lupus erythematosus; RA, Rheumatoid arthritis; pSS, primary Sjögren’s syndrome. |
Role of m6A Modification in Autoimmune Diseases
Role of m6A Modification in Th1-Related Autoimmune Diseases
IBD, an idiopathic intestinal inflammatory condition affecting the ileum, rectum, and colon, includes Crohn’s disease (CD) and ulcerative colitis (UC). The pathogenesis of IBD, which remains incompletely understood, may be related to individual genetic susceptibility, immune response, gut microbiota, and the external environment.44 There is a high degree of heterogeneity in m6A regulatory factors among healthy individuals and patients with UC and CD. The expression of IGF2BP2 was significantly reduced in UC tissues, while both IGF2BP1 and IGF2BP2 were downregulated in CD tissues. m6A modification may affect immune infiltration in IBD and the response to treatment.20 Selective knockout of Mettl14 in T cells leads to an increase in Th1 cytokines (TNF-α, IFN-γ) and Th17 cytokines (IL-17a, IL-17c), a decrease in RORγt expression in Treg cells, and the induction of spontaneous colitis in mice.45 A colon-specific mouse model with Mettl14 deficiency exhibits mucosal barrier dysfunction and colonic stem cell apoptosis, leading to severe colitis. Mettl14 may limit the death of colonic epithelial cells by regulating the stability of an inhibitor of NF-κB (Nfkbia).21 Therefore, m6A modification is essential for maintaining colonic epithelial homeostasis. In LPS-treated MODE-K cells (a mouse intestinal epithelial cell line), knockdown of METTL3 inhibits apoptosis, reduces caspase 3/9 cleavage, and lowers levels of inflammatory enzymes (iNOS and COX-2) as well as proinflammatory cytokines such as TNF-α, IL-1β, and IL-6. Similarly, overexpression of METTL3 promotes the phosphorylation of p65 and exacerbates DSS-induced IBD in mice, suggesting the involvement of the NF-κB signaling pathway.22 This demonstrates the important role of METTL3 in inflammation regulation. Additionally, the loss of METTL3 in T cells disrupts T cell homeostasis and differentiation. Naïve T cells lacking Mettl3 show a reduction in Th1 and Th17 cells and an increase in Th2 cells46 (Figure 3A). Taken together, these data highlight the pivotal role of m6A methylation in regulating CD4+ T cell proliferation and differentiation, positioning it as a potential new breakthrough for IBD treatment.
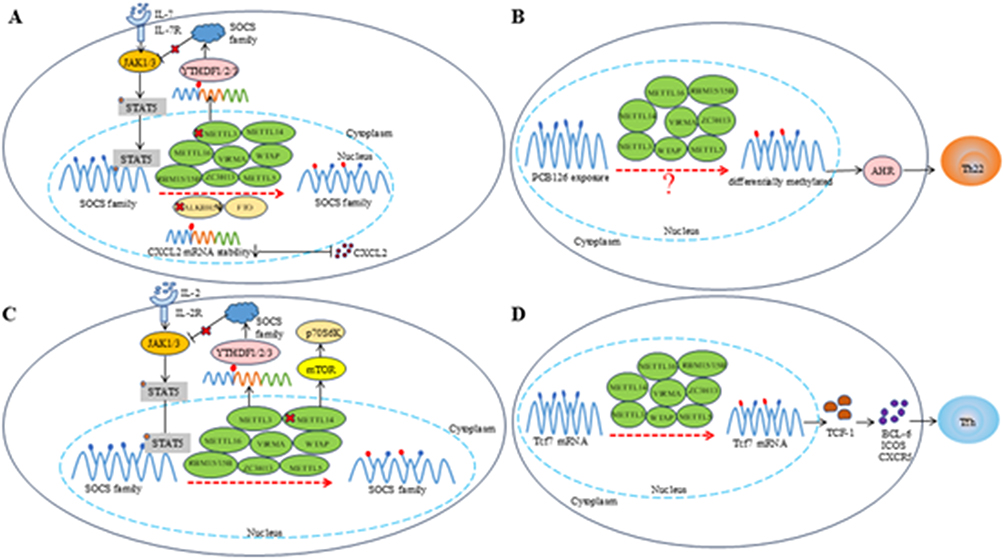 |
Figure 3 Schematic diagram of m6A modification. (A). m6A modification promotes the proliferation and differentiation of Th1 cells by degrading SOCS mRNA and further activating the IL-7-STAT5 pathway. METTL3 deficiency can enhance the stability of SOCS3 mRNA and reduce the differentiation of Th17 cells, and ALKBH5 deficiency can reduce the stability of CXCL2 and alleviate local infiltration; (B). m6A marked transcripts (AHR) exhibit differential methylation after exposure to a specific environment (PCB126), the exact mechanism of which is not yet clear; (C). METTL3 plays a key role in maintaining the inhibitory function of Tregs by degrading SOCS mRNA and activating the IL-2-STAT5 pathway. Mettl14 regulates iTreg inhibition through the mTOR pathway; (D). METTL3 stabilizes Tcf7 transcripts through m6A modification and plays a key role in promoting Tfh cell differentiation. |
Role of m6A Modification in Th17-Related Autoimmune Diseases
MS is a common and complex demyelinating disease of the central nervous system, with confirmed involvement of Th17 cells in its pathogenesis.47 Most patients experience a distinct pattern of relapse and remission, referred to as relapsing-remitting multiple sclerosis (RRMS).48 Studies have indicated that MS patients exhibit higher levels of 13 central m6A RNA methylation regulatory factors compared to non-MS individuals. Additionally, patients with progressive multiple sclerosis (PMS) demonstrate elevated cerebrospinal fluid (CSF) m6A methylation and enhanced expression of m6A-related genes relative to those with RRMS. Dynamic changes in m6A RNA methylation levels correlate with MS disease progression, enabling early differentiation between PMS and RRMS. Therefore, m6A RNA methylation level can serve as a novel CSF biomarker for MS diagnosis.25 Furthermore, while ALKBH5 does not affect T-cell development and function under homeostatic conditions, it controls the ability of naïve CD4+ T-cells to induce autoimmune colitis. The absence of ALKBH5 leads to reduced IFN-γ secretion in the central nervous system (CNS) and inhibits the trafficking of CD4+ T-cells to the CNS.26 By decreasing m6A modifications in IFN-γ and CXCL2, ALKBH5 enhances transcript protein expression and stability, resulting in augmented CD4+ T-cell responses and increased neutrophil infiltration into the CNS. In the commonly used animal model for MS, experimental autoimmune encephalomyelitis (EAE), the absence of ALKBH5 suppresses the IL-17 signaling pathway in CD4+ T-cells. Specific deletion of ALKBH5 confers protective effects in EAE.26,49 Additionally, the loss of METTL3 in T-cells diminishes Th17 cell differentiation, thereby mitigating the progression of EAE. Concurrently, the absence of METTL3 in Th17 cells enhances the stability of SOCS3 mRNA, attenuates the expression of CCR5 and IL-17A, reduces the migration of Th17 cells into the CNS, and consequently, alleviates the course of EAE46(Figure 3A).
Psoriasis is a chronic, recurrent immune-mediated inflammatory skin disease. Both mRNA and protein expression levels of WTAP were elevated in psoriatic cell models and lesioned skin of psoriasis patients, and this overexpression was primarily observed in the epidermal layer rather than the dermal layer.23 WTAP also affects the secretory function of keratinocytes. Overexpression of WTAP leads to increased expression of the inflammatory cytokine IL-1β, while knockdown of WTAP results in a decrease in IL-1β. WTAP also promotes keratinocyte proliferation, and excessive keratinocyte proliferation is a hallmark of psoriasis. WTAP may participate in the pathogenesis of psoriasis by accelerating cell cycle progression. Therefore, WTAP could potentially serve as a therapeutic target for psoriasis. Another study revealed methylation differences between uninvolved psoriatic skin, involved psoriatic skin, and healthy control (HC) skin. Specifically, involved psoriatic skin contained the fewest m6A peaks. Hypomethylated m6A was enriched in coding sequences (CDSs), 3’UTRs and 5’UTRs, while hypermethylated m6A was enriched in CDSs and 3’UTRs. Hypermethylation in involved psoriatic was particularly associated with olfactory transduction and cytokine production, while hypomethylation was primarily linked to the Wnt signaling pathway and developmental processes. Interestingly, as gene expression increases, m6A methylation also increases, further suggesting that m6A plays a regulatory role in gene expression in involved psoriatic skin.50 Additionally, the level of long noncoding RNA (lncRNA) AGAP2-AS1 in the skin tissue of psoriasis patients was higher than that in HC individuals. AGAP2-AS1, an endogenous competitive RNA, promotes keratinocyte proliferation and inhibits apoptosis. Downregulation of METTL3 expression increases AGAP2-AS1 expression in psoriasis patients. AGAP2-AS1 may promote keratinocyte proliferation through the miR-424-5p/AKT/mTOR axis.24 Studies have also shown that circular RNA (circRNA) Hsa_circ_0004287 is upregulated in PBMCs of patients with AD and psoriasis. In vitro inhibition of Hsa_circ_0004287 in macrophages upregulates IL-6, IL-1β, and TNF-α. Nuclear-localized circRNAs can function through interactions with their host genes. Knockdown of Hsa_circ_0004287 promotes the expression of its host gene MALAT1. Hsa_circ_0004287 binds to IGF2BP3 in an m6A-dependent manner, reducing MALAT1 stability, thereby inhibiting macrophage activation and suppressing psoriasis progression.51 In summary, m6A methylation modification plays a crucial role in the pathogenesis and progression of both MS and psoriasis. These findings not only suggest novel therapeutic strategies targeting m6A regulators (eg, WTAP or METTL3) but also indicate the potential of m6A methylation levels as diagnostic biomarkers for MS.
Role of m6A Modification in Th22-Related Autoimmune Diseases
AITD is considered an organ-specific autoimmune disease, in which thyroid autoantibodies can be detected in the blood of patients, including Graves’ disease and Hashimoto’s thyroiditis. Five genetic variations of ALKBH5 have been detected in patients with GD and HT, suggesting that ALKBH5 and METTL3 may be susceptible candidate genes for GD and HT.27,28 High-throughput microarray analysis has revealed abnormal expression of SOCS molecules and METTL3 in CD4+ T cells of GD patients. Upon METTL3 knockdown, the expression of certain SOCS family members increases, indicating that METTL3 may be involved in the pathogenesis of GD by inducing m6A methylation of SOCS mRNA.29 These studies collectively implicate m6A methylation in the etiology of AITD, and Th22 cells are also known to contribute to the disease progression of AITD.52,53 The transcription factor aryl hydrocarbon receptor (AhR) notably promotes the differentiation of naïve CD4+ T cells into Th22 cells54,55 (Figure 3B). Studies have shown that changes in m6A levels can affect the expression patterns of developmental genes, and m6A-marked transcripts undergo differential methylation upon exposure to certain environments. These transcripts include those activated by AHR agonists. Currently, there is limited research directly linking m6A to Th22 cells, necessitating further investigation to explore the specific mechanisms.
Role of m6A Modification in Treg-Related Autoimmune Diseases
SLE is an immune disease with complex progression and diverse clinical manifestations. lncRNAs are involved in regulating the development and pathogenesis of SLE, and abnormal expression of m6A-related lncRNAs was associated with the clinical manifestations of SLE patients.56 circRNA represents a novel type of non-coding RNA, among which circGARS was specifically overexpressed in SLE. By binding to microRNA-19a (miR-19a), circGARS downregulates the expression of YTHDF2, modulates the immune response through the NF-κB pathway, downregulates TNFAIP3 expression, and thereby affects the progression of SLE.57 Additionally, bioinformatics analysis suggests that the key regulatory factor of m6A (IGFBP3) and two immune genes (IDO1 and CD14) may be beneficial for the diagnosis and treatment of SLE.58 Studies have shown significantly reduced mRNA levels of METTL3, ALKBH5, YTHDF2, and FTO in the peripheral whole blood of SLE patients.30 Other research indicates lower levels of ALKBH5, METTL14, and YTHDF2 mRNA in the PBMCs of SLE patients compared to HCs. Decreased ALKBH5 mRNA correlates with CRP and fever. The reduced expression of YTHDF2 in PBMCs of SLE patients is a risk factor for SLE.31 One of the key features of SLE is the production of anti-double-stranded DNA antibodies (anti-dsDNA). ALKBH5 mRNA levels were inversely correlated with anti-dsDNA antibody levels, making ALKBH5 a risk factor for SLE pathogenesis.30 Both T cells and PBMCs from SLE patients show reduced ALKBH5 mRNA levels. Overexpression of ALKBH5 does not affect the T cell cycle but can inhibit T cell proliferation and promote apoptosis. The low levels of ALKBH5 in SLE patients may contribute to SLE pathogenesis by influencing T cell proliferation and apoptosis.32 Furthermore, both the post-transcriptional RNA modifification enzyme 5-methylcytosine (m5C) and NSUN2 expression were reduced in CD4+ T cells of SLE patients.59 METTL3 expression was also downregulated in CD4+ T cells of SLE patients. Inhibiting METTL3 in SRBC-immunized mice significantly promotes CD4+ T cell activation, weakens Treg and Tfh cell differentiation, and increases the proportion of Th1 and Th17 cells, with a particularly pronounced effect on Treg cells. Suppression of METTL3 elevates antibody production in a chronic graft-versus-host disease (cGVHD) mouse model, exacerbating the lupus-like phenotype. METTL3-mediated m6A modification of Foxp3 mRNA maintains Foxp3 protein expression and transcript stability.33
RA is a chronic immune disease characterized by synovial inflammation and progressive destruction of articular cartilage. The imbalance between Treg and Th17 cells is one of the important factors affecting the progression of RA. In recent years, with the deepening of research on m6A, there have been many reports on m6A-related factors in RA. Jiang et al obtained the transcriptional profile of the fibroblast-like synoviocyte cell line MH7A through high-throughput sequencing, suggesting that m6A modification was related to the occurrence of RA.60 The total m6A content in the peripheral blood of RA patients was remarkably increased, while the mRNA expression of FTO, ALKBH5, and YTHDF2 was significantly reduced. The degree of reduction was correlated with disease activity, and there was no distinct difference in METTL3 compared with the HC group.34 The expression of METTL3 in PBMCs of RA patients was notably increased, while there was no obvious difference in METTL14 and YTHDF2 compared to the HC group.35 Other studies have shown that the mRNA expression of YTHDF2 in PBMCs of RA patients was noticeably lower than that of HC group.36 Tang et al found that the levels of m6A and METTL14 in PBMCs of active RA patients were lower than those patients in remission and HCs. Knockdown of METTL14 could down-regulate m6A and promote the secretion of IL-6 and IL-17.37 These inconsistencies in research results may be due to different cell types, but the conclusions were consistent regarding the impact of m6A on synovial tissue and fibroblast-like synoviocytes (FLS) in RA. Current research results suggest that m6A-related enzymes are involved in synovial hyperplasia and promote the proliferation, migration, and invasion of FLS. Knockdown of ALKBH5 inhibits these functions of RA FLSs, while overexpression of ALKBH5 shows the opposite effect. This phenomenon has also been confirmed in the CIA mouse model.38,39,41,61 Under hypoxic conditions, the expression of ALKBH5 in FLS increases, and the HIF1α/2α-ALKBH5-CH25H pathway may be key to FLS attack and inflammation in this environment.41 METTL14 may promote FLS activation through the LASP1/SRC/AKT signaling pathway and participate in related inflammatory responses.40 In addition, in RA FLS, modular calcium-binding protein 2 (SMOC2) increases obviously. SMOC2 leads to increased levels of myosin 1c (MyO1C) through ALKBH5-mediated m6A modification, which causes RA FLS migration and invasion.62 m6A can also mediate the binding of lncRNA MAPKAPK5-AS1 to miR-146a-3p, affecting the inflammatory response and apoptosis of co-cultured RA-FLSs by targeting SIRT1.63 The lack of METTL3 results in a decrease in the differentiation of naive T cells into Th1 and Th17 cells but has little effect on the number of Treg cells. The lack of METTL3 in Treg cells leads to an increase in SOCS expression, inhibiting the IL-2-STAT5 signaling pathway, which is crucial for Treg cells to exert their suppressive function.46,64 Treg cells are divided into many different types, and Induced regulatory T cells (iTregs) can be formed in vitro. The treatment of certain autoimmune diseases using iTregs has attracted increasing attention from researchers. However, iTregs cannot maintain the inhibitory activity and expression of FoxP3. Zhao et al’s research found that METTL14 can maintain the immunosuppressive function of iTregs and may regulate the inhibitory function through the mTOR pathway, which plays a significant role in maintaining the function and stability of iTregs to sustain FOXP3 expression65 (Figure 3C). Taken together, m6A methylation participates in disease pathogenesis by regulating immune-inflammatory responses and modulating T cell quantity and function. Despite discrepancies in research findings, m6A methylation plays a pivotal regulatory role in both SLE and RA.
Role of m6A Modification in Tfh-Related Autoimmune Diseases
pSS is a chronic autoimmune disease characterized by the hypofunction or loss of function of the human exocrine glands, especially the salivary and lacrimal glands.66 One of the characteristics of pSS is the production of autoantibodies anti-SSA and anti-SSB. It has been reported that Tfh cells are involved in the pathogenesis of pSS.67 Moreover, Tfh cells are a crucial component of humoral immunity. METTL3 is essential for maintaining the expression of characteristic genes in Tfh cells, including CXCR5, BCL-6, ICOS, and Tcf7, and plays a significant role in the proliferation and survival of Tfh cells68 (Figure 3D). Some studies have analyzed the GEO database and found downregulated expression of ALKBH5, YTHDF1, RBMX, and RBM15B in the peripheral blood of pSS patients, as well as downregulated expression of ALKBH5, YTHDF1, METTL3, and RBM15B in labial gland tissue. Additionally, METTL3 and ALKBH5 were closely associated with the infiltrating immune cells such as T cells, indicating that m6A methylation was involved in the autophagy processes and immune infiltration of pSS.43 Other research has shown significantly elevated expression of METTL3, YTHDF1, YTHDF2, YTHDF3, ALKBH5, FTO, YTHDC1, and YTHDC2 in the peripheral blood of pSS patients compared to HCs. Elevated levels of ALKBH5 were a risk factor for pSS. m6A methyltransferases play a role in the autoimmune initiation of pSS by stimulating ISG15 to activate the type I IFN signaling pathway.42 He et al analyzed 23 m6A-mediated RNA modification patterns in blood and parotid gland samples from pSS patients using bioinformatics. They found differential expression of m6A regulators between the pSS group and the HC group. FMR1 was most remarkably expressed in blood samples, while FMR1 and HNRNPC were most notably expressed in parotid gland samples. m6A regulators may play a role in immune cell infiltration in pSS parotid gland tissue.69 In summary, m6A methylation may become a new therapeutic target for pSS.
Clinical Value of m6A Modifications for Autoimmune Diseases
The diverse manifestations of m6A methylation modification in various autoimmune diseases and its significant role in the interaction with Th cells will promote research based on m6A for diagnosis and treatment (Figure 3A–D). As discussed above, m6A-related enzymes (METTL3, METTL14, FTO, ALKBH5, WTAP) exhibit paradoxical roles in autoimmune diseases, either promoting or inhibiting disease progression. The effects of m6A modification appear to vary across different animal models. For instance, reducing METTL3 or METTL14 levels in T cells can produce opposing outcomes, likely due to differences in animal models and signaling pathways involved.45,46 A similar phenomenon has been observed in RA research. Knockdown of METTL14 in the synovium of mice and rats also yielded contrasting effects.37,40 Even when analyzing PBMCs from the same disease, inconsistent levels of m6A-related enzymes suggest that m6A modification may function differently depending on disease stage or activity.36,37 Furthermore, the immune microenvironment may influence m6A modification. For example, hypoxia upregulates ALKBH5 expression in FLSs. Thus, the function of m6A enzymes likely depends on cellular context, disease status, and molecular mechanisms, highlighting the complexity and plasticity of epigenetic regulatory networks.
As mentioned earlier, the expression of m6A methylation levels in IBD, MS, Psoriasis, AITD, SLE, RA, and pSS differs from that of the HC group. This differential expression makes it possible to use the detection of m6A methylation levels in peripheral blood for early disease diagnosis. For instance, the level of m6A RNA methylation can distinguish between PMS and RRMS early in the disease process, serving as a CSF marker for MS diagnosis.25 The hypermethylated circRNA hsa_circ_0007259 activates the STAT3 signaling pathway via hsa_miR-21-5p. This circRNA plays a role in the pathophysiology of RA and can serve as a valuable biomarker for RA diagnosis.70 Furthermore, researchers have identified five differentially expressed m6A regulatory factors between healthy individuals and RA patients using the GEO database. Through software analysis, they constructed an m6Asig system capable of distinguishing between RA C1 and C2 subtypes (associated with higher inflammatory responses). This system could potentially become a biomarker for inflammatory activity in RA.71 Although researchers have made some progress in exploring the diagnostic value of m6A methylation in early-stage diseases, further studies are still needed to verify the feasibility and accuracy of these findings.
Beyond diagnostics, m6A modification impacts Th cell differentiation, function, and cytokine secretion, while interfering with RNA splicing, export, translation, and degradation through methyltransferases, demethylases, and binding proteins. This makes it possible to develop agonists or inhibitors of these enzymes and proteins to control disease progression. In 2012, some small-molecule inhibitors of FTO demethylase were discovered, with the natural product rhein emerging as the most potent by competitively binding to the FTO active site.72 The natural compound radicicol also acts as an FTO inhibitor, exhibiting dose-dependent inhibition of FTO demethylation activity.73 Furthermore, based on the protein-ligand interaction structure of FTO, researchers have identified two small-molecule inhibitors of FTO.74,75 Interestingly, entacapone, a drug used in combination with other medications to treat Parkinson’s disease, has also been found to inhibit FTO, which has been verified in experimental animals.76 Additionally, a non-steroidal anti-inflammatory drug Meclofenamic acid (MA) has been discovered as a highly selective inhibitor of FTO, inhibiting its demethylation of m6A. Mechanistic studies reveal that MA selectively inhibits FTO demethylation through competitive binding with m6A-containing substrates, exhibiting greater potency than ALKBH5.77 Currently, most reported inhibitors target FTO, possibly due to its early discovery as a demethylase in mammals. Researchers have screened two series of adenosine derivatives through high-throughput docking with METTL3, identifying ligands with good performance that selectively inhibit METTL3, although further studies are needed to confirm their specific effects.78 Chinese Ecliptae herba extract and its main component wedelolactone may enhance osteoblastogenesis of bone marrow mesenchymal stem cells (BMSCs) by targeting METTL3-mediated m6A methylation of VEGF-A and HIF-1α.79 Paracetamol can alter total m6A levels in IL-1β-treated chondrocytes (mimicking an inflammatory state), upregulating ALKBH5, downregulating METTL3, and inhibiting the secretion of inflammatory cytokines such as IL-6 and IL-8.80 Besides the use of natural compounds or small molecules to modulate the expression of m6A regulatory factors in disease treatment, the modification of these factors at the molecular level, as mentioned earlier in this review, also exhibits significant regulatory effects. While inhibitors of other methyltransferases have not been reported yet, these findings provide new insights and directions for drug development. Overall, m6A modification has the potential to offer new insights for the diagnosis of autoimmune diseases and the development of clinical drugs.
Challenges of m6A Methylation in Autoimmune Diseases
Accumulating evidence suggests that dynamic and reversible m6A methylation affects multiple aspects of immune responses and Th cells, such as development, differentiation, migration, cytokine secretion, and inflammatory responses. Based on these functions, m6A methylation has been linked to various immune-related diseases, including viral infections, inflammation, autoimmune diseases, and tumorigenesis.81,82 So far, the role of m6A methylation in tumorigenesis has garnered significant attention, leading researchers to conduct numerous studies and yield encouraging results. For instance, targeting METTL3 has demonstrated positive therapeutic effects in multiple tumors, including acute myeloid leukemia, breast cancer, lung cancer, liver cancer, and stomach cancer.83,84 However, despite autoimmune diseases also being one of the significant health issues worldwide, research on the role of m6A methylation in the development and progression of autoimmune diseases, therapeutic potential, and interaction with Th cells is still in its infancy. It is hoped that future research will further untangle the upstream mechanisms of these methylation enzymes, and interventions targeting the upstream factors of these enzymatic modifications have the potential to become innovative methods for improving disease treatment. Although some progress has been made in understanding the underlying mechanisms of m6A modification in the pathological and biological processes of autoimmune diseases, detailed studies on the role of m6A modification in clinical diagnosis, treatment, and prognosis remain inadequate. Furthermore, many enzymes are involved in the entire methylation process, and while current research has primarily focused on METTL3, METTL14, WTAP, FTO, and ALKBH5, there is a paucity of studies on enzymes that read m6A modifications. m6A readers have a significant impact on the stability of the target mRNA regulated by methylation, highlighting the need for more comprehensive research in this area.
Conclusion and Prospective
With the advancement of molecular biology techniques, our understanding of disease mechanisms has gradually extended to genetic and epigenetic processes. Consequently, the role of m6A methylation modification in autoimmune diseases has been uncovered with the development of life sciences and detection technology. In this review, we summarize the latest research progress on m6A methylation in IBD, MS, Psoriasis, AITD, SLE, RA, and pSS, elucidating its impact on Th cell development, differentiation, and cytokine secretion in these diseases. Taken together, the m6A modification has a critical impact on differentiation and function of Th cells that are involved into the pathogenesis of various autoimmune diseases, which deserves further investigation and discussion, and targeting m6A regulatory factors may emerge as a new strategy for the treatment of autoimmune diseases.
Data Sharing Statement
No data was used for the research described in the article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Medical Health Science and Technology Project of Zhejiang Province (Grant No. 2023RC105, 2023KY1235), National Natural Science Foundation of China (Grant No. 81871709), and Zhejiang Provincial Natural Science Foundation of China (Grant No. LMS25H200004, LBY21H190001).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Desrosiers R, Friderici K, Rottman F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc Natl Acad Sci USA. 1974;71(10):3971–3975. doi:10.1073/pnas.71.10.3971
2. Bokar JA, Shambaugh ME, Polayes D, Matera AG, Rottman FM. Purification and cDNA cloning of the AdoMet-binding subunit of the human mRNA (N6-adenosine)-methyltransferase. RNA. 1997;3(11):1233–1247.
3. Oerum S, Meynier V, Catala M, Tisné C. A comprehensive review of m6A/m6Am RNA methyltransferase structures. Nucleic Acids Res. 2021;49(13):7239–7255. doi:10.1093/nar/gkab378
4. Sun T, Wu R, Ming L. The role of m6A RNA methylation in cancer. Biomed Pharmacothe. 2019;112:108613. doi:10.1016/j.biopha.2019.108613
5. Zaccara S, Ries RJ, Jaffrey SR. Reading, writing and erasing mRNA methylation. Nat Rev Mol Cell Biol. 2019;20(10):608–624. doi:10.1038/s41580-019-0168-5
6. Zhang X, Li MJ, Xia L, Zhang H. The biological function of m6A methyltransferase KIAA1429 and its role in human disease. PeerJ. 2022;10:e14334. doi:10.7717/peerj.14334
7. Li Y, Su R, Deng X, Chen Y, Chen J. FTO in cancer: functions, molecular mechanisms, and therapeutic implications. Trends Cancer. 2022;8(7):598–614. doi:10.1016/j.trecan.2022.02.010
8. Chen H, Zhang J, Yan Y, et al. N6-methyladenosine RNA demethylase ALKBH5 is testis-specifically downregulated in hybrid male sterile dzo and is a target gene of bta-miR-200a. Theriogenology. 2022;187:51–57. doi:10.1016/j.theriogenology.2022.04.022
9. Seo KW, Kleiner RE. YTHDF2 Recognition of N(1)-Methyladenosine (m(1)A)-Modified RNA Is Associated with Transcript Destabilization. ACS Chem. Biol. 2020;15(1):132–139. doi:10.1021/acschembio.9b00655
10. Huang H, Weng H, Sun W, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20(3):285–295. doi:10.1038/s41556-018-0045-z
11. Raphael I, Nalawade S, Eagar TN, Forsthuber TG. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine. 2015;74(1):5–17. doi:10.1016/j.cyto.2014.09.011
12. Gomez-Bris R, Saez A, Herrero-Fernandez B, Rius C, Sanchez-Martinez H, Gonzalez-Granado JM. CD4 T-cell subsets and the pathophysiology of inflammatory bowel disease. Int J Mol Sci. 2023;24(3).
13. Wang X, Ji Y, Feng P, et al. The m6A Reader IGF2BP2 regulates macrophage phenotypic activation and inflammatory diseases by stabilizing TSC1 and PPARγ. Adv. Sci. 2021;8(13):2100209. doi:10.1002/advs.202100209
14. Li B, Huang L, Lv P, et al. The role of Th17 cells in psoriasis. Immunol Res. 2020;68(5):296–309. doi:10.1007/s12026-020-09149-1
15. Liu R, Du S, Zhao L, et al. Autoreactive lymphocytes in multiple sclerosis: pathogenesis and treatment target. Front Immunol. 2022;13:996469. doi:10.3389/fimmu.2022.996469
16. Janyga S, Marek B, Kajdaniuk D, Ogrodowczyk-Bobik M, Urbanek A, Ł B. CD4+ cells in autoimmune thyroid disease. Endokrynologia Polska. 2021;72(5):572–583. doi:10.5603/EP.a2021.0076
17. Scheinecker C, Göschl L, Bonelli M. Treg cells in health and autoimmune diseases: new insights from single cell analysis. J Autoimmun. 2020;110:102376. doi:10.1016/j.jaut.2019.102376
18. Jiang Q, Yang G, Liu Q, Wang S, Cui D. Function and role of regulatory t cells in rheumatoid arthritis. Front Immunol. 2021;626193. doi:10.3389/fimmu.2021.626193
19. Wei X, Niu X. T follicular helper cells in autoimmune diseases. J Autoimmun. 2023;134:102976. doi:10.1016/j.jaut.2022.102976
20. Chen Y, Lei J, He S. m(6)A modification mediates mucosal immune microenvironment and therapeutic response in inflammatory bowel disease. Front Cell Develop Biol. 2021;9:692160. doi:10.3389/fcell.2021.692160
21. Zhang T, Ding C, Chen H, et al. m(6)A mRNA modification maintains colonic epithelial cell homeostasis via NF-κB-mediated antiapoptotic pathway. Sci Adv. 2022;8(12):eabl5723. doi:10.1126/sciadv.abl5723
22. Yang L, Wu G, Wu Q, Peng L, Yuan L. METTL3 overexpression aggravates LPS-induced cellular inflammation in mouse intestinal epithelial cells and DSS-induced IBD in mice. Cell Death Disc. 2022;8(1):62. doi:10.1038/s41420-022-00849-1
23. Kong Y, Wu R, Zhang S, et al. Wilms’ tumor 1-associating protein contributes to psoriasis by promoting keratinocytes proliferation via regulating cyclinA2 and CDK2. Int Immunopharmacol. 2020;88:106918. doi:10.1016/j.intimp.2020.106918
24. Xian J, Shang M, Dai Y, et al. N(6)-methyladenosine-modified long non-coding RNA AGAP2-AS1 promotes psoriasis pathogenesis via miR-424-5p/AKT3 axis. J Dermatological Sci. 2022;105(1):27–36. doi:10.1016/j.jdermsci.2021.11.007
25. Ye F, Wang T, Wu X, Liang J, Li J, Sheng W. N6-Methyladenosine RNA modification in cerebrospinal fluid as a novel potential diagnostic biomarker for progressive multiple sclerosis. J Transl Med. 2021;19(1):316. doi:10.1186/s12967-021-02981-5
26. Zhou J, Zhang X, Hu J, et al. m(6)A demethylase ALKBH5 controls CD4(+) T cell pathogenicity and promotes autoimmunity. Sci Adv. 2021;7(25). doi:10.1126/sciadv.abg0470.
27. Song RH, Zhao J, Gao CQ, Qin Q, Zhang JA. Inclusion of ALKBH5 as a candidate gene for the susceptibility of autoimmune thyroid disease. Adv Med Sci. 2021;66(2):351–358. doi:10.1016/j.advms.2021.07.006
28. Song RH, Liu XR, Gao CQ, Du P, Zhang JA. METTL3 gene polymorphisms contribute to susceptibility to autoimmune thyroid disease. Endocrine. 2021;72(2):495–504. doi:10.1007/s12020-020-02503-1
29. Song RH, Du P, Gao CQ, Liu XR, Zhang JA. METTL3 Is involved in the development of graves’ disease by Inducing SOCS mRNA m6A modification. Front Endocrinol. 2021;12:666393. doi:10.3389/fendo.2021.666393
30. Luo Q, Fu B, Zhang L, Guo Y, Huang Z, Li J. Decreased peripheral blood ALKBH5 correlates with markers of autoimmune response in systemic lupus erythematosus. Dis. Markers. 2020;2020:8193895. doi:10.1155/2020/8193895
31. Luo Q, Rao J, Zhang L, et al. The study of METTL14, ALKBH5, and YTHDF2 in peripheral blood mononuclear cells from systemic lupus erythematosus. Mol. Genet. Genomic Med. 2020;8(9):e1298. doi:10.1002/mgg3.1298
32. Deng LJ, Fang XY, Wu J, et al. ALKBH5 expression could affect the function of t cells in systemic lupus erythematosus patients: a case-control study. Curr. Pharm. Des. 2022;28(27):2270–2278. doi:10.2174/1381612828666220617154204
33. Lu S, Wei X, Zhu H, et al. m(6)A methyltransferase METTL3 programs CD4(+) T-cell activation and effector T-cell differentiation in systemic lupus erythematosus. Mol Med. 2023;29(1):46. doi:10.1186/s10020-023-00643-4
34. Luo Q, Gao Y, Zhang L, et al. Decreased ALKBH5, FTO, and YTHDF2 in peripheral blood are as risk factors for rheumatoid arthritis. Biomed Res. Int. 2020;2020:5735279. doi:10.1155/2020/5735279
35. Wang J, Yan S, Lu H, Wang S, Xu D. METTL3 attenuates lps-induced inflammatory response in macrophages via nf-κb signaling pathway. Mediators Inflamm. 2019;2019:3120391. doi:10.1155/2019/3120391
36. Yao F, Xu C, Gao Y, et al. Expression and clinical significance of the m6A reader YTHDF2 in peripheral blood mononuclear cells from rheumatoid arthritis patients. J Immunotoxicol. 2022;19(1):53–60. doi:10.1080/1547691X.2022.2067916
37. Tang J, Yu Z, Xia J, et al. METTL14-Mediated m6A modification of TNFAIP3 involved in inflammation in patients with active rheumatoid arthritis. Arthritis Rheumatol. 2023;75(12):2116–2129. doi:10.1002/art.42629
38. Shi W, Zheng Y, Luo S, et al. METTL3 promotes activation and inflammation of FLSs through the NF-κB signaling pathway in rheumatoid arthritis. Front Med. 2021;8:607585. doi:10.3389/fmed.2021.607585
39. Kuang Y, Li R, Wang J, et al. ALKBH5-Mediated RNA m(6) A methylation regulates the migration, invasion, and proliferation of rheumatoid fibroblast-like synoviocytes. Arthritis Rheumatol. 2024;76(2):192–205. doi:10.1002/art.42676
40. Li X, Xu X, Zhang Q, Ling M, Li X, Tan X. METTL14 promotes fibroblast-like synoviocytes activation via the LASP1/SRC/AKT axis in rheumatoid arthritis. Am J Physiol Cell Physiol. 2023;324(5):C1089–c100. doi:10.1152/ajpcell.00575.2022
41. Fan D, Geng Q, Wang B, et al. Hypoxia-induced ALKBH5 aggravates synovial aggression and inflammation in rheumatoid arthritis by regulating the m6A modification of CH25H. Clin Immunol. 2024;261:109929. doi:10.1016/j.clim.2024.109929
42. Xiao Q, Wu X, Deng C, et al. The potential role of RNA N6-methyladenosine in primary Sjögren’s syndrome. Front Med. 2022;9:959388. doi:10.3389/fmed.2022.959388
43. Cheng L, Li H, Zhan H, et al. Alterations of m6A RNA methylation regulators contribute to autophagy and immune infiltration in primary Sjögren’s syndrome. Front Immunol. 2022;13:949206. doi:10.3389/fimmu.2022.949206
44. Ryan FJ, Ahern AM, Fitzgerald RS, et al. Colonic microbiota is associated with inflammation and host epigenomic alterations in inflammatory bowel disease. Nat Commun. 2020;11(1):1512. doi:10.1038/s41467-020-15342-5
45. Lu TX, Zheng Z, Zhang L, et al. A new model of spontaneous colitis in mice induced by deletion of an RNA m(6)A methyltransferase component METTL14 in T cells. CMGH. 2020;10(4):747–761. doi:10.1016/j.jcmgh.2020.07.001
46. Li HB, Tong J, Zhu S, et al. m(6)A mRNA methylation controls T cell homeostasis by targeting the IL-7/STAT5/SOCS pathways. Nature. 2017;548(7667):338–342. doi:10.1038/nature23450
47. Kaskow BJ, Baecher-Allan C. Effector T cells in multiple sclerosis. Cold Spring Harb Perspect Med. 2018;8(4):a029025. doi:10.1101/cshperspect.a029025
48. Lassmann H, van Horssen J, Mahad D. Progressive multiple sclerosis: pathology and pathogenesis. Nat Rev Neurol. 2012;8(11):647–656. doi:10.1038/nrneurol.2012.168
49. Constantinescu CS, Farooqi N, O’Brien K, Gran B. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br. J. Pharmacol. 2011;164(4):1079–1106. doi:10.1111/j.1476-5381.2011.01302.x
50. Wang YN, Jin HZ. Transcriptome-Wide m(6)A methylation in skin lesions from patients with psoriasis vulgaris. Front Cell Develop Biol. 2020;8:591629. doi:10.3389/fcell.2020.591629
51. Yang L, Fu J, Han X, et al. Hsa_circ_0004287 inhibits macrophage-mediated inflammation in an N(6)-methyladenosine-dependent manner in atopic dermatitis and psoriasis. J Allergy Clin Immunol. 2022;149(6):2021–2033. doi:10.1016/j.jaci.2021.11.024
52. Bai X, Sun J, Wang W, et al. Increased differentiation of Th22 cells in Hashimoto’s thyroiditis. Endocr J. 2014;61(12):1181–1190. doi:10.1507/endocrj.EJ14-0265
53. Song RH, Yu ZY, Qin Q, et al. Different levels of circulating Th22 cell and its related molecules in Graves’ disease and Hashimoto’s thyroiditis. Int J Clin Exp Pathol. 2014;7(7):4024–4031.
54. Aluru N, Karchner SI. PCB126 exposure revealed alterations in m6a RNA modifications in transcripts associated with AHR activation. Toxicol Sci. 2021;179(1):84–94. doi:10.1093/toxsci/kfaa158
55. Jiang Q, Yang G, Xiao F, et al. Role of Th22 Cells in the Pathogenesis of Autoimmune Diseases. Front Immunol. 2021;12:688066. doi:10.3389/fimmu.2021.688066
56. Wu J, Deng L-J, Xia Y-R, et al. Involvement of N6-methyladenosine modifications of long noncoding RNAs in systemic lupus erythematosus. Mol Immunol. 2022;143:77–84. doi:10.1016/j.molimm.2022.01.006
57. Zhao X, Dong R, Zhang L, et al. N6-methyladenosine-dependent modification of circGARS acts as a new player that promotes SLE progression through the NF-κB/A20 axis. Arthritis Res Ther. 2022;24(1):37. doi:10.1186/s13075-022-02732-x
58. Zhao X, Ge L, Wang J, et al. Exploration of potential integrated models of N6-methyladenosine immunity in systemic lupus erythematosus by bioinformatic analyses. Front Immunol. 2021;12:752736. doi:10.3389/fimmu.2021.752736
59. Guo G, Wang H, Shi X, et al. Disease Activity-Associated Alteration of mRNA m(5) C Methylation in CD4(+) T Cells of Systemic Lupus Erythematosus. Front Cell Develop Biol. 2020;8:430. doi:10.3389/fcell.2020.00430
60. Jiang H, Cao K, Fan C, Cui X, Ma Y, Liu J. Transcriptome-wide high-throughput m6a sequencing of differential m6a methylation patterns in the human rheumatoid arthritis fibroblast-like synoviocytes cell Line MH7A. J Inflamm Res. 2021;14:575–586. doi:10.2147/JIR.S296006
61. Xiao J, Cai X, Wang R, Zhou W, Ye Z. ALKBH5-YTHDF2 m6A modification axis inhibits rheumatoid arthritis progression by suppressing NLRP3. Biochem. Biophys. Res. Commun. 2023;668:70–76. doi:10.1016/j.bbrc.2023.05.087
62. Liu D, Li R, Xu S, et al. SMOC2 promotes aggressive behavior of fibroblast-like synoviocytes in rheumatoid arthritis through transcriptional and post-transcriptional regulating MYO1C. Cell Death Dis. 2022;13(12):1035. doi:10.1038/s41419-022-05479-0
63. Wen J, Liu J, Wan L, et al. m(6)A-mediated lncRNA MAPKAPK5-AS1 induces apoptosis and suppresses inflammation via regulating miR-146a-3p/SIRT1/NF-κB axis in rheumatoid arthritis. Cell Cycle. 2023;22(23–24):2602–2621. doi:10.1080/15384101.2024.2302281
64. Tong J, Cao G, Zhang T, et al. m(6)A mRNA methylation sustains Treg suppressive functions. Cell Res. 2018;28(2):253–256. doi:10.1038/cr.2018.7
65. Liu Y, Yuan Y, Zhou Z, et al. Mettl14 sustains FOXP3 expression to promote the differentiation and functions of induced-regulatory T cells via the mTOR signaling pathway. Immunol Lett. 2023;258:35–44. doi:10.1016/j.imlet.2023.04.008
66. Imgenberg-Kreuz J, Rasmussen A, Sivils K, Nordmark G. Genetics and epigenetics in primary Sjögren’s syndrome. Rheumatology. 2021;60(5):2085–2098. doi:10.1093/rheumatology/key330
67. Gong YZ, Nititham J, Taylor K, et al. Differentiation of follicular helper T cells by salivary gland epithelial cells in primary Sjögren’s syndrome. J Autoimmun. 2014;51:57–66. doi:10.1016/j.jaut.2013.11.003
68. Yao Y, Yang Y, Guo W, et al. METTL3-dependent m(6)A modification programs T follicular helper cell differentiation. Nat Commun. 2021;12(1):1333. doi:10.1038/s41467-021-21594-6
69. He F, Liu H, Yu C. N(6)-methyladenosine regulator-mediated RNA methylation is involved in primary sjögren’s syndrome Immunoinfiltration. Dis. Markers. 2022;2022:5242287. doi:10.1155/2022/5242287
70. Luo H, Wei J, Wu S, Zheng Q, Zhang N, Chen P. Exploring CircRNA N6-methyladenosine in human rheumatoid arthritis: hyper-methylated hsa_circ_0007259 as a potential biomarker and its involvement in the hsa_circ_0007259/hsa_miR-21-5p/STAT3 axis. Int Immunopharmacol. 2023;124:110938. doi:10.1016/j.intimp.2023.110938
71. Ma C, Wu J, Lei H, Huang H, Li Y. Significance of m6A in subtype identification, immunological evolution, and therapeutic sensitivity of RA. Immunobiology. 2024;229(1):152781. doi:10.1016/j.imbio.2023.152781
72. Chen B, Ye F, Yu L, et al. Development of cell-active N6-methyladenosine RNA demethylase FTO inhibitor. J Am Chem Soc. 2012;134(43):17963–17971. doi:10.1021/ja3064149
73. Wang R, Han Z, Liu B, et al. Identification of natural compound radicicol as a potent FTO inhibitor. Mol Pharmaceut. 2018;15(9):4092–4098. doi:10.1021/acs.molpharmaceut.8b00522
74. He W, Zhou B, Liu W, et al. Identification of A novel small-molecule binding site of the fat mass and obesity associated protein (FTO). J Med Chem. 2015;58(18):7341–7348. doi:10.1021/acs.jmedchem.5b00702
75. Qiao Y, Zhou B, Zhang M, et al. A novel Inhibitor of the obesity-related protein FTO. Biochemistry. 2016;55(10):1516–1522. doi:10.1021/acs.biochem.6b00023
76. Peng S, Xiao W, Ju D, et al. Identification of entacapone as a chemical inhibitor of FTO mediating metabolic regulation through FOXO1. Sci, trans med. 2019;11(488). doi:10.1126/scitranslmed.aau7116.
77. Huang Y, Yan J, Li Q, et al. Meclofenamic acid selectively inhibits FTO demethylation of m6A over ALKBH5. Nucleic Acids Res. 2015;43(1):373–384. doi:10.1093/nar/gku1276
78. Bedi RK, Huang D, Eberle SA, Wiedmer L, Śledź P, Caflisch A. Small-molecule inhibitors of METTL3, the major human epitranscriptomic writer. ChemMedChem. 2020;15(9):744–748. doi:10.1002/cmdc.202000011
79. Tian S, Li YL, Wang J, et al. Chinese Ecliptae herba (Eclipta prostrata (L.) L.) extract and its component wedelolactone enhances osteoblastogenesis of bone marrow mesenchymal stem cells via targeting METTL3-mediated m6A RNA methylation. J Ethnopharmacol. 2023;312:116433. doi:10.1016/j.jep.2023.116433
80. Gao J, Li Y, Liu Z, Wang D, Zhang H. Acetaminophen changes the RNA m(6)A levels and m(6)A-related proteins expression in IL-1β-treated chondrocyte cells. BMC Mol Cell Biol. 2022;23(1):45. doi:10.1186/s12860-022-00444-3
81. Xu L, Shen T, Li Y, Wu X. The Role of M(6)A modification in autoimmunity: emerging mechanisms and therapeutic implications. Clin Rev Allergy Immunol. 2025;68(1):29.
82. Li Z, Lao Y, Yan R, Li F, Guan X, Dong Z. N6-methyladenosine in inflammatory diseases: important actors and regulatory targets. Gene. 2025;936:149125. doi:10.1016/j.gene.2024.149125
83. Mao Z, Li M, Wang S. Targeting m(6)A RNA modification in tumor therapeutics. Curr Oncol. 2025;32(3):159. doi:10.3390/curroncol32030159
84. Meng W, Xiao H, Mei P, et al. Critical Roles of METTL3 in translation regulation of cancer. Biomolecules. 2023;13(2):243. doi:10.3390/biom13020243
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.